Hill's equation for muscle tension vs <velocity
of contraction>
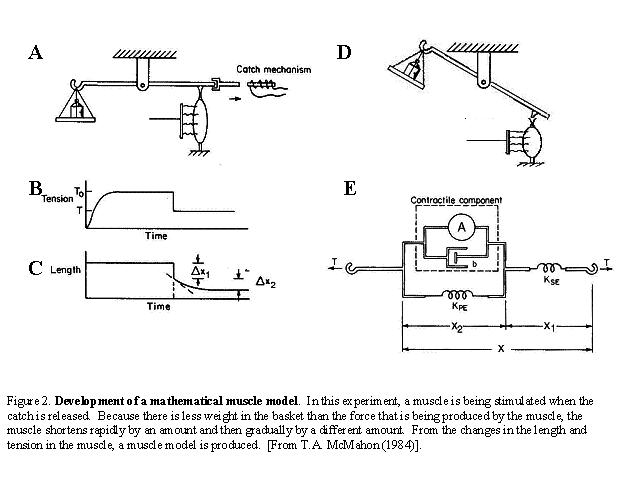
Quick release experiments: Years
before the protein constituents of muscle were known, experiments were done
on the mechanical properties of isolated whole muscle. On page 10 of Muscles
Reflexes and Locomotion (TA McMahon), dynamic ("quick-release")
experiments on isolated muscle are described. A muscle is stretched to a constant
length while stimulated to tetanus, then it is released against a variable load.
 www.shadmehrlab.org
www.shadmehrlab.org
Below are shown a series of quick release length changes as suggested by the
single graph of TAM's fig. 1.8(b).
Notice that there is a lever arm that rotates and that torque is involved in
computing the effective load. The catch mechanism is a solenoid. Such experiments
were started by A.V. Hill in the 1920s and continued by him after the interruption
of WWII.
How to explain the sudden length change Δ x1? It is the result of a series
elastic element (the tendon) that responds "instantly" to a change
in force. There is no "t" in F = k*x.
The slower change to shortening Δ x2 is result of the muscle engaging
and having a "dashpot-like" element, with a rate-of-change of shortening
factor that creates a time constant (like a parallel RC circuit). See
model from TAM Fig 6.7.
Digression to point out the gamma fiber feedback of muscle
tension by Golgi tendon organ and the Ia fiber length change detector wrapped
around the muscle spindle, or intrafusal muscle fiber. Propreoception!
This activity of muscle tension and length provides a "seventh sense"
which you however are not normally aware of. Conscious awareness of joint angle
position come from free nerve endings in joint capsules.
Ia fibers are the basis of the stretch reflex through
the spinal cord...which must be eliminated (example: Lab LT) to study muscle
in isolation. Consideration of the stretch reflex will take us to the next topic
of EN123, Automatic Control.
The v. weak intrafusal muscle spindles can be contracted
by motoneurons, allowing the Ia fiber to extend its range of action and
the extrafusal fibers are shorted and lengthed by external loads and motoneuron
stimulation.
The length Δ x2 below decreases more in
the top graph because the muscle has the opportunity to contract against the
light load. As TAM requests, pay particular attention to the (negative) slope
of the length change at time of release; that's the velocity of shortening,
v. A tricky slope to measure accurately!
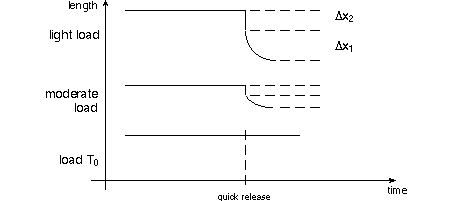
A series of quick release experiments will show an inverse relationship between
load and velocity of shortening. Try to lift a 50lb suitcase as fast as you
can and see how slow your muscles are! A.
V. Hill fitted the data empirically to the equation
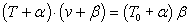
where v is shortening velocity (rate of contraction), T is the variable load
and T0 is the particular load at which the the muscle doesn't change length.
α and β are empirical constants. Since the term on the right contains
neither v nor T, a curve of v vs T has a hyperbolic shape. (What are
the asymptotes of the hyperbola? -α, -β)
It turns out that a wide variety of muscles tested by quick release result
in about the same values of a and b, and that is the mechanical importance
of Hill's equation.
When velocity v is zero then T = T0. (Considering how stimulated by nerve excitation
the muscle is...) .
Maximum velocity of shortening occurs when T=0. (Except when the muscle could
be pulled by an external force...)
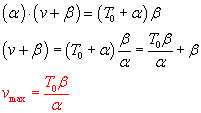
TAM says on p. 24, "For purposes of comparing different muscles, Hill's
equation can be written in a normalized form,"
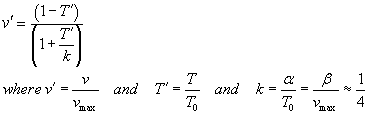
If T0 = Vmax = 1 then a = b = 0.25.
Both v' and T' will be between 0 and 1...
Can you see a way to get from Hill's equation to the normalized form? derivation
Can you show that 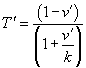 ?
?
This equation for v' results in the plot on TAM page 15. A MATLAB script for
such a plot is
tprime = [-.1 : .01 : 1.7];
k = 0.25;
vprime = (1 - tprime)./(1+tprime/k);
plot(vprime, tprime)
hold on, plot(tprime(11), vprime(11), 'r.')
grid on
Notice the period before the divide symbol; it's needed because tprime is a
vector.
The semicolons after each line suppress printout. In plot the first variable
is the "x-axis." On the normalized plot the tension is 1 at v = 0.
In the plot, a red dot marks v=0.
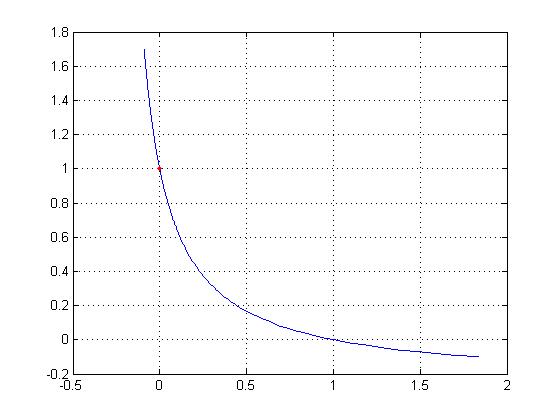
Negative velocity of muscle
Look at the plot of T vs v for Hill's equation, if v becomes negative. The force
continues to increase: individual muscle fibers are "stronger". What
does negative velocity mean? It means the muscle is being stretched during stimulation,
instead of being able to contract.
Building muscle. Such stretching during stimulation pulls apart actin-myosin
bridges and causes "micro-injury". The biochemical result of such
injury is a signal for the muscle to strengthen by increasing in size. Muscle
builders, like Arnold Schwarzenegger, are aware that "doing negatives"
is the only way to build up muscle mass. A body builder lying on his back, on
a bench, and pushing a weight up is bench pressing. While increasing the amount
of weight bench pressed is a goal, a weight lifter almost never lifts weights
in training. A spotter puts the weight in a cradle above the body builder, then
the body builder slowly lowers the weight to his chest, doing a negative. Likewise,
as you lower yourself during a pushup, you're doing a negative. The next day
the muscle forced to negative velocities will be sore, a side effect of the
micro-injuries and the process of developing more muscle.
We'll explore negative muscle velocity and muscle strength with the Tandem
Bike experiment Lab VT.
Power. Power is energy/sec. Energy is force X distance. Therefore power
is force X velocity. If a muscle is not moving it is generating no power. If
the muscle is contracting as fast as it can against no load (no force) it is
generating no power. Somewhere in-between 0 and maximum velocity is maximum
power output for the muscle. In Hill's equation the term T·v has units
of power.
Here's one way to isolate the power T·v in terms of v: Divide both sides
of Hill's equation by v+B:
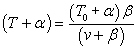
Subtract α from both sides and find a common denominator on the right:
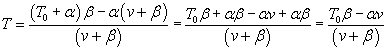
Since T·v = Power, we express power in terms of velocity:
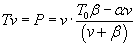
If we let a = b = .25, and T0 = 1, then power calculated and plotted in matlab,
by
vv = [0:0.01:1];
pp = (vv.*(1*.25-.25*vv))./(vv+.25);
plot(vv, pp); grid on
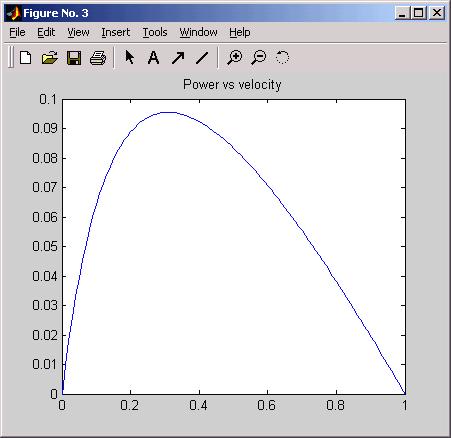
where it seems that maximum power occurs at about one third top speed.
From TAM:

where optimal tension for maximum power is about 3/4 max tension.
If you allow velocity to be negative, with
vv = [-0.1:0.01:1];
you can see that the power of the muscles fibers can be much greater than in
the positive direction.
Bicycling and power: When riding a bicycle with several gears you can
feel the power-velocity relationship as you go up hill, gearing down until you're
pedaling at an optimal rotation in cycles per minute to obtain maximum speed.
Gear down too much and your pedals spin too fast to deliver much power to the
bike chain.
Tandem bike demo: TAM says, "At
a soiree of the Royal Society conducted by Hill in 1952 two subjects mounted
a specially built tandem stationary bicycle. While one subject pedaled forwards
the other resisted him through directly coupled pedals. At a speed of 35 RPM
the subject pedaling forwards used 3.7 times as much oxygen per minute as the
one pedaling backwards. It was demonstrated that a small woman, pedaling
backwards, could rapidly exhaust a large man pedaling forward against her resistance.
Since oxygen consumption and heat production are closely tied ... the muscles
of the woman were presumably generating far less heat, even though the coupling
of the pedals guaranteed that the forces and displacements experienced by the
woman's feet were the same as those for the man's."
(Same amount of work done: force X distance, but woman's muscles were
"stronger", more efficient...)
See Lab VT write-up for more information.
Heat Generated by muscle: In chapter 2 of his book TAM shows how Hill
later was able to derive his equation from thermodynamic considerations (measuring
heat generated by muscle, finding that muscle is about 25% efficient converting
chemical to mechanical energy). Contracting muscle, where crossbridges are being
attached and detached frequently, generates the most heat. The energy-producing
reaction of high-energy ATP to ADP + phosphate produces the heat.
From TAM chpt 2: "Most of what is known about heat production in muscle
derives from the work of A. V. Hill and his collaborators, beginning in 1910
immediately after Hill finished his undergraduate course, and continuing for
sixty years." and more...
A single twitch increases the temperature of a muscle by about 3x10^-3 deg
C...
To begin with, all Hill knew were the changes in heat, not the underlying chemical
reactions...
Hill's glossary:
Resting heat
Initial and recovery heat
Activation heat
Shortening heat: The Fenn effect
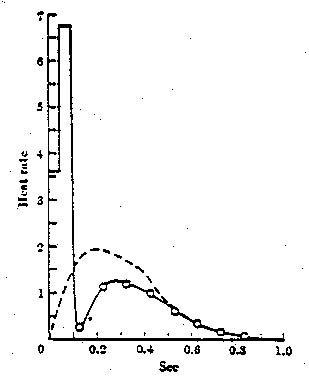
http://www.uic.edu/classes/phyb/phyb516/energeticsu3.htm
TAM p. 34: equation from thermodynamics rearranged
into the mechanical Hill equation. "Hill plotted the rate at which
total energy is liberated in shortening (T+a)*v where v = -xdot...against muscle
tension T..."
Muscle fatigue: We've already noted that fast twitch muscles are easily
fatigued because they use anaerobic metabolism to restore ATP.
We can also tell you that the six eye muscles of each eye are so over-engineered
(over-evolved?) that they cannot be fatigued except by starvation. As a consequence,
the signals from motoneurons to control eye position are an accurate measure
of where the eyes are looking (in the dark, for example) and we humans have
no apparent need for proprioception in the eye muscles (although tendon organs
and muscle spindles are present).
R.H. Fitts, "Cellular Mechanisms
of Muscle Fatigue," Physiological Reviews 14: 49-92 (Jan 1994).
"The major source of acid production in skeletal muscle is the anaerobic
production of lactic acid from glucose and glycogen..."
"It is now well known that fast motoneurons and fast motor units fatigue
more rapidly and to a greater extent than slow muscles and slow motor units..."
Fatigue of sprinters vs marathon runners...
{kind=link}
{kind=link}
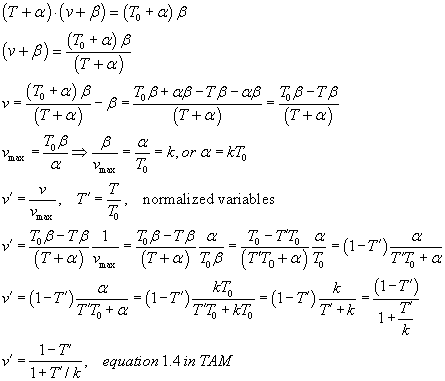{kind=link}