Pulse-Step Model for Saccade Generation
Up to now we have considered eye movement control which tries
to match eye velocity to target velocity, without worry that the fovea had to
be directed at the target. Now, with saccades, we consider the system that cares
about the fovea is looking at the (small) target.. So saccades, in conjuction
with fixation, are position-goal directed eye movementsl.
Think of a saccade as a command to move toward the target
at about 400 deg per sec (7 rad/sec). A saccades stops when the target is reached:
no overshoot, no undershoot. We know that the control signal must deal with
the mechanics of the eyeball in its orbit, by way of the OMN integrator and
"speed control".
Pulse-step control
The speed of eye movement is the derivative of E(t). Remember from Laplace
transforms that

The saccade control system must supply (in terms of degrees) a steady input of 400
to the OMN to achieve the required speed. Notice how the "s" term has
been absorbed to regenerate the expected OMN model. And yes, if you supply a steady
input to the OMN, the resulting eye movement will be a perfect ramp. (A
steady input is seen as a 1/s step and the OMN+plant collapses to 1/s; 1/s^2) is
the Laplace transform of the ramp.) In fact the OMN motoneurons will
generate a ramp+pulse discharge waveform (of spikes/sec) for the eye muscles:
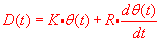 with
a position and a velocity component built in. The OMN waveform will look like
with
a position and a velocity component built in. The OMN waveform will look like
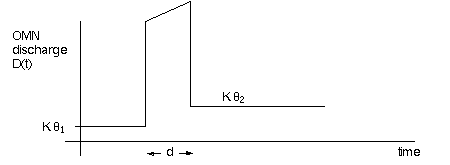
But as you can see from the graph above, going 400 deg/sec is
only half the problem. The other half is stopping precisely on time, when the
target is foveated. Saccade "pulse" timing must not involve visual
feedback (saccades can be made in the dark, to binaurally localized clicks).
And once stopped, the correct hold (fixation signal) must be there to maintain
gaze on the target (the step of pulse-step control).
A timed pulse
One approach: Design a system that converts retinal position error into a pulse
duration. But even if the correct pulse duration is found, the OMN needs at that
moment to reduce its output to the proper "hold discharge" to maintain
fixation at the target. In fact, one way to create a timer out of "analog"
parts is to send a step into an integrator and monitor the ramp output. That's just
what we'll do below, using the integrator in the OMN.
We will pursue an efference copy model, (internal feedback)
model, the one-dimensional bang-bang controller. Redrawing a figure from
RHSC2 chapter 12, p. 360, we have
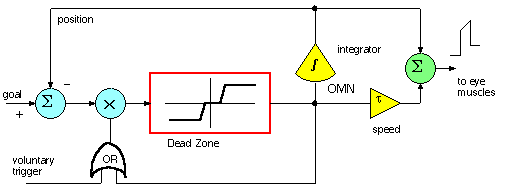
Now the external target position, computed from the retinal signal, is compared
to the internal OMN position integrator and gated by a "voluntary trigger"
as an input to a Dead Zone/Saturation module. If the goal is reached the dead zone
output is zero but the integrator in the OMN maintains postion as a NON-LEAKY integrator.
The saturation limits are +/- 400.
Once the voluntary trigger gets the saccade generator started, then
the feedback from the DZ/Sat unit will keep it going until the Goal-Efference Copy
difference is within the dead zone.
The part on the right, in yellow and green, is the OMN.
One question: If saccades can be made to remembered targets, how
is the recent goal remember? ANS: a sample-and-hold circuit. See Asgn 7 for details
of a Simulink S/H circuit. The S/H circuit will incorporate a pulse generator that
can be suppressed by a saccade generator "busy" signal to make sure nothing
interrupts the saccade in progress.
Single unit firing patterns and motorneuron recruitment
See RHSC2 figure 8.3 for illustration of high fusion frequency (tetanic
frequency) of extraocular muscle. 350 Hz vs 100 Hz for regular muscle. EOM are
fastest of all muscles: 10 msec contraction time vs 40 msec for gastrocnemius.
Next, see Fig 8.10. Look at the k slopes (position) and r slopes
(speed). Notice the thresholds for theta (q) on the position curves. Why aren't
all curves the same? Ans: The need for recruitment, from the smallest,
to the largest force of contraction. With the exception here being that the
mech. load is so small the fast twitch units don't fatigue.
The on direction for q is the same for all motoneurons, but
the threshold varies from cell to cell. In fact each cell has a different r
and k and q0 due to the division into SO and FG muscle types.
Question: does r/k always equal 150 msec for any particular
unit? Yes, because for a saccade all units are recruited for the movement
and the average of all units results in r/k = .6/4 = .150 seconds. each motoneuron
must control a different set of EOM's: comprising unique motor units.
What is q for any particular motoneuron?. Firing threshold
varies from one MN to another, and may be the basis for recruitment
in the oculomotor system. See figure 8.10 of RSHC2. Selecting lower values of
k & r may be part of OMN recruitment, too.
Timing signals for saccades in the CNS: What generates
the duration time of the saccade? It appears that both the superior colliculus
and the frontal eye fields of the frontal cortex can generate saccade timing
signals.
See Fig 12.17 and Fig 12.18: neurons. The nonlinearity ignores
small errors and saturates for larger errors. Try p 363. The model has a trigger
T. "...a brief pulse on T will initiate a saccade only if there is a sufficient
error, and the saccade will continue until the error is adequately dealt with,
when p is turned off."
Pulse-step matching & mismatching
Notice that there is no overshoot or undershoot in actual normal saccades reaching
their destination! Saccades are perfectly calibrated for the correct ballistic
duration. A control signal is needed to "permit" the pulse signal,
or to gate the pulse signal.
What would a saccade waveform look like if the pulse were
too long? too short? if the integrator were disabled? If the gain of the integrator
were too large, or too small? You'll find out by making a Simulink model of
the bang-bang controller. And when we study saccadic dysmetria, we will see
the saccade control system adapt to induced mechanical weakness in medial and
lateral rectus muscles.
The saccade pulse generator
Fig 12.15: RHSC compares a timing generator with an internal feedback mechanism.
Since saccades can be made in the dark, or to remembered targets,
or to moving targets, the second model seems likely. [The
horizontal displacement of the ears means that locating the azimuth of
snapping fingers is computationally straightforward... but what helps figure
out elevation of the sound? See
this website for background...]
Fig 12.16 shows a model for turning retinal position input into
a duration signal, but having weighted synapses so that further out from the
fovea causes a longer duration reaction in the cell.
Opening the loop on saccade control by stabilized images
A staircase of diminishing saccades can result.
Nerve cell correspondence for pulse-step model
See diagram in previous lecture. Of interest: where is the neural integrator?