 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 50 NUMBER 4 OCTOBER 2011
Printable (PDF) Version of this issue
CONTENTS Articles and Notes
Acute Pancreatic Necrosis in a Diabetic Long-Tailed Macaque (Macaca fascicularis), by I. Völker & R. Plesker......1
Death During Parturition of a Captive Adult Female Vervet Monkey (Chlorocebus aethiops) and its Social Consequences for a Captive Troop, by A. J. Guy & D. Curnoe ......4
Assessing Video Presentation as Enrichment for Captive Male Pigtailed Macaques (Macaca nemestrina), by G. H. Lee, M. J. Yi, & C. M. Crockett ......7
A Multi-Male Situation in a Population of Predominantly Unimale Bisexual Troops of Hanuman Langurs, Semnopithecus entellus, Around Jodhpur, Rajasthan, by G. Sharma, P. Vijay, Devilal, C. Ram & L. S. Rajpurohit ......10
News, Information, and Announcements
Editor’s Note ......3
Announcements from Publications: IUCN Newsletters and Journals......16
. . .
African Primates; Asian Primates; Lemur News; Neotropical Primates; Primate Conservation
News Briefs......17
. . .
A New Lease on Life for Dieting Orangutan Oshine; Primate Testing – Crucial or Cruel? Orangutans with iPads?
Positions Available......18
. . .
Research Position – Parkinsonism and Aging; Assistant Professor – U.C. Davis
EU Continues Support of European Network on Primate Research (EUPRIM.Net) ......19
Workshops......19
. . .
Pharmaceutical Toxicology Workshop; Environmental Enrichment for Laboratory Animals; EUPRIM-Net Course on “General Primate Biology”
Meeting Announcements......20
Letter from the American Society of Naturalists......21
Information Available......21
. . .
New Name, Same Organization; Animal Nutrition Information; Interesting Websites
Resources Available......22
. . .
All The World’s Primates; The Enrichment Record Poster Repository; Darwin Goes Digital; Orangutan Photos for Orangutan Protection; Chimpanzee Skeletal Digital Atlas (Pan troglodytes); AAALAC Adopts New Position Statements
Help Needed: Handbook on Welfare and Enrichment for Captive Wildlife......23
IPS Awards Granted......24
. . .
IPS Education Grants and Awards; IPS Conservation Grants; IPS Research Grants; IPS Captive Care Grants
Departments
Recent Books and Articles......26
* * *
Acute Pancreatic Necrosis in a Diabetic Long-Tailed Macaque
(Macaca fascicularis)
Iris Völker and Roland Plesker
Paul-Ehrlich-Institut, Langen, Germany
Introduction
Diseases of the exocrine pancreas have only rarely been described in nonhuman primates. Furthermore, terms such as pancreatitis, fibrosis, and necrosis are not always clearly distinguished.
In principle, pancreatic failure in macaques can be the result of viral or parasitic infections, neoplasms, or other reasons (Scott, 1993). Pathological, acute pancreatitis is usually mentioned as acinar necrosis with hemorrhage, cell atrophy, dilation of ducts as well as zymogen depletion. Clinically, indigestion with diarrhea, steatorrhea, dehydration or emaciation might be observed (Brady & Morton, 2008).
The following case study describes a diabetic adult female laboratory long-tailed macaque (Macaca fascicularis) which developed spontaneous acute pancreatitis and necrosis, assumed to have been induced by metabolic fatal fatty liver syndrome (FFLS).
Monkey and case history: The affected individual was a female 13-year-old long-tailed macaque, born on Mauritius and introduced into the primate colony at the Paul-Ehrlich Institute, where it served as a negative control in experiments investigating bovine spongiform encephalopathy. Housing, handling, and experimenta-tion were performed in accordance with European regulations.
Based on a blood glucose level of 336 mg/dl, the macaque was diagnosed at the age of seven with acquired diabetes mellitus type II and was therefore treated with ˝ tablet/day Euglucon N (Aventis Pharma, Germany). This successfully decreased blood glucose to 132 mg/dl, which fluctuated in subsequent years. Four months prior to death, sticky urine was observed, indicating glucosuria, and a blood glucose level of 451 mg/dl blood was found. Treatment was therefore increased to 2 x ˝ Euglucon tablets per day, resulting in a reduction to 170 mg/dl blood glucose (Table 1).
In its final days, the animal rejected medication and displayed loss of appetite, weight loss, and dehydration, and its vomit contained flecks of blood. The monkey was therefore euthanized by intracardial injection of a ketamine (Ketamin 10%, Bela-Pharm, Vechta, Germany)-xylazine (Rompun 5%, Bayer, Leverkusen, Germany) mixture.
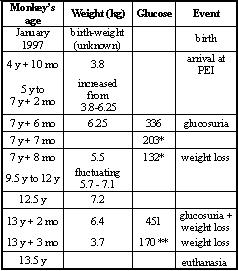
Table 1: Clinical data of a diabetic long-tailed macaque with acute pancreatic necrosis.
y = years; mo = months; Glucose = blood glucose in deciliters; * = treated with ˝ tablet Euglucon®/day; ** = treated with 2 x ˝ tablet Euglucon®/day
Immunohistochemistry for differentiation of B- and T- lymphocytes was performed with antibodies specific for the CD3 antigen using the PAP method (T-cells) and for CD79α using the ABC method (B-cells).
Clinical chemistry: Clinical chemistry was carried out with serum from the affected macaque (Vettest 8008, Westbrook, USA) and the blood glucose level was determined using strips for blood samples (Accu-Check® Comfort, Roche, Mannheim).
Hematology: The blood cell count was determined using a CellDym Abbott CD3500SL (Abbott Diagnostics, Vienna, Austria).
Necropsy: The carcass showed high-grade exsiccosis and moderate muscle atrophy. An acute pancreatic necrosis was seen with beige-yellow adherent masses in the upper abdomen that appeared to be covered in fibrin. Residual beige streaks of pancreatic tissue interrupted by yellow-beige-reddish dots were present on surfaces formed by cutting (Fig. 1). A discharge of white lymph was seen on some other cut surfaces in the body.
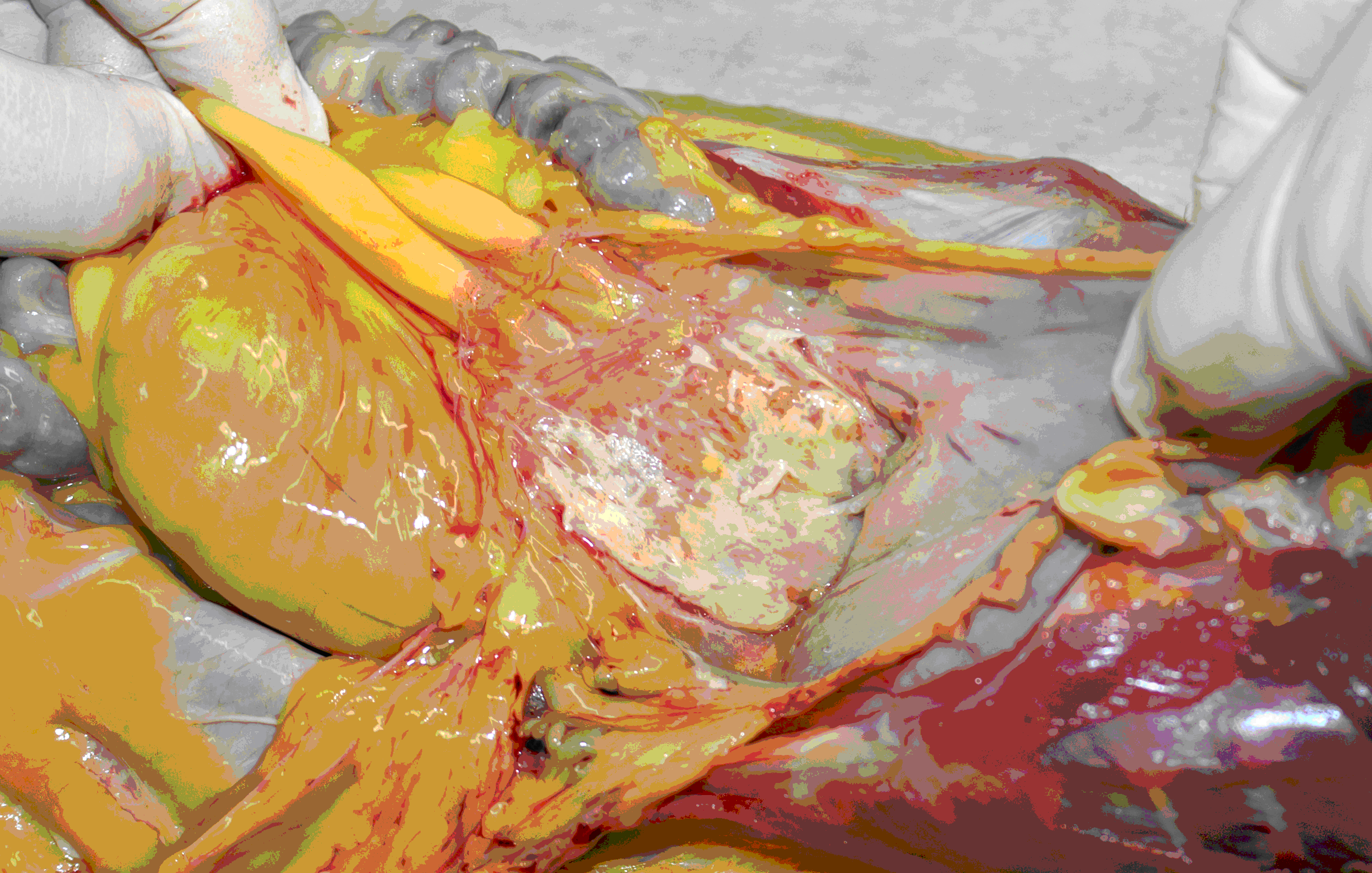
Figure 1: Pancreas; long-tailed macaque, macroscopic appearance (situs) of the upper abdomen.
Histology: In the pancreas, unaffected areas and those with high-grade acute necrosis were found adjacent to each other (Fig. 2). Edema and areas of fibrosis were observed. The perivascular regions and islets of Langerhans were moderately infiltrated with B- and T-cells. The serosa and submucosa of different abdominal organs displayed necrosis. An amyloid deposition, as determined using a congo red stain, was also present in the islets of Langerhans (Fig. 3). No adenovirus-induced inclusion bodies were detected.
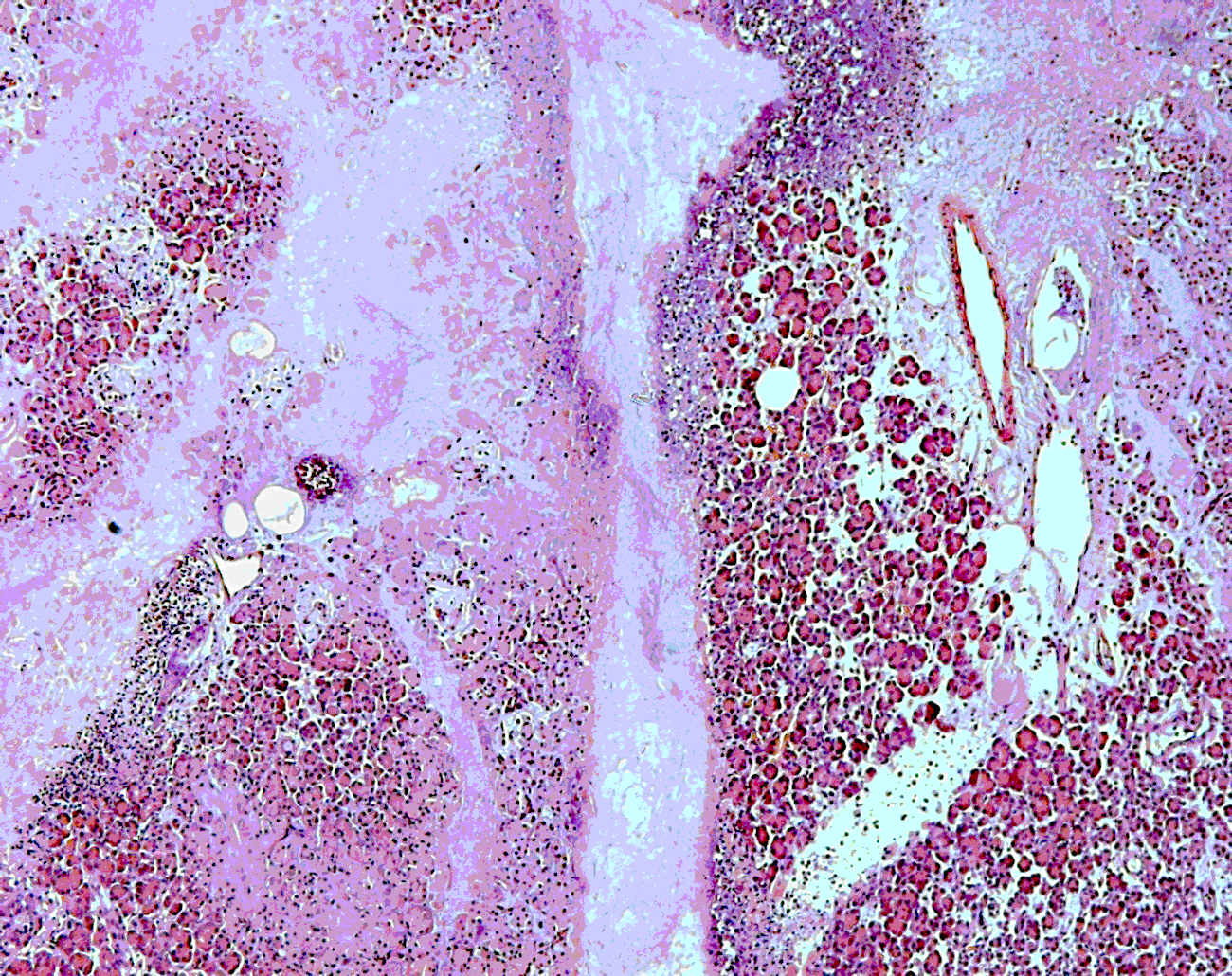
Figure 2: Pancreas; long-tailed macaque, homogenous eosinophilic masses due to necrosis: hematoxylin-eosin stain.
Spleen follicles were atrophic and the red pulp appeared rich in cells.
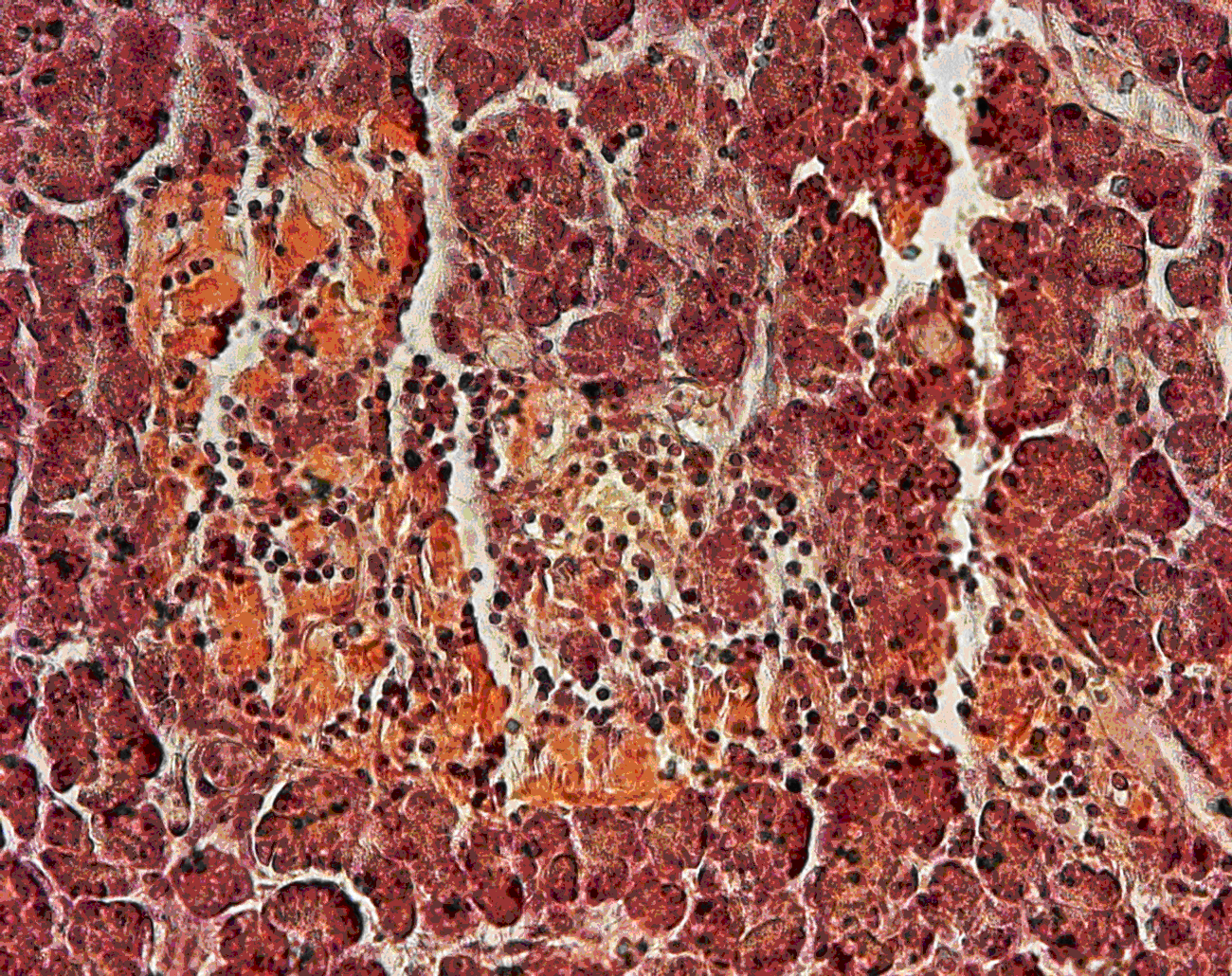
Figure 3: Pancreas; long-tailed macaque, amyloid deposition and lymphocyte infiltration in islets of Langerhans: congo red stain.
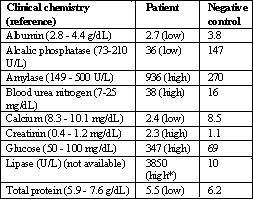
Table 2: Clinical chemistry of a diabetic long-tailed macaque with acute pancreatic necrosis.
dL = deciliter; U = units; L = liter; * = assumed high
Hematology: Lymphopenia, eosinophilia and low hematocrit were detected in hematology (Table 3).
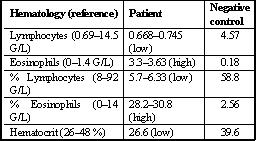
Table 3: Hematology of a diabetic long-tailed macaque with acute pancreatic necrosis.
G/L = gram/ liter (human references)
Discussion
This case had three main pathological characteristics: diabetes mellitus type II, a fatty liver, and a severely necrotic pancreas. Although a fatty liver is often linked to acquired diabetes mellitus, it is also the dominant feature of metabolic FFLS. In addition, this syndrome can also be responsible for an acute pancreatic necrosis (Gliatto & Bronson, 1993). Clinical observations such as depression and weight loss in connection with a predisposition with regard to species (cynomolgus), sex (female), and age (mean 9 years) might argue for FFLS and we therefore propose that this was the most likely cause of the disorder described here. Extremely elevated serum values of pancreatic enzymes such as amylase and lipase correlated with the pathology of the pancreas in this case and the hypoalbuminemia and protein loss can confirm alterations in the liver.
Although differential toxic agents might cause similar alterations, long-term treatment with Euglucon N is not known to induce such lesions. However, an infectious causative agent for pancreatic necrosis must also be considered.
In conclusion, the pancreatitis resulted in leakage of pancreatic enzymes into the abdominal cavity which in turn affected different organs. A severe acute pancreatic failure such as this generally has a poor prognosis.
Brady, A. G., & Morton, D. G. (2008). Digestive system. In: B. T. Bennet, C. R. Abee, & R. Henrickson (Eds.), Nonhuman primates in biomedical research (pp. 406-407). New York: Academic Press.
Gliatto, J. M., & Bronson, R. T. (1993). Fatal fasting syndrome of obese macaques. In T. C. Jones, U. Mohr, & R. D. Hunt (Eds.), Monographs on pathology of laboratory animals: Non-human primates I (pp. 198-202). Berlin and New York: Springer Verlag.
Scott, G. B. D. (1993). The exocrine pancreas. In G. B. D. Scott, Comparative Primate Pathology (pp. 183-194). Ames, Iowa: Iowa State Press.
-------------------------------------------
Roland Plesker, Section 4/ZT, Paul-Ehrlich-Institut, Paul-Ehrlich-Str. 51-59, 63225 Langen, Germany [e-mail: [email protected]].
Acknowledgments: Edgar Holznagel, Cheick Coulibaly, Carina Kruip, and Kernt Köhler.
-------------------------------------------
* * *
It is very mixed feelings that I have at this time: sadness that this interesting and satisfying job is over; relief that this difficult and worrisome task is finished; uncertainty about what the future will bring; and, mostly, gratitude to the many people who have helped me in so many ways. I cannot possibly mention all of you, and I don’t want to mention some names and leave others out, so, without naming names, I thank you all!
I must also mention my gratitude to the many nonhuman primates who have been the subjects (and objects) of this journal and of the studies and experiments described in it. My greatest hope has always been that we have been able to make their lives easier, especially because many of the results of those studies and experiments have been to make the lives of human primates, and other animals, easier and better. Let me extend my thanks not only to those animals, but to the humans whose work has been to make the lives of experimental animals easier!
* * *
Background
Vervet monkeys (Chlorocebus aethiops) live in multi-male troops where males disperse and females are philopatric (Cowlishaw & Dunbar, 2000). A troop of vervet monkeys can consist of several lineages of related females, their offspring, and relatively permanently associated males who are unrelated (Lee, 1989). Vervet troops are characterized by linear dominance hierarchies and coalitions (Struhsaker, 1967). Males have strong dominance relationships that determine access to mates, and females are strongly female-bonded, as demonstrated through frequent female–female grooming, most of which is between closely related females of similar rank (Dunbar & Barrett, 2000). Relative rank is quite stable for both sexes, but female ranks tend to be more stable than male ranks (Bramblett and Bramblett, 1982).
Females stay in their natal group and form grooming relationships with relatives. They are strongly territorial and will join forces to drive away rival groups (Dunbar & Barrett, 2000). Within a troop, females are ranked based on the direction of approach–retreat interactions as well as dyadic interactions including biting, avoiding, chasing, and supplanting behaviors (Bramblett & Bramblett, 1982). High-ranking females are consistently able to exclude lower-ranking females from access to food and water (Cheney et al., 1981). The largest and most aggressive animals are usually dominant, but female vervet monkeys also inherit their mothers’ ranks (Dunbar, 1988).
Coalitions form an important part of interactions between vervet females. Low-ranking females often attempt to interact with high-ranking females to improve their competitive abilities. These interactions can facilitate tolerance of the lower-ranking females at feeding sites and gain them support in aggressive encounters (Cheney et al., 1981). Kinship is an important basis for formation of coalitions, but these can also form between unrelated individuals when it is advantageous. Nonhierarchical alliances can also occur (Dunbar, 1988). Larger troops often consist of cliques that interact with and support each other more than with other group members (Dunbar, 1989). Relationships with members outside the clique may be more often antagonistic (Dunbar, 1989).
Aggression in vervet monkeys includes behaviors such as threatening facial expressions, ground slaps, lunges, chases, and occasionally bites (Cheney & Seyfarth, 1986). Most of these behaviors do not involve physical contact and form the majority of aggressive interactions. Grabbing or slapping toward each other without actual physical contact acts as both an aggressive and a defensive gesture. Actual slapping or grabbing is considered extreme aggression in vervet monkeys and is usually accompanied by bites (Struhsaker, 1967).
Aggression among female vervet monkeys has been reported by Fairbanks & McGuire (1986) to increase following social disruption such as the death of the alpha female. However, details of the circumstances of the death and the troop interactions that followed were not published. Observers and managers of captive troops should be trained to recognize such rare critical events and be prepared to act quickly in order to avoid or limit morbidity and mortality.
The current study reports the death of a captive female vervet monkey during parturition and the events that followed, with the aim of reporting in detail the behavior of the troop during the unusual parturition and the social consequences of the death of an alpha female.
A behavioral study of vervet monkeys was conducted by one of the authors (A. G.) in September, 2010, at Wild Animal Trauma Centre and Haven (WATCH), located in Vryheid, KwaZulu-Natal, South Africa. This Centre plays a key role in the rehabilitation and release of vervet monkeys in South Africa. The Centre is home to three troops of vervet monkeys at various stages of rehabilitation, and has recently released two troops to a conservation area in KwaZulu-Natal.
Two days after the study commenced a pregnant female in one troop died. This female was known to be the alpha, supported by behavioral cues such as supplanting other troop members from food, grooming, and resting sites. As the death occurred soon after formal data collection for the behavioral study had commenced, there was insufficient data to compare quantitative data on time budgets before and after. For this reason, data for the current study is limited to general observations made during the alpha female’s difficult labor and over the five weeks following her death. Aggressive interactions were identified directly from observed behaviors such as threatening facial expressions (in this case, raising eyebrows), chasing, lunges, and biting, and indirectly from injuries to individuals. Quantitative behavioral data for the troop will be presented in a later study of time budgets in vervet monkeys.
The subjects of this study were a troop of vervet monkeys, housed at WATCH. The troop consisted of 27 individuals including two adult females, one adult male, two subadult males, four subadult females, and 18 juveniles. These captive primates included hand-raised orphans and injured wild monkeys that had recovered and been integrated into the troop in preparation for release. The troop was housed in a natural enclosure enriched with grass, trees, and climbing structures, with a ground area of 306.72 m² and a height of 3.2 m. Diet consisted of a mixture of fresh fruit and vegetables, supplemented with protein-rich foods such as eggs and nuts. Water was available ad libitum.
The pregnant alpha female was first observed to be in labor early in the morning of the 15th of September, 2010. She appeared to be in the birthing process but was clearly uncomfortable and appeared distressed. The decision was made to monitor her condition as it was unknown if intervention might be necessary, since a daytime birth had not previously been observed at the Centre. Subadults and juveniles of both sexes were observed approaching and grooming the female, inspecting her genital area and lifting her tail. The female stayed in vocal contact with the troop, and the adult male exhibited threatening behaviors towards human observers. After approximately three hours, approaches by other troop members became more frequent. By this point, the female appeared to be exhausted and it was suspected that she had been in labor throughout the night, since vervet monkey births normally occur at night (Fairbanks & McGuire, 1984). Intervention was avoided at this point as it was thought that it might cause additional stress. By late morning, the birth had not progressed any further and the decision was made by Centre staff to capture her and consult a veterinarian, but the female died before medical intervention could occur.
Following this event, aggression began among the remaining troop members. There was only one adult female remaining. She was thirteen years old and had always been submissive, as indicated by her being supplanted from food and resting locations by all members of the troop. It was suspected that these factors contributed to her being unable to take over the troop. Instead, one of the subadult females became the apparent alpha, as was evidenced by her ability to easily displace the adult female, other subadult females, and juveniles from resting positions, grooming, and food; her ability to force grooming with apparently unwilling participants; and her priority of access to food – where she was also capable of excluding all troop members from one of the three food bowls, making it “her own” during a feeding. Aggression escalated and resulted in severe injury to three juveniles (two males and one female) who had to be removed from the group in order to recover. Injuries were sustained to the forearm, back, underarm, and leg. All these injuries were severe enough to require ongoing treatment, and all bled profusely and reopened when the monkeys ran around in the enclosure.
It appeared that there were at least two subgroups fighting for dominance within the troop. The primary aggressors appeared to be two subadult males, who were even seen directing aggression toward the only mature adult male in the troop. Subadult and juvenile females were also observed to be aggressors. This was still continuing six weeks after the alpha female’s death (when the primary study was complete). By ten weeks, reports from WATCH indicated that the increased level of aggression had subsided, but it was unclear which female was the new alpha of the troop. The subadult female mentioned earlier was still attempting to maintain dominance; however the adult female was seen frequently in the company of the adult male and one of the subadult males.
The death of the alpha adult female and the abnormality of the daytime labor would suggest that any vervet monkey observed giving birth during the day should be taken for veterinary assistance, unless it is obvious that the birth is progressing normally. This is supported by an eight-year study of captive vervet monkeys, in which it is recorded that, of 77 births, only two occurred during the day and both resulted in the death of the mother (Fairbanks & McGuire, 1984).
As vervet monkeys are characterized by linear hierarchies (Struhsaker, 1967), it was expected that the only other adult female would move up to take the alpha’s position. Due her age and submissive nature, this did not happen immediately and the subadult female moved up in rank instead. While it was expected that the subsequent aggression would return to normal levels soon afterwards, this did not occur. The aggression between members of the troop escalated and was considered to be extreme, as physical aggression in vervet monkey troops is uncommon (Struhsaker, 1967). Such severe aggression has rarely been seen at this Rehabilitation Centre, except when adult males are housed together. This level of aggression was sustained for ten weeks after the death of the alpha female.
We suspected that the dominance relationships could not be clearly reestablished in the absence of a dominant adult female. The subadult female attempted to take over the alpha position, but as aggression in the troop continued even after her dominant status seemed clear, it appears that this was not sufficient to reestablish an orderly hierarchy. Even after ten weeks, the female hierarchy remained unclear. The fact that subgroups appeared to be fighting for dominance supports previous studies which suggested that coalitions are very important in obtaining and maintaining dominance status (Cheney et al., 1981). It also suggests that a strong adult female is vital to maintaining order within the troop, and that multiple adult females may be required to form effective coalitions.
Subadult males were observed to be the aggressors on a number of occasions. It is possible that the males were assisting females that they preferred in aggressive encounters so that such support might be reciprocated. Alpha females are known to influence the ranking of male vervet monkeys (Raleigh & McGuire, 1989). So, by assisting a particular female to become the new alpha, a subadult male may improve his chances of becoming the alpha male of the group.
It is also possible that the artificial conditions of captivity have played a role in this situation. Captivity greatly limits opportunities to flee and hide and so may have contributed to the sustained high levels of aggressive behavior observed in this study. In addition, the lack of relatedness among troop members, as a result of individuals coming into care at different times, may have delayed the establishment of a new dominance hierarchy. Females in wild troops are closely related (Lee, 1989) and rank tends to be inherited – females tend to find their places in the hierarchy adjacent to their kin (Dunbar & Barrett, 2000). Kin will also form coalitions, supporting each other to increase their competitive capabilities (Dunbar & Barrett, 2000). However, while kinship is an important basis for formation of coalitions associated with dominance relationships, these can also form between unrelated individuals when it is advantageous (Dunbar, 1988). Clearly, unrelated individuals are capable of forming coalitions, since subgroups in the study troop were observed interacting aggressively. However, multiple adult females may be required in order for this to be effective. As only one adult female was present in the troop, this may have further delayed the establishment of a new dominance hierarchy.
In conclusion, the death of the alpha female was highly disruptive to the social hierarchy of this troop of captive vervet monkeys. Despite the formation of coalitions and the apparent dominance of one subadult female, the position of alpha female remained in question ten weeks after the death of the previous alpha. Multiple adult females may be required to reestablish a linear dominance hierarchy.
Bramblett, C. A., & Bramblett, S. S. (1982). Longitudinal stability in adult status heirarchies among vervet monkeys (Cercopithecus aethiops). American Journal of Primatology, 2, 43-51.
Cheney, D. L., Lee, P. C., & Seyfarth, R. M. (1981). Behavioral correlates of non-random mortality among free-ranging female vervet monkeys. Behavioural Ecology and Sociobiology, 9, 153-161.
Cheney, D. L., & Seyfarth, R. M. (1986). The recognition of social alliances by vervet monkeys. Animal Behaviour, 34, 1722-1731.
Cowlishaw, G., & Dunbar, R. (Eds.). (2000). Primate conservation biology. Chicago: The University of Chicago Press.
Dunbar, R., & Barrett, L. (2000). Cousins: Our primate relatives. Walton, B. (Ed.). London: BBC Worldwide, Ltd.
Dunbar, R. I. M. (1988). Primate social systems. New York: Cornell University Press.
Dunbar, R. I. M. (1989). Social systems as optimal strategy sets: The costs and benefits of sociality. In V. Standen & R. A. Foley (Eds.), Comparative socioecology: The behaviour and ecology of humans and other mammals (pp. 131-150). Oxford: Blackwell Scientific Publications.
Fairbanks, L. A., & McGuire, M. T. (1984). Determinants of fecundity and reproductive success in captive vervet monkeys. American Journal of Primatology, 7, 27-38.
Fairbanks, L. A., & McGuire, M. T. (1986). Age, reproductive value, and dominance-related behaviour in vervet monkey females: Cross-generational influences on social relationships and reproduction. Animal Behaviour, 34, 1710-1721.
Lee, P. C. (1989). Family structure, communal care and female reproductive effort. In V. Standen & R. A. Foley (Eds.), Comparative socioecology: The behaviour and ecology of humans and other mammals (pp. 323-340). Oxford: Blackwell Scientific Publications.
Raleigh, M. J., & McGuire, M. T. (1989). Female influences on male dominance acquisition in captive vervet monkeys, Cercopithecus aethiops sabaeus. Animal Behaviour, 38, 59-67.
Struhsaker, T. T. (1967). Social structure among vervet monkeys (Cercopithecus aethiops). Behaviour, 29, 83-121.
-------------------------------------------
Authors’ address: School of Biological, Earth and Environmental Sciences, University of New South Wales, Syd-ney, NSW, Australia 2052 [e-mail: [email protected]].
This research was approved by the Animal Ethics Committee of the University of New South Wales (approval number 10/70B). The study was also registered with Ezemvelo KZN Wildlife in South Africa.
We would like to thank Bruce and Sandi Cronk of WATCH Rehabilitation Centre for hosting Amanda Guy for field work and sharing their views on this topic. We would also like to thank the University of New South Wales for access to facilities and for financial support.
-------------------------------------------
* * *
Introduction
Environmental enrichment is an important part of animal husbandry. The goal is to improve the welfare of the animals by providing stimuli that increase mental engagement, alleviate boredom, and reduce stress. Enrichment also has the benefit of encouraging species-specific behaviors while reducing or extinguishing abnormal behaviors (Mellen & MacPhee, 2001). For any captive animal, administering enrichment that is relevant to the species and helpful to the individual can be challenging. In lab and research settings it is even more difficult because research protocols often place limitations on diet and social contact.
A device ubiquitous in modern human life, the television, can be utilized as environmental enrichment for captive primates. Platt and Novak (1996) found that group-housed rhesus macaques (Macaca mulatta) watched a television passively and would also manipulate a joystick to play a video game that dispensed food rewards. Also, they found that activity increased and that monkeys attended to their surrounding environment more.
Andrews and Rosenblum (2002) provided a joystick task for bonnet macaques (Macaca radiata) to choose between a food treat or watching a video of conspecifics in a social group. Two of the three monkeys preferred the video over the food treat. A compelling finding from their study is that over the 75-week study period the video rewards choice decreased and the food rewards choice increased.
Bloomsmith and Lambeth (2000) studied chimpanzees (Pan troglodytes) and their responses to different videotape conditions: conspecifics, humans and other animals, and television programs. The amount of time spent watching videos of chimpanzees was not significantly different from videos of humans. Chimpanzees watched any video significantly more than a blank screen. Individually housed chimpanzees watched the monitor more than socially housed chimpanzees.
We set out to assess the effectiveness of audio/video enrichment for pigtailed macaques (Macaca nemestrina) by exposing two singly housed males to a video of The Lion King (Disney, 1994). We evaluated whether the macaques would attend to the video and how the presence of the video affected their behavior. We hypothesized that the subjects would attend to the video and that it would decrease the rate of abnormal behavior initially but that over time the subjects would become habituated to the stimulus.
Subjects and Experimental Setup: The subjects were two nine-year-old male pigtailed macaques (Macaca nemestrina) housed at the Washington National Primate Research Center at the University of Washington. Neither subject had a history of serious abnormal behavior, but were being monitored for overgrooming (i.e., pulling or plucking hair with hands or mouth, resulting in removal of hair). Subjects A and B were individually housed in 0.95 x 0.8 x 1.0 m single-tier cages positioned next to each other. Prior to the study they had been housed next to each other for nine months. For 1.5 of those months, they were housed in grooming contact (tactile contact through widely spaced bars), but this contact ended due to incompatibility. Both had visual, auditory, and olfactory contact with each other and with other monkeys in the room. Animals had free access to water and were fed monkey chow biscuits twice daily, once before 9:00 a.m. and once after 2:00 p.m. For the duration of the experiment, subjects received their normal enrichment which included toys, a perch, and daily fresh produce or forage (provided via puzzle feeders, or items such as frozen treats in paper lunch sacks).
The experiment lasted five weeks and went through four phases: pre-baseline (1 week), habituation (1 week), video or blank (that is, power-off) screen exposure (2 weeks), and post-baseline (1 week). Four unobtrusive recorded observations were done each week between 12:00 p.m.–12:30 p.m., a quiet time of day between routine husbandry activities. A Sony Handycam on a tripod placed 2 m from the subjects was used to collect video recordings of both subjects. Baseline phase observations were made with no television present. In the habituation phase, a 13-inch color television/VCR combo was placed 1.5 m directly in front of the subjects atop a yellow barrel that raised the screen to the monkeys’ eye level, but no video was shown. This was to control for any novelty effect caused by the presence of the equipment. During the video exposure phase, subjects were shown either a videotape or a blank television screen on alternating days, starting with the video condition. On videotape days, the animated movie The Lion King (Disney, 1994) was shown at a low volume and always from the beginning of the movie. The video was shown for a half-hour twice per week for two weeks. All equipment specific to this experiment was set up immediately before each observation and removed afterwards.
Data Collection and Analysis: Behavioral data for each subject were sampled from twenty minutes of each recorded observation, using continuous sampling to record the duration of various behaviors. The first five minutes of each recording were excluded from sampling. In order to control for order effects, the subject that was coded first was alternated for each day’s data. One researcher coded all the video files and was blind to the phase condition because a second researcher had relabeled the filenames with random numbers.
The eight behavior categories used in this study were: abnormal (stereotypies, overgrooming, or self-abuse), inactive, other activity (locomoting, eating/drinking, miscellaneous active behaviors not included in other categories), self-grooming, manipulating toys or cage, affiliative, agonistic/tense (e.g., cage shakes, head bobs), and attending to television (eye gaze directed at the television).
During coding, we kept the recorded behaviors mutually exclusive in order to score the twenty minutes continuously; however, the subject sometimes presented multiple behaviors simultaneously. When this occurred, the new behavior interrupted the recording of the previous behavior (behaviors were recorded from their onset and not necessarily until they extinguished).
The data collected show differences in behavior between the two subjects. In general Subject A performed much more agonistic/tense behavior than Subject B. Subject B performed a mild cage shake only once during the entire study, during the exposure phase. Across all phases, subject A performed eleven cage shakes and two head bobs, including one head bob directed to the monitor while the video was showing. Behaviors within each subject varied across days and phases and did not show any obvious pattern of change from before, during, and after video exposure phases.
Attending to the Video: Both subjects attended to the video more than the blank screen initially (Day 1: Subject A 25% of the time and Subject B 34%, compared to 0% and 16% toward the blank screen, respectively). The amount of time spent attending to the video quickly declined during subsequent video exposures (Day 2: A = 8%, B = 15%; Day 3: A = 2%, B = 3%; Day 4: A = 2%, B = 1%) (Fig. 1).
Video Effects on Behavior: For both subjects, there was a decrease in self-grooming on Day 1 of video exposure. Other behaviors like affiliative, manipulate, and agonistic/tense did not follow any pattern. Subject A did direct a head bob to the video once; however that was the only agonistic/tense behavior that seemed to be related to the video. Abnormal behavior was never observed during video or blank phases.
Habituation: Both subjects exhibited rapid habituation to the video over the four days of exposure. Pearson’s correlation test between attending to video and days of exposure shows that the decrease approaches significance only for Subject B (Pearson r = -0.947, p ≤ 0.053 versus r = -0.887, p ≤ 0.113 for Subject A). By the last video exposure, attention to the television totaled less than half a minute (out of 20 coded minutes) for each subject. The two subjects’ video observation durations over the four exposures were highly correlated with each other (r= 0.986, p≤0.014).
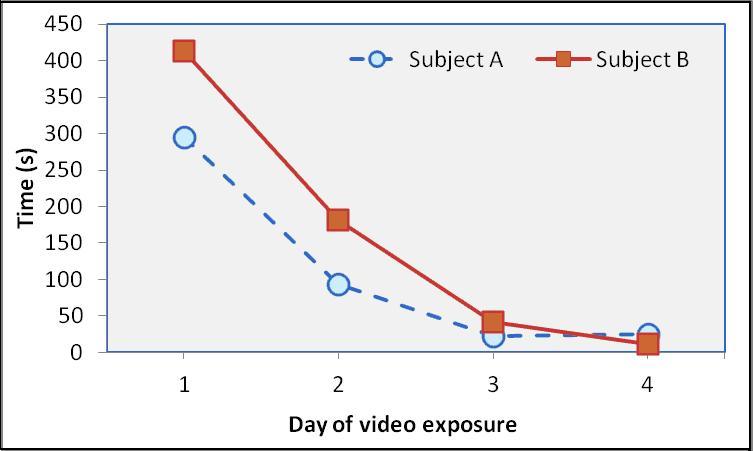
Figure 1: Comparison of amount of video observation between Subjects A and B.
Discussion
The results of this experiment support the hypothesis that the subjects would attend to the videos at first. But even during a quiet time of day with little distraction, our subjects did not redirect much of their activity budget towards the television. While there were no positive changes in behavior, video presentation of an animated movie did not increase any undesirable behaviors such as stereotypy or tension. Repeated animated video presentation did not appear to have any conspicuous long-lasting effects on behavior. Of the measured behaviors, only attention to video showed any consistent pattern. Both subjects showed rapid and dramatic habituation to video presentation over only four exposures; their significantly correlated response suggests there is little to no use of repeated animated video as enrichment for singly housed adult male pigtailed macaques.
Given that our sample size of four days of exposure was so small, it is not surprising that the patterns of declining time spent observing the video by each Subject did not prove significant. Four exposures to video over two weeks is a very short sample time for monkeys that may spend years in a cage. The fact that the subjects were tested in the room they are normally housed in increases the generalizability of this study. A follow up study with more subjects and more days of video versus blank exposure would increase statistical power to test the hypothesis. However, the habituation to repeated video exposure was so striking that a larger study should be not be done without changing some of the parameters.
Since other studies have found a difference in habituation in males vs. females (Platt and Novak, 1996) any follow up study should include both sexes. Another avenue for future exploration is to apply video presentation to a specific demographic such as infants or juveniles being raised in a limited social environment, or animals that need to be deprived of stimuli that they normally have access to (e.g., an animal requiring clinical treatment may be moved to a special housing area) as these animals may show greater benefit. O’Neill-Wagner (2005) reported that rhesus macaques in a hospital setting all directed some interest toward assorted videos (including an animated movie), but there was no control group for comparison and it was not known whether video had clear benefits such as distracting the macaques from picking at their bandages, etc. Another parameter to vary is type of video; the present study only showed animation and these monkeys might not have recognized animated characters as more than colorful moving shapes. Video of conspecifics or people might elicit more interest.
Our study intentionally repeated the same segment so as to examine habituation. A future study might try switching videos in a single session and seeing if attention increases when the video changes. This could help determine if the subjects are becoming habituated to the movie itself or video enrichment in general. Perhaps frequent rotation of varied video types will provide the aspect of novelty that is needed to maintain interest (Taylor et al., 1997). In a study of video presentation to gorillas where subjects viewed multiple types of videos on each occasion, habituation did not occur (Maloney, 2011). Bloomsmith (2000) found that chimps exposed to repeated presentation of videotapes habituated over time, but this was in the course of several months. Harris et al. (1999) spent six weeks training eight individually housed rhesus macaques to lever-press in order to activate a television. Only two of the eight monkeys learned the task, suggesting that a video of conspecifics and humans in familiar environments is not a very valuable reward. Further evaluation with the two subjects who had learned the task revealed that demand for television appeared highly elastic.
This preliminary study has demonstrated that our two subjects rapidly lost interest in a repeated animated video. Their clear and similar response suggests little usefulness of this form of video enrichment for adult male pigtailed macaques.
We would like to thank the Psychological Well-Being program at the Washington National Primate Research Center. Thanks to editors of LPN for constructive comments. M. Yi conducted this study in conjunction with receiving undergraduate research credits in the University of Washington Department of Psychology. Partially supported by NIH grant RR00166.
Andrews, M. W., & Rosenblum, L. A. (2002). Response patterns of bonnet macaques following up to 75 weeks of continuous access to social-video and food rewards. American Journal of Primatology, 57, 213-218.
Bloomsmith, M. A., & Lambeth, S. P. (2000). Videotapes as enrichment for captive chimpanzees (Pan troglodytes). Zoo Biology, 19, 541-551.
Harris, L. D., Briand, E. J., Orth, R., & Galbicka, G. (1999). Assessing the value of television as environmental enrichment for individually housed rhesus monkeys: A behavioral economic approach. Contemporary Topics in Laboratory Animal Science, 38, 48-53.
Maloney, M. A., Leighty, K. A., Kuhar, C. W., & Bettinger, T. L. (2011). Behavioral responses of silverback gorillas (Gorilla gorilla gorilla) to videos. Journal of Applied Animal Welfare Science, 14, 96-108.
Mellen, J., & MacPhee, M. S. (2001). Philosophy of environmental enrichment: Past, present, and future. Zoo Biology, 20, 211-226.
O’Neill-Wagner, P. (2005). Video entertainment may facilitate recovery for monkeys in a clinical setting. In F. F. McMahon, D. E. Lytle, & B. Sutton-Smith (Eds.), Play & Culture Studies: Vol. 6. Play: An Interdisciplinary Synthesis (pp. 43-51). Lanham, MD: University Press of America.
Platt, D. M., & Novak, M. A. (1997). Videostimulation as enrichment for captive rhesus monkeys (Macaca mulatta). Applied Animal Behaviour Science, 52, 139-155.
Taylor, W. J., Brown, D. A., Davis, W. L., & Laudenslager, M. L. (1997). Novelty influences use of play structures by a group of socially housed bonnet macaques (Macaca radiata). Laboratory Primate Newsletter, 36 [1], 4-6.
-------------------------------------------
Authors’ address: 1705 NE Pacific St, Seattle, WA 98195 [e-mail: [email protected]].
-------------------------------------------
* * *
Introduction
Hanuman langurs exhibit clear-cut sexual dimorphism. On average, adult males weigh 18.5 kg and adult females weigh 11.7 kg (Sommer 1985). Males defend infants but never carry or feed them.
Multi-male bisexual troops are very rare in the Jodhpur area. Each troop has a home range of about 0.5–1.3 km². The home ranges of multi-male bisexual troops are larger than those of unimale-bisexual troops. Females remain for life in their natal troops, but males emigrate, usually as juveniles, to unisexual all-male bands, whose home range can be as large as 20 km² (Rajpurohit, 1987; Rajpurohit & Sommer, 1993; Rajpurohit et al., 1994).
As multi-male bisexual troops form, males migrate from their troops and, after some time, return. The ranging area of these males is also larger in comparison to alpha males and females. The multi-male troop is created in two ways: first, when male juveniles stay in their own troop and alpha males have no objection and don’t try to chase them out; and second, when an outside male or group of males comes to take over the bisexual troop and a multi-male situation arises. Such a situation may go on for a few months to more than a year.
In Jodhpur langurs, a multi-male troop structure may develop during an alpha male change, but that is usually temporary, and the troop becomes one-male again. There have been some cases observed where troops developed semi-permanent multi-male structures (Srivastava, et al., 1986; Mohnot, et al., 1987; Rajpurohit, 1987).
In the multi-male societies of red colobus (Procolobus) and Himalaya Hanuman langurs, the dominant adult males achieved the most mating and presumably therefore sired the most offspring (Struhsaker, 1975; Boggess, 1980). However, Laws & Laws (1984) found that in the Himalayan foothills, Hanuman langur males, temporarily immigrating into bisexual troops, were as successful at mating as the alpha.
In matrilineal, multi-male colobine societies, there is little affiliative social contact among adult males. Within multi-male troops of Himalayan Hanuman langurs, agonistic vocal interaction occurs between males much more frequently than between females. Boggess (1980) found that only 0.3% of adult grooming interactions were between males. Dunbar (1988) suggested that in Ethiopian Colobus grereza there is a higher frequency of agonistic interactions in multi-male troops than in unimale troops.
A geographically isolated natural population of about 2,000 langurs near Jodhpur (Rajasthan) has been studied by various Indian and foreign researchers since 1967. In the open scrub habitat, langurs spend an average of 66% of their feeding time on natural food and the rest eating food provided by local people for religious reasons. The langurs here are well habituated to people and are visible on the ground for most of the day, and are therefore easy to observe.
Jodhpur is located in Rajasthan at the eastern edge of the Great Indian Desert. In and around this town, which is surrounded by semi-desert plateau, lives a geographically isolated population of about 2,007 langurs: 38 one-male bisexual troops and 15 unisexual, all-male bands (Rajpurohit et al., 2010). The climate is dry, with maximum temperatures about 48° C in May/June and minimum around 0° C in December/January. Jodhpur receives 90% of its scanty rainfall (annual average: 360 mm) during the monsoon (July to September).
The habitat used by these langurs includes open scrub forests, uncultivated fields, human habitations, parks, and orchards. Water is available to all groups throughout the year from man-made ponds, which collect rainwater. Physical growth, genital development, and incisor-canines were used as major criteria for age categorization (cf. Rajpurohit & Sommer, 1991, and Table 1). Physical marks, scars, deformities, or typical moving and sitting postures were used to identify focal animals individually.
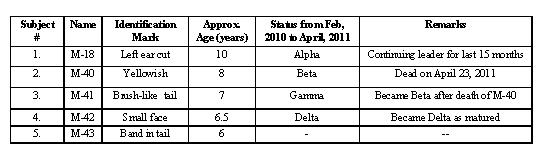
Table 1: Characteristics of adult and sub-adult males of the multi-male bisexual troop B-18.
Process of multi-male group formation: Most multi-male situations are created during inter-troop interaction and takeover. At the time of takeover, 3 or 4 males from an all-male band take part in aggressive interactions. After takeover, the intruders mostly stay in the troop for 2 or 3 days. If this situation continues for few more days, this troop temporarily changes to a multi-male bisexual troop. Sometimes the former alpha male stays in the troop as an interim resident for 5 to 10 days.
Interaction between the alpha and other males: This multi-male bisexual troop, B-18, lived in the Kailana Canal area about 9 km west of Jodhpur city. There are five males, (3 adult and 2 sub-adult) in this troop, including the alpha. They were all observed continuously staying in troop B-18. Sometimes the alpha male was observed to be aggressive toward the other adult males.
On Feb 10, 2010, at 10:30 a.m., the alpha male (M-18) was seen to be aggressive and approached the beta male (M-40) at feeding time when he was sitting with females. The alpha male took action: he whooped and ran toward the beta male and there was fighting for 3 or 4 minutes. The beta male received a bite wound about 6 cm long on his left leg (Fig. 1). He left the troop, but stayed within the troop’s home range. The alpha did not harm any other male, but they were keeping some distance from him. The alpha was noted to be aggressive often after this attack and was observed running behind the beta and gamma (M-41) males.

Figure 1: Cut on beta male’s leg.
On November 24, 2010, at 9:00 a.m., the alpha was observed being aggressive toward the beta male, who had a lower lip cut. The injury looked fresh – it was bleeding. But the beta and other males were staying in the troop, and nothing unusual was observed for the next six months.
On April 23, 2011, at 7:30 a.m., all the members of the troop were sitting along the canal, looking into it. There was a dead langur in the canal – it was the beta male (Fig. 2). The alpha appeared more aggressive, whooping and jumping from one branch to another. The other males were also sitting along the canal. They were continually teeth grinding toward the alpha. It was presumed that the beta male had been killed by the alpha on the previous day (April 22). All the members of the troop appeared afraid. But actual fighting between the alpha and the other males was not observed. After three days the troop was normal and the other males were still in the troop, with the gamma male in the beta position.

Figure 2: Dead beta male.
Change in the resting site: Before the death of the beta male all the males used a sleeping site by the canal, keeping some distance from the females, but after the death of the beta, the new beta and the young and subadult males all started sleeping on a tree near the canal, keeping a distance from the alpha.
Group leading in multi-male troop: At feeding time it was observed that when the alpha male feeds, all the males keep their distance; when the alpha completes his feeding, other males of the troop start. Among these latter males, the beta shows dominance. Sometimes the beta male leads his small bachelor troop. But during this study period the alpha always showed his leadership.
Changes of alpha male can be a rapid takeover (occurring in days) or a gradual process (up to several months). During gradual replacement, temporary multi-male stages and successions of short tenure may develop, which are typically labeled “interim residencies” (Sommer & Rajpurohit 1989). The social situation stabilizes after a male who is able to defend the troop for a longer period of time gains residency. Among various Hanuman langur field sites, with different percentages of one-male versus multi-male troop structure, Jodhpur, with an almost 100% harem structure, represents one extreme. Based on the ideas of Moore (1985), Newton (1988) suggested that a unimale troop structure will arise if a male is able to monopolize a group of females, and a multi-male troop if he is not.
Apart from the migration of immature males, a great variety of social changes have been recorded in the Hanuman langur. Change from unimale to multi-male troops and vice versa; troop formation from fragments of other troops and bands; gradual resident-male replacement and rapid resident-male replacement (or takeover), with or without infanticide, have been noted (Sugiyama, 1967; Roonwal & Mohnot, 1977; Hrdy 1977; Newton, 1987; Sharma et al., 2010). In Hanuman langurs at Dharwar and Jodhpur, temporary multi-male stages occur during male invasion (Sugiyama, 1964; Mohnot, 1984; Vogel & Loch, 1984;, Sharma, 2007).
Among the colobines, Marsh (1979) demonstrated a positive relationship between group size and the number of adult males in Senegalese red colobus (Procolobus), and Dunbar (1987) found a similar relationship in East African guereza (Colobus caudatus). Newton (1988a) also found a significant positive correlation between the number of adult males in troops of Hanuman langurs and both group size and the number of females in troops. But in Jodhpur at Kailana Canal (B-18) and Bijoloi (B-20), we observed two troops which have a negative correlation between the number of adult males and both group size and the number of females.
Three factors may affect a male’s ability to monopolize a troop. First, wide female dispersion within a troop will reduce a male’s ability to monopolize the females (Van Schaik & Van Hooff, 1983). Second, Terbergh & Janson (1986) suggest that males will be better able to monopolize females if they have ample time free from foraging. However, there is no clear association between the social organization and the proportion of fruit in a species’ diet; frugivores tend to spend less time resting and would therefore be predicted to be predominately unimale (Dunbar, 1988). Third, Emlen & Oring (1977) suggest that extreme synchrony or asynchrony in female reproductive cycles would reduce the potential for male monopolization, while moderate asynchrony would increase it.
Busse (1977) has argued that males in a multi-male troop are best able to defend themselves, their females, and their offspring. This explanation is consistent with the observation that red colobus live in multi-male troops in habitats where chimpanzees are sympatric (Gombe and Kibale: Stanford, 1998) and in unimale troops where they are not (Sare Demba Tana in Senegal: Struhsakar & Leland, 1987).
Hrdy (1977), Bishop (1979), and Laws & Laws (1984) have described a pattern of social change in some langur populations in which males in all-male bands associated with otherwise unimale troops. In the Rajaji population these influxes resulted in the formation of temporary multi-male troops during the mating season (Laws & Laws, 1984). In general, however, unimale troop populations of Hanuman langurs tend to show rapid, aggressive adult male replacement with evidence of infanticide (Sharma et al., 2010). In contrast, multi-male troop populations tend to show gradual male replacement with a staggered pattern of male introductions and exclusions (Boggess, 1984; Newton, 1986).
The sexual selection hypothesis appears to explain infanticide in both matrilineal harem society and patrilineal multi-male society. Why has infanticide not been found in colobine matrilineal multi-male troops? Leland et al. (1984) suggest that a unimale troop structure predisposes to infanticides because intermale reproduction competition and variance in mating success in such populations are greater than in multi-male societies. Infanticide will be facilitated as incoming males are unlikely to be closely related to troop infants and there are no other males present who might defend infants and contest post-takeover reproductive access. In contrast, in multi-male societies, competition for mating will occur within the troop, rather than between troop and band. Promiscuity will confuse paternity, increasing the probability that an infanticidal male would attack his own offspring and that another male would defend it. In addition, the chances that the infanticidal male would breed with the victim’s mother are reduced (Hrdy, 1979; Leland et al., 1984).
In the patrilineal, multi-male society of red colobus (Procolobus), Struhsakar (1975) described a pronounced dominance hierarchy among troop males that was expressed through priority to access to space, food, copulation, and grooming position.
Male calls of Hanuman langurs can be heard over 1 km from the source and occur predominantly in the first few hours after sundown (Newton, 1984), perhaps because thermal effects aid long-distance transmission (Waser & Waser, 1977). It is generally assumed that loud male calls are directed towards other males. Loud calls may also assist in maintaining troop cohesion and attracting females (Bennett, 1983).
The authors are grateful to S. M. Mohnot, Emeritus Professor of Zoology and chairman, Primate Research Center, Jodhpur, for continuous encouragement; to Dr. G. R. Jakher; and to Dr. Rajiv Gupta, Head, Department of Zoology, J.N.V. University, Jodhpur, for providing facilities and logistic support during this study. Thanks are due to UGC, New Delhi for financial support under major project (No. F.30-200/2004 (SR) dt.10 Nov 2004).
Altmann J. (1974). Observational study of behaviour: Sampling methods. Behaviour, 49, 227-267.
Bennett, E. L. (1983). The banded langur: Ecology of a colobine in a West Malaysian rain-forest. PhD thesis, University of Cambridge, England.
Bishop, N. H. (1979). Himalayan langurs: Temperate colobines. Journal of Human Evolution, 8, 251-281.
Boggess, J. (1980). Intermale relations and troop male membership changes in langurs (Presbytis entellus) in Nepal. International Journal of Primatology, 1, 233-274.
Boggess, J. (1984). Infant killing and male reproductive strategies in langurs (Presbytis entellus). In G. Hausfater & S. B. Hrdy (Eds.), Infanticide: Comparative and evolutionary perspectives (pp. 283-310). New York: Aldine.
Busse, C. D. (1977). Chimpanzee predation as a possible factor in the evoluation of red colobus monkey social organization. Evolution, 31, 907-911.
Dunbar, R. I. M. (1988). Primate social systems. Beckenham: Croom Helm.
Emlen, S., & Oring, L. (1977). Ecology, sexual selection and evolution of mating systems. Science, 197, 215-233.
Hrdy, S. B. (Ed.) (1977). The langurs of Abu: Female and male strategies of reproduction. Cambridge: Harvard University Press.
Hrdy, S. B. (1979). Infanticide among animals: A review, classification and examination of the implications for the reproductive strategies of females. Ethology and Sociobiology, 1, 13-40.
Jay, P. C. (1963). The social behaviour of the langur monkey. PhD thesis, University of Chicago, Illinois.
Laws, J. W., & Laws, J. V. H. (1984). Social interaction among adult male langurs (Presbytis entellus) at Rajaji Wildlife Sanctuary. International Journal of Primatology, 5, 31-50.
Leland, L., Struhsaker, T. T., & Butynski, T. M. (1984). Infanticide by adult males in three primate species of the Kibale Forest, Uganda: A test of hypotheses. In G. Hausfater & S. B. Hrdy (Eds.), Infanticide: Comparative and evolutionary perspectives (pp. 151-72). New York: Aldine.
Marsh, C. W. (1979). Female transference and mate choice among Tana River red colobus. Nature, 69, 568-569.
Mohnot, S. M. (1984). Some observations on all-male bands of the Hanuman langur (Presbytis entellus). In M. L. Roonwal, S. M. Mohnot, & N. S. Rathore (Eds.), Current primate researches (pp. 343-356). University of Jodhpur.
Mohnot, S. M., Agoramoorthy, G., Rajpurohit, L. S., & Srivastava, A. (1987). Ecobehavioural studies of Hanuman langur, Presbytis entellus. Technical Report (1983-86) (pp. 1-89). MAB Project, Department of Environment, Govt. of India, New Delhi.
Moore, J. (1985). Demography and sociality in primates. PhD thesis, Harvard University.
Newton, P. N. (1984). The ecology and social organization of Hanuman langurs (Presbytis entellus, Dufresne, 1797) in Kanha Tiger Reserve, Central Indian Highlands. PhD thesis, University of Oxford.
Newton, P. N. (1986). Infanticide in an undisturbed forest population of Hanuman langurs (Presbytis entellus). Animal Behaviour, 34, 785-789.
Newton, P. N. (1987). The social organization of forest Hanuman langurs (Presbytis entellus). International Journal of Primatology, 8, 199-232.
Newton, P. N. (1988). The variable social organization of Hanuman langur (Presbytis entellus), infanticide, and the monopolization of females. International Journal of Primatology, 9, 59-77.
Oppenheimer, J. R. (1977). Presbytis entellus, the Hanuman langurs. In H. S. H. Prince Rainier & G. H. Bourne (Eds.), Primate Conservation (pp. 459-512). New York: Academic Press.
Rajpurohit, L. S. (1987). Male social organisation in Hanuman langur, Presbytis entellus. PhD thesis, University of Jodhpur, Jodhpur.
Rajpurohit, L. S., Sharma, G., Devilal, Vijay, P., Swami, B., & Ram, C. (2010). Demography of free-ranging Hanuman langur (Semnopithecus entellus) in and around Jodhpur, Rajasthan (India). In Bioresources for Rural Livelihood, Vol. 3. Biodiversity and Ecology (pp. 311-315). New Delhi: Narendra Publishing House.
Rajpurohit, L. S., & Sommer, V. (1991). Sex differences in mortality among langurs (Presbytis entellus) of Jodhpur, Rajasthan. Folia Primatologica, 56, 17-27.
Rajpurohit, L. S., & Sommer, V. (1993). Juvenile male emigration from natal one-male troops in Hanuman langurs. In M. E. Pereira & L. A. Fairbanks (Eds.), Juvenile primates: Life history, development and behaviour (pp. 86-103). New York: Oxford University Press.
Rajpurohit, L. S., Srivastava, A., & Mohnot, S. M. (1994). Birth dynamics in Hanuman langur, Presbytis entellus of Jodhpur, India. Journal of Bioscience, 19, 315-324.
Roonwal, M. L., & Mohnot, S. M. (1977). Primates of south Asia: Ecology, sociobiology, and behavior. Cambridge, MA: Harvard University Press.
Sharma, G. (2007). Paternal care in Hanuman langur, Semnopithecus entellus entellus, around Jodhpur (India). PhD thesis, Jai Narain Vyas University, Jodhpur.
Sharma, G., Ram, C., & Rajpurohit, L. S. (2010). A case study of infanticide after resident male replacement in Semnopithecus entellus around Jodhpur (India). Laboratory Primate Newsletter, 49[4], 6-11.
Srivastava, A., Mohnot, S. M., & Rajpurohit, L. S. (1986). Existence of multi-male bisexual troops of the Hanuman langur (Semnopithecus entellus) in a predominantly one-male troop habitat. Abstract of paper presented at the International Symposium on Primates – The New Revolution, New Delhi, India, 26-31 December, 1986.
Sommer, V. (1985). Weibliche and mannliche reproduction strategien der Hanuman langur (Presbytis entellus) von Jodhpur, Rajasthan, India. PhD dissertation, Georg-August-Universität, Göttingen, Germany.
Sommer, V., & Rajpurohit, L. S. (1989). Male reproductive success in harem troops of Hanuman langur (Presbytis entellus). International Journal of Primatology, 10, 293-317.
Srivastava, A., & Mohnot, S. M. (1992). Existence of multimale troops and their transformation into unimale troops in Hanuman langurs. Primate Report, 34, 71-75.
Stanford, C. B. (1998). Chimpanzee and red colobus: The ecology of predator and prey. Cambridge, MA: Harvard University Press.
Struhsaker, T. T. (1975). The red colobus monkey. Chicago, IL: University of Chicago Press.
Struhsaker, T. T., & Leland, L. (1987). Colobines: Infanticide by adult males. In B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, & T. T. Struhsaker (Eds.), Primate societies (pp. 83-97). Chicago, IL: University of Chicago Press.
Sugiyama, Y. (1964). Group composition, population density and some sociological observations of Hanuman langurs (Presbytis entellus). Primates, 5, 7-37.
Sugiyama, Y. (1967). Social organization of Hanuman langurs. In S. A. Altmann (Ed.), Social communication among primates (pp. 221-236). Chicago, IL: University of Chicago.
Terborgh, J., & Janson, C. H. (1986). The sociology of primate groups. Annual Review of Ecology and Systematics, 17, 111-135.
Van Schaik, C. P., & Van Hooff, J. A. R. A. M. (1983). On the ultimate causes of primate social systems. Behaviour, 85, 91-117.
Vogel, C. (1977). Ecology and sociology of Presbytis entellus. In M. R. D. Prasad & T. C. Anand (Eds.), Use of nonhuman primates in biomedical research (pp. 24-45). New Delhi: Indian National Science Academy.
Vogel, C., & Loch, H. (1984). Reproductive parameters, adult male replacements, and infanticide among free-ranging langurs (Presbytis entellus) at Jodhpur (Rajasthan), India. In G. Hausfater & S. B. Hrdy (Eds.), Infanticide: Comparative and evolutionary perspectives (pp. 237-255). New York: Aldine Press.
Waser, P., & Waser, M. S. (1977). Experimental studies of primate vocalization: Specialization for long-distance propagation. Zeitschrift fur Tierpsychologie, 43, 239-263.
Wolfheim, J. H. (Ed.). (1983). Primates of the world: Distribution, abundance and conservation. Seattle: University of Washington Press.
-------------------------------------------
Dr. Goutam Sharma, Animal Behaviour Unit, Dept of Zoology, Jai Narain Vyas Univ., Jodhpur-342005 (Rajasthan), India [e-mail: [email protected]].
-------------------------------------------
* * *
So long, it’s been good to know you –
* * *
Announcements from Publications – IUCN Newsletters and Journals
The International Union for Conservation of Nature (IUCN) Species Survival Commission (SSC) Primate Specialist Group (PSG) publishes four regional news-letters, listed below, some of which have developed into small journals in their own right. Representing Africa, Asia, Madagascar, and the Americas, the four regional newsletter/journals are intended to share information among field researchers, conservationists and captive-care professionals.
In addition, the original newsletter of the PSG has itself evolved into a unique, overarching journal with a worldwide perspective. This journal, Primate Conservation, is published by Conservation International and the Margot Marsh Biodiversity Foundation, and provides an opportunity for researchers to publish longer, in-depth articles of interest to the global community of primate conservationists.
All of these publications are produced and distributed free of charge to authors and subscribers, on the premise that the free exchange of information is vital to effective conservation. This allows many researchers from habitat countries to continue receiving these publications – and contributing to them – without financial concern. Just as importantly, these publications are also available to students and young professionals in these countries, providing an important source of information and encouragement as they begin their conservation careers.
See <www.primate-sg.org/journals.htm> for information on all of these publications. To request a back issue of one of them, please contact Jill Lucena at [[email protected]].
African Primates
African Primates publishes information relevant to the conservation of nonhuman primates and their ecosystems in Africa. Its aim is to facilitate the exchange of information and ideas among primatologists and conservationists working with primates in Africa.
It is hoped that this newsletter will enhance the conservation of African primates by 1) increasing interest in their survival; 2) alerting people to situations where primate species and populations are under threat; and 3) providing a forum for useful debate on some of the more pressing, controversial and sensitive issues that have an impact on the conservation of these primates.
After a hiatus of six years, Carolyn L. Ehardt (University of Texas at San Antonio), the new Senior Editor, relaunched African Primates on 4 September 2010 in a new and upgraded electronic format. The journal is now a formal, peer-reviewed, fully indexed, scientific, international Open Resource E-Journal, freely accessible world-wide at: <journals.sfu.ca/afrprims>.
For submission of news, announcements, and manu-scripts: contact Carolyn Ehardt, Dept of Anthropology, Univ. of Texas, One UTSA Circle, San Antonio, TX 78249 [e-mail: [email protected]].
Asian Primates
Asian Primates is a new journal committed to disseminating information relating to research and conservation of nonhuman primates in Asia, and will also serve to highlight and draw the attention to issues relating to threatened primate species and their habitats.
Asian Primates will be an important source of information not only among the IUCN/SSC Primate Specialist Group members in the region, but also to other professionals and those with a keen interest in primates and primate conservation. The journal further aims to provide a venue for developing the capacities of young Asian nationals by encouraging them to submit manuscripts in English.
As no single discipline can encapsulate the many aspects of primates, Asian Primates thus encourages submissions that reflect inter- and multi-disciplinary perspectives about primates, thereby allowing the sharing of these perspectives, and the initiation of innovative and creative dialogues that help us learn more about our closest living relatives — and that help us to conserve them, their habitats, other denizens that share those habitats with them, and the ecosystem services these habitats provide us.
Please send all contributions to Dr. Jatna Supriatna, Conservation International, Jl. Pejaten Barat Raya No. 16 A, Kemang, Jakarta Selatan, 12550, Indonesia [e-mail: [email protected]]. For detailed information on the proper formatting of submissions, please consult the guidelines provided at <www.primate-sg.org/PDF /APJ1.l/contributors.pdf>.
Lemur News
Lemur News publishes manuscripts that deal largely or exclusively with lemurs and their habitat. The aims of the newsletter are 1) to provide a forum for exchange of information about all aspects of lemur biology and conservation, and 2) to alert interested people to particular threats to lemurs as they arise.
Lemur News welcomes the results of original research, field surveys, advances in field and laboratory techniques, book reviews, and informal status reports from research, conservation and management programs with lemurs in Madagascar and around the world. Manuscripts should be sent to: Christoph Schwitzer, Bristol Conservation and Science Foundation, Bristol Zoo Gardens, Clifton, Bristol BS8 3HA, U.K. [e-mail: [email protected]].
The 2010 issue of Lemur News is available at <www.primate-sg.org/PDF/LN15.pdf>. The address for earlier issues is the same, with the LN number different; the address for the 2011 issue will be the same, with LN16.pdf as the last part.
Neotropical Primates
“As the journal and newsletter of the Neotropical section of the PSG, Neotropical Primates helps to disseminate information on the biology and conservation of the New World monkeys. We welcome manuscripts dealing with any aspect of primate conservation, including research articles, news items, thesis abstracts, notices of recent publications, and the like.
“Anyone interested in submitting a manuscript or other item should please consult the author guidelines, at <www.primate-sg.org/NPguidelines2.1.doc>, beforehand. PLEASE NOTE: Neotropical Primates publishes articles in English, Spanish and Portuguese. If you are submitting an article in a language which is not your birth language, please have it thoroughly reviewed by a native speaker of that language before submitting it to us.
“The most recent issue currently on the Web can be seen at <www.primate-sg.org/PDF/NP17.1.pdf>. Earlier issues may be obtained by changing the number after NP in the last part.”
Primate Conservation
First published as a mimeographed newsletter in 1981, Primate Conservation has become a full journal devoted to sharing information on the world’s most threatened primates. Primate Conservation plays a central role in the publication of conservation research on primate species — in particular status surveys and studies on distribution, which are a fundamental component of conservation endeavor. Primate Conservation is also an ideal forum for longer articles, and has the flexibility to publish a variety of supporting materials such as color illustrations.
Anyone interested in submitting a manuscript or other item should please contact Anthony Rylands [e-mail: [email protected]].
PLEASE NOTE: All prospective authors should first review the instructions to contributors, at <www.primate-sg.org/PCguidelines1a.doc>. Manuscripts not received in proper house style may be delayed in publication.
UPDATE: Primate Conservation now publishes abstracts along with articles. Abstracts should be in English and no more than 250 words, with up to eight additional keywords. A second abstract may be included in another language, together with translated keywords, if the author prefers.
* * *
A New Lease on Life for Dieting Orangutan Oshine
Oshine, a former pet Bornean orangutan, was morbidly obese. When the team at Monkey World rescued her in August, 2010, she weighed 100 kg – more than double the natural weight of a female orangutan.
Since arriving at Monkey World – Ape Rescue Centre, the 14-year-old ape has been put on a health and fitness program designed to bring her weight down by losing fat while at the same time increasing her strength and agility. After 11 months the results are impressive: Oshine has lost a whopping 20 kg., is exercising and scaling a 20-m climbing frame, and has even adopted an orphaned baby orangutan named Silvestre!
“With Monkey World’s help, Oshine has turned her life around. When she arrived at the Rescue Centre she was morbidly obese and ran the risk of developing heart disease, blood clots, high blood pressure, and diabetes. We have been quite strict with her diet and have removed all sweets and processed foods that she used to get in addition to her normal diet, and it has worked,” said Dr. Alison Cronin, Director of Monkey World. “Oshine still has a long way to go and needs to lose another 20 to 30 kg before we will be happy with her fitness and health. I hope that by the end of this year Oshine will have lost enough weight so that we can introduce her to one of our adult groups of orangutans, where she will be able to have her own baby; but until she loses more weight it would not be healthy for her.”
Monkey World, in Dorset, U.K., is home to Europe’s orangutan crčche, where orphaned babies are sent to grow up with others of their own kind. For more information on Monkey World – Ape Rescue Centre, please visit <www.monkeyworld.org>. – Press Release from Monkey World – Ape Rescue Centre, July 28, 2011
Primate Testing – Crucial or Cruel?
Dr. Sebastien Farnaud of the Dr. Hadwen Trust and Professor Roger Lemon of University College, London, debated the ethics and uses of primate research. The Guardian (U.K.) printed their debate on July 29. It is now available at <www.guardian.co.uk/commentisfree/2011/jul/29/primate-testing-monkeys>.
Orangutans with iPads?
Orangutans at zoos nationwide may soon be able to connect with one another and create their own network of friends by using iPads and online chat services such as Skype.
The use of iPads by animals is not new, but researchers and orangutan advocates closely involved with an orangutan enrichment program recently launched at Milwaukee Zoo, in which orangutans are introduced to iPads, hope to take gadget use among orangutans nationwide and to a new level.
Richard Zimmerman, executive director of Orangutan Outreach, is working to get zoos across the country to introduce iPads in their orangutan enclaves and thus allow the apes to connect with their brethren thousands of miles away in “primate playdates” via Skype or other chat services. Zoos in Phoenix, Atlanta, and Toronto are among those that have agreed to participate. – From ComparativePsychNews, Digest #1861, Sept 2
* * *
Research Position – Parkinsonism and Aging
A research position is available at Thomas Jefferson University, Philadelphia, Pennsylvania, for a qualified individual to participate in a study of attention, memory and executive functioning in normal aged nonhuman primates and in aged nonhuman primates with early-stage Parkinsonism. The project is examining potential biomarkers associated with normal and pathological cognitive aging and will be examining various pharmacological strategies designed to improve age-related and Parkinson’s-related cognitive deficits. The researcher will be primarily responsible for behavioral testing of animals and for performing behavioral pharmacological studies, and will participate in the design and implementation of various neurochemical, anatomical, and molecular/biomarker studies.
Behavioral neuroscience experience is required, and previous experience with nonhuman primates is preferable. An MS or PhD in neuroscience or a related field is required. Qualified, experienced technicians will also be considered. All replies should be sent to Jay Schneider [e-mail: [email protected]].
Assistant Professor – U.C. Davis
The Department of Psychology at the University of California, Davis, invites applications for a tenure-track position in Psychobiology. The appointment will be at the Assistant Professor level, with a nine-month appointment. The proposed beginning date is July 1, 2012. We are seeking an outstanding scholar who studies the underlying biological mechanisms of behavior (e.g., neurobiological, genetic, epigenetic approaches). We are especially interested in candidates who study these mechanisms at the circuit, cellular, and/or molecular levels and who also have sophisticated approaches to behavior. Research on alternative animal model species is encouraged. The psychobiology area has dedicated facilities for small animal research. In addition, the California National Primate Research Center <www.cnprc.ucdavis.edu> is located three miles from the main campus; other campus resources include the Center for Neuroscience <neuroscience.ucdavis.edu> and the Genome Center <www.genomecenter.ucdavis.edu>. Candidates must have a PhD and also have a demonstrated capability or exceptional promise for developing an independent, extramurally-funded research program at the cutting edge of psychobiological research. Interested applicants should submit curriculum vitae, statements of research and teaching interests, representative reprints and/or preprints, and at least three letters of recommendation. Review of applications will commence on November 1, 2011, and will continue until the position is filled. Applications must be received online at <psych. facultysearch.ucdavis.edu>.
For information, contact the Search Committee Chair, Dr. Karen Bales [e-mail: [email protected]]. This position is subject to final administrative approval. To learn more about the rapidly expanding programs of research in psychobiology at the University of California, Davis, see <psychology.ucdavis.edu/psychareas/?AreaID=3&link=25>. The University of California, Davis, and the Department of Psychology are interested in candidates who are committed to the highest standards of scholarship and professional activities, and to the development of a campus climate that supports equality and diversity. The University of California is an affirmative action/equal opportunity employer.
* * *
EU Continues Support of European Network on Primate Research
(EUPRIM-Net)
The Network Activities focus on the training and education of the technical and scientific personnel interested in the biology and husbandry of primates. In a series of workshops and courses, knowledge about the morphology, physiology, and behavior of primates, as well as ecological aspects, is imparted by renowned international experts.
Another Network Activity provides a platform to discuss and define best practices in experimental procedures and husbandry of primates in biomedical research and breeding colonies. Topics addressed in a series of workshops are: Breeding programs – the pros and cons of F2 (second generation cross-breeding); Veterinary practices: A) Development of training materials and modules, and B) General techniques and best practice using NHP; Noninvasive imaging techniques in NHP research; Methods and practices for professional banking of tissues and material; and Diagnostic tools and setting up of a diagnostics validation system. The results of each workshop will be codified as official white papers and will provide important guidelines for the EUPRIM-Net partners and other primate centers.
During the past four years of work in EUPRIM-Net, Positive Reinforcement Training (PRT) has been established as a successful tool in reducing stress in connection with husbandry and scientific procedures in all participating centers. A training DVD, showing the benefits of PRT in different handling situations, has been widely distributed. The continuation of this Network Activity now includes ethological aspects of the other factors contributing to psychological well-being of the animals. The aim of this activity is to spread knowledge and understanding about Animal Behavioral Management (ABM) and PRT of laboratory primates to as many primate facilities as possible.
Biobanking is a direct measure to reduce the number of animals used in research. Within the scope of Access Activities, European researchers are offered easy access to a range of biological materials (e.g. tissue, blood, serum, DNA, RNA, cDNA) of different NHP species. The EUPRIM-Net BioBank has been appreciated as a small but valuable resource.
A number of Research Activities support the overall aims of EUPRIM-Net by delivering solid results that can be directly translated in the Network Activities. The research projects are about development of new and improved tools to measure infection and disease in NHP; telemetry; development of in vitro technologies to replace, reduce, and refine nonhuman primate studies; and research activities in ABM and PRT.
If you are interested in EUPRIM-Net and would like to know more about the project or specific activities, please contact Prof. Stefan Treue (Project Coordinator), or Dr. Björg Pauling (Project Manager), EUPRIM-Net, German Primate Center (DPZ), Kellnerweg 4, D-37077 Göttingen, Germany [e-mail: [email protected]]; and see <www.euprim-net.eu>.
* * *
Pharmaceutical Toxicology Workshop
Charles River and the National Centre for the Replacement, Refinement and Reduction of Animals in Research (NC3Rs) are sponsoring a pharmaceutical toxicology workshop, “Designing Studies to Reduce Animal Use While Maximizing Human Translation”, October 24–25 in Lansdowne, Virginia. Topics to be addressed include:
Environmental Enrichment for Laboratory Animals
The Institute of Animal Technology (IAT) Continuing Professional Development (CPD) Board announces its first Practical Environmental Enrichment Workshop for Laboratory Animals, which will be held November 8–10, 2011, at a central U.K. location. This workshop will have practical sessions, as well as group work and theory. Contact Sabrina Brando [e-mail: animalconcepts@ me.com] if you are interested and she will send you the relevant paperwork.
Other workshops in the near future include an Animal Welfare & Behaviour Seminar at the Zurich Zoo, Switzerland, October 19–21, 2011; and an Advanced Animal Training Workshop in Chessington, Surrey, U.K., November 15–18, 2011. See <www.animalconcepts.eu/AnimalConcepts/Events/Events.html> for links to details of all these workshops.
EUPRIM-Net Course on “General Primate Biology”
A EUPRIM-Net Course on “General Primate Biology” (for scientists, veterinarians, and students) will be held November 14–17, 2011, at the German Primate Center in Göttingen, <www.dpz.eu>, with lectures and interactive seminar sessions on a variety of different topics related to primate biology.
Experts have been invited to convey the latest knowledge in the fields of
On the Website <www.euprim-net.eu/network/courses.htm>, you will find more detailed information on the course program and a form for registration, as well as information for travel arrangements (site map and options for accomodations). There are substantial course-fee discounts for students and academics working at government-financed institutions. Course fees are not applicable for EUPRIM-Net partner institutes. The registration deadline is October 28.
If you have any questions or requests concerning the EUPRIM-Net course series in general or this course in particular, please contact Thomas Ziegler [e-mail: [email protected]].
* * *
The Orangutan SSP Husbandry Workshop 2011 will be held October 16–21 at the Cameron Park Zoo, Waco, Texas. See <www.cameronparkzoo.com/learnplay/conservation/orangutan-workshop/>.
A symposium, “Primatology in Peru: History, Current Status, and Perspectives”, will take place in Lima, Peru, October 18–21, 2011, with workshops and pre- and post-symposia on October 17, 18, and 22. In addition to workshops, the symposium will include invited speakers and oral and poster sessions. This symposium, organized by Yunkawasi, the Natural History Museum of the Principal University of San Marcos (Universidad Mayor de San Marcos), and the German Primate Center, is open to all interested persons. The site of the symposium is the School of Biological Sciences of Ricardo Palma University in Lima. For further information and to contact the organizers, see the symposium Website: <yunkawasiperu.org/evento-primates.html>.
The 3nd Leonardo Automation Club Symposium on “Lean Enterprise applied to vivarium operations and husbandry – improved operational efficiency, doubling the outcomes with the same human and physical resources; An ‘on time’ supply of required services” will be held in Buguggiate, Italy, October 24 to 25, 2011. This Symposium will focus on the Lean organization and management techniques as a proven approach to improving the ergonomics and working conditions of the staff and to cutting operating expenses in laboratory animal facilities, without jeopardizing research objectives, regulatory requirements, or animal care programs. The increasing pressure to optimize the efficient deployment of resources inevitably means reducing operating budgets, without neglecting maintenance of services for research and laboratory animal care programs. At the same time we experience a growing awareness and concern over repetitive and hazardous activities and the need to improve working conditions. The Leonardo Automation Club is an initiative of the Fondazione Guido Bernardini, the main objectives of which are to promote safety and ergonomics, hygiene standards and husbandry techniques in laboratory animal facilities, and, through the activities of the Club, to bring together a group of professionals with the same interests. For details, click the calendar at <www.fondazioneguidobernardini.org>.
The 29th Annual Nonhuman Primate Models for AIDS meeting will be October 25–28, 2011, at the Bell Harbor International Conference Center, Seattle, Washington, sponsored by the Washington National Primate Research Center. See <nhp2011.wanprc.org/>.
In cooperation with AALAS and AAALAC, the 23rd Annual Conference Animal Research Facilities 2011 (Planning * Construction * Operations * Sustainability) will be held November 7–8, 2011, at the Renaissance Las Vegas Hotel (Nevada). See <www.TradelineInc.com/Animal2011>. There will also be three pre-conference courses. See <www.TradelineInc.com/Animal2011PreCon2>:
* * *
“The American Society of Naturalists (ASN) typically holds a joint conference each year (the ‘Evolution Meeting’) with the Society for the Study of Evolution (SSE) and Society of Systematic Biologists (SSB). Starting in 2014, we plan to begin biennial independent meetings of the American Society of Naturalists (ASN would continue to also meet with SSE/SSB). The independent meeting is intended to bring together a broader spectrum of biologists, including ecologists and animal behaviorists, who might not typically attend the Evolution meeting. The purpose of these meetings will be to foster interdisciplinary interactions among biologists who share ASN’s goals: “to advance and to diffuse knowledge of organic evolution and other broad biological principles so as to enhance the conceptual unification of the biological sciences.” The current plan is to initiate the conference series with a meeting at the Asilomar Conference Center on the Monterey Peninsula, California, in 2014.
“To help us plan the best possible meeting, I am writing to ask you to fill out a brief survey. This will help us evaluate the level of interest in this meeting, choose the optimal time, and gather ideas for themes and formats that might help interdisciplinary conversations and communication between students, postdocs, and faculty at all levels.
“The survey can be found at: <www.surveymonkey.com/s/YNCHZ9V>. Please also forward this message to any students or colleagues who might be interested.”
* * *
New Name, Same Organization
Effective February 1, 2011, the name of Southwest Foundation for Biomedical Research has changed to the Texas Biomedical Research Institute to better reflect the organization and its mission. There is no change in management, tax exempt status, tax ID, or ownership.
Animal Nutrition Information
The Canadian Association of Zoos and Aquariums Nutrition Advisory and Research Group (CAZA-NARG) is a nutrition advisory and research group serving Canadian zoological institutions, in English and in French. Their material is also useful for wildlife and laboratory workers. See <www2.ville.montreal.qc.ca/biodome/narg>.
Interesting Websites
* * *
All The World’s Primates
Primate Conservation, Inc. (PCI), has announced the opening of its All The World’s Primates Website at <alltheworldsprimates.org>. Based on submissions from hundreds of scientists, conservationists, and wildlife photographers, and edited by Noel Rowe and Marc Myers, All The World’s Primates is a groundbreaking, comprehensive resource for researchers, students, and anyone interested in humans’ closest relatives. Each species and subspecies is described separately through an extensive information database, interactive range maps, photographs, illustrations, video and audio, overview articles, and other useful tools. All The World’s Primates members can share information and insights with other members through blogs and forums.
PCI director Noel Rowe says, “Education is a crucial part of conservation, so we think it’s important to have an in-depth online primate resource that stays current, is easy to use, and — for the first time — shows the beauty and diversity of the entire primate order.” All The World’s Primates Website encompasses all primates, including those that are newly described. It is updated regularly by its contributors, most of whom have done original research, and is fully referenced with over 10,000 citations.
For those who wish to simply look at the apes, monkeys, and prosimians, there are hundreds of videos and more than 3,000 photos and illustrations to enjoy. The site’s interface allows quick organization of primates by name, country, risk of extinction, or several other criteria while accessing each primate’s description, taxonomy, and range from the database. Other detailed information includes conservation status, diet, life history, behavior, social systems, and physical measurements. Complete data sets are available for researchers, while students and laypersons will appreciate overviews and introductory articles on topics in primatology and physical anthropology.
All The World’s Primates Website is a membership benefit of joining Primate Conservation, Inc. Proceeds support site maintenance and PCI’s grants program. Individual memberships start at $29.95 per semester, and institutional memberships are also offered. In keeping with PCI’s policy of support for work in primate habitat countries, a number of complimentary memberships are available. Reporters and bloggers may request a media pass to the site by e-mailing <[email protected]>.
Primate Conservation, Inc., is a U.S. 501(c)(3) non-profit foundation dedicated to studying, preserving, and maintaining the habitats of the least-known and most endangered primates in the world. Since 1993, PCI has supported more than 450 projects in 28 countries.
The Enrichment Record Poster Repository
“The Enrichment Record, together with the Veterinary Bioscience Institute, is excited to introduce The Enrichment Record Poster Repository. This open access poster repository will provide a structured and safe environment for the deposition of enrichment posters. Often, enrichment information presented as posters at conferences is not published and is lost once a conference is over. If it is published, it is published at a much later date. This repository will provide laboratory animal science community members with early access to enrichment information presented at conferences while allowing authors to maximize the value of their posters by dramatically increasing availability and by helping to prevent lengthy delays before others can benefit from their new enrichment research.
“The Enrichment Record Poster Repository is subdivided into categories by species, making it easy for the viewer to locate information. The submission process is simple and has safeguards in place to ensure applicability and maintain quality standards. After a poster is submitted, there is a 24-hour delay, which allows time for review of submitted information to confirm that it is appropriate for the site.
“The Enrichment Record Poster Repository is an open access repository, meaning that anyone will be able to view these posters. We believe open access promotes transparency and supports outreach efforts that help educate both internal and external audiences. Sharing is the optimal way to distribute information to the laboratory animal science community and beyond. You may post posters anonymously as long as you provide your authorship information to The Enrichment Record
“We are looking forward to your submissions and feedback. Visit the site: <www.vetbiotech.com/posters2.php>.”
Darwin Goes Digital
Books from Charles Darwin’s personal library –annotated by him – are now available online. For the first time in history, Charles Darwin’s scribbled notes contained within the pages of books from his library are available – online and free – to the general public.
“The chief interest of the Darwin books lies in the pencil notes scribbled on their pages, or written on scraps of paper and pinned to the last page,” Darwin’s son Francis said in 1908.
In April, 2011, the Biodiversity Heritage Library (BHL) digitized and published online 330 of the most heavily annotated resources in Darwin’s library. Every annotation, including underlines and crossed-out passages, has been painstakingly transcribed and catalogued and is searchable by keyword, at <www.biodiversitylibrary.org/collection/darwinlibrary>.
This is the first of two releases. The second, which is yet to be scheduled, will include all remaining items in the library, 413 of which also include his annotations. – By Jessica P. Johnson, June 24, 2011
Orangutan Photos for Orangutan Protection
Glenn Alderson, a photographer, is donating 100% of the profits of his orangutan photos to the Center for Orangutan Protection. See his Website: <glennalderson.com/borneo-project.php>.
Chimpanzee Skeletal Digital Atlas (Pan troglodytes)
A free digital academic atlas comprising most of the osteology of our closest living relative in anatomically oriented high-resolution photographs has recently been published. Anatomical terminology is in three languages: English, Spanish, and Latin. The folder, at <www.atapuerca.tv/descargas/docencia/20.zip>, is very large.
AAALAC Adopts New Position Statements
The Association for Assessment and Accreditation of Laboratory Animal Care International (AAALAC)’s Council on Accreditation has adopted six Position Statements to address several points in the 8th edition of the Guide for the Care and Use of Laboratory Animals (National Research Council, 2011) that required interpretation for institutions participating in the accreditation program.
The new Position Statements address:
* * *
Help Needed: Handbook on Welfare and Enrichment for Captive Wildlife
Halit Khoshen [e-mail: [email protected]] writes to Zoo-Biology:
“I am in the process of writing a handbook on welfare and enrichment for captive wildlife in Central America, which will be freely distributed in Spanish and English to Central American zoos, rescue centers, and sanctuaries.
“The handbook is divided into three main parts:
“If the information were available in an easy-to-read, fully illustrated fashion, and in their native language, they would be able to care for and elevate the quality of life of the animals under their care more easily. And what is more important than the day-to-day life?
“In order to design the most comprehensive handbook, I need the help of all of you.
“The first handbook is for mammals. It includes the coati, kinkajou, raccoon, jaguar, cougar, jaguaroundi, ocelot, margay cat, tayra, crab-eating fox, grey fox, tapir, spider monkey, night monkey, capuchin monkey, howler monkey, tamarin monkey, 2- and 3-toed sloths, agouti, paca, capybara, squirrel, northern tamandua, white-tailed deer, red brocket deer, white-lipped peccary, collared peccary, and Rothschild’s porcupine.
“If you or your organization would be willing to help in this enormous task, please do not hesitate to contact me at [email protected] for more information. If you know any organization or person I should contact, please let me know this as well.
“All contributions will be fully acknowledged.
“Aid is needed in:
* * *
IPS Education Grants and Awards
Elizabeth Lonsdorf, International Primatological Society (IPS) Vice-President for Education, announces the 2011 Lawrence Jacobsen Education Development Grants and Charles Southwick Conservation Education Commitment Awards.
Seven Jacobsen grants were awarded:
“Previous awardees for any IPS grant/award – please send me your name, project title, award year, and award type, as we are trying to track our awardees and grantees in a database. Thank you!
“If any members are interested in serving on the Education Committee, as a student competition judge, or have specific issues they would like addressed, please contact me at <[email protected]>.”
IPS Conservation Grants
Janette Wallis, IPS Vice-President for Conservation, writes: “This year, we received 50 applications for an IPS Conservation Grants and, as usual, it was a very difficult task to narrow our list of winners from so many outstanding applications. After careful review and deliberation, we selected eleven projects for funding. The following list provides (alphabetically) the principle investigator(s), country where the work will be carried out, and the title of the project. Those with “(CCI)” were selected to receive an additional $500 from the Conservation through Community Involvement initiative. We congratulate these winners and thank them for their work for primate conservation!”
IPS Research Grants
Jo Setchell, IPS Vice-President for Research, announces the winners of the 2011 Research Grants competition: “The competition was very strong this year, with 80 applications (approximately twice the number than last year) from 22 countries." The 10 successful applicants were:
Debby Cox, IPS Vice-President for Captive Care writes: “This year we received 20 grant submissions for captive care and while the committee would like to fund all worthwhile submissions, unfortunately, we could only select seven for this year. The seven selected stood out from the rest as seen in the scoring by committee members, and were approved by council. The successful applicants were:
* * *
Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
Books
* Among African Apes: Stories and Photos from the Field. M. Robbins & C. Boesch (Eds.). Berkeley: University of California Press, 2011. 182 pp.
* Animal Tool Behavior: The Use and Manufacture of Tools by Animals, Revised and Updated Edition. R. W. Shumaker, K. R. Walkup, & B. Beck. Baltimore: Johns Hopkins University Press, 2011. 282 pp.
* Born in Africa: The Quest for the Origins of Human Life. M. Meredith. New York: Public Affairs, 2011. 230 pp.
* The Chimpanzees of Bossou and Nimba. T. Matsuzawa, T. Humle, & Y. Sugiyama (Eds.). Japan: Springer Verlag, 2011. 490 pp.
* Gorillas: Living on the Edge. A. Rouse. Trethomas, Wales: Electric Squirrel Publishing, 2011. 96 pp.
* Monkeys on the Edge: Ecology and Management of Long-Tailed Macaques and Their Interface with Humans. M. D. Gumert, A. Fuentes, & L. Jones-Engel (Eds.). New York: Cambridge University Press, 2011. 382 pp.
* Primate Communication and Human Language: Vocalisation, Gestures, Imitation and Deixis in Humans and Non-humans. A. Vilain, J.-L. Schwartz, C. Abry, & J. Vauclair (Eds.). Philadelphia: John Benjamins, 2011. 246 pp.
* The Road to Primatology: A Naturalist’s Memoir. E. Mathiesen. Available from the author: 45 Lauriston St, Providence, RI 02906. About 26 pp. [$1 for postage]
. . . Originally conceived as a supplement to the Laboratory Primate Newsletter, this autobiographical memoir tells about the Assistant Editor’s fascination with animals and nature in childhood, discouragement with science in high school, and the serendipitous meeting with Judith Schrier, the Editor, that led to the author’s helping with the LPN for 24 years. Not only has the author enjoyed learning about primates during this time, but in addition primatology has informed her care of her three pet parrots. Finally, she describes the added bonus of getting to know the LPN’s authors and other primatologists at meetings, and the pleasure of working with some of them as editor and translator. Eventually this memoir will be published via Amazon.com. Prepublication copies are available from the author for the price of postage.
* Role of EAA in the Regulation of Prolactin in Nonhuman Primates: Involvement of Endogenous EAA and Their Interaction with Opioidergic and Adrenergic Pathways. S. Sajjad. Saarbrücken, Germany: LAP Lambert Academic Publishing, 2011. 173 pp.
* American Journal of Physical Anthropology, 2011, 145 [2], <onlinelibrary.wiley.com/doi/10.1002/ajpa.v145.2/issuetoc>.
. . . Contents include: Assessing endocranial variations in great apes and humans using 3D data from virtual endocasts, by T. Bienvenu, F. Guy, W. Coudyzer, E. Gilissen, G. Roualdčs, P. Vignaud, & M. Brunet; Comparing Dirichlet normal surface energy of tooth crowns, a new technique of molar shape quantification for dietary inference, with previous methods in isolation and in combination, by J. M. Bunn, D. M. Boyer, Y. Lipman, E. M. St. Clair, J. Jernvall, & I. Daubechies; On dental wear, dental work, and oral health in the type specimen (LB1) of Homo floresiensis, by W. L. Jungers & Y. Kaifu; and Timely estrus in wild brown mouse lemur females at Ranomafana National Park, southeastern Madagascar, by M. B. Blanco.
* American Journal of Physical Anthropology, 2011, 145 [3], <onlinelibrary.wiley.com/doi/10.1002/ajpa.v145.3/issuetoc>.
. . . Contents include: Locomotor behavior of wild orangutans (Pongo pygmaeus wurmbii) in disturbed peat swamp forest, Sabangau, Central Kalimantan, Indonesia, by K. L. Manduell, H. C. Morrogh-Bernard, & S. K. S. Thorpe; Female “dispersal” in hamadryas baboons: Transfer among social units in a multilevel society, by L. Swedell, J. Saunders, A. Schreier, B. Davis, T. Tesfaye, & M. Pines; The influence of experimental manipulations on chewing speed during in vivo laboratory research in tufted capuchins (Cebus apella), by C. L. Thompson, E. M. Donley, C. D. Stimpson, W. I. Horne, & C. J. Vinyard; Pattern and pace of dental eruption in Tarsius, by E. H. Guthrie & S. R. Frost; A new brain endocast of Homo erectus from Hulu Cave, Nanjing, China, by X. Wu, R. L. Holloway, L. A. Schepartz, & S. Xing; The postcranial dimensions of the La Chapelle-aux-Saints 1 Neandertal, by E. Trinkaus; and Feeding behavior and nutrient intake in spiny forest-dwelling ring-tailed lemurs (Lemur catta) during early gestation and early to mid-lactation periods: Compensating in a harsh environment, by L. Gould, M. L. Power, N. Ellwanger, & H. Rambeloarivony.
* Animal Behaviour, 2011, 82[1], <www.sciencedirect.com/science/journal/00033472>.
. . . Contents include: The impact of social networks on animal collective motion, by N. W. F. Bode, A. J. Wood, & D. W. Franks; Female voice frequency in the context of dominance and attractiveness perception, by B. Borkowska & B. Pawlowski; The evolution of superstition through optimal use of incomplete information, by K. R. Abbott & T. N. Sherratt; The effect of innovation and sex-specific migration on neutral cultural differentiation, by S. Yeaman, R. Bshary, & L. Lehmann; Correlates of birth origin effects on the development of stereotypic behaviour in striped mice, Rhabdomys, by M. A. Jones, G. J. Mason, & N. Pillay; and Which is the best experimental design in animal choice tests? By O. A. Bruzzone & J. C. Corley.
* Animal Behaviour, 2011, 82[3], <www.sciencedirect.com/science/journal/00033472>.
. . . Contents include: Animal behaviour meets microbial ecology, by E. A. Archie & K. R. Theis; Chimpanzees, Pan troglodytes, share food in the same way after collaborative and individual food acquisition, by A. P. Melis, A.-C. Schneider, & M. Tomasello; Paternity and relatedness in a polyandrous nonhuman primate: Testing adaptive hypotheses of male reproductive cooperation, by S. L. Díaz-Muńoz; and Molecular approaches in behavioural research: A cautionary note regarding mitochondrial transfers to the nucleus (numts), by D. A. Triant & L. D. Hayes.
* Comparative Medicine, 2011, 61[3], <tinyurl.com/6f3p6qg>.
. . . Contents include: Minimizing animal numbers: The variable-criteria sequential stopping rule, by D. A. Fitts; Psychogenic alopecia in rhesus macaques presenting as focally extensive alopecia of the distal limb, by J. A. Kramer, K. G. Mansfield, J. H. Simmons, & J. A. Bernstein; Alterations in peripheral blood B-cell populations in SHIV89.6P-infected macaques (Macacca fascicularis), by H. M. Kling, T. W. Shipley, & K. A. Norris; and Colonization with nontuberculous mycobacteria is associated with positive tuberculin skin test reactions in the common marmoset (Callithrix jacchus), by L. M. Wachtman, A. D. Miller, D. Xia, E. H. Curran, & K. G. Mansfield.
* The Gibbon’s Voice, November, 2010, 12[1], <www.gibboncenter.org>. (Gibbon Conservation Center, P.O. Box 800249, Santa Clarita, CA 91380.)
. . . Contents include Reintroducing Javan gibbons (Hylobates moloch): An assessment of behavioral preparedness, by J. Smith; and a photo of a newborn H. moloch, born at the Gibbon Conservation Center.
* International Journal of Primatology, 2011, 32[4],
<www.springerlink.com/content/0164-0291>.
. . . Contents: Factors affecting urine washing behavior in tufted Capuchins (Cebus apella), by G. Schino, M. Palumbo, & E. Visalberghi; Density and spatial distribution of buffy-tufted-ear marmosets (Callithrix aurita) in a continuous Atlantic forest, by D. Norris, F. Rocha-Mendes, R. Marques, R. de Almeida Nobre, & M. Galetti; Taxonomic implications of a field study of morphotypes of Hanuman langurs (Semnopithecus entellus) in peninsular India, by K. S. Chetan Nag, P. Pramod, & K. Praveen Karanth; Predicting capacity demand on sanctuaries for African chimpanzees (Pan troglodytes), by L. J. Faust, D. Cress, K. H. Farmer, S. R. Ross, & B. B. Beck; Anatomical contributions to hylobatid taxonomy and adaptation, by A. L. Zihlman, A. R. Mootnick, & C. E. Underwood; Population density estimates of the critically endangered yellow-tailed woolly monkeys (Oreonax flavicauda) at La Esperanza, northeastern Peru, by S. Shanee & N. Shanee; Impact of habitat fragmentation on the demography of lion-tailed macaque (Macaca silenus) populations in the rainforests of Anamalai Hills, Western Ghats, India, by G. Umapathy, S. Hussain, & S. Shivaji; Gray-brown mouse lemurs (Microcebus griseorufus) as an example of distributional constraints through increasing desertification, by Y. E.-M. B. Bohr, P. Giertz, Y. R. Ratovonamana, & J. U. Ganzhorn; Copulation calls in female chimpanzees (Pan troglodytes schweinfurthii) convey identity but do not accurately reflect fertility, by S. W. Townsend, T. Deschner, & K. Zuberbühler; Extension of the geographical range of white-browed titi monkeys (Callicebus discolor) and evidence for sympatry with San Martin titi monkeys (Callicebus oenanthe), by J. Vermeer, J. C. Tello-Alvarado, S. Moreno-Moreno, & F. Guerra-Vásquez; Expression of myosin heavy chain isoforms in the supraspinatus muscle of different primate species: Implications for the study of the adaptation of primate shoulder muscles to different locomotor modes, by J. M. Potau, R. Artells, G. Bello, C. Muńoz, M. Monzó, J. F. Pastor, F. de Paz, M. Barbosa, R. Diogo, & B. Wood; Of least concern? Range extension by rhesus macaques (Macaca mulatta) threatens long-term survival of bonnet macaques (M. radiata) in peninsular India, by R. Kumar, S. Radhakrishna, & A. Sinha; Seasonal differences in activity patterns of Geoffroyi´s spider monkeys (Ateles geoffroyi) living in continuous and fragmented forests in southern Mexico, by Ó. M. Chaves, K. E. Stoner, and V. Arroyo-Rodríguez; The equivocal relationship between territoriality and scent marking in wild saddleback tamarins (Saguinus fuscicollis), by Y. Lledo-Ferrer, F. Peláez, & E. W. Heymann; Estrogen and progestogen correlates of the structure of female copulation calls in semi-free-ranging Barbary macaques (Macaca sylvanus), by D. Pfefferle, M. Heistermann, R. Pirow, J. K. Hodges, & J. Fischer; Distribution patterns of slender loris subspecies (Loris lydekkerianus) in Kerala, southern India, by S. Radhakrishna, H. N. Kumara, & R. Sasi; and Troop size, habitat use, and diet of chacma baboons (Papio hamadryas ursinus) in commercial pine plantations: Implications for management, by S. P. Henzi, L. R. Brown, L. Barrett, & A. J. Marais.
* IPPL News, May, 2011, 38[1], <www.ippl.org/news.php>. (Intl Primate Protection League, P.O. Box 766, Summerville, SC 29484 [e-mail: [email protected]]).
. . . Contents include: Bolivian grassroots refuges benefit rescued primates; “A Day in the Jungle” links Indonesian students with native wildlife; Bringing up Baby Betsy: A record-breaking colobus monkey rescue; and Sanctuary caregivers give chimp orphans lots of love (about the Centre de Réhabilitation des Primates de Lwiro in the Democratic Republic of Congo).
* Journal of the American Association for Laboratory Animal Science, 2011, 50[2], <www.ncbi.nlm.nih.gov/pmc/issues/195230>.
. . . Contents include: Determination of hemoglobin A1c and fasting blood glucose reference intervals in captive chimpanzees (Pan troglodytes), by M. S. McTighe, B. C. Hansen, J. J. Ely, & D. R. Lee; Pharmacokinetics of oxymorphone in titi monkeys (Callicebus spp.) and rhesus macaques (Macaca mulatta), by K. R. Kelly, B. H. Pypendop, J. K. Grayson, S. D. Stanley, K. L. Christe, L. M. Summers, & N. W. Lerche; Characterization of cardiac time intervals in healthy bonnet macaques (Macaca radiata) by using an electronic stethoscope, by H. Kamran, L. Salciccioli, S. Pushilin, P. Kumar, J. Carter, J. Kuo, C. Novotney, & J. M. Lazar; Use of p63, a myoepithelial cell marker, in determining the invasiveness of spontaneous mammary neoplasia in a rhesus macaque (Macaca mulatta), by M. J. Williams-Fritze, J. A. Carlson Scholz, V. Bossuyt, & C. J. Booth; Primary hepatic Mycobacterium tuberculosis complex infection with terminal dissemination in a pig-tailed macaque (Macaca nemestrina), by D. E. Stockinger, K. M. Roellich, K. W. Vogel, K. L. Eiffert, A. E. Torrence, J. L. Prentice, K. G. Stephens, C. K. Wallis, C. E. Hotchkiss, & R. D. Murnane; and Diagnosis and treatment of degenerative joint disease in a captive male chimpanzee (Pan troglodytes), by E. N. Videan, M. L. Lammey, & D. R. Lee.
* Lemur News: The Newsletter of the Madagascar Section of the IUCN/SSC Primate Specialist Group, December, 2010, No. 15, <www.primate-sg.org/PDF/LN15.pdf>.
. . . Contents: Madagascar’s Environmental Crisis. Madagascar’s illegal logging crisis: An update and discussion of possible solutions, by E. R. Patel; Ongoing threats to lemurs and their habitat inside the Sahamalaza – Iles Radama National Park, by M. Seiler, G. H. Randriatahina, & C. Schwitzer; News and Announcements; Preliminary conservation status assessment for the Data Deficient northern giant mouse lemur Mirza zaza, by E. J. Rode, K. A.-I. Nekaris, & C. Schwitzer; An observation of the hairy-eared dwarf lemur, Allocebus trichotis, in the Lakato region, eastern Madagascar, by E. Lagadec & S. M. Goodman; When big lemurs swallow up small ones: Coquerel’s dwarf lemur as a predator of grey mouse lemurs and endemic rodents, by S. Schliehe-Diecks, M. Markolf, & E. Huchard; Collective mobbing of a boa by a group of red-fronted lemurs (Eulemur fulvus rufus), by L. Pyritz & T. Andrianjanahary; Response of two nocturnal lemurs (Microcebus murinus and Lepilemur leucopus) to a potential boiidae (Sanzinia madagascariensis) predator, by K. Fish; Effective predation defence in Cheirogaleus medius, by K. H. Dausmann; Lepilemur feeding observations from Northern Madagascar, by A. J. Lowin; Hypotheses on ecological interactions between the aye-aye (Daubentonia madagascariensis) and microhylid frogs of the genus Platypelis in Tsaratanana bamboo forest, by A. Rakotoarison, S. A. Rasamison, E. Rajeriarison, D. R. Vieites, & M. Vence; Discovery of crowned sifaka (Propithecus coronatus) in Dabolava, Miandrivazo, Menabe Region, by J. Razafindramanana & R. Rasamimanana; Inferences about the distant past in Madagascar, by E. L. Simons; Husbandry guidelines for mouse lemurs at Paris Zoo, by D. Roullet; Diurnal lemur density in the national park parcel Ivontaka Nord, UNESCO Biosphere Reserve of Mananara-Nord, by M. Polasky Lyons; Distribution of Prolemur simus north of the Mangoro-Nosivolo River – How far north do we really have to look? by R. Dolch, E. R. Patel, J. H. Ratsimbazafy, C. D. Golden, T. Ratolojanahary, J. Rafalimandimby, & J. L. Fiely; Enquęte préliminaire de la distribution des lémuriens de bambou dans et autour du Corridor forestier Fandriana-Vondrozo, Madagascar, by A. Rajaonson, M. Ratolojanahary, J. Ratsimbazafy, A. Feistner, & T. King; Effect of red ruffed lemur gut passage on the germination of native rainforest plant species, by O. H. Razafindratsima & E. Razafimahatratra; Feeding ecology of the crowned sifaka (Propithecus coronatus) in a coastal dry forest in northwest Madagascar (SFUM, Antrema), by C. Pichon, R. Ramanamisata, L. Tarnaud, F. Bayart, A. Hladik, C. M. Hladik, & B. Simmen; Effet de la dégradation de l’habitat sur la consommation alimentaire d’Eulemur rubriventer dans deux sites: Talatakely et Vatoharanana, du Parc National de Ranomafana, by L. H. Rakotonirina, G. J. Spiral, J. H. Ratsimbazafy, S. Ravelonjanahary, R. Ralaiarison, S. Tecot, A. Hall, T. Calhoon, & G. R. Randria; Observations of terrestrial latrine behaviour by the southern gentle lemur Hapalemur meridionalis in the Mandena littoral forest, southeast Madagascar, by T. M. Eppley & G. Donati; Conservation des lémuriens via la protection de leurs habitats et le développement communautaire dans les corridors de Betaolana et Tsaratanana-Betaolana, région de SAVA, by L. Razafy Fara & I. Andriamarosolo; Genetic diversity in ten Indri (Indri indri) populations compared to other lemur species, by J. Zaonarivelo, R. Brenneman, R. Andriantompohavana, & E. E. Louis, Jr.; Verreaux’s sifaka fur condition in the spiny forest of southern Androy, by I. Norscia, J. Lambotsimihampy, & E. Palagi; and Rediscovery of Sibree’s dwarf lemur in the fragmented forests of Tsinjoarivo, central-eastern Madagascar, by M. B. Blanco.
* Primate Conservation, Inc. UPDATE, 2011, <www.primate.org/news.htm>. (1411 Shannock Rd, Charlestown, RI 02813).
. . . Contents include: Mapping habitat loss and availability for Callicebus modestus and Callicebus olallae in the Department of Beni, Bolivia, by L. Lopez & A. Reinaga; and Hoolock gibbon survey in Assam, India, by R. Chetry & D. Chetry.
* Proceedings of the National Academy of Sciences, U.S.A., 2011, 108[25], <www.pnas.org/content/108/25.toc>.
. . . Contents include: Morphology, body proportions, and postcranial hypertrophy of a female Neandertal from the Sima de las Palomas, southeastern Spain, by M. J. Walker, J. Ortega, K. Parmová, M. V. López, & E. Trinkaus; Bidirectional plasticity in the primate inferior olive induced by chronic ethanol intoxication and sustained abstinence, by J. P. Welsh, V. Z. Han, D. J. Rossi, C. Mohr, M. Odagiri, J. B. Daunais, & K. A. Grant; and Maturation time of new granule cells in the dentate gyrus of adult macaque monkeys exceeds six months, by S. J. Kohler, N. I. Williams, G. B. Stanton, J. L. Cameron, & W. T. Greenough.
* Proceedings of the National Academy of Sciences, U.S.A., 2011, 108[34], <www.pnas.org/content/108/34.toc>.
. . . Contents include: Craniometric ratios of microcephaly and LB1, Homo floresiensis, using MRI and endocasts, by R. C. Vannucci, T. F. Barron, & R. L. Holloway; and Maternal separation produces lasting changes in cortisol and behavior in rhesus monkeys, by X. Feng, L. Wang, S. Yang, D. Qin, J. Wang, C. Li, L. Lv, Y. Ma, & X. Hu.
* 28th Annual Symposium on Nonhuman Primate Models for AIDS. Ronald S. Veazey & Preston A. Marx, Guest Editors. Journal of Medical Primatology, 2011, 40[4],
<onlinelibrary.wiley.com/doi/10.1111/jmp.2011.40.issue-4/issuetoc>.
. . . Contents: Introduction, by R. S. Veazey & P. A. Marx; A divergent myeloid dendritic cell response at virus set-point predicts disease outcome in SIV-infected rhesus macaques, by S. M. Barratt-Boyes & V. Wijewardana; Development of a pigtail macaque model of sexually transmitted infection/HIV coinfection using Chlamydia trachomatis, Trichomonas vaginalis, and SHIVSF162P3, by T. Henning, Y. Fakile, C. Phillips, E. Sweeney, J. Mitchell, D. Patton, G. Sturdevant, H. D. Caldwell, W. E. Secor, J. Papp, R. M. Hendry, J. McNicholl, & E. Kersh; High-throughput single-nucleotide polymorphism discovery and the search for candidate genes for long-term SIVmac nonprogression in Chinese rhesus macaques (Macaca mulatta), by J. Satkoski Trask, W. T. Garnica, R. S. Malhi, S. Kanthaswamy, & D. G. Smith; Reactivation of latent tuberculosis in rhesus macaques by coinfection with simian immunodeficiency virus, by S. Mehra, N. A. Golden, N. K. Dutta, C. C. Midkiff, X. Alvarez, L. A. Doyle, M. Asher, K. Russell-Lodrigue, C. Monjure, C. J. Roy, J. L. Blanchard, P. J. Didier, R. S. Veazey, A. A. Lackner, & D. Kaushal; SIV-infected Chinese-origin rhesus macaques express specific MHC class I alleles in either elite controllers or normal progressors, by D. Wambua, R. Henderson, C. Solomon, M. Hunter, P. Marx, A. Sette, & B. R. Mothé; and Abstracts.
* Diversity in Cortical Organization. 22nd Annual Karger Workshop, San Diego, California, Nov 11, 2010. Brain, Behavior and Evolution, 2011, 78[1].
* Anatomical pathways for auditory memory in primates. Munoz-Lopez, M. M., Mohedano-Moriano, A., & In-sausti, R. (Monica M. Munoz-Lopez, Human Neuroanatomy Lab., Dept of Health Sciences, School of Med., Univ. of Castilla-La Mancha, Ave. Almansa, 14, 02006 Albacete, Spain. [e-mail: [email protected]]). Frontiers in Neuroanatomy, 2010, 4:129, doi: 10.3389/fnana.2010.00129 <www.frontiersin.org/neuroanatomy/10.3389/fnana.2010.00129/full>.
. . . Episodic memory or the ability to store context-rich information about everyday events depends on the hippocampal formation (entorhinal cortex, subiculum, presubiculum, parasubiculum, hippocampus proper, and dentate gyrus). A substantial amount of behavioral-lesion and anatomical studies have contributed to our understanding of the organization of how visual stimuli are retained in episodic memory. However, whether auditory memory is organized similarly is still unclear. One hypothesis is that, like the “visual ventral stream” for which the connections of the inferior temporal gyrus with the perirhinal cortex are necessary for visual recognition in monkeys, direct connections between the auditory association areas of the superior temporal gyrus and the hippocampal formation and with the parahippocampal region (temporal pole, perirhinal, and posterior parahippocampal cortices) might also underlie recognition memory for sounds. Alternatively, the anatomical organization of memory could be different in audition. This alternative “indirect stream” hypothesis posits that, unlike the visual association cortex, the majority of auditory information makes one or more synapses in intermediate, polymodal areas, where they may integrate information from other sensory modalities, before reaching the medial temporal memory system. This review considers anatomical studies that can support either one or both hypotheses – focusing on anatomical studies on the primate brain, primarily in macaque monkeys, that have reported not only direct auditory association connections with medial temporal areas, but, importantly, also possible indirect pathways for auditory information to reach the medial temporal lobe memory system.
* Projections to early visual areas V1 and V2 in the calcarine fissure from parietal association areas in the macaque. Borra, E., & Rockland, K. S. (Elena Borra, Dip. di Neuroscienze, Sez. di Fisiologia, Univ. di Parma, Via Volturno 39, I-43100 Parma, Italy [e-mail: [email protected]]). Frontiers in Neuroanatomy, 2011, 5, <www.frontiersin.org/neuroanatomy/10.3389/fnana.2011.00035/full>.
. . . “Non-extrastriate projections to area V1 in monkeys, now demonstrated by several anatomical studies, are potential substrates of physiologically documented multisensory effects in primary sensory areas. The full network of projections among association and primary areas, however, is likely to be complex and is still only partially understood. In the present report, we used the anterograde tracer biotinylated dextran amine to investigate projections to areas V1 and V2 from subdivisions of the parietal association cortex in macaque. Parietal cortex was chosen to allow comparisons between projections from this higher association area and from other previously reported areas. In addition, we were interested in further elucidating pathways to areas V1 and V2 from parietal areas, as potentially contributing to attention and active vision. Of eight cases, three brains had projections only to area V2, and the five others projected to both areas V1 and V2. Terminations in area V1 were sparse. These were located in supragranular layers I, II, upper III; occasionally in IVB; and in layer VI. Terminations in V2 were denser, and slightly more prevalent in the supragranular layers. For both areas, terminations were in the calcarine region, corresponding to the representation of the peripheral visual field. By reconstructions of single axons, we demonstrated that four of nine axons had collaterals, either to V1 and V2 (n = 1) or to area V1 and a ventral area likely to be TEO (n = 3). In area V1, axons extended divergently in layer VI as well as layer I. Overall, these and previous results suggest a nested connectivity architecture, consisting of multiple direct and indirect recurrent projections from association areas to area V1. Terminations in area V1 are not abundant, but could be potentiated by the network of indirect connections.”
* Are wet-induced wrinkled fingers primate rain treads? Changizi, M., Weber, R., Kotecha, R., & Palazzo, J. (2AI Labs, 917 Lusk St, Ste 369, Boise, ID 83706 [e-mail: [email protected]]). Brain, Behavior and Evolution, 2011, 77, 286-290, <www.ncbi.nlm.nih.gov/pubmed/21701145>.
. . . “Wet fingers and toes eventually wrinkle, and this is commonly attributed by lay opinion to local osmotic reactions. However, nearly a century ago surgeons observed that no wrinkling occurs if a nerve to the finger has been cut. Here we provide evidence that, rather than being an accidental side effect of wetness, wet-induced wrinkles have been selected to enhance grip in wet conditions. We show that their morphology has the signature properties of drainage networks, enabling efficient removal of water from the gripped surface.”
* Selective attention from voluntary control of neurons in prefrontal cortex. Schafer, R. J., & Moore, T. (T. M., Stanford Univ. Sch. of Med., Stanford, CA 94305 [e-mail: [email protected]]). Science, 2011, 332, 1568-1571,
<www.sciencemag.org/content/332/6037/1568.full>.
. . . “Animals can learn to voluntarily control neuronal activity within various brain areas through operant conditioning, but the relevance of that control to cognitive functions is unknown. We show that monkeys can control the activity of neurons within the frontal eye field (FEF), an oculomotor area of prefrontal cortex. However, operantly driven FEF activity was primarily associated with selective visual attention and not oculomotor preparation. Attentional effects were untrained and were observed both behaviorally and neurophysiologically. Furthermore, selective attention correlated with voluntary, but not spontaneous, fluctuations in FEF activity. Our results reveal a specific association of voluntarily driven neuronal activity with ‘top-down’ attention and suggest a basis for the use of neurofeedback training to treat disorders of attention.”
* Spinogenesis and pruning in the anterior ventral infero-temporal cortex of the macaque monkey: An intracellular injection study of layer III pyramidal cells. Elston, G. N., Oga, T., Okamoto, T., & Fujita, I. (G. N. E., Centre for Cog. Neurosci., 60 Duke Rd, Doonan, Sunshine Coast, QLD 4562, Australia [e-mail: [email protected]]). Frontiers in Neuroanatomy, 2011, 5:42.
<www.frontiersin.org/neuroanatomy/10.3389/fnana.2011.00042/full>.
. . . “Pyramidal cells grow and mature at different rates among different cortical areas in the macaque monkey. In particular, differences across the areas have been reported in both the timing and magnitude of growth, branching, spinogenesis, and pruning in the basal dendritic trees of cells in layer III. Presently available data suggest that these different growth profiles reflect the type of functions performed by these cells in the adult brain. However, to date, studies have focused on only a relatively few cortical areas. In the present investigation we quantified the growth of the dendritic trees of layer III pyramidal cells in the anterior ventral portion of cytoarchitectonic area TE (TEav) to better comprehend developmental trends in the cerebral cortex. We quantified the growth and branching of the dendrities, and spinogenesis and pruning of spines, from post-natal day 2 (PND2) to four and a half years of age. We found that the dendritic trees increase in size from PND2 to 7 months of age and thereafter became smaller. The dendritic trees became increasingly more branched from PND2 into adulthood. There was a two-fold increase in the number of spines in the basal dendritic trees of pyramidal cells from PND2 to 3.5 months of age and then a 10% net decrease in spine number into adulthood. Thus, the growth profile of layer III pyramidal cells in the anterior ventral portion of the inferotemporal cortex differs to that in other cortical areas associated with visual processing.”
* Task-dependent and independent synchronous activity of monkey hippocampal neurons in real and virtual translocation. Hori, E., Tabuchi, E., Matsumura, N., Ono, T., & Nishijo, H. (H. N., System Emotional Science, Univ. of Toyama, Sugitani 2630, Toyama 930-0194, Japan [e-mail: [email protected]]). Frontiers in Behavioral Neuroscience, 2011, 5:36, <www.ncbi.nlm.nih.gov/pmc/articles/PMC3139221>.
. . . Previous neurophysiological and behavioral studies relate hippocampal functions to place learning and memory, and encoding of task- (or context-) specific information. Encoding of both task-specific information and own location is essential for episodic memory and for animals to navigate to reward-related places. It is suggested that different neural circuits with different assemblies of different hippocampal neurons are created in different environments or behavioral contexts for the hippocampal formation (HF) to encode and retrieve episodic memory. To investigate whether synchronous activity of hippocampal neurons, suggesting functional connectivity between those neurons, is task and position dependent, multiple single unit activities were recorded during performance of real and virtual translocation (VT) tasks. The monkey moved to one of four reward areas by driving a cab (real translocation) or by moving a pointer on a monitor. Of 163 neuron pairs, significant peaks in cross-correlograms (CCGs) were observed in 98 pairs. Most CCGs had positive peaks within 50 ms. Task-dependent cross-correlations (CCRs) were observed in 44% of the neuron pairs, and similarly observed in both the real and VT tasks. These CCRs were frequently observed in pyramidal vs. pyramidal neuron pairs with positive peak and peak shift. However, no consistent patterns of peak polarity, peak shift, and neuronal types were seen in task-independent CCRs. There was no significant difference in frequency of CCG peaks between real and VT tasks. These results suggest that the task-dependent information may be encoded by interaction among pyramidal neurons, and the common information across tasks may be encoded by interaction among pyramidal neurons and interneurons in the HF. These neuronal populations could provide a neural basis for episodic memory to disambiguously guide animals to places associated with reward in different situations.
* Encoding of complexity, shape and curvature by macaque infero-temporal neurons. Kayaert, G., Wagemans, J., & Vogels, R. (Lab. of Exp. Psych., Dept of Psych., K.U. Leuven, Tiensestraat 102, Box 3711, B3000 Leuven, Belgium [e-mail: [email protected]]). Frontiers in Systems Neuroscience, 5:51.
<www.ncbi.nlm.nih.gov/pmc/articles/PMC3131530>.
. . . “We recorded responses of macaque infero-temporal (IT) neurons to a stimulus set of Fourier boundary descriptor shapes wherein complexity, general shape, and curvature were systematically varied. We analyzed the response patterns of the neurons to the different stimuli using multidimensional scaling. The resulting neural shape space differed in important ways from the physical, image-based shape space. We found a particular sensitivity for the presence of curved versus straight contours that existed only for the simple but not for the medium and highly complex shapes. Also, IT neurons could linearly separate the simple and the complex shapes within a low-dimensional neural shape space, but no distinction was found between the medium and high levels of complexity. None of these effects could be derived from physical image metrics, either directly or by comparing the neural data with similarities yielded by two models of low-level visual processing (one using wavelet-based filters and one that models position and size invariant object selectivity through four hierarchically organized neural layers). This study highlights the relevance of complexity to IT neural encoding, both as a neurally independently represented shape property and through its influence on curvature detection.”
* Limited or no protection by weakly or nonneutralizing antibodies against vaginal SHIV challenge of macaques compared with a strongly neutralizing antibody. Burton, D. R., Hessell, A. J., Keele, B. F., Klasse, P. J., Ketas, T. A., Moldt, B., Dunlop, D. C., Poignard, P., Doyle, L. A., Cavacini, L., Veazey, R. S., & Moore, J. P. (Dept of Immunol. & Micro. Sci., Scripps Research Inst., La Jolla, CA 92037 [e-mail: [email protected][). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 11181-11186, <www.pnas.org/content/108/27/11181.full>.
. . . “To guide vaccine design, we assessed whether human monoclonal antibodies (MAbs) b12 and b6 against the CD4 binding site (CD4bs) on HIV-1 gp120 and F240 against an immundominant epitope on gp41 could prevent vaginal transmission of simian HIV (SHIV)-162P4 to macaques. The two anti-gp120 MAbs have similar monomeric gp120-binding properties, measured in vitro, but b12 is strongly neutralizing and b6 is not. F240 is nonneutralizing. Applied vaginally at a high dose, the strongly neutralizing MAb b12 provided sterilizing immunity in seven of seven animals, b6 in zero of five animals, and F240 in two of five animals. Compared with control animals, the protection by b12 achieved statistical significance, whereas that caused by F240 did not. For two of three unprotected F240-treated animals there was a trend toward lowered viremia. The potential protective effect of F240 may relate to the relatively strong ability of this antibody to capture infectious virions. Additional passive transfer experiments also indicated that the ability of the administered anti-gp120 MAbs to neutralize the challenge virus was a critical influence on protection. Furthermore, when data from all of the experiments were combined, there was a significant increase in the number of founder viruses establishing infection in animals receiving MAb b6, compared with other nonprotected macaques. Thus, a gp120-binding, weakly neutralizing MAb to the CD4bs was, at best, completely ineffective at protection. A nonneutralizing antibody to gp41 may have a limited capacity to protect, but the results suggest that the central focus of HIV-1 vaccine research should be on the induction of potently neutralizing antibodies.”
* Maternal high-fat diet disturbs uteroplacental hemodynamics and increases the frequency of stillbirth in a nonhuman primate model of excess nutrition. Frias, A. E., Morgan, T. K., Evans, A. E., Rasanen, J., Oh, K. Y., Thornburg, K. L., & Grove, K. L. (Dept of Obstetrics & Gynecology, Oregon Health & Science Univ., L458, 3181 S.W. Sam Jackson Park Rd, Portland, OR 97239 [e-mail: [email protected]]). Endocrinology, 2011, 152, 2456-2464, <endo.endojournals.org/content/152/6/2456.full>.
. . . “Prepregnancy maternal obesity confers an increased risk of stillbirth, but the mechanisms are unknown. Maternal obesity is associated with placental inflammation. We considered that maternal diet may predispose to the increased risk of placental inflammation and stillbirth. We hypothesized that a chronic high-fat diet (HFD) is associated with abnormal uteroplacental circulation and placental inflammation. Here we used a nonhuman primate model to determine the effect of chronic HFD on the uterine and placental hemodynamics, placental histology, and inflammation in a prospective, observational study of 24 Japanese macaques. Overall, there was a statistically significant (38–56%) reduction in uterine volume blood flow from HFD animals, whether they were lean or obese. Consumption of a HFD, independent of obesity, increased placental inflammatory cytokines and the expression of Toll-like receptor 4. We show that HFD consumption by obese mothers with hyperinsulinemia also reduced volume blood flow on the fetal side of the placenta and significantly increased the frequency of both placental infarctions and stillbirth. These results suggest that a HFD, independent of obesity, decreases uterine volume blood flow. Maternal obesity and insulin resistance further exacerbates the placental dysfunction and results in an increased frequency of stillbirth.”
* Functional correlates of the anterolateral processing hierarchy in human auditory cortex. Chevillet, M., Riesenhuber, M., & Rauschecker, J. P. (J. P. R., Georgetown Univ. Med. Ctr, New Research Bldg, Rm WP-19, 3970 Reservoir Rd NW, Washington, DC 20057 [e-mail: [email protected]]). Journal of Neuroscience, 2011, 31, 9345-9352, <www.jneurosci.org/content/31/25/9345.full>.
. . . “Converging evidence supports the hypothesis that an anterolateral processing pathway mediates sound identification in auditory cortex, analogous to the role of the ventral cortical pathway in visual object recognition. Studies in nonhuman primates have characterized the anterolateral auditory pathway as a processing hierarchy, composed of three anatomically and physiologically distinct initial stages: core, belt, and parabelt. In humans, potential homologs of these regions have been identified anatomically, but reliable and complete functional distinctions between them have yet to be established. Because the anatomical locations of these fields vary across subjects, investigations of potential homologs between monkeys and humans require these fields to be defined in single subjects. Using functional MRI, we presented three classes of sounds (tones, band-passed noise bursts, and conspecific vocalizations), equivalent to those used in previous monkey studies. In each individual subject, three regions showing functional similarities to macaque core, belt, and parabelt were readily identified. Furthermore, the relative sizes and locations of these regions were consistent with those reported in human anatomical studies. Our results demonstrate that the functional organization of the anterolateral processing pathway in humans is largely consistent with that of nonhuman primates. Because our scanning sessions last only 15 min/subject, they can be run in conjunction with other scans. This will enable future studies to characterize functional modules in human auditory cortex at a level of detail previously possible only in visual cortex. Furthermore, the approach of using identical schemes in both humans and monkeys will aid with establishing potential homologies between them.”
* Dosage thresholds for AAV2 and AAV8 photoreceptor gene therapy in monkey. Vandenberghe, L. H., Bell, P., Maguire, A. M., Cearley, C. N., Xiao, R., Calcedo, R., Wang, L., Castle, M. J., Maguire, A. C., Grant, R., Wolfe, J. H., Wilson, J. M., & Bennett, J. (J. B., F. M. Kirby Center for Molecular Ophthalmology, Scheie Eye Inst., Univ. of Pennsylvania, Philadelphia, PA 19104 [e-mail: [email protected]]). Science Translational Medicine, 2011, 3, 88ra54, <stm.sciencemag.org/content/3/88/88ra54.full>.
. . . Gene therapy is emerging as a therapeutic modality for treating disorders of the retina. Photoreceptor cells are the primary cell type affected in many inherited diseases of retinal degeneration. Successfully treating these diseases with gene therapy requires the identification of efficient and safe targeting vectors that can transduce photoreceptor cells. One serotype of adeno-associated virus, AAV2, has been used successfully in clinical trials to treat a form of congenital blindness that requires transduction of the supporting cells of the retina in the retinal pigment epithelium (RPE). Here, we determined the dose required to achieve targeting of AAV2 and AAV8 vectors to photoreceptors in nonhuman primates. Transgene expression in animals injected subretinally with various doses of AAV2 or AAV8 vectors carrying a green fluorescent protein transgene was correlated with surgical, clinical, and immunological observations. Both AAV2 and AAV8 demonstrated efficient transduction of RPE, but AAV8 was markedly better at targeting photoreceptor cells. These preclinical results provide guidance for optimal vector and dose selection in future human gene therapy trials to treat retinal diseases caused by loss of photoreceptors.
* Online adaptation and over-trial learning in macaque visuomotor control. Braun, D. A., Aertsen, A., Paz, R., Vaadia, E., Rotter, S. & Mehring, C. (Dept of Engineering, Univ. of Cambridge, Cambridge CB2 1PZ, U.K. [e-mail: [email protected]]). Frontiers in Computational Neuroscience, 2011, 5, <www.frontiersin.org/computational_neuroscience/10.3389/fncom.2011.00027/full>.
. . . “When faced with unpredictable environments, the human motor system has been shown to develop optimized adaptation strategies that allow for online adaptation during the control process. Such online adaptation is to be contrasted to slower over-trial learning that corresponds to a trial-by-trial update of the movement plan. Here we investigate the interplay of both processes, i.e., online adaptation and over-trial learning, in a visuomotor experiment performed by macaques. We show that simple non-adaptive control schemes fail to perform in this task, but that a previously suggested adaptive optimal feedback control model can explain the observed behavior. We also show that over-trial learning as seen in learning and aftereffect curves can be explained by learning in a radial basis function network. Our results suggest that both the process of over-trial learning and the process of online adaptation are crucial to understand visuomotor learning.”
* Intrasulcal electrocorticography in macaque monkeys with minimally invasive neurosurgical protocols. Matsuo, T., Kawasaki, K., Osada, T., Sawahata, H., Suzuki, T., Shibata, M., Miyakawa, N., Nakahara, K., Iijima, A., Sato, N., Kawai, K., Saito, N., & Hasegawa, I. (I. H., Dept of Physiology, Niigata Univ. Sch. of Med., Asahimachi St 1-757, Chuo-ku, Niigata 951-8510, Japan [e-mail: [email protected]]). Frontiers in Systems Neuroscience, 2011, 5, <www.ncbi.nlm.nih.gov/pmc/issues/193815/>.
. . . “Electrocorticography (ECoG), multichannel brain-surface recording and stimulation with probe electrode arrays, has become a potent methodology not only for clinical neurosurgery but also for basic neuroscience using animal models. The highly evolved primate’s brain has deep cerebral sulci, and both gyral and intrasulcal cortical regions have been implicated in important functional processes. However, direct experimental access is typically limited to gyral regions, since placing probes into sulci is difficult without damaging the surrounding tissues. Here we describe a novel methodology for intrasulcal ECoG in macaque monkeys. We designed and fabricated ultra-thin flexible probes for macaques with micro-electro-mechanical systems technology. We developed minimally invasive operative protocols to implant the probes by introducing cutting-edge devices for human neurosurgery. To evaluate the feasibility of intrasulcal ECoG, we conducted electrophysiological recording and stimulation experiments. First, we inserted parts of the Parylene-C-based probe into the superior temporal sulcus to compare visually evoked ECoG responses from the ventral bank of the sulcus with those from the surface of the inferior temporal cortex. Analyses of power spectral density and signal-to-noise ratio revealed that the quality of the ECoG signal was comparable inside and outside of the sulcus. Histological examination revealed no obvious physical damage in the implanted areas. Second, we placed a modified silicone ECoG probe into the central sulcus and also on the surface of the precentral gyrus for stimulation. Thresholds for muscle twitching were significantly lower during intrasulcal stimulation compared to gyral stimulation. These results demonstrate the feasibility of intrasulcal ECoG in macaques. The novel methodology proposed here opens up a new frontier in neuroscience research, enabling the direct measurement and manipulation of electrical activity in the whole brain.”
* Neighboring pallidal neurons do not exhibit more synchronous oscillations than remote ones in the MPTP primate model of Parkinson’s disease. Mitelman, R., Rosin, B., Zadka, H., Slovik, M., Heimer, G., Ritov, Y., Bergman, H., & Elias, S. (Dept of Med. Neurobiology, Inst. of Med. Res. Israel-Canada, Hebrew Univ. – Hadassah Med. Sch., P.O.B. 12272, Jerusalem 91120, Israel [e-mail: [email protected]]). Frontiers in Systems Neuroscience, 2011, 5, <www.ncbi.nlm.nih.gov/pmc/articles/PMC3129135/>.
. . . “In the healthy primate, neurons of the external and internal segments of the globus pallidus (GP) present a primarily irregular firing pattern, and a negligible level of synchrony is observed between pairs of neurons. This holds even for neighboring cells, despite their higher probability to receive common inputs and to innervate each other via lateral connectivity. In the Parkinsonian primate, this changes drastically, and many pairs of GP cells show synchronous oscillations. To address the relation between distance and synchrony in the Parkinsonian state, we compared the synchrony of discharge of close pairs of neurons, recorded by the same electrode, with remote pairs, recorded by different ones. However, spike trains of neighboring cells recorded by the same extracellular electrode exhibit the shadowing effect; i.e., lack of detection of spikes that occur within a few milliseconds of each other. Here, we demonstrate that the shadowing artifact can both induce apparent correlations between non-correlated neurons, as well as conceal existing correlations between neighboring ones. We therefore introduced artificial shadowing in the remote pairs, similar to the effect we observed in the close ones. After the artificial shadowing, neighboring cells did not show a higher tendency to oscillate synchronously than remote ones. On the contrary, the average percentage (over all sessions) of artificially shadowed remote pairs exhibiting synchronous oscillations was 35.4% compared to 17.2% in the close ones. A similar trend was found when the unshadowed remote pairs were separated according to the estimated distance between electrode tips: 29.9% of pairs at approximate distance of less than 750 μm were significantly synchronized, in comparison with 28.5% of the pairs whose distance was more than 750 μm. We conclude that the synchronous oscillations in the GP of MPTP-treated primates are homogenously distributed.”
* Dynamics of the primate ovarian surface epithelium during the ovulatory menstrual cycle. Wright, J. W., Jurevic, L., & Stouffer, R. L. (Div. of Repro. Sci., Oregon NPRC, Beaverton, OR 97006 [e-mail: wrightj@ ohsu.edu]). Human Reproduction, 2011, 26, 1408-1421, <www.ncbi.nlm.nih.gov/pubmed/21421660>.
. . . “Epithelial ovarian cancer (EOC) risk correlates strongly with the number of ovulations that a woman experiences. The primary source of EOC in women is the ovarian surface epithelium (OSE). Mechanistic studies on the etiology of OSE transformation to EOC cannot be realistically performed in women. Selecting a suitable animal model to investigate the normal OSE in the context of ovulation should be guided by the model’s reproductive similarities to women in natural features that are thought to contribute to EOC risk. We selected the nonhuman primate, rhesus macaque, as a surrogate to study the normal OSE during the natural menstrual cycle. We investigated OSE morphology and marker expression, plus cell proliferation and death in relation to menstrual cycle stage and ovulation. OSE cells displayed a morphological range from squamous to columnar. Cycle-independent parameters and cycle-dependent changes were observed for OSE histology, steroid receptor expression, cell death, DNA repair and cell adhesion. Contrary to findings in nonprimates, primate OSE cells were not manifestly cleared from the site of ovulation, nor were proliferation rates affected by ovulation or stage of the menstrual cycle. DNA repair proteins were more highly expressed in OSE than in other ovarian cells. This study identifies significant differences between primate and nonprimate OSE. In contrast to established views, ovulation-induced death and proliferation are not indicated as prominent contributors to EOC risk, but disruption of OSE cadherin-mediated adhesion may be, as could the loss of ovary-mediated chronic suppression of proliferation and elevation of DNA repair potential.”
* Ovarian surface epitheliectomy in the nonhuman primate: Continued cyclic ovarian function and limited epithelial replacement. Wright, J. W., Pejovic, T., Jurevic, L., Bishop, C. V., Hobbs, T., & Stouffer, R. L. (Address same as above). Human Reproduction, 2011, 26, 1422-1430, <www.ncbi.nlm.nih.gov/pubmed/21421661>.
. . . “The fifth leading cause of cancer deaths among women is ovarian cancer (OC), which originates primarily in the ovarian surface epithelium (OSE) that surrounds the ovary. Permanent removal of the OSE could provide a novel strategy to substantially reduce OC risk, while retaining the benefits of ovarian function, including gameto- and steroidogenesis. It must be determined whether ovarian surface epitheliectomy (OSEx) carries deleterious side effects, including loss of menstrual cyclicity, infertility, or scarring (e.g. adhesions), prior to any clinical application of this strategy. To achieve this, we selected the nonhuman primate, rhesus macaque, for long-term (12 month) studies on the effects of OSEx. Rhesus macaque females underwent OSEx by detergent treatment and were then monitored for menstrual cyclicity (menstruation, steroidogenesis and follicle development) and adverse side effects (tissue scarring or adhesions). Ovaries were collected at 6 or 12 months and examined for evidence of tissue damage, follicle rupture, and regression of the corpus luteum. The ovarian surface was examined immuno-histologically for signs of epithelial replacement, using markers for OSE and fimbrial epithelium (FE), a possible alternative source of pelvic tumors diagnosed as OC. After OSEx, menstrual cycle length, estrogen and progesterone production, follicle rupture, and luteal regression appeared normal. No evidence of adhesions was seen. At 6 and 12 months post-OSEx, the ovarian surface was sparsely populated by cells expressing OSE and FE markers. Proliferative activity in this population was notably low. OSEx may provide a novel method to reduce the risk of OC, without sacrificing ovarian function, although the effects on fertility remain to be tested. The absence of epithelial replacement via enhanced proliferation suggests OSEx does not increase malignant potential. Complete and permanent OSEx may be feasible.”
* A prime-boost strategy using virus-like particles pseudotyped for HCV proteins triggers broadly neutralizing antibodies in macaques. Garrone, P., Fluckiger, A.-C., Mangeot, P. E., Gauthier, E., Dupeyrot-Lacas, P., Mancip, J., Cangialosi, A., Du Chéné, I., LeGrand, R., Mangeot, I., Lavillette, D., Bellier, B., Cosset, F.-L., Tangy, F., Klatzmann, D., & Dalba, C. (C. D., Epixis S.A., F-69007 Lyon, France [e-mail: [email protected]]. Science Translational Medicine, 2011, 3, 94ra71, <stm.sciencemag.org/content/3/94/94ra71.full>.
. . . “Chronic hepatitis C virus (HCV) infection, with its cohort of life-threatening complications, affects more than 200 million persons worldwide and has a prevalence of more than 10% in certain countries. Preventive and therapeutic vaccines against HCV are thus much needed. Neutralizing antibodies (NAbs) are the foundation for successful disease prevention for most established vaccines. However, for viruses that cause chronic infection such as HIV or HCV, induction of broad NAbs from recombinant vaccines has remained elusive. We developed a vaccine platform specifically aimed at inducing NAbs based on pseudotyped virus-like particles (VLPs) made with retroviral Gag. We report that VLPs pseudotyped with E2 and/or E1 HCV envelope glycoproteins induced high-titer anti-E2 and/or anti-E1 antibodies, as well as NAbs, in both mouse and macaque. The NAbs, which were raised against HCV 1a, cross-neutralized the five other genotypes tested (1b, 2a, 2b, 4, and 5). Thus, the described VLP platform, which can be pseudotyped with a vast array of virus envelope glycoproteins, represents a new approach to viral vaccine development.”
* Differential prefrontal white matter development in chimpanzees and humans. Sakai, T., Mikami, A., Tomonaga, M., Matsui. M., Suzuki. K., Hamada, Y., Tanaka, M., Miyabe-Nishiwaki, T., Makishima, H., Nakatsukasa, M., Matsuzawa, T. (Primate Res. Inst., Kyoto Univ., Inuyama, Aichi 484-8506, Japan [e-mail: [email protected]]). Current Biology, 2011, 21, 1397-1402, <www.sciencedirect.com/science/journal/09609822>.
. . . A comparison of developmental patterns of white matter (WM) within the prefrontal region between humans and nonhuman primates is key to understanding human brain evolution. WM mediates complex cognitive processes and has reciprocal connections with posterior processing regions. Although the developmental pattern of prefrontal WM in macaques differs markedly from that in humans, this has not been explored in our closest evolutionary relative, the chimpanzee. The present longitudinal study of magnetic resonance imaging scans demonstrated that the prefrontal WM volume in chimpanzees was immature and had not reached the adult value during prepuberty, as observed in humans but not in macaques. However, the rate of prefrontal WM volume increase during infancy was slower in chimpanzees than in humans. These results suggest that a less mature and more protracted elaboration of neuronal connections in the prefrontal portion of the developing brain existed in the last common ancestor of chimpanzees and humans, and that this served to enhance the impact of postnatal experiences on neuronal connectivity. Furthermore, the rapid development of the human prefrontal WM during infancy may help the development of complex social interactions, as well as the acquisition of experience-dependent knowledge and skills to shape neuronal connectivity.
* A chimpanzee recognizes synthetic speech with significantly reduced acoustic cues to phonetic content. Heimbauer, L. A., Beran, M. J., & Owren, M. J. (Dept of Psych., Georgia State Univ., P.O. Box 5010, Atlanta, GA 30302-5010 [e-mail: [email protected]]). Current Biology, 2011, 21, 1210-1214, <www.sciencedirect.com/science/article/pii/S096098221100652X>.
. . . “A long-standing debate concerns whether humans are specialized for speech perception, which some researchers argue is demonstrated by the ability to understand synthetic speech with significantly reduced acoustic cues to phonetic content. We tested a chimpanzee (Pan troglodytes) that recognizes 128 spoken words, asking whether she could understand such speech. Three experiments presented 48 individual words, with the animal selecting a corresponding visuographic symbol from among four alternatives. Experiment 1 tested spectrally reduced, noise-vocoded (NV) synthesis, originally developed to simulate input received by human cochlear-implant users. Experiment 2 tested “impossibly unspeechlike” sine-wave (SW) synthesis, which reduces speech to just three moving tones. Although receiving only intermittent and noncontingent reward, the chimpanzee performed well above chance level, including when hearing synthetic versions for the first time. Recognition of SW words was least accurate but improved in experiment 3 when natural words in the same session were rewarded. The chimpanzee was more accurate with NV than SW versions, as were 32 human participants hearing these items. The chimpanzee’s ability to spontaneously recognize acoustically reduced synthetic words suggests that experience rather than specialization is critical for speech-perception capabilities that some have suggested are uniquely human.”
* Spontaneous prosocial choice by chimpanzees. Horner, V., Carter, J. D., Suchak, M., & de Waal, F. B. M. (Psych. Dept, Emory Univ., Atlanta, GA 30322 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 13847-13851, <www.pnas.org/content/108/33/13847.full>.
. . . “The study of human and primate altruism faces an evolutionary anomaly: there is ample evidence for altruistic preferences in our own species and growing evidence in monkeys, but one of our closest relatives, the chimpanzee (Pan troglodytes), is viewed as a reluctant altruist, acting only in response to pressure and solicitation. Although chimpanzee prosocial behavior has been reported both in observational captive studies and in the wild, thus far Prosocial Choice Tests have failed to produce evidence. However, methodologies of previous Prosocial Choice Tests may have handicapped the apes unintentionally. Here we present findings of a paradigm in which chimpanzees chose between two differently colored tokens: one “selfish” token resulting in a reward for the actor only (1/0), and the other “prosocial” token rewarding both the actor and a partner (1/1). Seven female chimpanzees, each tested with three different partners, showed a significant bias for the prosocial option. Prosocial choices occurred both in response to solicitation by the partner and spontaneously without solicitation. However, directed requests and pressure by the partner reduced the actor’s prosocial tendency. These results draw into question previous conclusions indicating that chimpanzees have a limited sensitivity to the needs of others and behave prosocially only in response to significant prompting.”
* Aping expressions? Chimpanzees produce distinct laugh types when responding to laughter of others. Davila-Ross, M., Allcock, B., Thomas, C., & Bard, K. A. (Centre for Comp. & Evol. Psych., Dept of Psych., King Henry Bldg, Univ. of Portsmouth, PO1 2DY, Portsmouth, U.K. [e-mail: [email protected]]). Emotion, 2011, DOI: 10.1037/a0022594, 1528-3542, <emo-aping-expressions-022811[1].pdf>.
. . . Humans have the ability to replicate the emotional expressions of others even when they undergo different emotions. Such distinct responses of expressions, especially positive expressions, play a central role in everyday social communication of humans and may give the responding individuals important advantages in cooperation and communication. The present work examined laughter in chimpanzees to test whether nonhuman primates also use their expressions in such distinct ways. The approach was first to examine the form and occurrence of laugh replications (laughter after the laughter of others) and spontaneous laughter of chimpanzees during social play and then to test whether their laugh replications represented laugh-elicited laugh responses (laughter triggered by the laughter of others) by using a quantitative method designed to measure responses in natural social settings. The results of this study indicated that chimpanzees produce laugh-elicited laughter that is distinct in form and occurrence from their spontaneous laughter. These findings provide the first empirical evidence that nonhuman primates have the ability to replicate the expressions of others by producing expressions that differ in their underlying emotions and social implications. The data further showed that the laugh-elicited laugh responses of the subjects were closely linked to play maintenance, suggesting that chimpanzees might gain important cooperative and communicative advantages by responding with laughter to the laughter of their social partners. Notably, some chimpanzee groups of this study responded more with laughter than others, an outcome that provides empirical support of a socialization of expressions in great apes similar to that of humans.
* Endoparasites in wild animals at the zoological garden in Skopje, Macedonia. Atanaskova, E., Kochevski, Z., Stefanovska, J., & Nikolovski, G. (Dept of Preventive Med. for Pets & Ungulates, Fac. of Vet. Med., LazarPop-Trajkov 5-7, 1000 Skopje, Republic of Macedonia [e-mail: [email protected]]). Journal of Threatened Taxa, 2011, 3, 1955-1958, <www.threatenedtaxa.org/ZooPrintJournal/2011/July/elena.htm>.
. . . “The health status of animals held in captivity depends on many factors, like: diet, environment, enclosure conditions and animal management. Since one of the main causes of death in wild animals are parasitic infections, regular parasite control and antiparasitic treatment are of great importance in zoo animals. Our study presents an overview of parasites found in animals kept at Skopje Zoo, Macedonia, from 2007 to 2009. Parasite detection was done by fecal analysis using ZnSO4 flotation method. The most common parasites identified were Toxocara sp., Toxascaris leonina, Strongyloides sp., Trichostrongylus sp. and Eimeria sp. In 2007 the percentage of infected animals was 21.4%. In 2008 the percentage of infected animals was about 32.14% and in 2009 it was 28.57%, which is relatively high (and which included Macaca sylvanus infected with Oesophagostomum sp.). The reconstruction of the enclosures during the past three years, when many animals were transferred from one enclosure to another and were mixed with other species of animals, probably caused such a high percentage of parasitic infection. With an appropriate program of antiparasitic treatment and measures to prevent parasitic infection, this percentage can decrease.”
* Organochlorine insecticide poisoning in golden langurs Trachypithecus geei. Pathak, D. C. (Dept of Pathology, College of Vet. Sci., Assam Agricultural Univ., Khanapara, Guwahati, Assam 781022, India [e-mail: [email protected]]). Journal of Threatened Taxa, 2011, 3, 1959-1960, <www.threatenedtaxa.org/ZooPrintJournal/2011/July/pathak.htm>.
. . . Organochlorine insecticide poisoning was recorded in three golden langurs in Chakrashila Wildlife Sanctuary in Kokrajhar district of Assam during the month of December, 2008. The poisoning was due to prolonged ingestion of rubber plant leaves sprayed with the insecticide in a rubber plantation adjacent to the sanctuary. Though no specific gross lesions were observed, histopathologically, centilobular hepatic necrosis, mild renal degeneration, necrotic enteritis, pulmonary congestion and neuronal degeneration were recorded in all three animals.
* Community structure and diversity of tropical forest mammals: Data from a global camera trap network. Ahumada, J. A., Silva, C. E. F., Gajapersad, K., Hallam, C., Hurtado, J., Martin, E., McWilliam, A., Mugerwa, B., O’Brien, T., Rovero, F., Sheil, D., Spironello, W. R., Winarni, N., & Andelman, S. J. (Conservation International, Arlington, VA 22202 [e-mail: j.ahumada@ conservation.org]). Philosophical Transactions of the Royal Society, B, 2011, 366, 2703-2711, <rstb.
royalsocietypublishing.org/content/366/1578/2703.full>.
. . . “Terrestrial mammals are a key component of tropical forest communities as indicators of ecosystem health and providers of important ecosystem services. However, there is little quantitative information about how they change with local, regional and global threats. In this paper, the first standardized pantropical forest terrestrial mammal community study, we examine several aspects of terrestrial mammal species and community diversity (species richness, species diversity, evenness, dominance, functional diversity, and community structure) at seven sites around the globe using a single standardized camera trapping methodology approach. The sites—located in Uganda, Tanzania, Indonesia, Lao PDR, Suriname, Brazil and Costa Rica—are surrounded by different landscape configurations, from continuous forests to highly fragmented forests. We obtained more than 51,000 images and detected 105 species of mammals with a total sampling effort of 12,687 camera trap days. We find that mammal communities from highly fragmented sites have lower species richness, species diversity, functional diversity and higher dominance when compared with sites in partially fragmented and continuous forest. We emphasize the importance of standardized camera trapping approaches for obtaining baselines for monitoring forest mammal communities so as to adequately understand the effect of global, regional and local threats and appropriately inform conservation actions.”
* Induced pluripotent stem cells from highly endangered species. Ben-Nun, I. F., Montague, S. C., Houck, M. L., Tran, H. T., Garitaonandia, I., Leonardo,T. R., Wang, Y.-C., Charter, S. J., Laurent, L. C., Ryder, O. A., & Loring, J. F. (J. F. L., Ctr for Regenerative Med., 4122 Sorrento Valley Blvd, Ste 107, SP30-3021, San Diego, CA 92121 [e-mail: [email protected]]). Nature Methods, 2011, <www.nature.com/nmeth/journal/vaop/ncurrent/full/nmeth.1706.html>.
. . . “For some highly endangered species there are too few reproductively capable animals to maintain adequate genetic diversity, and extraordinary measures are necessary to prevent extinction. We report generation of induced pluripotent stem cells (iPSCs) from two endangered species: a primate, the drill, Mandrillus leucophaeus, and the nearly extinct northern white rhinoceros, Ceratotherium simum cottoni. iPSCs may eventually facilitate reintroduction of genetic material into breeding populations.”
* Fever from the forest: Prospects for the continued emergence of sylvatic dengue virus and its impact on public health. Vasilakis, N., Cardosa, J., Hanley, K. A., Holmes, E. C., & Weaver, S. C. (Dept of Pathology, Univ. of Texas Med. Branch, Galveston, TX 77555 [e-mail: [email protected]]). Nature Reviews Microbiology, 2011, 9, 532-541, <www.nature.com/nrmicro/journal/v9/n7/full/nrmicro2595.html>.
. . . “The four dengue virus (DENV) serotypes that circulate among humans emerged independently from ancestral sylvatic progenitors that were present in non-human primates, following the establishment of human populations that were large and dense enough to support continuous inter-human transmission by mosquitoes. This ancestral sylvatic-DENV transmission cycle still exists and is maintained in non-human primates and Aedes mosquitoes in the forests of Southeast Asia and West Africa. Here, we provide an overview of the ecology and molecular evolution of sylvatic DENV and its potential for adaptation to human transmission. We also emphasize how the study of sylvatic DENV will improve our ability to understand, predict and, ideally, avert further DENV emergence.”
* Cross-species transmission of a novel adenovirus associated with a fulminant pneumonia outbreak in a New World monkey colony. Chen, E. C., Yagi, S., Kelly, K. R., Mendoza, S. P., Maninger, N., Rosenthal, A., Spinner, A., Bales, K. L., Schnurr, D. P., Lerche, N. W., & Chiu, C. Y. (C. Y. C., Dept of Lab. Med., UCSF, San Francisco, CA 94122 [e-mail: [email protected]]). PLoS Pathogens, 2011, 7[7], e1002155.
<dx.doi.org/10.1371%2Fjournal.ppat.1002155>.
. . . “Adenoviruses are DNA viruses that naturally infect many vertebrates, including humans and monkeys, and cause a wide range of clinical illnesses in humans. Infection from individual strains has conventionally been thought to be species-specific. Here we applied the Viro-chip, a pan-viral microarray, to identify a novel adenovirus (TMAdV, titi monkey adenovirus) as the cause of a deadly outbreak in a closed colony of New World monkeys (titi monkeys; Callicebus cupreus) at the California National Primate Research Center (CNPRC). Among 65 titi monkeys housed in a building, 23 (34%) developed upper respiratory symptoms that progressed to fulminant pneumonia and hepatitis, and 19 of 23 monkeys, or 83% of those infected, died or were humanely euthanized. Whole-genome sequencing of TMAdV revealed that this adenovirus is a new species and highly divergent, sharing < 57% pairwise nucleotide identity with other adenoviruses. Cultivation of TMAdV was successful in a human A549 lung adenocarcinoma cell line, but not in primary or established monkey kidney cells. At the onset of the outbreak, the researcher in closest contact with the monkeys developed an acute respiratory illness, with symptoms persisting for 4 weeks, and had a convalescent serum sample seropositive for TMAdV. A clinically ill family member, despite having no contact with the CNPRC, also tested positive, and screening of a set of 81 random adult blood donors from the western United States detected TMAdV-specific neutralizing antibodies in 2 individuals (2/81, or 2.5%). These findings raise the possibility of zoonotic infection by TMAdV and human-to-human transmission of the virus in the population. Given the unusually high case fatality rate from the outbreak (83%), it is unlikely that titi monkeys are the native host species for TMAdV, and the natural reservoir of the virus is still unknown. The discovery of TMAdV, a novel adenovirus with the capacity to infect both monkeys and humans, suggests that adenoviruses should be monitored closely as potential causes of cross-species outbreaks.”
* African monkeys are infected by Plasmodium falciparum nonhuman primate-specific strains. Prugnolle, F., Ollomo, B., Durand, P., Yalcindag, E., Arnathau, C., Elguero, E., Berry, A., Pourrut, X., Gonzalez, J.-P., Nkoghe, D., Akiana, J., Verrier, D., Leroy, E., Ayala, F. J., & Renaud, F. (Inst de Recherche pour le Développement Montpellier, BP 64501, 34394 Montpellier Cedex 5, France [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 11948-11953, <www.pnas.org/content/108/29/11948.full>.
. . . “Recent molecular exploration of the Plasmodium species circulating in great apes in Africa has revealed the existence of a large and previously unknown diversity of Plasmodium. For instance, gorillas were found to be infected by parasites closely related to Plasmodium falciparum, suggesting that the human malignant malaria agent may have arisen after a transfer from gorillas. Although this scenario is likely in light of the data collected in great apes, it remained to be ascertained whether P. falciparum-related parasites may infect other nonhuman primates in Africa. Using molecular tools, we here explore the diversity of Plasmodium species infecting monkeys in Central Africa. In addition to previously described Hepatocystis and Plasmodium species (Plasmodium gonderi and Plasmodium sp DAJ-2004), we have found one African monkey to be infected by a P. falciparum-related parasite. Examination of the nuclear and mitochondrial genomes of this parasite reveals that it is specific of nonhuman primates, indicating that P. falciparum-related pathogens can naturally circulate in some monkey populations in Africa. We also show that at least two distinct genetic entities of P. falciparum infect nonhuman primates and humans, respectively. Our discoveries bring into question the proposed gorilla origin of human P. falciparum.”
* Low levels of SIV infection in sooty mangabey central memory CD4+ T cells are associated with limited CCR5 expression. Paiardini, M., Cervasi, B., Reyes-Aviles, E., Micci, L., Ortiz, A. M., Chahroudi, A., Vinton, C., Gordon, S. N., Bosinger, S. E., Francella, N., Hallberg, P. L., Cramer, E., Schlub, T., Chan, M. L., Riddick, N. E., Collman, R. G., Apetrei, C., Pandrea, I., Else, J., Munch, J., Kirchhoff, F., Davenport, M. P., Brenchley, J. M., & Silvestri (Yerkes NPRC, Emory University, Atlanta, GA 30322 [e-mail: [email protected]]). Nature Medicine, 2011, 17, 830-836, <www.nature.com/nm/journal/vaop/ncurrent/full/nm.2395.html>.
. . . “Naturally simian immunodeficiency virus (SIV)-infected sooty mangabeys do not progress to AIDS despite high-level virus replication. We previously showed that the fraction of CD4+CCR5+ T cells is lower in sooty mangabeys compared to humans and macaques. Here we found that, after in vitro stimulation, sooty mangabey CD4+ T cells fail to upregulate CCR5 and that this phenomenon is more pronounced in CD4+ central memory T cells (TCM cells). CD4+ T cell activation was similarly uncoupled from CCR5 expression in sooty mangabeys in vivo during acute SIV infection and the homeostatic proliferation that follows antibody-mediated CD4+ T cell depletion. Sooty mangabey CD4+ TCM cells that express low amounts of CCR5 showed reduced susceptibility to SIV infection both in vivo and in vitro when compared to CD4+ TCM cells of rhesus macaques. These data suggest that low CCR5 expression on sooty mangabey CD4+ T cells favors the preservation of CD4+ T cell homeostasis and promotes an AIDS-free status by protecting CD4+ TCM cells from direct virus infection.”
* Right handed Neandertals: Vindija and beyond. Frayer, D. W., Fiore, I., Lalueza-Fox, C., Radovcic, J., & Luca Bondioli, L. (Dept of Anthropology, Univ. of Kansas, Lawrence, KN 66045. [e-mail: [email protected]]). Journal of Anthropological Sciences, 2010, 88, 113-127.
. . . Seven Vindija (Croatia) Neandertal teeth, dated ~32,000 years ago, were analyzed to determine patterning of scratches on the anterior teeth. Oblique scratches exclusively on the labial faces of incisors and canines represent a distinctive pattern, characteristic of hand directed, non-masticatory activities. At Vindija and elsewhere these scratches reveal activities which were performed primarily with the right hand. The late Neandertals from Vindija, combined with other studies, show that European Neandertals were predominately right-handed with a ratio 15:2 (88.2%), a frequency similar to living people. Studies of teeth from Atapuerca extend this modern ratio to more than 500,000 years ago and increase the frequency of right-handers in the European fossil record to almost 94%. Species-wide, preferential right-handedness is a defining feature of modern Homo sapiens, tied to brain laterality and language with the ~9:1 ratio of right- to left-handers – a reflection of the link between left hemispheric dominance and language. Up-to-date behavioral and anatomical studies of Neandertal fossils and the recent discovery of their possession of the FOXP2 gene indicate Neandertals (and, very likely, their European ancestors) had linguistic capacities similar to living humans.
* Late Mousterian persistence near the Arctic Circle. Slimak, L., Svendsen, J. I., Mangerud, J., Plisson, H., Heggen, H. P., Brugčre, A., & Pavlov, P. Y. (CNRS, UMR 5608, TRACES, Univ. de Toulouse le Mirail, Maison de la Recherche, 5 Allées Antonio Machado, 31058 Toulouse Cedex 9, France [e-mail: [email protected]]). Science, 2011, 332, 841-845, <www.sciencemag.org/content/332/6031/841.full>.
. . . “Palaeolithic sites in Russian high latitudes have been considered as Upper Palaeolithic and thus representing an Arctic expansion of modern humans. Here we show that at Byzovaya, in the western foothills of the Polar Urals, the technological structure of the lithic assemblage makes it directly comparable with Mousterian Middle Palaeolithic industries that so far have been exclusively attributed to the Neandertal populations in Europe. Radiocarbon and optical-stimulated luminescence dates on bones and sand grains indicate that the site was occupied during a short period around 28,500 carbon-14 years before the present (about 31,000 to 34,000 calendar years ago), at the time when only Upper Palaeolithic cultures occupied lower latitudes of Eurasia. Byzovaya may thus represent a late northern refuge for Neandertals, about 1000 km north of earlier known Mousterian sites.”
* Strontium isotope evidence for landscape use by early hominins. Copeland, S. R., Sponheimer, M., de Ruiter, D. J., Lee-Thorp, J. A., Codron, D., le Roux, P. J., Grimes, V., & Richards, M. P. (Dept of Anthropology, Univ. of Colorado, 233 UCB, Boulder, CO 80309). Nature, 2011, 474, 76-78, <www.nature.com/nature/journal/v474/n7349/full/nature10149.html>.
. . . “Ranging and residence patterns among early hominins have been indirectly inferred from morphology, stone-tool sourcing, referential models and phylogenetic models. However, the highly uncertain nature of such reconstructions limits our understanding of early hominin ecology, biology, social structure and evolution. We investigated landscape use in Australopithecus africanus and Paranthropus robustus from the Sterkfontein and Swartkrans cave sites in South Africa using strontium isotope analysis, a method that can help to identify the geological substrate on which an animal lived during tooth mineralization. Here we show that a higher proportion of small hominins than large hominins had non-local strontium isotope compositions. Given the relatively high levels of sexual dimorphism in early hominins, the smaller teeth are likely to represent female individuals, thus indicating that females were more likely than males to disperse from their natal groups. This is similar to the dispersal pattern found in chimpanzees, bonobos and many human groups, but dissimilar from that of most gorillas and other primates. The small proportion of demonstrably non-local large hominin individuals could indicate that male australopiths had relatively small home ranges, or that they preferred dolomitic landscapes.”
* New adapiform primate of Old World affinities from the Devil’s Graveyard Formation of Texas. Kirk, E. C., & Williams, B. A. (Dept of Anthropology, Univ of Texas at Austin, 1 University Station C3200, Austin, TX 78712 [[email protected]]). Journal of Human Evolution, 2011, 61, 158-168, <www.ncbi.nlm.nih.gov/pubmed/21571354>.
. . . “Most adapiform primates from North America are members of an endemic radiation of notharctines. North American notharctines flourished during the Early and early Middle Eocene, with only two genera persisting into the late Middle Eocene. Here we describe a new genus of adapiform primate from the Devil’s Graveyard Formation of Texas. Mescalerolemur horneri, gen. et sp. nov., is known only from the late Middle Eocene (Uintan) Purple Bench locality. Phylogenetic analyses reveal that Mescalerolemur is more closely related to Eurasian and African adapiforms than to North American notharctines. In this respect, M. horneri is similar to its sister taxon Mahgarita stevensi from the late Duchesnean of the Devil’s Graveyard Formation. The presence of both genera in the Big Bend region of Texas after notharctines had become locally extinct provides further evidence of faunal interchange between North America and East Asia during the Middle Eocene. The fact that Mescalerolemur and Mahgarita are both unknown outside of Texas also supports prior hypotheses that low-latitude faunal assemblages in North America demonstrate increased endemism by the late Middle Eocene.”
* Late Miocene to Pliocene carbon isotope record of differential diet change among East African herbivores. Uno, K. T., Cerling, T. E., Harris,, J. M., Kunimatsu, Y., Leakey, M. G., Nakatsukasa, M., & Nakaya, H. (Dept of Geology & Geophysics, Univ. of Utah, Salt Lake City, UT 84112 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 6509-6514, <www.pnas.org/content/108/16/6509.full>.
. . . “Stable isotope and molecular data suggest that C4 grasses first appeared globally in the Oligocene. In East Africa, stable isotope data from pedogenic carbonate and fossil tooth enamel suggest a first appearance between 15–10 Ma and subsequent expansion during the Plio-Pleistocene. The fossil enamel record has the potential to provide detailed information about the rates of dietary adaptation to this new resource among different herbivore lineages. We present carbon isotope data from 452 fossil teeth that record differential rates of diet change from C3 to mixed C3/C4 or C4 diets among East African herbivore families at seven different time periods during the Late Miocene to the Pliocene (9.9–3.2 Ma). Significant amounts of C4 grasses were present in equid diets beginning at 9.9 Ma and in rhinocerotid diets by 9.6 Ma, although there is no isotopic evidence for expansive C4 grasslands in this part of the Late Miocene. Bovids and hippopotamids followed suit with individuals that had C4-dominated (>65%) diets by 7.4 Ma. Suids adopted C4-dominated diets between 6.5 and 4.2 Ma. Gomphotheriids and elephantids had mostly C3-dominated diets through 9.3 Ma, but became dedicated C4 grazers by 6.5 Ma. Deinotheriids and giraffids maintained a predominantly C3 diet throughout the record. The sequence of differential diet change among herbivore lineages provides ecological insight into a key period of hominid evolution and valuable information for future studies that focus on morphological changes associated with diet change.”
* Diet of Paranthropus boisei in the early Pleistocene of East Africa. Cerling, T. E., Mbua, E., Kirera, F. M., Manthi, F. K., Grine, F. E., Leakey, M. G., Sponheimer, M., & Uno, K. T. (Dept of Geology & Geophysics, Univ. of Utah, Salt Lake City, UT 84112-0111 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 9337-9341, <www.pnas.org/108/23/9337.full>.
. . . “The East African hominin Paranthropus boisei was characterized by a suite of craniodental features that have been widely interpreted as adaptations to a diet that consisted of hard objects that required powerful peak masticatory loads. These morphological adaptations represent the culmination of an evolutionary trend that began in earlier taxa such as Australopithecus afarensis, and presumably facilitated utilization of open habitats in the Plio-Pleistocene. Here, we use stable isotopes to show that P. boisei had a diet that was dominated by C4 biomass such as grasses or sedges. Its diet included more C4 biomass than any other hominin studied to date, including its congener Paranthropus robustus from South Africa. These results, coupled with recent evidence from dental microwear, may indicate that the remarkable craniodental morphology of this taxon represents an adaptation for processing large quantities of low-quality vegetation rather than hard objects.”
* Evolution of the V, D, and J gene segments used in the primate γδ T-cell receptor reveals a dichotomy of conservation and diversity. Kazen, A. R., & Adams, E. J. (E. J. A., Dept of Biochem. & Molecular Biol., Univ. of Chicago, Chicago, IL 60637 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, E332-E340, <www.pnas.org/content/108/29/E332.full>.
. . . “γδ T cells are an immunological enigma in that both their function in the immune response and the molecular mechanisms behind their activation remain unclear. These cells predominate in the epithelia and can be rapidly activated to provide an array of responses. However, no homologous γδ T-cell populations have been identified between humans and mice, and our understanding of what these cells recognize as ligands is limited. Here we take an alternative approach to understanding human γδ T-cell ligand recognition by studying the evolutionary forces that have shaped the V, D, and J gene segments that are used during somatic rearrangement to generate the γδ T-cell receptor. We find that distinctly different forces have shaped the γ and δ loci. The Vδ and Jδ genes are highly conserved, some even through to mouse. In contrast, the γ-locus is split: the Vγ9, Vγ10, and Vγ11 genes represent the conserved region of the Vγ gene locus whereas the remaining Vγ genes have been evolving rapidly, such that orthology throughout the primate lineage is unclear. We have also analyzed the coding versus silent substitutions between species within the V and J gene segments and find a preference for coding substitutions in the complementarity determining region loops of many of the V gene segments. Our results provide a different perspective on investigating human γδ T-cell recognition, demonstrating that diversification at particular γδ gene loci has been favored during primate evolution, suggesting adaptation of particular V domains to a changing ligand environment.”
* An X-linked haplotype of Neandertal origin is present among all non-African populations. Yotova, V., Lefebvre, J.-F., Moreau, C., Gbeha, E., Hovhannesyan, K., Bourgeois, S., Bédarida, S., Azevedo, L., Amorim, A., Sarkisian, T., Avogbe, P. H., Chabi, N., Dicko, M. H., Kou’ Santa Amouzou, E. S., Sanni, A., Roberts-Thomson, J., Boettcher, B., Scott, R. J., & Labuda, D. (D, L., Dépt de Pédiatrie, Univ. de Montréal, Montreal, Canada [e-mail: [email protected]]). Molecular Biology and Evolution, 2011, 28, 1957-1962,
<mbe.oxfordjournals.org/content/28/7/1957.full>.
. . . “Recent work on the Neandertal genome has raised the possibility of admixture between Neandertals and the expanding population of Homo sapiens who left Africa between 80 and 50 Kya (thousand years ago) to colonize the rest of the world. Here, we provide evidence of a notable presence (9% overall) of a Neandertal-derived X chromosome segment among all contemporary human populations outside Africa. Our analysis of 6,092 X-chromosomes from all inhabited continents supports earlier contentions that a mosaic of lineages of different time depths and different geographic provenance could have contributed to the genetic constitution of modern humans. It indicates a very early admixture between expanding African migrants and Neandertals prior to or very early on the route of the out-of-Africa expansion that led to the successful colonization of the planet.”
* Patterns of cranial shape diversification during the phylogenetic branching process of New World monkeys (Primates: Platyrrhini). Perez, S. I., Klaczko, J., Rocatti, G., & Dos Reis, S. F. (Museo de La Plata, Univ. Nac. de La Plata, CONICET, Paseo del Bosque s/n, 1900, La Plata, Argentina [e-mail: [email protected]]). Journal of Evolutionary Biology, 2011, 24, 1826-1835,
<onlinelibrary.wiley.com/doi/10.1111/j.1420-9101.2011.02309.x/full>.
. . . “One of the central topics in evolutionary biology is understanding the processes responsible for phenotypic diversification related to ecological factors. New World monkeys are an excellent reference system to investigate processes of diversification at macroevolutionary scales. Here, we investigate the cranial shape diversification related to body size and ecology during the phylogenetic branching process of platyrrhines. To investigate this diversification, we used geometric morphometric techniques, a molecular phylogenetic tree, ecological data and phylogenetic comparative methods. Our statistical analyses demonstrated that the phylogenetic branching process is the most important dimension to understand cranial shape variation among extant platyrrhines and suggested that the main shape divergence among the four principal platyrrhine clades probably occurred during the initial branching process. The phylogenetic conservatism, which is the retention of ancestral traits over time within the four principal platyrrhine clades, could be the most important characteristic of platyrrhine cranial shape diversification. Different factors might have driven early shape divergence and posterior relative conservatism, including genetic drift, stabilizing selection, genetic constraints owing to pleiotropy, developmental or functional constraint, lack of genetic variation, among others. Understanding the processes driving the diversification among platyrrhines will probably require further palaeontological, phylogenetic and comparative studies.”
* Aging of the cerebral cortex differs between humans and chimpanzees. Sherwood, C. C., Gordon, A. D., Allen, J. S., Phillips, K. A., Erwin, J. M., Hof, P. R., & Hopkins, W. D. (Dept of Anthropology, George Washington Univ., Washington, DC 20052 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 13029-13034, <www.pnas.org/content/108/32/13029.full>.
. . . “Several biological changes characterize normal brain aging in humans. Although some of these age-associated neural alterations are also found in other species, overt volumetric decline of particular brain structures, such as the hippocampus and frontal lobe, has only been observed in humans. However, comparable data on the effects of aging on regional brain volumes have not previously been available from our closest living relatives, the chimpanzees. In this study, we used MRI to measure the volume of the whole brain, total neocortical gray matter, total neocortical white matter, frontal lobe gray matter, frontal lobe white matter, and the hippocampus in a cross-sectional sample of 99 chimpanzee brains encompassing the adult lifespan from 10 to 51 y of age. We compared these data to brain structure volumes measured in 87 adult humans from 22 to 88 y of age. In contrast to humans, who showed a decrease in the volume of all brain structures over the lifespan, chimpanzees did not display significant age-related changes. Using an iterative age-range reduction procedure, we found that the significant aging effects in humans were because of the leverage of individuals that were older than the maximum longevity of chimpanzees. Thus, we conclude that the increased magnitude of brain structure shrinkage in human aging is evolutionarily novel and the result of an extended lifespan.”
* Phylogenetic rate shifts in feeding time during the evolution of Homo. Organ, C., Nunn, C. L., Machanda, Z., & Wrangham, R. W. (C. O., Dept of Organismic & Evol. Bio., Harvard Univ., Cambridge, MA 02138 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 14555-14559, <www.pnas.org/content/108/35/14555.full>.
. . . “Unique among animals, humans eat a diet rich in cooked and nonthermally processed food. The ancestors of modern humans who invented food processing (including cooking) gained critical advantages in survival and fitness through increased caloric intake. However, the time and manner in which food processing became biologically significant are uncertain. Here, we assess the inferred evolutionary consequences of food processing in the human lineage by applying a Bayesian phylogenetic outlier test to a comparative dataset of feeding time in humans and nonhuman primates. We find that modern humans spend an order of magnitude less time feeding than predicted by phylogeny and body mass (4.7% vs. predicted 48% of daily activity). This result suggests that a substantial evolutionary rate change in feeding time occurred along the human branch after the human–chimpanzee split. Along this same branch, Homo erectus shows a marked reduction in molar size that is followed by a gradual, although erratic, decline in H. sapiens. We show that reduction in molar size in early Homo (H. habilis and H. rudolfensis) is explicable by phylogeny and body size alone. By contrast, the change in molar size to H. erectus, H. neanderthalensis, and H. sapiens cannot be explained by the rate of craniodental and body size evolution. Together, our results indicate that the behaviorally driven adaptations of food processing (reduced feeding time and molar size) originated after the evolution of Homo but before or concurrent with the evolution of H. erectus, which was around 1.9 Mya.
* Living archaeology: Artefacts of specific nest site fidelity in wild chimpanzees. Stewart, F. A., Piel, A. K., & McGrew, W. C. (Leverhulme Ctr. for Human Evol. Studies, Dept of Biol. Anthro., Univ. of Cambridge, Henry Wellcome Bldg, Fitzwilliam St, Cambridge CB2 1QH, U.K. [e-mail: [email protected]]). Journal of Human Evolution, 2011, 61, 388-395, <www.sciencedirect.com/science/article/pii/S0047248411001357>.
. . . “Savanna chimpanzees are known to re-use areas of the landscape for sleep, and patterns of chimpanzee sleeping site re-use are proposed as a referential model for early hominin archaeological site formation. We recorded the prevalence of deformed but healed branches and remnants of dead branches found around fresh nests at the savanna site of Issa in Ugalla, Tanzania. These old nest scars were found in 79% of 112 beds. We also randomly selected potential nesting locations for a subset of 32 beds within the same trees, and found nest scars in only 19% of these ‘control’ locations. We then monitored 275 nests for up to 19 months for decay, regeneration of new branches, and re-use. Of these 275 nest locations, 24% were re-used within the first nine months of monitoring, and most re-use occurred when the nest had already decayed and was not easily visible from the ground. After 18 months, the proportion of specific nest positions re-used increased to 48%. This fidelity is likely a result of the creation of ideally-shaped support structures and supple new growth for mattress material with successive use of nest locations. We propose that specific nest site re-use may not be a direct product of environmental determination, but a result of ‘niche construction’ through formation of good building sites within trees. Environmental modification through construction behavior may have influenced both chimpanzee and early hominin ranging, and thus leaves behind recognisable patterns of artefact deposition across the landscape.”
* Australopithecus sediba: A new species of Homo-like Australopith from South Africa. Berger, L. R., de Ruiter, D. J., Churchill, S. E., Schmid,, P., Carlson, K. J., Dirks, P. H. G. M., & Kibii, J. M. (Inst. for Human Evol., Univ. of the Witwatersrand, P. O. Wits, 2050 Wits, South Africa [e-mail: [email protected]]). Science, 2011, 333, 195-204, <www.sciencemag.org/content/328/5975/195.full>.
. . . “Despite a rich African Plio-Pleistocene hominin fossil record, the ancestry of Homo and its relation to earlier australopithecines remain unresolved. Here we report on two partial skeletons with an age of 1.95 to 1.78 million years. The fossils were encased in cave deposits at the Malapa site in South Africa. The skeletons were found close together and are directly associated with craniodental remains. Together they represent a new species of Australopithecus that is probably descended from Australopithecus africanus. Combined craniodental and postcranial evidence demonstrates that this new species shares more derived features with early Homo than any other australopith species and thus might help reveal the ancestor of that genus.”
* The endocast of MH1, Australopithecus sediba. Carlson, K. J., Stout, D., Jashashvili, T., de Ruiter, D. J., Tafforeau, P., Carlson, K., & Berger, L. R. (Inst. for Human Evol., Univ. of the Witwatersrand, Palaeosciences Centre, Private Bag 3, Wits 2050, South Africa [e-mail: [email protected]]). Science, 2011, 333, 1402-1407, <www.sciencemag.org/content/333/6048/1402.full>.
. . . The virtual endocast of MH1 (Australopithecus sediba), obtained from high-quality synchrotron scanning, reveals generally australopith-like convolutional patterns on the frontal lobes but also some foreshadowing of features of the human frontal lobes, such as posterior repositioning of the olfactory bulbs. Principal component analysis of orbitofrontal dimensions on australopith endocasts (MH1, Sts 5, and Sts 60) indicates that among these, MH1 orbitofrontal shape and organization align most closely with human endocasts. These results are consistent with gradual neural reorganization of the orbitofrontal region in the transition from Australopithecus to Homo, but given the small volume of the MH1 endocast, they are not consistent with gradual brain enlargement before the transition.
* A partial pelvis of Australopithecus sediba. Kibii, J. M., Churchill, S. E., Schmid, P., Carlson, K. J., Reed, N. D., de Ruiter, D. J., & Berger, L. R. (S. E. C., Dept of Evol. Anthro., Box 90383, Duke Univ., Durham, NC 27708 [e-mail: [email protected]]). Science, 2011, 333, 1407-1411, <www.sciencemag.org/content/333/6048/1407.full>.
. . . The fossil record of the hominin pelvis reflects important evolutionary changes in locomotion and parturition. The partial pelves of two individuals of Australopithecus sediba were reconstructed from previously reported finds and new material. These remains share some features with australopiths, such as large biacetabular diameter, small sacral and coxal joints, and long pubic rami. The specimens also share derived features with Homo, including more vertically oriented and sigmoid-shaped iliac blades, greater robusticity of the iliac body, sinusoidal anterior iliac borders, shortened ischia, and more superiorly oriented pubic rami. These derived features appear in a species with a small adult brain size, suggesting that the birthing of larger-brained babies was not driving the evolution of the pelvis at this time.
* Australopithecus sediba hand demonstrates mosaic evolution of locomotor and manipulative abilities. Kivell, T. L., Kibii, J. M., Churchill, S. E., Schmid, P., & Berger, L. R. (J. M. K., Inst. for Human Evol., Univ. of the Witwatersrand, Private Bag 3, Wits 2050, South Africa [e-mail: [email protected]]). Science, 2011, 333, 1411-1417, <www.sciencemag.org/content/333/6048/1411.full>.
. . . “Hand bones from a single individual with a clear taxonomic affiliation are scarce in the hominin fossil record, which has hampered understanding the evolution of manipulative abilities in hominins. Here we describe and analyze a nearly complete wrist and hand of an adult female [Malapa Hominin 2 (MH2)] Australopithecus sediba from Malapa, South Africa (1.977 million years ago). The hand presents a suite of Australopithecus-like features, such as a strong flexor apparatus associated with arboreal locomotion, and Homo-like features, such as a long thumb and short fingers associated with precision gripping and possibly stone tool production. Comparisons to other fossil hominins suggest that there were at least two distinct hand morphotypes around the Plio–Pleistocene transition. The MH2 fossils suggest that Au. sediba may represent a basal condition associated with early stone tool use and production.”
* The foot and ankle of Australopithecus sediba. Zipfel, B., DeSilva, J. M., Kidd, R. S., Carlson, K. J., Churchill, S. E. , & Berger, L. R. (Inst. for Human Evol., Univ. of the Witwatersrand, P. O. Wits, 2050 Wits, South Africa [e-mail: [email protected]]). Science, 2011, 333, 1417-1420, <www.sciencemag.org/content/333/6048/1417.full>.
. . . A well-preserved and articulated partial foot and ankle of Australopithecus sediba, including an associated complete adult distal tibia, talus, and calcaneus, have been discovered at the Malapa site, South Africa, and reported in direct association with the female paratype Malapa Hominin 2. These fossils reveal a mosaic of primitive and derived features that are distinct from those seen in other hominins. The ankle (talocrural) joint is mostly humanlike in form and inferred function, and there is some evidence for a humanlike arch and Achilles tendon. However, Au. sediba is apelike in possessing a more gracile calcaneal body and a more robust medial malleolus than expected. These observations suggest, if present models of foot function are correct, that Au. sediba may have practiced a unique form of bipedalism and some degree of arboreality. Given the combination of features in the Au. sediba foot, as well as comparisons between Au. sediba and older hominins, homoplasy is implied in the acquisition of bipedal adaptations in the hominin foot.
* Australopithecus sediba at 1.977 Ma and implications for the origins of the genus Homo. Pickering, R., Dirks, P. H. G. M., Jinnah, Z., de Ruiter, D. J., Churchill, S. E., Herries, A. I. R., Woodhead, J. D., Hellstrom, J. C., & Berger, L. R. (R. P., Sch. of Earth Sci., Univ. of Melbourne, Victoria, 3010, Australia [e-mail: [email protected]]). Science, 2011, 333, 1421-1423, <www.sciencemag.org/content/333/6048/1421.full>.
. . . Newly exposed cave sediments at the Malapa site include a flowstone layer capping the sedimentary unit containing the Australopithecus sediba fossils. Uranium-lead dating of the flowstone, combined with paleomagnetic and stratigraphic analysis of the flowstone and underlying sediments, provides a tightly constrained date of 1.977 ± 0.002 million years ago (Ma) for these fossils. This refined dating suggests that Au. sediba from Malapa pre-dates the earliest uncontested evidence for Homo in Africa.
* Short term pharmacological immobilization in macaque monkeys. Votava, M., Hess, L., Schreiberová, J., Málek, J., & Štein, K. (Dept of Pharmacology, 2nd Fac. of Med., Charles Univ. in Prague, V Úvalu 84, Prague 5, 15 006 Czech Republic [e-mail: [email protected]]). Veterinary Anaesthesia and Analgesia, 2011, 38, 490-493, <onlinelibrary.wiley.com/doi/10.1111/j.1467-2995.2011.00637.x/full>.
. . . In order to develop a safe and effective immobilization protocol in rhesus monkeys, which is not based on dissociative anaesthetic agent, 20 rhesus monkeys, weighing 2.6–8.0 kg, 1–3 years of age, of both sexes, received 50 µg/kg medetomidine, 0.25 mg kg/1 midazolam and 5 µg/kg fentanyl with 150 IU hyaluronidase intramuscularly. The animals were closely observed for behavioral changes and reaction to sound stimulus. Pulse rate and oxygen saturation of hemoglobin (SpO2) were monitored every 5 minutes, for 20 minutes. After this period, 250 µg/kg atipamezole or a placebo was administered IM and behavioral changes were closely observed. Full immobilization was observed after mean 269 ± SD 116 seconds. Ten minutes after injection mean arterial oxygen saturation of hemoglobin was 94 ± 4%, but did not fall significantly further. The median pulse rate was 116 beats/ minute 5 minutes after the administration of the drug. This level further decreased to a median level of 108 beats/minute 20 minutes after the drug’s administration. The median time to recover from immobilization was significantly shorter after atipamezole administration compared to placebo (2.7 versus 55 minutes). All animals awoke smoothly and no side effects such as vomiting or agitation were observed. The present study demonstrates that 20-minute pharmacological immobilization with a combination of midazolam, medetomidine, and fentanyl is feasible in rhesus monkeys with minimal effect on heart rate.
* Sex identification using the ZFX and ZFY genes in common marmosets (Callithrix jacchus). Takabayashi, S., & Katoh, H. (Exp. Animals Inst., Hamamatsu Univ. Sch. of Med., Shizuoka, 431-3192, Japan). Experimental Animals, 2011, 60, 417-420, <www.jstage.jst.go.jp/article/expanim/60/4/60_417/_article>.
. . . “We investigated sex determination via the ZFX and ZFY genes using PCR-RFLP in the common marmoset. We designed a novel primer set to detect ZFX and ZFY. A 483-bp band from the ZFX gene and a 471-bp band from the ZFY gene were amplified. Sequencing data of the products amplified from ZFX and ZFY showed the recognition sites of two restriction enzymes, DdeI and MseI, respectively. After digestion of the products using each enzyme, we found that the band patterns between females and males were different. PCR-based sex identification might provide a tool for further breeding studies and experimental embryological studies using marmosets.”
* Life at the top: Rank and stress in wild male baboons. Gesquiere, L. R., Learn, N. H., Simao, M. C. M., Onyango, P. O., Alberts, S. C., & Altmann, J. (Dept of Ecol. & Evol. Biol., Princeton Univ., Princeton, NJ 08544 [e-mail: [email protected]]). Science, 2011, 333, 357-360, <www.sciencemag.org/content/333/6040/357.full>.
. . . In social hierarchies, dominant individuals experience reproductive and health benefits, but the costs of social dominance remain a topic of debate. Prevailing hypotheses predict that higher-ranking males experience higher testosterone and glucocorticoid (stress hormone) levels than lower-ranking males when hierarchies are unstable but not otherwise. In this long-term study of rank-related stress in a natural population of savannah baboons (Papio cynocephalus), high-ranking males had higher testosterone and lower glucocorticoid levels than other males, regardless of hierarchy stability. The singular exception was for the highest-ranking (alpha) males, who exhibited both high testosterone and high glucocorticoid levels. In particular, alpha males exhibited much higher stress hormone levels than second-ranking (beta) males, suggesting that being at the very top may be more costly than previously thought.
* * *

The road is long, and the road can be hard –
but good friends make the road worthwhile!
And yet, the road always has an end –
* * *