 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 28 NUMBER 2 APRIL 1989
Articles and Notes
Training Female Rhesus Monkeys to Cooperate during In-Homecage Venipuncture, by R. Vertein & V. Reinhardt ...... 1
Chimps, Ethics, and Numbers: Appropriate Sample Sizes for Tests of Vaccines, by B. Holland ...... 4
Heart Rate and Activity of Rhesus Monkeys in Response to Routine Events, by S. W. Line, K. N. Morgan, H. Markowitz, & S. Strong ...... 9
Reproductive Biology at the Yerkes Regional Primate Research Center and the Nature of Animal Welfare Extremism, by R. D. Nadler & J. F. Dahl ...... 13
Long-term Effects of a Natural Foraging Task on Aggression and Stereotypies in Socially Housed Pigtail Macaques, by M. L. Boccia ...... 18
The Use of Toys for Primate Environmental Enrichment, by D. S. B. Watson, B. J. Houston, & G. E. Macallum ...... 20
Kong Toys as Enrichment Devices for Singly-Caged Macaques, by C. Crockett, J. Bielitzki, A. Carey, & A. Velez ...... 21
News, Information, and Announcements
Information Available ...... 8
. . PSIC, WRPRC FAX Number, MMWR, PIC,
IATA Publishes Regulations, A.P.E. Project
AAPA Membership Drive ...... 12
Regulations Proposed on Animal Welfare Act ...... 15
News Briefs ...... 16
. . Ruling on Silver Spring Monkeys, Immuno Lawsuit Dismissed,
Yellow Fever Virus Activity in Trinidad
Animal Care Courses Offered ...... 22
Research and Education Opportunities ...... 24
. . Digit Fund, Earthwatch, Wildlife Preservation Trust,
ASP Summer Internship List, School for Field Studies,
Madagascar Tour, University Research Expeditions
Editors' Notes ...... 25
Departments
Address Changes ...... 23
Position Available ...... 23
Recent Books and Articles ...... 26
* * *
Russell Vertein and Viktor Reinhardt
University of Wisconsin
Introduction
Blood sampling via venipuncture is a common procedure with laboratory rhesus monkeys, but it can be distressing for the individual animal and frustrating for the animal caretaker, animal technician, or veterinarian. The animal may show signs of fear (e.g., fear-grinning, screaming, acute diarrhea) and/or physical resistance (e.g., refusing to enter the squeeze cage, struggling), while caretakers often use anesthetics (Weller, 1981; Schiffer et al., 1984; Braunstein & Asch, 1986; Daniel et al., 1988; Johnson et al., 1988) and/or mechanical squeeze devices (Elvidge et al., 1976; Line et al., 1987; Loomis et al., 1980; Fuller et al., 1984; Monfort et al., 1987; Clarke et al., 1988) to control the animal.
Two publications indicate that such unsatisfactory encounters can be avoided. In a study by Rosenblum et al. (1981), "All animals [juvenile and adult rhesus monkeys] were trained to present an arm through a small cage opening. In this manner blood samples were drawn from untranquilized fully alert animals." In a study involving female rhesus monkeys, Billiard et al. (1985) reported that, "Blood samples were obtained by venipuncture from an animal two or three times each week by the volunteer method," in which an animal learned to present a leg out of a transfer cage for venipuncture. Unfortunately no details are given in either report about how the animals were trained to cooperate during venipuncture.
The present paper describes a training schedule for caged adult female rhesus monkeys living in compatible pairs. The partners of each pair were trained to cooperate during venipuncture in their homecage.
Figure 1: Restraint training of 9-year-old Fox and her 14-year-old companion Xantha. Both animals share a food reward offered by the animal caretaker. Note that the animals are restrained but not squeezed.
Methods
Eight healthy, 5- to 16-year-old female rhesus monkeys (Macaca mulatta) were transferred into stainless steel squeeze cages. Each cage was 85x85x85 cm large and equipped with a movable back wall permitting optional reduction of cage space. None of the 8 animals had been exposed to such a cage before. They were all familiar with the attendant caretaker (senior author), out of whose hand they took favored food, such as raisins, peanuts, grapes, apples, and bananas. After one week of habituation the monkeys were trained by the caretaker once a day on 24 consecutive work days according to the following schedule:
Days 1-5: 15-minute restraint training sessions
Step A: The caretaker gently pulls the movable back wall
toward the front of the cage until the cage space available to the
animals is reduced by about 75%. The two monkeys are
not squeezed in this restraint condition but
have room enough to turn freely around, to walk back and forth in the
front of the cage, and to climb up the back and front walls of
the cage. The caretaker offers favored food in his hand
for 2 minutes (Figure 1).
Step B: The animals are left alone in the reduced cage for
5 minutes.
Step C: The caretaker offers food for 2 minutes.
Step D: Same as Step B.
Step E: The caretaker pushes the back wall into its normal
position and rewards the animals with favored food for 1 minute.
Days 6-7: 15-minute touch-restraint training sessions
Step F: The caretaker restrains the animals as in
Step A,
offers them food for 1/2 minute, and finally touches each animal's legs
several times during 1 1/2 minutes.
Step G: Same as Step B.
Step H: Same as Step F.
Step I: Same as Step B.
Step J: Same as Step E.
Days 8-23: 8-minute leg-out restraint training sessions
Step K: Same as Step A.
Step L: The caretaker gets hold of an animal's
leg through the partly opened cage door. He then gently pulls the leg
toward himself and caresses the animal's thighs and calves
(Figure 2). He keeps
the animal in this position for approximately 20 seconds before releasing
its leg and offering it some food in his hand. He repeats this
with the other animal. Each pair is trained in this way
for 5 minutes each session.
Step M: Same as Step E.
Day 24: Venipuncture
Step N: The caretaker restrains the animals as in Step A and collects a blood sample from each animal via puncture of its saphenous vein (Figure 3). He then pushes the back wall into its normal position and offers both animals a food reward during a few seconds.
Figure 2: Leg-out restraint training of Fox. Note that Fox is restrained but not squeezed.
Results
All 8 animals took favored food from the caretaker's hand in Step A, C, and E (reduced cage space) by Day 4 of the training.
The caretaker successfully went through Step L (leg-holding) for the first time with all 8 animals on Day 9 of the training.
On Day 24 of the training, blood collection via venipuncture was successfully accomplished in all 8 animals within a total of 17 minutes. Three monkeys actively presented a leg to the caretaker on this occasion. The other 5 monkeys were less cooperative but showed no signs of fear or resistance to having a leg gently pulled through the cage door and their saphenous vein punctured. All monkeys accepted the food reward from the caretaker's hand after the venipuncture.
The training of the 8 rhesus monkeys was completed within 24 work days. It required a total of 250 minutes, that is 31 minutes per individual animal.
The trained animals have been assigned to research protocols requiring regular blood collection; in-cage venipuncture has become routine for them. It is important to note that the animals are cooperative not only with their attendant caretaker but with anyone with whom they are familiar and who is experienced in gently handling nonhuman primates.
Figure 3: Fox shows no signs of fear or resistance while being venipunctured. Xantha waits for her turn. Note that the animals are restrained but not squeezed.
Discussion
The in-cage venipuncture training of rhesus monkeys described in this report is a simple means to minimize distress in animals that must be handled. The little time required to train an animal pays off for the caretaker, animal technician, or veterinarian who is more satisfied working with cooperative animals than with fearful, resisting animals. Good cooperation between animal and personnel during venipuncture in an animal's homecage, with a compatible companion present for psychological support, is not only beneficial for the animal and satisfactory for the personnel, but is also of value to the scientist who is concerned that her/his research animal is not subjected to undue distress.
We are currently adapting our training schedule for in-homecage venipunture of adult male rhesus monkeys.
References
Billiard, R.B., Richardson, D., Anderson, E., Mahajan, D., & Little, B. (1985). The effect of chronic and acyclic elevation of circulating androstenedione or estrone concentrations on ovarian function in the rhesus monkey. Endocrinology, 116, 2209-2220.
Braunstein, G. D., & Asch, R.H. (1986). Pregnancy-specific beta-glycoprotein concentrations throughout pregnancy in the rhesus monkey. Journal of Clinical Endocrinology and Metabolism, 62, 1264-1270.
Clarke, A.S., Mason, W.A., & Moberg, G.P. (1988). Interspecific contrasts in responses of macaques to transport cage training. Laboratory Animal Science, 38, 305-309.
Daniel, M.D., Letvin, N.L., Sehgal, P.K., Schmidt, D.K., Silvia, D.P., Solomon, K.R., Hodi, F.S., Ringer, D.J., Hunt, R.D., King, N.W., & Desrosiers, R.C. (1988). Prevalence of antibodies to 3 retroviruses in a captive colony of macaque monkeys. International Journal of Cancer, 41, 601-608.
Elvidge, H., Challis, J.R.G., Robinson, J.S., Roper, C., & Thorburn, G.D. (1976). Influence of handling and sedation on plasma cortisol in rhesus monkeys (Macaca mulatta). Journal of Endocrinology, 70, 324-326.
Fuller, G. B., Hobson, W. C., Reyes, F. I., Winter, J. S. D., & Faiman, C. (1984). Influence of restraint and ketamine anesthesia on adrenal steroids, progesterone, and gonadotropins in rhesus monkeys. Proceedings of the Society for Experimental Biology and Medicine, 175, 487-490.
Johnson, M.S., Ottobre, A.C., & Ottobre, J.S. (1988). Prostaglandin production by corpora lutea of rhesus monkeys: Characterization of incubation conditions and examination of putative regulators. Biology of Reproduction, 39, 839-846.
Line, S.W., Clarke, A., & Markowitz, H. (1987). Plasma cortisol of female rhesus monkeys in response to acute restraint. Laboratory Primate Newsletter, 26, 1-4.
Loomis, M.R., Henrickson, R.V., & Anderson, J.H. (1980). Effects of ketamine hydrochloride on the hemogram of rhesus monkeys (Macaca mulatta). Laboratory Animal Science, 30, 851-853.
Monfort, S.L., Hess, D.L., Shideler, S.E., Samuels, S.J., Hendrickx, A.G., & Lasley, B.L. (1987). Comparision of serum estradiol to urinary estrone conjugates in the rhesus macaque (Macaca mulatta). Biology of Reproduction, 37, 832-837.
Rosenblum, I.Y., & Coulston, F. (1981). Normal range of longitudinal blood chemistry and hematology values in juvenile and adult rhesus monkeys (Macaca mulatta). Ecotoxicology and Environmental Safety, 5, 401-411.
Schiffer, S.P., Gillett, C.S., & Ringler, D.H. (1984). Activated coagulation time for rhesus monkeys (Macaca mulatta). Laboratory Animal Science, 34, 191-193.
Weller, W. (1981). Normalwerte von Rhesusaffen bei langfristiger kontrollierter Haltung. Zeitschrift für Versuchstierkunde, 23, 30-41.
-------------------------------------------------------------------
Authors' address: Wisconsin Regional Primate Research Center, 1223
Capitol Court, Madison, WI 53715.
We are grateful to Mr. Robert Dodsworth for preparing the photographs of
Figures 1-3 and to Mr. John Wolf for editing this manuscript.
This project was supported by NIH grant RR-00167 to the Wisconsin
Regional Primate Research Center.
* * *
Bart Holland
UMDNJ--New Jersey Medical School
Introduction
"Physiological experiment on animals is justifiable for real investigation," wrote Charles Darwin (1881), "but not for mere damnable and detestable curiosity." Current research on animals is carried out in an atmosphere of tension between two conflicting ethical imperatives. Experiments are carried out on undeniably sentient animals such as primates, in order to investigate innovations in medical thinking which are expected to advance human therapy; an ethical imperative to advance the human condition as much as possible is offered as justification. Respect for all living things and attentiveness to animal welfare are also ethical imperatives, however. In current practice, an uneasy balance is struck, whereby some knowledge is considered worthwhile enough that experimentation on animals is justified. Even so, the government can and does regulate standards for the humane treatment of animals (U.S. Department of Health and Human Services, 1986). Moreover, not everyone agrees that animal experimentation can be ethical.
Given the ethical issues which surround animal experimentation, it is important to be aware that due care for the biostatistical aspects of experiments can contribute greatly to the humane conduct of research, by minimizing the exposure of animals to harm. Suppose that an experiment is planned which would incontestably settle a physiological point having great bearing on the relief of human suffering. Suppose further that there really were no scientifically valid substitutes for animal testing in that particular case, and that the animals were truly to be treated as well as possible given the experimental protocol. This may indeed be a lot to suppose. But would it necessarily follow that the experiment would be as ethical as possible? No, because an experiment, to be ethical, must be adequate to test the hypothesis of interest. In particular, the sample size must be neither wastefully large nor so small that important findings are likely to be missed.
Chimpanzees and HIV Vaccine Tests
The question of what constitutes an adequate sample size for an experiment has recently come to the fore because of the anticipated demand for the chimpanzee in research on infection with the human immunodeficiency virus (HIV). Such infection ultimately results in acquired immune deficiency syndrome (AIDS) in most, if not all, human patients. As Zarling et al. (1987) note, "Both chimpanzees and humans can be infected with HIV, and both subsequently develop antibodies to various HIV antigens; however, thus far, no HIV-infected chimpanzees have been reported to develop AIDS." It is possible, though, for infected chimps to develop at least "lymphadenopathy, transient depression of the ratio of T4 to T8 lymphocytes, and impaired blastogenic responses" (Alter et al., 1984).
Since the chimp can be infected with the virus and can summon up an antibody response to it, potential vaccines have been and will probably continue to be tested in that animal, as was the hepatitis B vaccine prior to its testing in human subjects. So far, all potential vaccine candidates tested have failed in the chimpanzee model. As Prince et al. (1988) point out, "Basic research is needed to explain these failures, otherwise a long string of abortive chimpanzee trials can be anticipated." It would indeed be unethical to carry out a long string of vaccine trials with little hope of success. Moreover, the chimp is a uniquely important creature whose numbers are severely limited: the U.S. Fish and Wildlife Service has announced that "it will upgrade the status of wild chimpanzees from threatened to endangered" (Ezzell, 1988), although even that may not be adequate protection. We must not squander a precious resource such as this animal, nor, for ethical reasons, can we undertake an inadequate experiment. If it has been decided that an experiment is justifiable and will be done, it must be done correctly.
The size of an experiment--that is, the number of animals involved--clearly is important. The use of more animals than absolutely necessary to test a given hypothesis or to delineate or demonstrate a particular phenomenon is obviously wrong because it may unnecessarily expose animals to risk or suffering. It is also a waste of resources which could be better spent. Less often recognized is the problem of too small a sample size. An experiment with too small a sample is unethical because it exposes animals to risk or suffering with an inadequate prospect of an informative outcome; they are placed at risk for no good reason. "Much primate research is based on minuscule sample sizes" (Gochfeld, 1988), and in some circumstances such work cannot be a reliable basis for statistical inference (Holland, 1988).
Example: Sample Sizes for Binomial Outcomes
How can we set suitable sample sizes for vaccine trials? There are many possible experimental designs which could be used in testing, and sample size calculation will vary according to how the experiment is performed. For the purposes of illustration, we will discuss the simplest case. This is called a binomial experiment, because the outcome for each individual experimental animal is binary and categorical: the animal ends up classified as either protected or not protected from infection.
Let us say that a worthwhile vaccine candidate will be administered to a group of chimpanzees and the animal will then be challenged by exposure to the virus. Each animal will then be assessed to determine whether the vaccine's immunogenicity was adequate to evoke neutralizing antibodies conferring protection against infection. The outcome for each animal, then, is whether it was or was not protected from infection by the virus, and we will assume that all cases in which infection was prevented were due to protection by the candidate vaccine.
The classic model of the binomial experiment is the tossing of a coin. The two possible outcomes, head or tail, are mutually exclusive and exhaustive. Each toss of the coin is independent of each other toss, in the sense that the result on one toss has no bearing on the result of any subsequent toss. For any long string of coin tosses, there is an underlying "true" probability of a given outcome, because of the properties of the coin. Individual outcomes cannot be predicted, so statisticians say outcomes are determined by a random process with a certain underlying probability. This does not mean necessarily that the outcomes could not be predicted if we were to examine in minute detail all the physical forces operating on the coin, just that we cannot predict outcomes for particular coin tosses or groups of tosses. Similarly, a vaccine may be effective in a certain percentage of cases, and this percentage may be a reproducible figure, even though we cannot predict with certainty the outcome for any given animal or experimental group.
Using the binomial theorem, the likelihood of a given set of observations can be calculated. For example, on the assumption that the true probability of getting a "head" is 0.5, we can estimate how often we will get any particular atypical sample, such as one consisting largely or exclusively of "heads". By analogous calculations, we can also obtain the probability of getting any specified number of successfully protected chimps in a vaccine trial, given any assumed underlying probability of success. These calculations help us estimate the sample size we need, because we can make various assumptions about the true protective effect of a vaccine, and then estimate the likelihood that we will get an atypical sample in trials of different sizes. Such likelihood estimates represent the chance that we will draw an erroneous conclusion about the vaccine from a specified trial, due to sampling fluctuation. As sample size increases, sampling fluctuation diminishes and the risk of an erroneous conclusion based on an atypical sample decreases.
Let us make some assumptions about vaccine efficacy and about sample size, and see how the risk of an erroneous conclusion changes as sample size changes. The probabilities we need for this exercise are given by the binomial formula. Suppose we want to know the probability of finding x successes in n subjects, and that the true underlying probability of successful vaccination is given by p. The chance that we will observe x successes is:
x n-x
n! p q
--------
x!(n-x)!
where q = 1-p and x! is read "x factorial". Note that x! = (x)(x-1)( x-2)...(1), and that both 0! and 1! are defined to be 1.
Let's compare the results we can expect from experiments using two versus five chimpanzees, assuming three different vaccine efficacies. Efficacy does in fact vary among the different types of vaccines on the market today. According to the Physicians' Desk Reference (Medical Economics Company, 1988), a single injection of a live mumps virus vaccine "has been shown to induce mumps neutralizing antibodies in approximately 97 percent of susceptible children." Studies of a virus-inactivated hepatitis B vaccine, cited in the same reference, showed that the product "induced protective levels of antibody in greater than 90% of healthy individuals who received the recommended 3 dose regimen." Virus-inactivated influenza vaccines are less effective (Arden et al., 1986), and certain attenuated live vaccines even less so. For example, in one clinical trial, a two-dose regimen of a particular bivalent live attenuated influenza vaccine was estimated to be 50 to 70% effective (Couch et al., 1986). For the purposes of demonstration, we will take 0.97, 0.90, and 0.50 as estimates of the probability of vaccine success.
Table 1 shows the probability (P) of the different results which can occur at each combination of sample size (n) and underlying vaccine effectiveness (p). Note that if a vaccine is 97% effective, you are extremely unlikely to have the vaccine fail in both members of a pair of experimental animals. The chance that you will see only one of them protected is between five and six percent. Most of the time both will be protected. With the sample size of five, almost all the time four or five of the five animals will be protected and you will rightly conclude that the vaccine is very effective.
n = 2, p = .97 n = 5, p = .97
X P X P
0 .0009 0 <.0001
1 .0582 1 <.0001
2 .9409 2 <.0003
TOTAL 1.0000 3 <.0082
4 <.1328
5 <.8587
TOTAL 1.0000
n = 2, p = .90 n = 5, p = .90
X P X P
0 .0100 0 <.0001
1 .1800 1 <.0005
2 .8100 2 <.0081
TOTAL 1.0000 3 <.0729
4 <.3281
5 <.5905
TOTAL 1.0000
n = 2, p = .50 n = 5, p = .50
X P X P
0 .2500 0 <.0313
1 .5000 1 <.1563
2 .2500 2 <.3125
TOTAL 1.0000 3 <.3125
4 <.1563
5 <.0313
TOTAL 1.0000
As the probability of success declines, the problem of sampling variability becomes more pronounced. When p = .90, and n = 2, you have an 18% chance of seeing only one success in your experimental pair. Finally, with a 50% effective vaccine and n = 2, you have a 25% chance of both animals being successfully protected, and also a 25% chance that neither will be protected. There is a high probability that an erroneous conclusion about the vaccine will be drawn from this small experiment. By contrast, the larger experiment with p = .50 and five subjects is much less likely to produce a result leading the experimenter to conclude that the vaccine is totally effective or totally ineffective. The larger the sample size, the more likely it is that the experimenter will see an accurate reflection of the vaccine's characteristic efficacy. This property of sampling variability is proven by statistical theorems, and is even more marked when the contrast is between sample sizes more disparate than two and five.
Discussion
Before an experiment is conducted, the relative reliability of conclusions from the proposed experiment should be examined. One can select various levels of vaccine efficacy and see how reliably these would be reflected in the results, given the sample size permitted by current resources. Alternatively, one might vary the sample size until an acceptably small risk of an erroneous conclusion is obtained, and set the sample size accordingly. Here we would need a specific hypothesis about the level of vaccine efficacy to start with. In some situations a specific estimate is justifiable on theoretical grounds or because only a very high degree of efficacy is worth detecting (i.e., it would not matter if we were erroneously to conclude that a vaccine were ineffective when in fact it did have some moderate protective effect).
Note that most of the information needed for the calculation of sample size must come from the experimenter. The experimenter must thoughtfully set in advance the level of vaccine efficacy that the experiment should be able to detect; the precision of efficacy estimates may also be set in advance. The experimenter must also accept that there is always some chance of an erroneous conclusion due to sampling variability in even the best-designed experiment, and must determine the risk of error which he or she considers tolerable under the circumstances. The sample size necessary can then be calculated, and no more animals than that number should be used.
Many experimental situations are far more complicated than those discussed here, and many details of an experiment need to be considered before the relevant sample size can be determined correctly. Many complicated study designs require quite involved calculations of sample size; some study designs are notably efficient, "squeezing" much information from small samples, while others are not (Cohen, 1988). It is advisable to consult a professional statistician before embarking upon any study, most especially research involving animals.
If animals are to be used at all in research, they must be used wisely. Once it has been decided that an experiment is to be done, it must be done in such a way that useful and valid information is obtained in exchange for the minimum possible exposure of animals to risk or suffering. The biostatistician can be a valuable ally of the experimenter by helping to maintain the most vigilant attention to the ethical aspects of animal experimentation.
References
Alter, H. J., Eichberg, J. W., Masur, H., Saxinger, W. C., Gallo, R., Macher, A. M., Lane, H. C., & Fauci, A. S. (1984). Transmission of HTLV-III infection from human plasma to chimpanzees: An animal model for AIDS. Science, 226, 549-552.
Arden, N. H., Patriarca, P. A., & Kendal, A. P. (1986). Experiences in the use and efficacy of inactivated influenza vaccine in nursing homes. In A. P. Kendal & P. A. Patriarca (Eds.), Options for the Control of Influenza (pp. 155-168). New York: Alan R. Liss.
Cohen, J. (1988). Statistical Power Analysis for the Behavioral Sciences (2nd ed.). Hillsdale, NJ: Lawrence Erlbaum.
Couch, R. B., Quarles, J. M., Cate, T. R., & Zahradnik, J. M. (1986). Clinical trials with live cold-reassortant influenza virus vaccines. In A. P. Kendal & P. A. Patriarca (Eds.), Options for the Control of Influenza (pp. 223-241). New York: Alan R. Liss.
Darwin, C. (1881). Letter to E. Ray Lankester. In Bartlett, J. (Ed.), Familiar Quotations (15th ed., p 515). Boston: Little, Brown, 1980.
Ezzell, C. (1988). Wild chimpanzees upgraded to endangered species. Nature, 336, 511.
Gochfeld, M. (1988). Research with primates. American Scientist, 76, 548.
Holland, B. (1988). Two chimps, too few. Nature, 334, 478.
Medical Economics Company (1988). Physicians' Desk Reference (42nd ed., pp. 1336, 1372). Oradell, NJ: Medical Economics.
Prince, A. M., Moor-Jankowski, J., Eichberg, J. W., Schellekens, H., Mauler, R. F., Girard, M., & Goodall, J. (1988). Chimpanzees and AIDS research. Nature, 333, 513.
United States Department of Health and Human Services (1986). Public Health Service Policy on Humane Care and Use of Laboratory Animals. Washington: U.S. Public Health Service.
Zarling, J. M., Eichberg, J. W., Moral, P. A., McClure, J., Sridhar, P., & Hu, S.-L. (1987). Proliferative and cytotoxic T cells to AIDS virus glycoproteins in chimpanzees immunized with a recombinant vaccinia virus expressing AIDS virus envelope glycoproteins. Journal of Immunology, 139, 990.
-------------------------------------------------------------------
Author's address: Division of Biostatistics and Epidemiology, Department of Preventive Medicine and Community Health (F-596), UMDNJ-New Jersey Medical School, Newark, NJ 07103-2757.
* * *
PSIC
The Primate Supply Information Clearinghouse writes that one of the most important resources for information sharing is the individual who has knowledge of a particular primate species, problem, or has contacts for further referrals. They would like to hear from those who would be willing to occasionally speak with another researcher about a specific problem or experiences with a species or colony. In this way, more progress can be made toward their (and all of our) goals of decreasing importation of nonhuman primates, improving domestic breeding and research programs, and improving the quality of life of those research animals that must be used. Contact Cathy A. Johnson-Delaney, Primate Information Center, Regional Primate Research Center, SJ-50, University of Washington, Seattle, WA 98195 [206-543-5178].
WRPRC FAX Number
The Washington Regional Primate Research Center, which includes the Primate Supply Information Clearinghouse and the Primate Information Center, now can be reached by FAX at 206-545-0305.
MMWR
Morbidity and Mortality Weekly Report, a publication of the Centers for Disease Control, announces a new phone number for their Production Offices. The number, 404-332-4555, provides information on subscriptions, published or submitted articles, HIV-related articles, statistics, supplements, surveillance summaries, summaries of notifiable diseases, and their annual index. .cb;PIC
The Primate Information Center has published its 1989 Fee Schedule and list of topical bibliographies in print. For a copy, contact the Primate Information Center, Regional Primate Research Center (SJ-50), University of Washington, Seattle, WA 98195 [206-543-4376].
IATA Publishes Regulations
The International Air Transport Association (IATA) has published the 14th edition of its "Live Animals Regulation," which entered into force on July 1, 1988. The publication is available in English, French, or Spanish for $40 [SFr. 70]. Order from IATA, 26 chemin de Joinville, P.O. Box 160, CH-1216, Cointrin, Switzerland; or IATA, 2000 Peel Street, Montreal, PQ H3A 2R4, Canada.
A.P.E. Project
The A.P.E. (Appropriate Primate Enrichment) Project is preparing an annotated bibliography of research findings on captive primate enrichment, to be ready soon. For a copy, contact Sarah T. Boysen, A.P.E. Project Coordinator, Dept. of Psychology, Ohio State Univ., Columbus, OH 43210-1222 [614-292-6512].
* * *
Scott W. Line, Kathleen N. Morgan, Hal Markowitz, and Sharon Strong
California Primate Research Center and San Francisco State University
Much of the research on psychological well-being for nonhuman primates has centered on ways to increase the behavioral opportunities for primates in a laboratory environment. An additional way to enhance their well-being is to seek husbandry methods that minimize pain and distress. Current regulations require that animals undergoing painful procedures be given treatments that relieve pain unless such treatments interfere with the research. The guidelines for relieving distress are more ambiguous, reflecting the lack of agreement among scientists about how to define stress.
Some of the normal daily human activities associated with keeping primates are likely to be stressful to the animals. Some activities such as changing cages create high levels of noise. Collection of blood samples generally requires physical restraint and the transient pain of venipuncture, and may be otherwise aversive. The disorientation and loss of control caused by anesthesia may itself lead to fear and distress.
The purposes of this project are twofold: 1) to establish normative values of heart rate and activity for singly caged rhesus monkeys (Macaca mulatta) , and 2) to document changes in these parameters as a function of routine husbandry procedures such as feeding, cleaning, cage changing, and tuberculin (TB) testing. Once normative values are obtained, and the reactions of the animals to routine procedures are recorded, then events that are potentially stressful can be identified. Procedural changes can then be adopted that will help minimize the distress that the animals experience. This will also promote better research by reducing the number of potential extraneous variables.
This report presents preliminary data on the normal daily cycles of heart rate and activity of singly housed adult female rhesus monkeys. It also includes some of their reactions to people performing routine tasks.
Methods
The subjects were 6 adult female rhesus macaques, aged 4 years 11 months to 5 years 1 month at the beginning of the study. Each weighed between 4.8 and 5.5 kg. Two were born into 0.2-hectare outdoor multi-male, multi-female corrals at the California Primate Research Center (CPRC). They remained in their natal cages for 28 months, were transferred to single-male groups for 21 and 18 months respectively, and subsequently housed indoors in single cages for 9 and 13 months prior to the present experiment. The remaining 4 subjects were born in outdoor single-male groups at the Hazleton primate facility in Alice, TX. They were placed in peer groups at 6 months and brought to the CPRC at 21 months of age. After a 3-month quarantine in indoor single cages, they were housed in a 0.2-hectare corral for 20 months. They were again housed indoors in single cages for 13 months before this study.
During the investigation the monkeys were housed individually in squeeze-back stainless steel hanging cages measuring .61x.66x.81 m, in an indoor room containing approximately 50 rhesus monkeys. Visual and auditory contact occurred with other monkeys across a central aisle. The room was illuminated from 0600 to 1800 hours daily, and cleaned at 0900-1000 hours. The monkeys were fed commercial monkey chow at 0700-0800 and 1400-1500 hours. Water was continually available from automatic devices.
Heart rate and activity levels were monitored automatically by a biotelemetry system.&S'1 Each subject had a transmitter surgically implanted subcutaneously along the dorsal midline. Each transmitter broadcast a radio signal encoding the electrocardiogram to a receiver mounted on the top of the subject's cage. Variations in the signal strength occurred as the monkey moved and were translated into a relative index of activity by the receiver. A computer in an adjacent room recorded the signals from each receiver at 2-minute intervals. Telemetry data were collected from 1000-1700 hours four days per week, for 24 hours continuously one day per week, and subsequently averaged at 1-hour intervals for each day.
Types of human activity in the room were identified by means of a panel mounted on the door to the animal room. The panel contained ten buttons listing different types of personnel and activities (animal technician feeding, animal technician cleaning, research technician, veterinarian, etc.). People were asked to press the appropriate button as they entered the room. A magnetic switch attached to the top of the door registered when they actually entered and when they left. Data from the door panel and the magnetic switch were also recorded at 2-minute intervals by the telemetry computer.
Results
Figure 1: Typical 24-hour patterns of heart rate and activity of six singly-caged adult female rhesus monkeys. Each point represents the mean (+/- SEM) of 30 samples taken during a one-hour period on 14 different days.
The mean heart rate and activity counts of 14 different 24-hour recordings of the six subjects over a three-month period are presented in Figure 1. Heart rate ranged from a low of 96 bpm at night to a high of 149 bpm at 0700. A second peak heart rate of 142 bpm was observed at 1400. The activity count varied from less than 2 counts/hr at night to peaks of 125 counts/hr at 0800 and 85 counts/hr at 1300. A one-way analysis of variance for heart rate during the hours that lights were on (0600-1800) showed a highly significant effect of time of day (p < .0001). A Friedman test of the variation in activity count during the same hours was also significant (p - .0001). Post-hoc paired comparisons of individual hours revealed that heart rate was significantly higher at 0700 and lower at 1800 than at any other hour, and that the hours from 1300 to 1500 were higher than the preceding or following hours (p < .05 in all cases). Activity was higher at 0800 than at any other hour (p = .0277, Wilcoxon test). The secondary activity peak at 1300 was higher than any of the hours from 1000 to 1700 (p = .0277, Wilcoxon test).
Figure 2: Heart rate of the subjects on a day the cages were changed and a normal day. The cage change took place between 0900 and 1100 hours. Each point represents the mean (+/- SEM) of 30 samples taken during a one-hour period for the six monkeys.
The heart rate response of the monkeys to having their cages changed is presented in Figure 2. This graph compares heart rate on a day that all the cages in the room were changed to heart rate on the following day. The procedure took place from 0900 to 1100 and involved placing the monkeys in transfer boxes while their dirty cages were replaced with clean ones. On the cage change day a peak heart rate of 175 bpm was recorded at 1000. The mean heart rate remained elevated for two hours after completion of the procedure. A 2-way analysis of variance resulted in a significant effect for both time of day and cage changing, and a significant interaction (p < .005 in all cases).
Figure 3: Heart rate of the subjects on a day they were TB tested and a normal day. The monkeys were still sedated with ketamine at 1000 and were fully awake by 1100.
The heart rate response of the monkeys to TB testing is shown in Figure 3. All the monkeys in the room were sedated with ketamine so that a technician could weigh them and administer the tuberculin. The peak heart rate of 192 was recorded at 1000 while the monkeys were still sedated. They appeared to be fully awake by 1100; nevertheless the mean heart rate remained elevated until 1400. Heart rate was significantly higher on the TB test day (2-way ANOVA, p < .025). There was also a significant effect of time of day, and a significant interaction (p < .005 in both cases).
Figure 4: Heart rate and activity during the morning health check. Each point represents the mean (+/- SEM) for the six subjects sampled at two-minute intervals on nine different days. The shaded bar denotes the period of time that a human was in the animal room.
Cage changing and tuberculin testing are relatively infrequent events. The animals also respond to some of the tasks that people perform on a daily basis with significant changes in heart rate or activity. For example, the first person to enter the room each day examines the animals for signs of disease or other health problems. Heart rate did not change while a technician was in the room performing the morning health check, but activity increased significantly (Figure 4, Friedman test, p = .0004). When a technician entered the room at night to administer antibiotics or other treatments to one of the other monkeys in the room, both heart rate and activity increased (Figure 5). The transient increase in heart rate was not significant (1-way ANOVA, p > .1), while the increase in activity was (Friedman test, p = .0042).
Figure 5: Heart rate and activity during an evening treatment. Each point represents the mean (+/- SEM) for the six subjects sampled at two-minute intervals on four different days. The shaded bar denotes the period of time that a human was in the animal room.
Discussion
Variations in heart rate and activity are closely associated with human activity in the room. Heart rate increases, though not significantly, when an animal technician enters the room in the morning for a routine health check, or in the evening to administer treatments. Activity increases significantly in both of these situations. However, these effects are of relatively short duration, with both heart rate and activity returning to basal levels after the human leaves the room. Both heart rate and activity increase rapidly when the lights come on in the morning. Peaks in heart rate occur just prior to the twice-daily feedings, and may be a reflection of the animals' anticipation of one of the major events in their day. These findings are consistent with a previous report of heart rate in freely moving, singly caged rhesus monkeys (Malinow et al., 1974). In that study, heart rate increased coincident with human activity in the animal room, reaching peaks during feeding and cleaning. In the present experiment the highest activity was recorded during the hour that the room was cleaned. Due to the increased noise and chance of being sprayed by water, cleaning is probably a mildly aversive event. Another activity peak happened at 1300, just prior to the second feeding. The level of human activity in the room is low after the afternoon feeding, and both heart rate and activity gradually decline from this point until the lights go off at 1800.
The persistent elevation in heart rate for several hours after cage changing and TB testing were completed was an unexpected finding. Though we anticipated that these might be aversive procedures, we assumed that the monkeys would have habituated to them through repeated exposure. We expected heart rate would return to basal levels soon after the event ended, as it does following its increase in response to the other routine events described above. One possible explanation is that events such as feeding and health checks are acute and of brief duration, while exposure to humans and to stimuli that may be aversive is greater and more prolonged in situations such as TB testing with ketamine sedation and cage changing. However, cage changes at this facility are often completed within an hour, and TB testing may be completed in less time than it takes to clean cages in the morning. Cage changing and TB testing are relatively rare events (taking place every other week, or every few months) as opposed to the daily occurrence of some of the other events mentioned. Though TB tests and cage changes happen repeatedly, the length of time between occurrences may preclude the monkeys' habituating to these events.
Animals continue to respond to daily routine experiences such as feeding and health checks, although these responses disappear rapidly following the event. The overall effect is a consistent and predictable daily pattern of heart rate and activity that is synchronized to these procedures. For other, less frequent events, heart rate and activity increase and remain elevated for several hours after the event's completion. These observations are important to note if one is to avoid confounding the physiological and behavioral effects of experimental manipulations with those induced by routine husbandry. Events that we tend to dismiss as potentially stress-inducing on the basis of their regularity may have more influence on our subjects than we suppose. Further research will help to identify the extent to which the effects of specific procedures need to be addressed, and how deleterious effects might be diminished.
Reference
Malinow, M. R., Hill, J. D., & Ochsner, A. J. (1974). Heart rate in caged rhesus monkeys (Macaca mulatta). Laboratory Animal Science, 24, 537-540.
-------------------------------------------------------------------
First author's address: California Primate Research
Center, University of California, Davis, CA 95616.
This work was supported in part by NIH grant RR00169 to the
California Primate Research Center. The authors thank Warren Miller of
Willoughby Electronics for design and construction of the door monitor
panel.
(1) Mini-Mitter Corp., Sun River, OR.
* * *
The Membership Committee of the American Association of Physical Anthropologists (AAPA) has issued an invitation to all primatologists to consider joining their organization. Membership includes a regular subscription to the American Journal of Physical Anthropology, a large portion of which is devoted to articles dealing with the biology of living and fossil nonhuman primates. Beginning in April, 1989, the editor of AJPA will be the primatologist Matt Cartmill of Duke University. Formerly the editor of the International Journal of Primatology, he will be guiding the AJPA for the next six years. Also included in membership is the Yearbook of Physical Anthropology as well as reduced registration fees at the annual meetings of the Association each spring. For more information, contact the Chairman of the Membership Committee, John G. Fleagle, at the Department of Anatomical Sciences, School of Medicine, Health Sciences Center, SUNY, Stony Brook, NY 11794-8081.
* * *
Ronald D. Nadler and Jeremy F. Dahl
Yerkes Regional Primate Research Center
The purpose of this article is to clarify the rationale of the research project, "Reproductive behavior and physiology of the gibbon," currently being conducted at the Yerkes Regional Primate Research Center of Emory University. One does not usually write about one's research before a significant amount of data are in hand, but in this case, one must refute a campaign of misinformation, distortion, and ridicule which has been directed against the study by animal welfare extremists.
The research, which was described derisively as "project penis" by one extremist group (Anon., 1988a), was also a focus of a recent protest held at the Yerkes Center by another such group. The accusation: "They're measuring penises at Yerkes." The clear implication of the accusation is that research funds are being spent on some perverse and perverted activity that could have no possible value to society.
The research is, in fact, a study in reproductive biology, designed to address several related hypotheses regarding the reproductive behavior, physiology, and anatomy of the gibbon, derived from our earlier research on the great apes (Nadler, Herndon, & Wallis, 1986; Dahl, 1987). The hypotheses to be tested concern 1) the sex of the animal which initiates sexual interactions, 2) the degree to which a male courtship display is exhibited, 3) the size and visibility of the penis, 4) the duration of estrus, 5) the testis weight-body weight ratio, 6) the frequency and pattern of copulation during the menstrual cycle a) when the male and the female have free access to each other, b) when primarily the female determines whether and when copulation may occur, and c) when primarily the male determines whether and when copulation may occur, 9) the pattern of female ano-genital swelling during the menstrual cycle, 10) the pattern of female sex hormone levels during the menstrual cycle, and 11) the pattern of male sex hormone levels during the cycle of the female with which the male is being tested. Particular interest is directed toward the relationships among several of the aforementioned variables, especially the relationships between hormone levels and ano-genital swelling of the female, hormone levels and sexual behavior, ano-genital swelling and ovulation, and sexual behavior and ovulation. The behavioral relationships, moreover, are of particular interest with respect to the varied test conditions employed, i.e., conditions that favor primarily male or primarily female regulation of sexual interactions, and as they relate to the concepts of attractivity, proceptivity, and receptivity (Beach, 1976). This reasonably complex study on a rare species, especially interesting from the comparative perspective, in a traditional area of research with wide and varied implications for captive breeding as well as basic science issues, was summarily dismissed by extremists as "measuring gibbons' penises" (Anon., 1988b). We submit that the measurement of genitalia does not differ from the scientific measurement of any other body structure in relation to the functioning of an organism and that such measurement is, in any event, a relatively small portion of the present research on gibbons.
Clearly this area of research on the reproductive biology of primates has a long history which cannot be described in this article. It is relatively straightforward, however, to place into perspective the measurement of penises, for those who resist simplistic distortions and who acknowledge that such a procedure could have scientific relevance. Several years ago, the eminent reproductive biologist, R. V. Short, noted that the difference in size and visibility of the penis in common chimpanzee and gorilla appears to represent an example of "form reflects function" (Short, 1981). The penis of the chimpanzee is relatively long and pink when erect, making it very conspicuous against a background of the chimpanzee's black hair. The gorilla, by contrast, although considerably larger in body size than the chimpanzee, has a much smaller penis which is black in color and, hence, quite inconspicuous. Short (1981) proposed that this morphological difference seems to reflect the function of the penis in courtship. Whereas the chimpanzee has an elaborate male courtship display in which the erect penis is a prominent feature, the male gorilla shows little, if any, courtship behavior and, in fact, females primarily initiate copulation. A. H. Harcourt (1981) noted that these differences in behavior, as well as several other differences among the great apes related to sex and reproduction, appear to reflect differences in the mating systems of the species; differences primarily in the number of males that compete for estrous females. The greater the intermale competition for estrous females, the more pronounced is male sexual initiative, including the performance of a courtship display. Among chimpanzees, with a multimale mating system, male sexual initiative and courtship behavior are prominent, whereas among gorillas, with a one-male mating system, male sexual initiative is minimal and male courtship essentially lacking. The gibbon, therefore, is an especially interesting species to study comparatively since 1) it is closely related to the great apes, 2) it has a one-male mating system similar to the gorilla, but 3) it differs from the gorilla in that it lives in monogamous pairs, in contrast to a polygynous, harem-type of social structure. The gibbon, in other words, is an ideal species for testing Short's and Harcourt's hypotheses regarding the variables that influence several parameters of reproductive function. One of the questions, therefore, is: Do these hypotheses accurately characterize the gibbon or, by implication, does social structure (monogamy vs polygyny) have a prepotent influence on the reproductive parameters? It is clear that the male gibbon has a relatively short, dark penis. The measurements being made, which are only a small portion of the actual research being conducted, represent the documentation required to confirm that fact (measurements are made under anesthesia associated with routine health examinations).
It is noteworthy that there has been no mention by the animal welfare extremists of the five especially designed gibbon enclosures that were built specifically to facilitate this research. The enclosures are located in 1.3 ha of forest adjacent to the Yerkes Center, positioned such that each is at least 50 m from its nearest neighbor, the median boundary size between territories of gibbon groups in the wild (Ellefson, 1974). The enclosures include indoor/outdoor compartments with a total volume of 130 cubic meters each, providing the animals with a spacious and thermally diverse environment. The enclosures were built in accordance with appropriate technology to minimize the costs of construction and maintenance (see Dahl, 1982; in press). In addition, they contain bars and perches specifically positioned and spaced to permit maximum opportunities for brachiation and comfortable rest, consistent with the morphological specializations, size, and predominant positional behavior of the gibbon (sensu Prost, 1965).
Extremist animal welfare organizations are waging a war of intimidation, obstruction, and even terrorism against animal researchers and research institutions, unparalleled historically. Their tactics of insinuation and intimidation require serious, effective response. They will not go away until they are exposed to the public for the dishonest, distorted, and anti-intellectual positions they represent. Researchers, through the news media and other public forums, must speak out in defense of themselves, their science, and the role of science in the enlightenment and intellectual progress of society.
In Victorian times, and for some time afterwards, it may have been understandable for people to exclude the word "penis" from "polite conversation". It has never been, and it certainly is not today, understandable or acceptable to exclude the penis from scientific investigation. To imply that measuring the penis in a scientific study is somehow inappropriate, perverse, or silly must be exposed as a socially reactionary position that cannot be tolerated in an enlightened society. Science is not attractive to people whose intellectual interests and curiosity are limited and for whom knowledge is a threat rather than a delight. It would be utter nonsense to allow the antiquated intellectual and sexual conservatism of an outspoken few to determine the appropriate topics and subject matter for scientific investigation.
References
Anon. (1988a). Yerkes researchers embark on "project penis". IPPL Newsletter, 15[3], 21.
Anon. (1988b). Yerkes director denounces research critics. IPPL Newsletter, 15[3], 21.
Beach, F. A. (1976). Sexual attractivity, proceptivity, and receptivity in female mammals. Hormones and Behavior, 7, 105-138.
Dahl, J. F. (1982). The feasibility of improving the captive environments of the Pongidae. American Journal of Primatology, Suppl. 1, 77-85.
Dahl, J. F. (1987). Sexual initiation in a captive group of pygmy chimpanzees (Pan paniscus). Primate Report, 16, 43-53.
Dahl, J. F. (In Press). An inexpensive, climate-controlled enclosure for gibbons utilizing appropriate technology. In E. F. Segal (Ed.), Psychological Wellbeing of Captive Primates. Park Ridge, NJ: Noyes.
Ellefson, J. O. (1974). A natural history of white-handed gibbons in the Malayan peninsula. In D. M. Rumbaugh (Ed.), Gibbon and Siamang, Vol. 3 (pp. 1-136). Basel: Karger.
Harcourt, A. H. (1981). Inter-male competition and the reproductive behavior of the great apes. In C. E. Graham (Ed.), Reproductive Biology of the Great Apes: Comparative and Biomedical Perspectives (pp. 301-318). New York: Academic Press.
Nadler, R. D., Herndon, J. G., & Wallis, J. (1986). Adult sexual behavior: hormones and reproduction. In G. Mitchell & J. Erwin (Vol. Eds.), Vol. 2A (Behavior, Conservation, and Ecology), J. Erwin (Series Ed.), Comparative Primate Biology (pp. 363-407). New York: Alan R. Liss.
Prost, J. H. (1965). Definitional system for the classification of primate locomotion. American Anthropologist, 67, 1198-1214.
Short, R. V. (1981). Sexual selection in man and the great apes. In C. E. Graham (Ed.), Reproductive Biology of the Great Apes: Comparative and Biomedical Perspectives (pp. 319-314). New York: Academic Press.
-------------------------------------------------------------------
Authors' address: Yerkes Regional Primate Research
Center, Emory University, Atlanta, GA 30322.
Preparation of this article was supported in part by NIH Grant
RR-00165 from the Division of Research Resources to the Yerkes Regional
Primate Research Center and by NSF Grant BNS 87-08406 to RDN. The Yerkes
Center is fully accredited by the American Association for Accreditation
of Laboratory Animal Care.
* * *
Proposed regulations implementing the 1985 amendments to the Animal Welfare Act were published in the Federal Register (54, 10822-10954) on March 15.
For nonhuman primates, the following requirements are notable: In order of preference, nonhuman primates must be housed in primary enclosures with compatible members of their own or other nonhuman primate species, in pairs, family groups, or other compatible social groupings; if this is not possible, individually housed animals must be able to see and hear nonhuman primates of their own or compatible species; and if even that is not possible, the individually housed animals must have positive physical contact or other interaction with their keeper or other familiar and knowledgeable person for at least one hour each day. The minimum space that must be provided in research facilities for each animal is determined by the typical weight of its species, except for brachiating species, according to the following table:
GROUP Weight Floor Area Height
/Animal
kg sq-m cm
1 (e.g., tamarins) 1 0.15 50.8
2 (e.g., capuchins) 1-3 0.28 76.2
3 (e.g., macaques) 3-10 0.40 76.2
4 (e.g., large African species) 10-15 0.56 81.28
5 (e.g., baboons) 15-25 0.74 91.44
6 (smaller apes and brachiating species) 25-40 2.33 213.36
7 (larger great apes) *new group >40 4.65 213.36
To allow time for rebuilding or remodeling to meet these requirements, variances of up to 2 years, with a possible extension of 1 more year, could be granted. Impervious surfaces are no longer required, but materials that cannot be adequately sanitized must be removed or replaced as necessary. Spot cleaning is permitted for scent-marking species. In order to promote the psychological well-being of nonhuman primates in research facilities, the physical environment in primary enclosures must be enriched by providing means of expressing species-typical activities, such as perches, swings, mirrors, and toys or objects to manipulate; and using foraging or task-oriented feeding methods. Individually housed animals must be released for at least 4 hours of exercise and social interaction per week into an area at least 3 times the area and twice the height required for that species (see table above). Additional environmental enrichment, exercise, and social interaction may be necessary, in accordance with the instructions of the attending veterinarian, for infants and juveniles, animals used in research which does not provide enough activity, and those that show signs of being in psychological distress. Documentation of such release of each animal, and of environmental enrichment, must be kept by the attending veterinarian and is subject to inspection by APHIS and, in the case of federal research facilities, any federal funding agency. The diet for nonhuman primates must consist of varied food items, and the method of feeding must be varied daily (e.g., task-oriented feeding or foraging). All animal research areas, as well as housing, will be subject to inspection and compliance with these regulations.
Requirements for dealers and exhibitors differ from those for research facilities. In particular, dealers must keep individually housed nonhuman primates in cages with twice the minimum floor area and twice the minimum height required for research facilities.
All persons who will be affected by these regulations should read them in their entirety, and consider responding with comments on their probable impact. Written comments are requested until May 15, 1989. Send an original and 3 copies to Helene R. Wright, Chief, Regulatory Analysis & Development Staff, PPD, APHIS, USDA, Room 1000, Federal Bldg., 6505 Belcrest Rd., Hyattsville, MD 20782, referring to Docket No. 88-013. It is suggested that copies of comments be sent to the National Association for Biomedical Research (NABR, 818 Connecticut Ave., Suite 303, Washington, DC 20006) and the American Society of Primatologists (c/o Nancy Caine, Dept. of Psychology, Bucknell Univ., Lewisburg, PA 17837), so that those organizations can analyze their content and develop comprehensive comments. Readers who belong to other involved organizations will want to send copies to them, as well.
Parts 1 (definitions) and 2 (administrative matters) were originally published March 31, 1987. Revisions of those Parts are still in proposal form, and the comment period on them will close May 15. According to the APHIS public notice accompanying the current revision, republication is "for the sole purpose of soliciting comments on the narrow issue of the interrelationship of the definitions and regulations in Parts 1 and 2 with the standards we are proposing in Part 3...The public is therefore invited to comment exclusively on this issue. We will not consider comments going beyond this issue." NABR "believes all significant provisions of Parts 1 and 2 are closely connected to the Part 3 Standards. Beyond this, [they] believe that many important features of the revised regulations are completely new. The public has never had the opportunity to comment upon the specifics of these new regulatory ideas being seen for the first time." A portion of the NABR Conference, April 30 to May 2 (see p.14 of the January, 1989, issue of this Newsletter), will be devoted to finalizing the Association's official submission on Parts 1 and 2.
* * *
Ruling on Silver Spring Monkeys
The euthanasia of some of the Silver Spring monkeys has been delayed by a temporary restraining order. Three monkeys were to be euthanized for humane reasons at Delta Regional Primate Center based on recommendations of a "blue ribbon" panel of animal care experts who determined the animals were suffering and in danger of serious life-threatening injuries due to their deteriorating health. A Louisiana SPCA committee also recommended the animals' euthanasia almost two years ago. A hearing has been scheduled for April 12 in federal court in Louisiana.
The suit for the restraining order was filed by numerous animal rights groups, including People for the Ethical Treatment of Animals (PETA). The activists contend that the animals' condition does not warrant euthanasia and that they could live "safely, humanely, and comfortably if transferred to a suitable facility." The New England Anti-Vivisection Society and PETA ran identical ads in The New York Times on December 26 and The Washington Post on December 27, respectively, requesting President Reagan to "save" the Silver Spring monkeys. On January 3, PETA ran the same ad in The Washington Times, but as a full page and told "concerned citizens to call the White House," providing the direct phone number for the office of the Chief of Staff.-- From the NABR Update, 1989, 10 [1].
Immuno Lawsuit Dismissed
The Appellate Division of the Supreme Court of New York has dismissed a lawsuit filed by the Austrian corporation Immuno A.G. against Dr. Jan Moor-Jankowski and others in 1984. The lawsuit followed publication in the Journal of Medical Primatology of a letter from Shirley McGreal, Chairwoman of the International Primate Protection League. In her letter, Dr. McGreal raised questions about Immuno's plans to set up a laboratory in Sierra Leone that would use wild-caught chimpanzees.
The judgement stated that "Without exception, the statements at issue were either opinion absolutely privileged under the First Amendment, or statements which the plaintiff utterly failed to show susceptible of being proved false. Indeed, most of the factual statements claimed by the plaintiff to be defamatory were, on the record before us, demonstrably true." The court also ruled that, "The plans of a party to undertake a course of activity which would possibly frustrate efforts of the international community to safeguard an endangered species, are of public concern."
The 5-judge panel deplored the delays in dismissing Immuno's complaint, commenting that, "To unnecessarily delay the disposition of a libel action is not only to countenance waste and inefficiency but to enhance the value of such actions as instruments of harassment and coercion inimical to the exercise of First Amendment rights." In its decision, announced on 17 January 1989, the court held unanimously (5-0) that Immuno's lawsuit should be dismissed, with costs.
Yellow Fever Virus Activity in Trinidad
In December 1988, the Ministry of Health, Trinidad and Tobago, and the Caribbean Epidemiology Center received reports of howler monkeys (Alouatta sp.) dying in the Trinity Hills area of the southeastern forests of Trinidad. Surveillance of monkeys in forested areas is a component of the Ministry's ongoing surveillance of sylvatic yellow fever. Field and laboratory investigations did not establish specific evidence of yellow fever in either of two dead monkeys, one decomposing and one freshly shot. However, mosquito surveillance using human landing collections (mosquitoes collected as they landed on humans) yielded two isolates of yellow fever virus from 29 pools of Haemagogus janthinomys collected in the forests between December 8 and January 19. Three additional yellow fever isolates were detected in the same pools.
Surveillance for sick or dying monkeys and of forest mosquitoes was intensified after dying monkeys were reported. Island-wide immunization has been undertaken. In Trinidad and Tobago, although intensified fever surveillance for human cases was implemented when reports of dying monkeys were confirmed, no human cases have been detected. As of February 1, 1989, no yellow fever virus has been isolated from monkeys. A program of local spraying of adulticides, identification and elimination of peridomestic breeding sites for Aedes aegypti, and application of larvicides has been initiated in villages near the forest edge. Although virus activity remains confined to one forested area, the Ministry of Health has made a public statement advising visitors to forested areas to be immunized.
No evidence of virus activity in the island of Tobago exists, and the port areas and airports of both islands are free of Ae. aegypti. Yellow fever virus has never been isolated from Ae. aegypti or mosquitoes in Tobago, and the monkeys and mosquitoes associated with sylvatic yellow fever are not found there. --[From Morbidity and Mortality Weekly Reports, 1989, 38 [4], 57-59.]
* * *
Maria L. Boccia
University of Colorado Health Sciences Center
Introduction
There are significant advantages in using social companions rather than inanimate objects to provide environmental enrichment and enhance the psychological well-being of primates. For example, the likelihood of boredom or habituation is greatly lessened, and the variety of stimulation is substantial, with an ever-changing, although predictable social partner. Recognizing this, laboratories have increasingly begun to use social companions for enrichment purposes (e.g., O'Neall, 1988; Reinhardt, Eisele, & Houser, 1988). Provision of social companions, however, can introduce several potential hazards. Of special concern is the possibility of aggression due to competition for food, water, or preferred perch sites, with the attendant risk of injury. We recently reported a study which demonstrated a reduction in aggression and stereotypies by providing a natural foraging task in a group of socially housed pigtail monkeys (Boccia, 1989). That study was, however, limited to two weeks following the introduction of the foraging task. It was possible that the animals would habituate to the foraging task and show a return to baseline levels of aggression and stereotypies. The study presented here examines the behavior of this group of monkeys two months after the initiation of the foraging task, to determine if this occurred.
Methods
A group of socially housed pigtail monkeys (Macaca nemistrina) , consisting of an adult male, 7 adult females, and 7 juveniles and infants, was studied. Water was freely available and food was provided at 0900 hr. by scattering monkey biscuits and fresh fruit across the floor of the living pen. The pen was 2.1x2.5x4 m., with cinderblock walls, wire mesh ceiling, and woodchip bedding on the cement floor. Additional space was provided with shelving and pipes.
A supplementary feeding of approximately one cup of sunflower seeds were dispersed throughout the cage in the woodchip bedding in the middle of the afternoons, 4-6 hours after the group was fed their daily ration of chow and fruit. Animals were thus given the opportunity to forage, searching through the bedding to find the seeds. Behavioral data were collected before introducing this task, immediately after beginning this regimen, and 2 months later. Behavioral observations consisted of 5 minute focal animal samples from 5 females 9-10 year old, 3 females 4-5 years old, and one juvenile 1.5 years old. These data, including social and exploratory behaviors, were subjected to Analyses of Variance with condition and age as factors. The conditions were baseline, first test, and 2-month follow-up. Between the first and second phases, two of the three juveniles in the original study were culled for another project. Analyses were, therefore, limited to the adults and subadults.
Results
Two months following the initial introduction of the foraging task, social, exploratory, and abnormal behaviors continued to show the changes that were seen immediately after introduction of the task. Stereotypies remained depressed p = .027), and hairpulling remained rare. In addition, bedding p = .022) and other types of exploration (p = .0007) remained elevated, and agonistic behaviors remained low (fight: p = .031) (see Figure 1). After an initial increase only one behavior returned to baseline levels: proximity to others: (p = .01).
Figure 1: Behavioral responses of all monkeys observed in response to the introduction of sunflower seeds (OE = explore objects; BE = explore bedding; SM = stereotypic motor behavior).
Several behaviors initially were differentially affected, depending on the age class of the monkeys (including play, locomotion, bedding explore, and affiliative behavior; see Boccia, 1989). By the two month follow-up observations, the juveniles had been removed from the group and most of these differences, which were primarily due to these juveniles, were no longer seen. Only social grooming exhibited an Age x Phase interaction: adults showed an immediate decline in grooming and this remained lower at the two month followup, while the subadults exhibited an initial increase, followed by a return to baseline levels (see Figure 2). The single remaining juvenile continued to show the higher levels of play behavior.
Figure 2: Differences in social grooming of adults and subadults to introduction of sunflower seeds.
Discussion
Given the trend toward social housing as an enrichment strategy for laboratory primates, it is imperative to discover techniques which can be implemented to minimize aggressive behavior, and the attendant risk of physical injury. The presence of a food which allows a natural foraging activity reduced agonistic behavior and stereotypies, including hairpulling behavior, for the two months of the study, in this group of macaques. It thus seems possible to employ such a strategy on a relatively long term basis without a loss of effectiveness. The initial study demonstrated that these effects were somewhat affected by the age class of the monkeys. The current study, due to the loss of the juvenile subjects, was unable to determine if the age effects persisted.
These results suggest that habituation is not a problem with this foraging task. Therefore, it seems possible to use such a task for enrichment purposes in socially housed primates, to reduce levels of stereotypies and agonistic interactions on a long term basis.
References
Boccia, M. L. (1989). Preliminary report on the use of a natural foraging task to reduce aggression and stereotypies in socially housed pigtail macaques. Laboratory Primate Newsletter, 28[1], 3-4.
O'Neill, P. (1988). Developing effective social and environment enrichment strategies for macaques in captive groups. Lab Animal, 17[4], 23-36.
Reinhardt, V., Eisele, S., & Houser, D. (1988). Environmental enrichment program for caged macaques: A review. Laboratory Primate Newsletter, 27[2], 5-6.
-------------------------------------------------------------------
Author's address: Department of Psychiatry, C-268R,
Univ. of Colorado Health Sciences Center, 4200 East Ninth Ave., Denver,
CO 80262.
This research supported by USPHS grant MH 44131.
* * *
D. S. B. Watson, B. J. Houston, and G. E. Macallum
Parke-Davis Research Institute
Concern for providing a wider range of activity for individually housed cynomolgus monkeys (Macaca fascicularis) has led this laboratory to search for items which provide the monkeys an opportunity for manipulation of objects which are durable, nontoxic, relatively inexpensive, and can be cleaned through normal cagewash procedures. We have investigated a variety of objects and toys, and have discovered that all monkeys do not like all toys, and that what interests one monkey today may hold no interest for him next week. The best approach to this problem appears to be to provide a variety of objects on a rotating basis, so interest is maintained, and new articles are available for handling (Wallace, 1988).
Hard nylon balls have been used here with moderate success (Renquist & Judge, 1985). Although some of our animals ignore these balls, others have been occupied for hours rolling them down the cage bars, holding them, spinning them in their food bins, or chewing on them.
We have had success with two stainless steel objects--a tray (7 x 9-3/4 x 1/2 in.) and a bowl (6 in. in diameter and 2 in. high). These items may be purchased at stores selling restaurant supplies, and are sturdy enough for heavy usage. Both of these objects provide monkeys with something to carry, to place food biscuits in, and to reflect back images. Monkeys with trays will often manipulate them in and out of the front cage bars. A less desirable feature is the noise generated by animals sliding or pounding the stainless steel across the cage walls or floor.
We have also found that some baby toys are well suited for monkey manipulation. Fisher-PriceR vinyl teething rings, "Ring of Beads", with four attached colored smaller rings of varying sizes and colors, is a favorite of most monkeys. This object provides the animals with something to chew on, as well as rings which can be maneuvered around and through each other. Extended use does lead to wear from teeth scratches, but they are remarkably durable and can be replaced at a low cost.
Another toddler toy which the monkeys have enjoyed has been the PlaywellTM Snuggly Duckling Mirror. This toy provides 2 handles for carrying, 3 plastic mirrors set at slightly different angles, and a duck face which rotates while making clicking noises, thus providing several different ways to occupy a monkey's attention.
We have tried several children's toys which have not proven to be "monkey proof." In general, objects to be avoided are those made of relatively thin or brittle plastic, those which have seams which can be pried apart by monkey teeth, or objects of clear plastic which contain items to be viewed through them. The latter are likely to be defaced by teeth marks or demolished by monkeys trying to get at what is inside.
Our search for toys has been limited to those which will not interfere with the use of the squeeze back cage. We continue to look for more variety in usable toys, as variety appears to be an important factor in the interest a monkey shows in an item. We have not done an analysis of the activity of our singly caged monkeys to determine just what changes these objects have made to their behavior, but it is subjectively evident that providing these playthings permits monkeys to occupy themselves with the manipulation of varying shapes and textures, providing increased environmental complexity. We feel that these toys are useful in enriching the cage environment of individually housed cynomolgus monkeys and have proven to be durable, nontoxic, inexpensive, and easily sanitized.
References
Renquist, D. M. & Judge, F. J. (1985). Use of nylon balls as behavioral modifier for caged primates. Laboratory Primate Newsletter, 24[4], 4.
Wallace, G. (1988). Improving life for primates. Caring for Animals, 5[1], 3.
-------------------------------------------------------------------
Authors' address: Parke-Davis Research Institute, Division of Warner-Lambert Canada Inc., 2270 Speakman Drive, Mississauga, Ontario, Canada L5K 1B4.
* * *
Carolyn Crockett, Joe Bielitzki, Ann Carey and Alex Velez
Regional Primate Research Center, University of Washington
Introduction
The Animal Welfare Act of 1985 mandated standards to promote the psychological well-being of laboratory primates. One approach to improving well-being has been to provide manipulable objects. In this study we investigated the responses of macaques housed in single cages to Kong toys (The Kong Company, Lakewood, CO 80226). The Kong is an autoclavable natural rubber toy that comes in a variety of sizes and colors. It resembles three different-sized donuts stacked one on top of the other with a bump at the small end. The large end has an opening to a hollow interior. Our study investigated short-term and long-term responses to a 3-1/2" x 2-1/8" Kong, whether it was more attractive when filled with an edible substance, and whether it produced any changes in abnormal behavior.
Subjects and Methods
The subjects were 22 macaques: 3 male and 8 female adult Macaca nemestrina (Mn), 4 adult female, 2 adult male, and 4 juvenile male M. fascicularis (Mf), and 1 adult female M. mulatta. Some monkeys had prior experience with the empty Kong, but none had frozen-juice-filled Kongs previously. The M. mulatta and six female Mn were used in pilot observations. The remaining 15 subjects were observed on four days: Day 0: No Kong ("Control," 1 hr); Day 1: First presentation of empty Kong (1 hr); Day 2: Next day, same empty Kong (20 min) followed by presentation of Kong filled with frozen apple juice and removal of empty one (1 hr); Day 3: Usually the next day, same Kong, now empty (1 hr). To measure long-term interest, 2 to 7 weeks later 8 subjects were observed for an additional hour to see if they continued to interact with the Kong originally introduced in the "Juice" condition.
At 1-min intervals the observer recorded one of twelve mutually exclusive categories of behavior including manipulating Kong and abnormal behavior. When the subject was interacting with the Kong, the specific type of behavior was also recorded on the minute. Wilcoxon tests compared percentages for the same subjects between different conditions, and Mann-Whitney U tests were used for all other comparisons. Two-tailed probabilities of 0.05 or less were considered significant.
Results
The response to the Kong was highly variable. The maximum percentage of time spent manipulating the Kong during a single observation session varied from 0% (an adult male Mf) to 100% (male and female Mn). The overall manipulation of the Kong was significantly higher among those subjects whose response to initial presentation of the Kong was to take it from the observer's hand or cage floor within 5 sec after presentation compared to those that ignored it (p = .004). Figure 1 shows the mean percent of time spent manipulating the Kong in the different conditions. The dashed line shows the mean for the 11 subjects that initially took the Kong and the solid line presents the data for the 4 that ignored it. Overall, there was a significant decline in interest from the first presentation to the next day. Presenting a fresh Kong filled with frozen juice resulted in a significant increase in manipulation, but this level also had declined by the following day. However, the percent of manipulation had not significantly declined further for the eight subjects retested several weeks later. There was no significant difference in the amount of manipulation of the empty Kong when first presented and its presentation when juice-filled.
Figure 1: Mean percentage of time spent manipulating Kong (empty in all but Juice condition; see text). Wilcoxon: * p < .01.
Ten subjects consumed the 1.25 oz. of frozen juice in less than an hour (mean = 46 min, range 34 to 53 min). Seven others had eaten part of it. Four consumed none or very little. When the toy was filled with juice, 27% of the manipulation involved holding the Kong while gnawing at the frozen contents and 23% involved digging into the object with a finger. When the Kong was empty, 44% of the interactions were gnawing or licking it, most often on the pointed end. Mean abnormal behavior during the Control condition was 2.3% compared with 2.9% during Day 3 (p = .35, not significant (NS)).
Other comparisons were: (1) Males vs. females: NS. (2) Captive vs. Wild-born (most of the captive-born animals had been primarily group-housed): NS. (3) Mn > Mf: trend, p = .053. (4) Mn > Mf: adults: p = .004. (5) Previous experience with Kong > none: p = .007.
Discussion
Most of the monkeys manipulated the Kong toy, and several were extremely interested in it. Only 1 of 22 subjects never touched the toy. Adult Mn interacted with the toy significantly more than did adult Mf but this result is confounded because the majority of Mn were captive-born with previous Kong experience whereas most Mf were wild-born with no experience. Although initial high interest waned, monkeys that showed considerable interest in the beginning continued to interact with the toy at a lower level over time. Some animals showed a renewed interest in the object when it was made wet by cage washing. Attempts to remove Kong toys from cages were often met with resistance by their "owners" (see also Bayne, 1989). Presenting the toy filled with frozen juice definitely increased its attractiveness to those animals that were interested in it at initial presentation. However, the percentage of time spent manipulating it was not significantly greater compared with presentation of a fresh empty Kong at the beginning of the study. One important conclusion from this study is that initial response to the object was a good predictor of subsequent manipulation.
Providing objects such as Kong toys to macaques in single-animal housing with little or no opportunity for manipulation is mildly enriching to some of the monkeys. Periodically removing and reintroducing the toys would increase their enrichment value. Immediately replacing them with Kongs filled with frozen juice or some other food item extends their attractiveness. Some animals fail to show any interest in the toy, and we found no evidence that Kongs affected the level of abnormal behavior. Compared to a nylon ball, the Kong provides opportunity for more complex manipulation and is not noisy. Its contents can be varied, and gnawing it makes the surface irregular and potentially more interesting.
Reference
Bayne, K. A. L. (1989). Nylon balls re-visited. Laboratory Primate Newsletter, 28(1), 5-6.
-------------------------------------------------------------------
Authors' address: Primate Center SJ-50, University of Washington, Seattle, WA 98195. This research was supported by NIH grants RR04515 and RR00166.
* * *
The Hunterian Institute of the Royal College of Surgeons of England has announced its schedule of courses for 1989. Those which may be of interest to readers of the Laboratory Primate Newsletter are "Named Vets Course" (May 31), "Veterinary Course" (June 20-2l), and "Laboratory Animals for Licensees and Research Workers" (October 4). Details are available from the Institute Office, The Hunterian Institute, Royal College of Surgeons of England, 35-43 Lincoln's Inn Fields, London WC2A 3PN, England.
* * *
M. Chimes, Abbott Labs D403, 1400 Sheridan Rd., N. Chicago, IL 60064.
John J. Ely, Dept. of Biostatistics, Human Genetics Div., Univ. of Pittsburgh, Pittsburgh, PA 15261.
Laura Gillespie, 46 West Hanover, Cape Girardeau, MO 63701.
Michael Kaplan, Childrens Hospital, Enders Building, 300 Longwood Ave., Boston, MA 02115.
Michale E. Keeling, Univ. Texas System Cancer Center, Environmental Science Park, Rt. 2, Box 151-B1, Bastrop, TX 78602.
Paul Kirk, Schering Corp., P.O. Box 32, Lafayette, NJ 07848.
Steven M. Kuhlman, Dept. Lab. Animal Medicine, TRW Hazleton Lab., 9200 Leesburg Tpk, Vienna, VA 22180.
Scott Line, California Primate Research Center, University of California, Davis, CA 95616.
Dan Lyons, Thomas Jefferson Univ., 644A JAH, 120 Locust St., Philadelphia, PA 19107.
LaVonne Meunier L-70, Smith Kline & French, P.O. Box 1539, King of Prussia, PA 19406-0939.
Susan Morgenstern, Yale Univ. School of Med., Section of Comparative Med., P.O. Box 3333, 333 Cedar St., New Haven, CT 06510. ,cb
Richard K. Nakamura, Room 11-105, Parklawn Bldg, 5600 Fishers Lane, Rockville, MD 20857.
Mike Parker, University of Iowa, Animal Care Unit, 400 Medical Laboratories, Iowa City, IA 52242.
Amy Morris-Smith, 1120 E. 38th Place, Tulsa, OK 74105.
Richard Nelson, Lab. Animal Resource Center, Washington State Univ., Pullman, WA 99164-1165.
Arthur W. Rowe, New York Univ. Medical Center, LEMSIP, Long Meadow Rd., Tuxedo, NY 10987.
M. E. Stephens, Dept. of Anthropology, University of Calgary, 2500 University Drive NW, Calgary, Alta. T2N 1N4 Canada.
David A. Valerio, Hazleton Res. Products, P.O. Box 7200, Denver, PA 17517.
Janet D. Thomas, 870 East El Camino Real #131, Mountain View, CA 94040.
Pat Ward, American Cyanamid, 56B/236, Middletown Rd., Pearl River, NY 10965.
Vern Weitzel, Dept. of Prehistory & Anthropology, Australian National University, Canberra, ACT 2601 Australia.
* * *
The National Zoological Park (Smithsonian Institution) is offering a Curatorial Internship, which will provide practical experience in the daily management of an exhibit collection of exotic animals (mammals, birds, reptiles, amphibians, and invertebrates), and experience in zoo administration, education, and conservation, as well as the opportunity to conduct original research in a zoo. Candidates must have a recent doctorate in life sciences; research or employment experience in zoology, environmental biology, or conservation and a career interest in conservation, zoo administration, or conservation education. The position will not exceed 2 years beginning 1 October 1989, at $18,000 per year. Send introductory letter, curriculum vitae, reprints, and three letters of reference by 30 June 1989 to: Elizabeth Franke Stevens, Curatorial Intern Search Coordinator, National Zoological Park, Washington, DC 20008.
* * *
Digit Fund
The Digit Fund is dedicated to continuing the program of research on mountain gorillas and their habitat begun by the late Dr. Dian Fossey at the Karisoke Research Center, Parc National des Volcans, Rwanda. The Center, founded by Dr. Fossey in 1967, served as a base for her research and conservation efforts on behalf of mountain gorillas for the next 18 years. The Digit Fund is now seeking to expand the scope of research and conservation activities at the Center to study not only of mountain gorillas, but also other rare and endangered animal species present in the Parc National des Volcans and the park's unique flora and ecology. The gorilla population in the study area has been monitored continuously since 1967 and a large body of background data is available on both groups and individuals.
The Karisoke Research Center can accommodate six researchers in rustic cabins. Electricity is available at the Center by generator only and laboratory facilities are limited. All researchers are expected to contribute $150/month to offset Center operating and maintenance expenses; other subsistence expenses can be expected to run a minimum of $200/month.
Each year The Digit Fund plans to appoint one to three Census and Research Interns, who will assist the Director of the Center in carrying out periodic censuses of the study population as well as in collecting daily demographic and ranging data on individual study groups. As time permits, Interns may also have the opportunity to assist resident researchers in ongoing behavioral or ecological studies of gorillas and other species or to carry out short-term studies of their own design. Census and Research Interns will be provided with a round-trip airfare to Rwanda and will also receive a small monthly subsistence allowance during their period of residence at Karisoke. This program is intended for individuals having completed at least two years of undergraduate study through recent graduates.
For further information on research opportunities, sponsorship of proposed projects, and internships, contact: The Digit Fund, c/o Morris Animal Foundation, 45 Inverness Drive East, Englewood, CO 80112-5480.
Earthwatch
Earthwatch is a non-profit organization which provides volunteers, funds, and recognition to scholars doing fieldwork in the sciences and humanities. Among the projects and scholars listed in their 1989 catalogue are studies of howler monkeys in Costa Rica, by Kenneth Glander and Margaret Clarke; rain forest ecology in Costa Rica, by Douglas Levey; paleontology in Zaire, by Noel Boaz; and orangutans in Borneo, by Biruté Galdikas. Volunteers learn to participate in fieldwork. They pay their own transportation expenses, plus a share of the costs of the expedition (all these costs are usually tax-deductible). Interested scientists should submit a two-page outline of the research subject and design, dates, budget, and plans for the use of volunteers to the Center for Field Research, 680 Mount Auburn Street, Box 403, Watertown, MA 02272. For more information, call 617-926-8200. Prospective volunteers should write to Earthwatch at the same address.
Wildlife Preservation Trust
The Wildlife Preservation Trust, an international non-profit organization, develops and maintains captive breeding programs worldwide, as well as conducting research, educational programs, and professional training. Their Training Program is a 10 or 16 week internship designed for postgraduates and zoo personnel to give them intensive training in propagation techniques with a variety of bird, mammal, and reptile species. The trainees work side-by-side with zoo staff on a daily basis. They also participate in weekly seminars on topics ranging from pathology to animal behavior to zoo design. For application and further information contact Training Program, Wildlife Preservation Trust International, 34th Street & Girard Ave., Philadelphia, PA 19104 [215-222-3636].
The Summer Course is designed for advanced undergraduates and graduate students. This year's course will run from July 22 to August 12. The deadline for applications is April 30. Further details from Mr. John McNicholas, Assistant Training Officer, Jersey Wildlife Preservation Trust, Les Augres Manor, Trinity, Jersey, Channel Islands, British Isles.
ASP Summer Internship List
The Education Committee of the American Society of Primatologists (ASP) has compiled a list of summer internships and assistantships for students interested in Primatology. For information about its preparation, or for copies, contact Reinhold J. Hutz, Chair of the Education Committee of ASP, Dept. of Biological Sciences, Univ. of Wisconsin, P.O. Box 413, Milwaukee, WI 53201.
School for Field Studies
The School for Field Studies offers 13-week semester programs in the fall and spring, and month-long summer and January courses. Northeastern University, Boston, accredits all SFS courses. This year's courses include semester and summer courses in Wildlife Ecology and Management in Kenya, Tropical Ecology in Costa Rica, and Social Ecology of Stumptail Macaques at an island colony in Mexico. For information contact the Admissions Department, The School for Field Studies, 16 Broadway, Beverly, MA 01915 [508-927-7777].
Madagascar Tour
Wildlife Preservation Trust International is sponsoring an educational tour of Madagascar, led by Alison Jolly. The 18-day tour departs October 21, 1989, and will cost approximately $5900. For information, contact WPTI, 34th Street & Girard Ave., Philadelphia, PA 19104.
University Research Expeditions
The University Research Expeditions Program of the University of California encourages public participation in and helps fund university research in many disciplines throughout the world. Participants are invited to "share the challenges and rewards of field research as an active member of a University of California expedition team." This year's expeditions include two to study the proboscis monkeys of Borneo, April 19 to May 11, and May 3 to May 15, led by Carey Yeager, a Ph.D. candidate in the Psychology Department at U.C.-Davis. For a catalog, contact Desk DP1, University Research Expeditions Program, Univ. of California, Berkeley, CA 94720 [415-642-6586].
* * *
Newsletter Funding Renewed
We were very pleased to learn that PHS, through the Animal Resources Branch, Division of Research Resources, NIH, will continue funding the Laboratory Primate Newsletter for an additional five years. We thank our readers for their strong support in the form of letters and contributions over the years, and particularly in the difficult period after the death of the founding editor. The new editors are especially grateful for this show of support from our readers and the NIH reviewers. The best way to continue support for the Newsletter is to send contributions--notices, requests for information, notes, and articles of general or practical interest. Keep in mind that the Newsletter is a fast way of disseminating information to most of the people carrying out research on nonhuman primates (in the field as well as in the laboratory), who are involved in the care of these animals, or who are otherwise interested in research on these animals.
It was also gratifying to receive a donation of $100 from Dr. David Milton Taub of LABS, in Yemassee, SC. This money will be used to help defray the cost of mailing the Newsletter to overseas subscribers. If we receive several more such donations, we could discontinue charging those subscribers, which would simplify our bookkeeping here.
Schrier Laboratory Dedicated
The fourth floor of the Hunter Laboratory of Psychology, at Brown University, which previously housed the Primate Behavior Laboratory, has been dedicated as the Allan M. Schrier Research Laboratory. In a ceremony on March 10, attended by colleagues, family, and friends, a plaque was unveiled by Mrs. Schrier. After a brief speech by Departmental Chairman Julius Kling, visitors toured the laboratory, which had been designed by Professor Schrier in 1962, and built with matching funds from the Public Health Service and the University.
* * *
Books
*An Atlas of Renal Disease in Aotus Monkeys with Experimental Plasmodial Infection. M. Aikawa, J. R. Broderson, I. Igarashi, G. Jacobs, M. Pappaioanou, W. E. Collins, & C. C. Campbell. Arlington, VA: American Institute of Biological Sciences, 1988. 97pp. [Available from AIBS, 730 11th Street NW, Washington, DC 20001-4584]
*Project on the Reproduction and Conservation of Nonhuman Primates, Iquitos, Peru. Final Report, January 1982-June 1985. Pan American Health Organization & World Health Organization. Washington, DC: PAHO/WHO, 1985. 250 pp.
*Zoonoses and Communicable Diseases Common to Man and Animals. P. N. Acha & B. Szyfres. Washington, DC: PAHO, 1987. 963 pp. [Price: $20 in paper. Order from PAHO, Pan American Sanitary Bureau, Regional Office of WHO, 525 23rd Street NW, Washington, DC 20037.]
*Manipulatory Activity of Primates (Ethological Analysis in Connection with Problems of Anthropogenesis). M. A. Deryagina. Moscow: Izdatel'stvo "Nauka," 1986. 109pp. [In Russian. Price: 1.10 Rubles]
*Primate Phylogeny. G. E. Grine, J. G. Fleagle, & L. B. Martin
(Eds.). New York: Academic Press, 1988. 146 pp. [Price: $15.95, paper].
. . Papers from a symposium held at the American Museum of Natural
History, November 9, 1986.
*Field Primatology: A Guide to Research. Wolfe, L. D. New York: Garland, 1987. 288 pp.
Bibliographies
*Retroviruses in Nonhuman Primates: A Bibliography, 1983-1988. C. A. Johnson-Delaney. Seattle: Primate Information Center, 1988. (293 citations, primate and subject indexes) [Price: $10. Send order to Primate Information Center, Regional Primate Research Center, SJ-50, Univ. of Washington, Seattle, WA 98195.]
Bulletins
ICLAS Bulletin, Number 63, Autumn, 1988. (International Council for Laboratory Animal Science, CANTAB Ltd., Alexandra House, Hinchingbrooke Hospital, Huntindon, Cambridgeshire, PE18 8NT, England)
*PsyETA Bulletin. Fall, 1988, 8[1] (Psychologists for the
Ethical Treatment of Animals, P.O. Box 87, New Gloucester, ME 04260).
. . Includes a pair of articles presenting very different views of the
Canadian Council on Animal Care, the nongovernmental agency that
regulates the use of research and other animals in Canada, by H. C.
Rowsell and by J. B. Tatum.
Directories
*Resources for Comparative Biomedical Research: A Directory of the DRR Animal Resources Program. Research Resources Information Center. Bethesda: National Institutes of Health, 1988. 74 pp. [A single free copy may be obtained from the Research Resources Information Center, 1601 Research Blvd., Rockville, MD 20850]
New Journals
*The Animals Voice Magazine. [Charter subscription: $18 from
P.O. Box 1649, Martinez, CA 94553-9868]
. . This magazine intends to be "the most comprehensive source of
information available covering the broadest scope of animal defense
issues."
Reports
*Primate Report, No. 21, October, 1988. [Published in
cooperation with the German Primate Center (DPZ). Price: $8]
. . This issue consists of a census for captive primates in Europe,
edited by J. Wilde, M. H. Schwibbe, and A. Arsene.
*Primate News, 1988, Volume 22[2]. (Published by the Oregon
Regional Primate Research Center, 505 NW 185th Ave., Beaverton, OR
97006)
. . This issue focuses on the Center's in vitro fertilization/embryo
transfer program.
*REP: Annual Report 1987. Rijswick, The Netherlands: Division
for Health Research TNO, 1987.
. . This is the annual report of the REP Institutes, which stands for
the Radiobiological Institute TNO, Institute for Experimental
Gerontology TNO, and Primate Center TNO, Rijswick Z.H., The Netherlands.
Of the many short notes describing the accomplishments of the
organization, the following are concerned with primates: Partial hand
transplantation in the rhesus monkey. Phase I: technical feasibility of
the model, by S. E. R. Hovius, H. P. J. D. Stevens, M. Jonker, & J. C.
van der Meulen. Variation in epitope expression forms the basis for the
polymorphism of the rhesus monkey CD3 antigen, by F. J. M. Nooij.
Treatment of experimental allergic encephalomyelitis in rhesus monkeys
with anti-MHC class II specific monoclonal antibodies, by M. Jonker & L.
Steinman. Effects of naltrexone on alcohol drinking in rhesus monkeys,
by M. Kornet, C. Goosen, L. G. Ribbens, & J. M. van Ree. Influence of
amphetamine on mother/infant interactions in rhesus monkeys, by H.
Dienske, P. Haccou, & E. Meelis. Behaviour development of infant
chimpanzees with the mother and in peer groups, by R. P. Spijkerman.
Graffiti in rhesus monkeys: environmental deficiency, discomfort or
artistic expression, by C. Goosen, D. Wesseling, & V. Baumans.
*NABR Annual Report. Washington, DC: National Association for Biomedical Research, 1988. [Available from NABR, 818 Connecticut Avenue, N.W., Suite 303, Washington, DC 20006]
*Program Highlights, 1987. Division of Research Resources, NIH. Rockville, MD: Research Resources Information Center, 1988. NIH Publication No. 88-2309. [RRIC, 1601 Research Boulevard, Rockville, MD, 20850]
Videotapes
Our Threatened Heritage. Washington, DC: National Wildlife
Federation. 18:50 min. [Price: one copy, $20; two or more, $15 each.
Order from the N.W.F., 1400 Sixteenth St., N.W., Washington, DC
20036-2266]
. . Description of, and proposed solutions for, tropical deforestation.
A free 20-page pamphlet with the same title is available from the same
source.
Animal Models
Nonhuman primate models of human hematological malignancies.
Lapin, B.
A. (Inst. of Experimental Pathology & Therapy, USSR Acadamy of Medical
Science, Sukhumi, USSR). In Neth, Gallo, Greaves, & Kabisch (Eds.),
Haematology and Blood Transfusion: Vol. 31. Modern Trends in Human
Leukemia VII (pp. 440-443). Berlin: Springer-Verlag, 1987.
. . Some members of a
colony of baboons, which were in their 11th generation in
captivity, and which had never shown lymphoma, were innoculated with
blood from leukemia patients. After 2 years those baboons, and others
which had been exposed to them, developed malignant lymphoma.
The discovery of HVP and STLV-I in baboon malignancy, with integration
of STLV-I provirus into the DNA of lymphomatous tissue, and the
characteristic dynamics of antibody titers with their elevation in the
prelymphoma period, make it possible to conclude that baboon malignant
lymphoma is associated with DNA and RNA oncogenic viruses.
*Behavioral models of aging in nonhuman primates. Dean, R. L. &
Bartus, R. T. (Dept of CNR Research, Lederle Labs., Medical Research
Division, American Cyanamid, Pearl River, NY 10965).
Psychopharmacology of the Aging Nervous System (pp. 325-392).
Iversen, Iversen, & Snyder (Eds.). New York: Plenum, 1988.
. . Evidence is presented which supports the notion that it is possible
to use aged nonhuman primates to develop valid and reliable animal
models of age-related memory impairments.
Research using drugs to improve recent memory in aged monkeys has
produced a pattern of results that has confirmed or corroborated the
results of numerous clinical studies in aged and demented humans.
*Primate models of senile dementia. Rupniak, N. M. J. & Iversen, S.
D. In S. M Stahl, S. D. Iversen, & E. C. Goodman (Eds.),
Cognitive neurochemistry (pp. 57-72). NY: Oxford University Press,
1987.
. . A review of several different approaches to the development of
primate models of dementia and their utility for pharmaceutical
research.
*Effects of aging on visual recognition memory in the rhesus monkey.
Moss, M. B., Rosene, D. L., & Peters, A. (Dept. of Anatomy, Boston Univ.
School of Medicine, 80 E. Concord St., Boston, MA 02118).
Neurobiology of Aging, Vol. 9 (pp. 495-502). Pergamon Press, 1988.
. . Performance by 6 rhesus monkeys 26 to 27 years of age was compared
to that of six monkeys 4 to 5 years of age on a trial unique delayed
nonmatching to sample task. The finding of a recognition impairment with
age is in parallel with studies of normal human aging and lends support
to the notion that the rhesus monkey is a suitable animal model of human
aging.
*The human and simian immunodeficiency viruses HIV-1, HIV-2 and SIV
interact with similar epitopes on their cellular receptor, the CD4
molecule. Sattentau, Q. J., Clapham, P. R., Weiss, R. A., Beverley, P.
C. L., Montagnier, L., Alhalabi, M. F., Gluckmann, J.-C., & Klatzmann,
D. (Academic Dept. of Genito-Urinary Medicine, Middlesex Hospital
and University College Medical School, James Pringle House, Charlotte
Street, London, UK).
AIDS, 1988, 2, 101-105.
. . It is demonstrated, by inhibition of virus infection, blocking of
syncytium formation, and inhibition of pseudotype infection with a panel
of CD4 MAbs, that HIV-1, HIV-2, and SIV isolates share the same cellular
receptor, the CD4 glycoprotein. It is also shown that very similar
epitopes of this molecule are involved in virus binding. It is inferred
that the binding sites on these viruses are highly conserved regions and
may make good targets for potential vaccines.
*The anatomy of adipose tissue in captive
Macaca monkeys and its implications for human biology. Pond, C. M.
& Mattacks, C. A. (Dept. of Biology, The Open University, Milton Keynes,
MK7 6AA, U.K.).
Folia Primatologica, 1987, 48, 164-185.
. . In a sample of 31 sedentary, ad libitum-fed monkeys, most
specimens had less than 5% adipose tissue by weight. Total fatness
correlated closely with the number of adipocytes per kg. lean body mass,
but not at all with mean adipocyte volume, except in specimens below 5%
fat. The total number of adipocytes per kg. of lean body mass increased
more than tenfold in the most obese specimens. These data suggest that,
like humans but in contrast to laboratory rodents, adipocyte
proliferation, not adipocyte enlargement, is the chief mechanism of
adipose tissue expansion except in very lean monkeys. The distribution
and cellularity of adipose tissue in normal humans were similar to those
of exceptionally obese monkeys.
*Regional obesity and coronary artery atherosclerosis in females: A
non-human primate model. Shively, C. A. & Clarkson, T. B.
(Arteriosclerosis Research Center, Bowman Gray School of Medicine of
Wake Forest Univ., Winston-Salem, NC).
Acta Medica Scandinavica, 1988, Suppl. 723, 71-78.
. . Three experiments demonstrated relationships between amount and
location of fat deposits and coronary artery atherosclerosis,
hyperglycemia, and blood pressure. These findings suggest that female
cynomolgus macaques may serve as an animal model of the health impact of
regional obesity.
*Non-A, non-B hepatitis-related hepatocellular carcinoma in a
chimpanzee. Muchmore, E., Popper, H., Peterson, D. A., Miller, M. F., &
Lieberman, H. M. (LEMSIP, NY Univ. Medical Center, RR
1, Longmeadow Rd., Tuxedo, NY 10987).
Journal of Medical Primatology, 1988, 17, 235-246.
. . Epidemiology has indicated the possible association of non-A,
non-B hepatitis (NANBH) with hepatocellular carcinoma (HCC) in man, but
there are no means for confirmation. Chimpanzees are recognized models
for studying hepatitis B and NANBH, and may become carriers of both. The
first case of HCC to be reported in chimpanzees was found after
longitudinal study of a hepatitis B-free chimpanzee 7 years after
inoculation with human plasma from a patient reported to have chronic
NANBH.
Animal Welfare
*Inherent stress: The tough life in lab routine. Barnard, N. & Hou,
S. (Physicians Committee for Responsible Medicine, P.O. Box 6322,
Washington, DC 20015).
Lab Animal, 1988, 17[9], 21-23, 26-27.
. . A review of environmental factors which may affect animals' health
and well-being, as well as experimental results. This article was
originally published in the March, 1988, issue of
PCRM Update.
Behavior
*Party composition and dynamics in
Pan paniscus. White, F. J. (Dept. of Biological Anthropology &
Anatomy, Duke Univ., Durham, NC 27706).
International Journal of Primatology, 1988, 9, 179-193.
. . Pygmy champanzees, or bonobos, associate in parties that vary in
size and composition. Data from a 2-year field study of nonprovisioned
bonobos show that party composition varies with party size. Although
females, on average, outnumber males, the proportion of males in the
party increases in larger parties. Both females and males will join and
leave a party in the company of others, but only males appear frequently
to join or leave as lone individuals. All-male parties were not
observed, but all-female (nonnursery) parties were relatively common.
*Social structure in a wild group of
Macaca thibetana at Mount Emei, China. Deng, Z. & Zhao, Q. (Kunming
Inst. of Zoology, Acad. Sinica, Kunming, Yunnan, P.R.C.).
Folia Primatologica, 1987, 49, 1-10.
. . Data on social interactions were collected, mainly in the
non-mating season, for one wild but partly provisioned group of
Macaca thibetana whose members were individually recognized. A
linear dominance hierarchy existed among individuals as in most macaque
species studied to date.
*Group fission among wild toque macaques as a consequence of female
resource competition and environmental stress. Dittus, W. P. J. (4/4
Galkanda Road, Anniwatte, Kandy, Sri Lanka).
Animal Behaviour, 1988, 36, 1626-1645.
. . At Polonnaruwa, Sri Lanka, 4 of 29 groups of
Macaca sinica divided in a period of 16 years. It is hypothesized
that low-ranking females secede to form new groups when the costs,
especially of intragroup competition for food resources, outweigh the
benefits of group membership. Final division resulted from a coalition
of subordinate females and males acting according to their respective
interests.
*How rhesus monkey mothers and infants keep in touch when infants
are at risk from social companions. Simpson, M. J. A. (Univ. Sub-Dept.
of Animal Behaviour, High Street, Madingley, Cambridge CB3 8AA,
England).
International Journal of Primatology, 1988, 9, 257-274.
. . Vigilance in the mother-infant dyad can be studied in terms of
contacts initiated by an infant with its mother and by a mother with
her infant. One can ask if the dyads that are more vigilant in terms of
high rates and/or positive predispositions for interrupting contact with
others are ones in which the infants are at higher "risk". However,
vigilant behavior, presumed to respond to levels of risk, also reduces
the likelihood of events that allow direct measurement of risk. Mothers
may be faced with the problem of finding optimum balances between the
costs of vigilance on behalf of current infants and the costs of
investing in the next--e.g. by procuring the best father for it. Results
are discussed within the framework of a model which is progressively
elaborated in order to reveal the problems that must be faced in
exploring vigilance in a situation where the dyad's interactions are
affected by the infants' interactions with others.
*Vigilance during play in squirrel monkeys. Biben, M., Symmes, D., &
Bernhards, D. (NIHAC, Building T-18, NIH, Bethesda, MD 20892).
American Journal of Primatology, 1989, 17, 41-49.
. . Play by young
Saimiri boliviensis may put them and other troop members at risk
for predation because youngsters are noisy, separated from adults, and
not vigilant when at play. Adult female squirrel monkeys become more
vigilant during periods of spontaneous play among juveniles. Five times
as much vigilance activity was directed toward an area from which threat
or disturbance was likely to come as was directed toward the juveniles
themselves. These results suggest both an adaptive, compensatory
increase in adult vigilance during play and a function for play
vocalizations.
*Color discrimination by the cotton-top tamarin
(Saguinus oedipus oedipus) and its relation to fruit coloration.
Savage, A., Dronzek, L. A., & Snowdon, C. T. (Psychology Dept., 1202 W.
Johnson St., Univ. of Wisconsin, Madison, WI 53706).
Folia Primatologica, 1987, 49, 57-69.
. . Tamarins were trained to discriminate a set of Munsell color chips,
both within the same hue category and from the 2 hue categories on
either side of the training hue. Human subjects were tested under
similar conditions. Results indicated that the cotton-top tamarin can
make accurate discriminations across the visible spectrum. Tamarins and
human subjects had the most difficulty discriminating the same hues.
The mature coloration of fruits commonly eaten by various species of New
World monkeys was noted, and it was found that the colors range
across most of the spectrum.
*Kinship and behavior in nonhuman primates. Bernstein, I. S. (Yerkes
Regional Primate Research Center of Emory Univ., Athens, GA 30602).
Behavior Genetics, 1988, 18, 511-524.
. . Nonhuman primates show significantly more social interaction with
matrilineal kin than nonkin. There is little evidence for kin
recognition per se, and long-term association, fostered by long periods
of biological dependency, strong sociality, and impressive cognitive
capacities, is sufficient to account for the correlation between kinship
and sociality. Association, as a proximal mechanism, in no way detracts
from the adaptive significance of sociality strongly biased toward kin.
Care
*Experience in breeding free baboons on a Black Sea coast of the
Caucasus. Lapin, B. A. & Chalyan, V. G. (Inst. of Experimental Pathology
& Therapy, USSR Academy of Medical Sciences, Sukhumi, USSR).
Vestnik Akademii Medicinskih Nauk SSSR, 1987, 10, 65-70 [In
Russian, English abstract].
. . The authors have been breeding baboons in the mountainous regions
of the Caucasus not far from the towns of Sukhumi and Tuapse for 15
years. Monkeys were brought directly from their native habitat and,
after quarantine and examination, were released into the forest.
Practically all the animals adapted and acclimatized to the new, much
more severe conditions with cold, snowy winters. Natural vegetation
served as food for the monkeys except during snowy periods, when
granulated fodder was provided. There has been no substantial unfavorable
effect on the surrounding flora and fauna.
*Maintenance of cotton-top tamarins fed an experimental pelleted
diet versus a highly diverse sweetened diet. Tardif, S. D., Clapp, N.
K., Henke,
,cb
M. A., Carson, R. L., & Knapka, J. J. (Marmoset Research
Program, Oak Ridge Associated Universities, Oak Ridge, TN 37831-0117).
Laboratory Animal Science, 1988, 38, 588-59l.
. . Individually housed
Saguinus oedipus were given either a "regular" diet (monkey chow
slurry, canned diet, and supplements), a similar slurry using an
experimental natural ingredient diet plus supplements, or the
experimental diet without supplements. Neither dry food consumption,
body weight, fecal output, nor histological evaluation of the colon were
affected by these diets. Daily intake of protein and calories were
higher than previously reported estimates for the species. Results
demonstrate that a natural ingredient non-sweetened pelleted diet
is palatable for cotton-top tamarins for a period of 3.5 months.
However, further testing over longer time periods is necessary. The
nonnutritional (e.g. psychological) advantages of providing a highly
diverse diet to primates housed in a relatively monotonous environment
should be considered before adopting such a diet for an entire colony.
*Behaviour, housing and welfare of non-human primates. Poole, T. B.
(University Federation for Animal Welfare, 8 Hamilton Close, South
Mimms, Potters Bars, Herts EN6 3QD, England).
New Developments in Biosciences: Their Implications for Laboratory
Animal Science (pp. 231-237). A. C. Beynen & H. A. Solleveld (Eds.).
Dordrecht, Netherlands: Nijhoff, 1988.
. . Suggested methods for improving the captive environment, as well
as assessing the need for and the results of such changes.
*Effects of different environmental enrichment devices on cage
stereotypies and autoaggression in captive cynomolgus monkeys. Bryant,
C. E., Rupniak, N. M. J., & Iversen, S. D. (N. M. J. Rupniak, Merck
Sharp & Dohme Research Labs, Neuroscience Research Centre, Terlings
Park, Eastwick Rd., Harlow, Essex CM20 2QR, England).
Journal of Medical Primatology, 1988, 17, 257-269.
. . Autoaggression and stereotypies in individually housed cynomolgus
monkeys were compared in a standard primate cage and an enriched playpen
environment. Stereotypy and autoaggression were markedly reduced in the
playpen, but reappeared on return to the home cage. Some, but not all,
of the various activities available in the playpen engaged the animals'
attention.
Conservation
*Preliminary survey of the distribution and population of
chimpanzees in the Republic of Guinea. Sugiyama, Y. & Soumah, A. G.
(Primate Research Inst., Kyoto Univ., Inuyama, Aichi, 484 Japan).
Primates, 1988, 29, 569-574.
. . On the basis of a questionnaire, at least 20 of the 36 prefectures
in the country were confirmed to be inhabited by chimpanzees, and the
estimated total number of animals in the present population amounted to
6,625, an approximately 50% reduction from the 13,940 estimated by the
same informants on the same questionnaire based on assumptions for the
past (20-60 years ago). The authors find these data apparently
overestimated, compared with their own observations in some localities.
Urgent efforts need to be directed towards the conservation of the
chimpanzees and their habitat, especially through international
cooperative programs.
Development
*Early development of locomotor behavior in vervet monkeys.
Vilensky, J. A. & Gankiewicz, E. (Fort Wayne Center for Medical
Education, 2101 Coliseum Blvd. East, Fort Wayne, IN 46805).
American Journal of Primatology, 1988, 17, 11-25.
. . The locomotor development of three vervet infants across
approximately the first 2 months of life is described. It is suggested
that locomotor control mechanisms are probably fairly mature at birth
but that weight support and postural control problems explain initial
locomotor difficulties exhibited by these infants.
*Effects of a natural
versus artificial environment on the tempo of maturation in female
rhesus monkeys. Wilson, M. E., Gordon, T. P., Rudman, C. G., & Tanner,
J. M. (Yerkes Regional Primate Research Center of Emory Univ., Field
Station, 2409 Taylor Road, Lawrenceville, GA 30243).
Endocrinology, 1988, 123, 2653-2661.
. . Spring-born females were raised from 12 months of age under
natural outdoor conditions (OH; n = 6) or indoors (IH; n = 9) under a
controlled photoperiod and temperature. Ages at the initial increases in
serum bioactive LH levels, menarche, and first ovulation were
significantly advanced in IH compared to OH females. The data suggest
that exposure to an outdoor environment moderates the tempo of both
sexual and skeletal maturation, suggesting that common factors may be
responsible for the initiation of both events.
Disease
*Leprosy as a zoonosis: An update. Walsh, G. P., Meyers, W. M.,
Binford, C. H., Gormus, B. J., Baskin, G. B., Wolf, R. H., & Gerone, P.
J. (Leonard Wood Memorial Leprosy Research Center, Cebu City,
Phillippines).
Acta Leprologica, 1988, 6, 51-60.
. . Leprosy occurs naturally in at least 3 animal species, including
Pan troglodytes and Cercocebus atys. This suggests that nonhuman
primates may be a source of infection in humans in areas where human
leprosy is endemic and nonhuman primates abound.
*Physicochemical properties of Marburg virus: Evidence for three
distinct virus strains and their relationship to Ebola virus. Kiley, M.
P., Cox, N. J., Elliott, L. H., Sanchez, A., DeFries, R., Buchmeier, M.
J., Richman, D. D., & McCormick, J. B. (Special Pathogens Branch, Div.
of Viral Diseases, CDC, Atlanta, GA 30333).
Journal of General Virology, 1988, 69, 1957-1967.
. . The physicochemical and antigenic properties of 3 groups of
Marburg (MBG) virus isolates, separated temporally and geographically,
were compared to each other and to another member of the same family,
Ebola (EBO) virus. MGB viruses are similar to EBO viruses in morphology
and other physicochemical properties and are very similar to each other
in RNA and protein composition. Each of the distinct MBG virus outbreaks
appears to have been due to a genetically distinguishable, but
antigenically closely related virus strain. These studies confirm the
belief that MBG and EBO viruses are members of a new virus family, the
Filoviridae.
*Failure of a human immunodeficiency virus (HIV) immune globulin to
protect chimpanzees against experimental challenge with HIV. Prince, A.
M., Horowitz, B., Baker, L., Shulman, R. W., Ralph, H., Valinsky, J.,
Cundell, A., Brotman, B., Boehle, W., Rey, F., Piet, M., Reesink, H.,
Lelie, N., Tersmette, M., Miedema, F., Barbosa, L., Nemo, G., Nastala,
C. L., Allan, J. S., Lee, D. R., & Eichberg, J. W. (New York Blood
Center, 310 E. 67th St., NY, NY 10021).
Proceedings of the National Academy of Science, 1988, 85,
6944-6948.
. . To assess the possible efficacy of passive immunization against
HIV an immune globulin (termed HIVIG) was prepared from plasma of
HIV-seropositive donors selected to be among those having the top 12.5%
of virus-neutralizing antibody titers. Two chimpanzees inoculated with
HIVIG at 1 ml/kg and 2 inoculated with 10 ml/kg were challenged 1 day
later with the same strain of HIV. All animals became infected.
Incubation periods did not differ significantly from that of a control
animal. These findings may have implications for understanding the
failure of experimental vaccines to protect against HIV challenges in
chimpanzee experiments.
*Simian immunodeficiency virus/Delta-induced immunodeficiency
disease in rhesus monkeys: Relation of antibody response and
antigenemia. Zhang, J.-y., Martin, L. N., Watson, E. A., Montelaro, R.
C., West, M., Epstein, L., & Murphey-Corb, M. (M. Murphey-Corb, Delta
Regional Primate Research Center, Three Rivers Rd., Covington, LA
70433).
Journal of Infectious Disease, 1988, 158, 1277-1286.
. . Infection of the rhesus monkey with SIV induces a disease similar
to AIDS. The authors compared SIV-specific antibody and antigenemia with
the progression of disease in monkeys experimentally infected with
SVI/Delta isolates that varied in pathogenicity. The data suggest that
SIV/Delta-induced disease is dependent upon antigenemic episodes that,
particularly in animals with CNS infection, appear coincident with
diminished antibody.
*Isolation and characterization of simian immunodeficiency virus
from mandrills in Africa and its relationship to other human and simian
immunodeficiency viruses. Tsujimoto, H., Cooper, R. W., Kodama, T.,
Fukasawa, M., Miura, T., Ohta, Y., Ishikawa, K.-I., Nakai, M., Frost,
E., Roelants, G. E., Roffi, J., & Hayami, M. (Dept. of Animal Pathology,
Inst. of Medical Science, Univ. of Tokyo, Minato-ku, Tokyo 108, Japan).
Journal of Virology, 1988, 62, 4044-4050.
. . Two isolates of simian retrovirus related to HIV, obtained from
apparently healthy
Papio sphinx, in western equatorial Africa and designated
SIVMND., appeared morphologically similar to HIV by electron
microscopy, showed
Mg&S'2 -dependent reverse transcriptase activity, and induced
cytopathic effect in human CD4-positive cells. Comparisons to other SIVs
indicated that SIVMND is a new member of the HIV-SIV group,
suggesting that recent interspecies transfer of HIV-SIV is unlikely in
nature.
*Molecular evolution of the human and simian immunodeficiency
viruses. Yokoyama, S. (Dept. of Ecology, Ethology, & Evolution, Univ.
of Illinois, Shelford Vivarium, 606 East Healey St., Champaign, IL
61820).
Molecular Biology and Evolution, 1988, 5, 645-659.
. . When the entire genomes of a HIV1, a HIV2, and two SIVs were
compared, the SIVAGM showed a unique pattern of mutation
accumulations. The SIVAGM was apparently derived from cell cultures
infected with a macaque isolate, SIVMAC-251. The contamination
provides an opportunity to measure the maximum rate of evolution in the
SIVAGM by comparing its DNA sequence to those of SIVMAC-251 and
SIVMAC-142.
Ecology
*Social organization and ecology of proboscis monkeys
(Nasalis larvatus) in mixed coastal forest in Sarawak. Bennett, E.
L. & Sebastian, A. C. (World Wildlife Fund Malaysia, 7, Jalan Ridgeway,
93200 Kunching, Sarawak, Malaysia).
International Journal of Primatology, 1988, 9, 233-255.
. . Population density, social organization, and feeding and ranging
behavior, based on a 16-month study, are compared with those from other
primates in an attempt to understand why females of certain species
(including proboscis monkeys) transfer between social groups.
*Habitat use by squirrel monkeys
(Saimiri oerstedi) in Costa Rica. Boinski, S. (Lab. of Comparative
Ethology, NICHHD, NIH-Animal Center, P.O. Box 289, Bldg. 112,
Poolesville, MD 20837).
Folia Primatologica, 1987, 49, 151-167.
. . Analyses of movement patterns, habitat preferences, activity
schedules, and dispersion of squirrel monkey troop members in
relation to seasonal changes in food abundance.
*Sex differences in the foraging behavior of squirrel monkeys in a
seasonal habitat. Boinski, S. (Address same as above).
Behavioral Ecology and Sociobiology, 1988, 23, 177-186.
. . Over an 11-month period females searched for and ate food at
significantly greater frequencies than did males. The reduction of time
spent by males in foraging gives them more time for other activities,
especially anti-predator vigilance. Overall, there was little evidence
of any direct foraging benefits for a squirrel monkey from being social.
*Ecology of the saddle-back tamarin
Saguinus fuscicollis illigeri on the Río Pacaya, Northeastern
Peru. Soini, P. (P.O. Box 341, Iquitos, Peru).
Folia Primatologica, 1987, 49, 11-32.
. . Quantitative data on the diet, habitat use, ranging, and daily
activities of one focal troop, gathered between November, 1979, and
January, 1982.
*Flexibility in diets of three species of Costa Rican primates.
Chapman, C. (Dept. of Biology, McGill Univ., 1205 Dr. Penfield Ave.,
Montreal, PQ H3A 1B1, Canada).
Folia Primatologica, 1987, 49, 90-105.
. . Variability in diet and dietary overlap were documented for
Ateles geoffroyi, Alouatta palliata, and Cebus capucinus in Santa
Rosa National Park, Costa Rica. All three species exhibited great
flexibility in diet, such that in 1 month a species could be considered
one type (e.g. frugivorous), while in a subsequent month it
would be classified differently (e.g. folivorous or insectivorous).
*Baboon diet: A five-year study of stability and variability in the
plant feeding and habitat of the yellow baboons
(Papio cynocephalus) of Mikumi National Park, Tanzania. Norton, G.
W., Rhine, R. J., Wynn, G. W., & Wynn, R. D. (R. J. Rhine, Psychology
Dept., Univ. of California, Riverside, CA 92521).
Folia Primatologica, 1987, 48, 78-120.
. . The habitat and plant feeding of 64 well-habituated, individually
identified adult male and female yellow baboons was studied for 5 years.
Variation across the years showed that a study of only one or two years
would have been incomplete and misleading. The list of baboon food
species obtained is considerably larger and more diverse than any
previously reported. Baboons are eclectic feeders that appear to be
optimizing their diet by selective feeding from among a wide array of
available foods in an ever-changing floristic environment.
*Habitats of three groups of chimpanzees
(Pan troglodytes) in western Tanzania compared. Collins, D. A. &
McGrew, W. C. (Dept. of Psychology, Univ. of Sterling, Stirling FK9 4LA,
Scotland).
Journal of Human Evolution, 1988, 17, 553-574.
. . Data on the habitats of K Group at Kasoje, B Group at Bilenge
(both in the Mahale Mountains), and the main study group at Gombe. Both
Kasoje and Gombe appear particularly favorable for chimpanzees because
they have a close mosaic of forest (providing wet-season foods) and
woodland (providing foods in the dry). It is suggested that their
habitats differ from the relatively drier environments in which, on
present fossil evidence, the known Pliocene hominids are thought to have
lived, but that the ecology of these chimpanzees may provide appropriate
models for the adaptations of hominoids in the Miocene forest-woodland
mosaics.
*Influence of artificial food supply on population parameters and
dispersal in the Hakone T troop of Japanese macaques. Fukuda, F.
(2-27-12, Shohnandai, Fujisawa, Kanagawa, 252 Japan).
Primates, 1988, 29, 477-492.
. . A free-ranging troop of Japanese monkeys, artifically provisioned
at two different levels at different periods, was observed for seven
years. Clear differences between the two periods were found in
primiparous age, ratio of non-natal resident males to all resident
males, ratio of newcomers to non-natal resident males, age-specific
disappearance rate, and proportion of deserters observed in the study
area.
*An ecological comparison of two sympatric primates:
Saguinus fuscicollis and Callicebus moloch of Amazonian Peru.
Crandlemire-Sacco, J. (Dept. of Anthropology, Univ. of Pittsburgh,
Pittsburgh, PA 15260).
Primates, 1988, 29, 465-475.
. . Tamarins and titis showed very little overlap in their choices of
food plants or animal foods and used distinctive foraging styles,
exploiting very different parts of the habitat for animal prey.
*Forest monkey populations in the Central African Republic: The
northern limits. A census in Manovo-Gounda-St. Floris National Park.
Fay, J. M. (Dept. of Anthropology, Washington University, Box 299, St.
Louis, MO 63166-0299).
Mammalia, 1988, 52, 57-74.
. . Eight species of diurnal anthropoids were recorded during a
3-month census, covering about 17,000 hectares with 89.08 km of
transects. Twenty-two percent of all observations were of mixed groups
with a maximum of 3 species observed together.
Evolution
*Finite social space, evolutionary pathways, and reconstructing
hominid behavior. Foley, R. A. & Lee, P. C.
Science, 1989, 243, 901-906.
. . Changes in social behavior were a key aspect of human evolution,
yet it is notoriously difficult for paleobiologists to determine
patterns of social evolution. By defining the limited number of
distributional strategies available to members of each sex of any
species and investigating the conditions under which they may occur and
change, the social behavior of different hominid taxa may be
reconstructed.
Genetics
*The genetic structure of a primate species: Rhesus macaques and
other cercopithecine monkeys. Melnick, D. J. (Genetics Lab.,
Anthropology Dept., Columbia Univ., New York, NY 10027).
International Journal of Primatology, 1988, 9, 195-231.
. . The author's objectives are to describe the distribution of
genetic diversity among five hierarchical levels of animal aggregation
across the species range of the rhesus monkey; to compare the genetic
structure of the rhesus with that of other cercopithecine monkeys,
particularly the closely related
M. fascicularis; and to examine alternative explanations for the
existing structure of primate genetic variation.
Instruments and Techniques
*Ultrasonic assessment of pregnancy and fetal development in three
species of macaque monkeys. Shimizu, K. (Primate Research Inst.,
Inuyama, Aichi 484, Japan).
Journal of Medical Primatology, 1988, 17, 247-256.
. . Serial ultrasonic assessments of gestational sac and fetal
biparietal diameter were performed in the Japanese monkey, rhesus
monkey, and crab-eating monkey.
*Pregnancy diagnosis by the ultrasonographical device and
observation of fetal growth in the squirrel monkey
(Saimiri sciureus). Narita, H., Hamano, M., & Cho, F. (Corp. for
Production and Research of Laboratory Primates, National Inst. of
Health, Hachimandai, Tsukuba-shi, Ibaraki-ken 305, Japan).
Experimental Animals, 1988, 37, 393-397 (Japanese, with English
summary, figures, and table).
. . Pregnancy was diagnosed on the basis of the detection of the
.tr ` 9E
gestational sac (GS), which was first detected 127`10 days before
delivery. Conditions of fetal growth were judged by measuring the GS
and the biparietal diameter.
*Subcutaneous implantable infusion device for chronic intravenous
pulsatile gonadotropin-releasing hormone treatment. Schmutzler, R. van
Uem, J. F. H. M., Elkind-Hirsch, K. Hahn, D.-W., McGuire, J. L.,
Danforth, D. R., & Hodgen, G. D. (G. D. Hodgen, Jones Inst. for
Reproductive Medicine, Dept. of OB/GYN, Eastern Virginia Medical School,
Medical College of Hampton Roads, Norfolk, VA 23510).
Journal of Medical Primatology, 1988, 17, 281-285.
. . Preliminary data indicate the potential utility of an implantable
subcutaneous device that facilitates chronic intravenous infusion of
pulsatile gonadotropin-releasing hormone (GnRH) for ovulation induction.
GnRH distribution curves were congruent in control monkeys and those
with implanted devices. Tissue tolerance was good in this brief trial.
*Head restraint device for chronic recording of neural activity in
the awake monkey. Buchholz, S. R. & Montgomery, E. B. Jr. (Dept. of
Neurology, Washington Univ. School of Medicine, 660 S. Euclid Ave., Box
8111, St. Louis, MO 63110).
Journal of Neuroscience Methods, 1988, 25, 139-141.
. . An adjustable, stainless steel head restraining device and small
screw-tightened skull clips are described which enable the recording of
single unit neuronal activity chronically from awake, behaving primates
of various sizes. This method has been used over the past 4 years on 8
primates, for periods up to 18 months without any mechanical failures.
The equipment reduces problems stemming from bone necrosis and is well
tolerated.
Pharmacology & Anesthesia
*Rearing paradigm in a nonhuman primate affects response to beta-CCE
challenge. Insel, T. R., Scanlan, J., Champoux, M., & Soumi, S. J. (Lab.
of Clinical Science, NIMH/NIHAC, P.O. Box 289, Poolesville, MD 20837).
Psychopharmacology, 1988, 96, 81-86.
. . Two groups of socially housed rhesus monkeys were raised under
conditions differing in the degree of control over food, water, and
treats in the first year of life. At age 18-24 months, a benzodiazepine
receptor inverse agonist, beta-carboline-3-carboxylic acid ethyl ester
(beta-CCE), was administered to both social groups to investigate the
effects of differential rearing on the response to a putative anxiogenic
drug. In both groups beta-CCE was followed by increases in aggression and
plasma cortisol, but the subjects reared with control showed greater
increases in aggression and less increase in plasma cortisol.
These results suggest that early experience with controllability may
have long term consequences which can be revealed by challenge with
beta-CCE.
Physiology
*Androgen and estrogen dynamics in the female baboon
(Papio anubis) . Longcope, C., Femino, A., & Johnston, J. O. (Dept.
of Obstetrics/Gynecology & Medicine, Univ. of Massachusetts Medical
School, Worcester, MA 01605).
Journal of Steroid Biochemistry, 1988, 31, 195-200.
. . Androgen and estrogen dynamics were studied in 5 female baboons,
by infusion with radioactively marked hormones. The general pattern of
androgen and estrogen dynamics is similar to that in women. This
similarity is due, in part, to the presence of sex-hormone binding
globulin in both baboons and women.
*Circadian pattern of plasma melatonin concentrations in the
marmoset monkey
(Callithrix jacchus). Webley, G. E., Abbott, D. H., George, L. M.,
Hearn, J. P., & Mehl, H. (Inst. of Zoology, Regent's Park, London NW1
4RY, UK).
American Journal of Primatology, 1989, 17, 73-79.
. . A circadian pattern of melatonin secretion was apparent in plasma
samples taken from marmosets every 4 hours over 3 24-hour periods.
Significantly higher levels were recorded at night than during the day.
Concentrations were similar over the 3 24-hour periods, in males and
females, and in dominant and subordinant females. The marmoset's
circadian profile is less pronounced than that seen in the human,
but is similar to that of the seasonally breeding rhesus monkey.
Reproduction
*Effects of hypothalamic lesions upon the sexual and social
behaviour of the male common marmoset
(Callithrix jacchus). Lloyd, S. A. C. & Dixson, A. F. (MRC
Reproductive Biology Unit, Centre for Reproductive Biology, 37 Chalmers
St., Edinburgh EH3 9EW, U.K.).
Brain Research, 1988, 463, 317-329.
. . Damage to the anterior hypothalamus or its junction with the
preoptic area in male marmosets causes a profound suppression of sexual
'arousal' and copulatory behavior. Such effects are not due to androgen
insufficiency or other, nonspecific, side effects of neural damage.
*Reproduction in the vervet monkey
(Cercopithecus aethiops): III. The menstrual cycle. Eley, R. M.,
Tarara, R. P., Worthman, C. M., & Else, J. G.
American Journal of Primatology, 1989, 17, 1-10.
. . The menstrual cycles of 17 multiparous vervet monkeys were
studied. Based on estradiol, progesterone, and LH profiles, ovulation is
predicted to occur on day 13 of the 32.4-day menstrual cycle. Estradiol
peaked on the day preceding the LH peak in 75% of cycles. Average luteal
phase length was 18 days, with progesterone rising above 4 nmol/l on the
day of the LH peak. Vaginal cytology and perianal skin coloration
exhibited too much within- and among-animal variability to be reliable
indicators of menstrual cycle stages. Uterine biopsies of the
proliferative phase were characterized by mild pseudostratification of
the columnar epithelium and absence of glandular secretion; in contrast,
those of the luteal phase had marked pseudostratification of the tall
columnar epithelium with glandular secretions in the lumen.
*Periparturitional behavior of a bonobo
(Pan paniscus). Bolser, L. & Savage-Rumbaugh, S. (Language Research
Center, Dept. of Biology, Georgia State Univ., Atlanta, GA 30303).
American Journal of Primatology, 1989, 17, 93-103.
. . The periparturitional behaviors of a mature bonobo female during
the pregnancy and birth of two infants are described in detail.
Information not previously reported for the species is included.
Periparturitional behaviors were similar for both births.
*Mating patterns in the golden lion tamarin
(Leontopithecus rosalia): Continuous receptivity and concealed
estrus. Stribley, J. A., French, J. A., & Inglett, B. J. (Dept. of
Psychology, Univ. of Nebraska, Omaha, NE 68182).
Folia Primatologica, 1987, 49, 137-150.
. . Sociosexual behavior and urinary estrogen levels were monitored on
a daily basis for 3 months in 5 pairs of golden lion tamarins. Mounts
and copulations were observed during all phases of the estrogen cycle,
and there were no significant changes in affiliative behavior associated
with changes in urinary estrogen levels. The lack of a conspicuous
sexual signal in these animals may be related to a pattern of continuous
sexual receptivity.
*Female sexual swellings in the Asian colobine
Simias concolor. Tenaza, R. R. (Biology Dept., Univ. of the
Pacific, Stockton, CA 95211).
American Journal of Primatology, 1989, 17, 81-86.
. . The pig-tailed langur is the first Asiatic colobine found to have
prominent sexual swellings and the only colobine with sexual swellings
that lives in one-male groups.
*Copulatory patterns of primates as viewed in broad mammalian
perspective. Dewsbury, D. A. & Pierce, J. D., Jr. (Dept. of Psychology,
Univ. of Florida, Gainesville, FL 32611).
American Journal of Primatology, 1989, 17, 51-72.
. . Literature on copulation in 67 primate species is reviewed, and a
tentative classification is proposed for 33 primate species, in relation
to the presence or absence of 4 defining attributes: a lock, thrusting
during intromissions, multiple intromissions, and multiple ejaculations.
Although neither social organization nor genital morphology provides a
fully satisfactory relationship, there are indications that variations
in copulatory behavior are related to variations in genital morphology.
*Metachlopromide-induced hyperprolactinemia fails to affect ovarian
cyclicity in the common marmosets
(Callithrix jacchus) . Kholkute, S. D., Patil, R. P., Desai, M. P.,
& Puri, C. P. (Inst. for Research in Reproduction (ICMR), Parel, Bombay
400012, India).
Primates, 1988, 29, 549-555.
. . Intramuscular administration of metachlopromide (2.5, 5, and 10
mg) induced a dose-dependent increase in plasma prolactin. The magnitude
and duration of metachlopromide-induced hyperprolactinemia were also
dose related, but metachlopromide treatment (5 mg/day) for 60 days
failed to affect ovarian function in the common marmoset as evidenced by
ovulatory plasma estradiol and progesterone profiles.
Taxonomy
*Evolution of immunoglobulin allotypes and phylogeny of apes.
Dugoujon, J.-M. & Hazout, S. (Centre de Recherche sur le Polymorphisme
Génétique des Populations Humaines du CNRS, CHU Purpan, F-31300
Toulouse, France).
Folia Primatologica, 1987, 49, 187-199.
. . Serum samples from 72
Pan troglodytes, 5 P. paniscus, 22 Gorilla gorilla, 23 Pongo
pygmaeus abelii, 5 P.p. pygmaeus, 2 hybrids P.p. abelli X
P.p. pygmaeus, and 13 Hylobates lar were tested for Gm and Bm
immunoglobulin allotypes by the classical hemagglutination inhibition
method. The distribution of the various alleles and phenotypes makes it
possible to distinguish each species or subspecies. Common chimpanzees
have the richest polymorphism. Pygmy chimpanzees share common phenotypes
with gorillas. Bornean and Sumatran orangutans have their own patterns
of polymorphism, as do gibbons. Principal component plot and dendrogram
are compatible with the traditional classification of Hominoidea in 3
families: Hominidae, Pongidae, and Hylobatidae.
*A new species of guenon (genus
Cercopithecus) from Gabon. Harrison, M. J. S. (Dept. of Zoology,
Univ. of Edinburgh, Edinburgh EH9 3JT, UK).
Journal of the Zoological Society of London, 1988, 215, 561-575.
. . A previously undescribed member of the
Cercopithecus lhoesti superspecies, with distinctive external
characteristics, is described and named
C. solatus.
*Anthropoid affinities of
Tarsius supported by lens beta-A-crystallin sequences. deJong, W. W. &
Goodman, M. (Dept of Biochemistry, Univ. of Nijmegen, P.O. Box 9101,
6500 HB Nijmegen, Netherlands).
Journal of Human Evolution, 1988, 17, 575-582.
. . The amino acid sequences of the eye lens protein alpha-A-crystallin of
Tarsius syrichta and Aotus trivirgatus have been determined and
compared with those of 3 lemuriform and 2 cattarrhine primates and 40
other mammalian species. The
Aotus alpha-A sequence is clearly grouped with those of rhesus monkey
and human on the basis of 3 shared derived amino acid replacements.
Tarsier alpha-A has one residue, 146 Ile, in common with the Anthropoidea,
where the Lemuriformes have 146 Val.
-------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
-------------------------------------------------------------------
* * *
Zoo Animal Management
The Dallas Zoo will host this year's national workshop on "Applying Behavioral Research to Zoo Animal Management," April 29 to May 6, 1989. Participants will learn how to develop and participate in behavioral research projects and apply the results to specific management problems. This year's workshop will include topics and projects in mammalian, avian, and reptilian behavior. A nationally recognized behavioral scientist will provide a keynote lecture and many additional activities are planned. Enrollment will be limited to 40 participants, with priority given to zoo staff from AAZPA accredited institutions. For further information, contact Ron Kagan, General Curator, Dallas Zoo, 621 East Clarendon Drive, Dallas, TX 75203 [214-670-6833].
International Ethological Conference
The XXIst International Ethological Conference, dedicated to the integration and synthesis of all aspects of animal behavior, will meet August 9-17, 1989, in Utrecht, The Netherlands. The Conference is open to all, including students. For further information, write to XXIst International Ethological Conference, c/o QLT Convention Services, Keizergracht 792, 1017 EC Amsterdam, The Netherlands.
Animal Behavior Society
The 25th Anniversary meetings of the ABS will be held at Northern Kentucky University, Highland Heights, KY, June 11 to 17, 1989. Besides papers, symposia, and posters, there will be a full-day symposium on the history of the study of animal behavior, organized by Donald A. Dewsbury. For further information about the meetings, contact Tom C. Rambo, Dept. of Biological Sciences, Northern Kentucky Univ., Highland Heights, KY 41076 [606-572-5305]. Concerning the history symposium, contact Dr. Dewsbury, Dept. of Psychology, Univ. of Florida, Gainesville, FL 32611.
Asociacon Mexicana de Primatología
The Asociacion Mexicana de Primatología (AMP) is a young organization whose goals include fostering scientific exchange among primatologists and the enhancement of conservation efforts in Mesoamerica. A joint meeting of the American Society of Primatologists and AMP is tentatively scheduled for 1992 in Cancun, Mexico. Those interested in learning more about AMP should contact the President, Alejandro Estrada, Instituto de Biologia, Universidad Nacional Autonoma de Mexico, Apartado Postal 176, San Andres Tuxtla, Veracruz, Mexico.
PHS Policy Workshop
Two workshops on the implementation of Public Health Service policy on laboratory animals, sponsored by the Office for Protection from Research Risks of NIH, have been scheduled. These workshops are open to institutional administrators, members of animal care and use committees, laboratory animal veterinarians, investigators, and those with responsibility for institutional animal care and use programs. For the workshop at Davis, CA, May 8-9, contact Julie Lamoree, Office of Campus Veterinarian, Univ. of California, Davis, CA 95616 [916-752-2364]. For June 8-9 at Memphis, TN, contact Jean Littlejohn, Animal Resources, St. Jude's Children's Research Hospital, Memphis, TN 38101 [901-522-0385].
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. (Phone: 401-863-2511)
Judith_Schrier@brown.edu
ACKNOWLEDGMENTS
The Newsletter is supported by U. S. Public Health
Service Grant RR-00419 from the Animal Resources Program,
Division of Research Resources, N. I. H.
Cover drawing of an infant marmoset (Callithrix jacchus) by Penny Lapham.
Copyright © 1989 by Brown University
Editor: Judith E. Schrier, M. Sc.
Associate Editor: James S. Harper, D.V.M.
Consulting Editor: Morris L. Povar, D.V.M.
Founding Editor: Allan M. Schrier, Ph.D.
{kind=link}
{kind=link}
{kind=link}
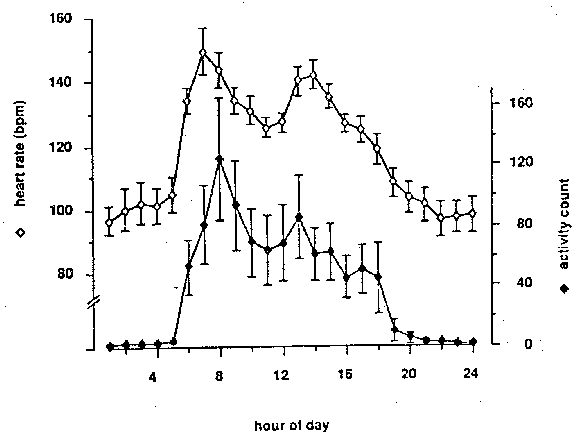{kind=link}
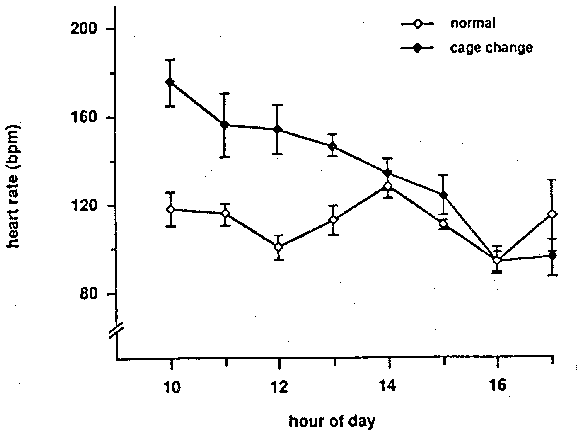{kind=link}
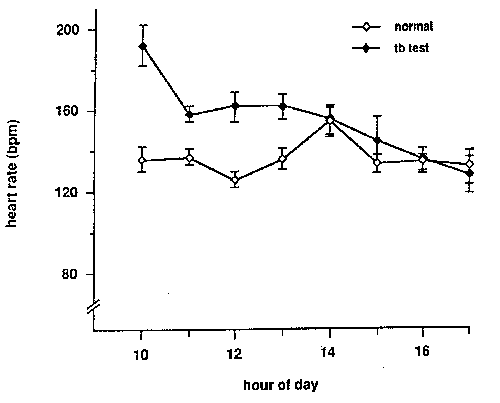{kind=link}
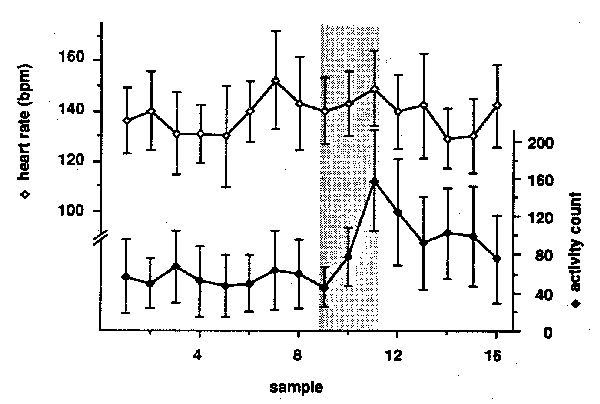{kind=link}
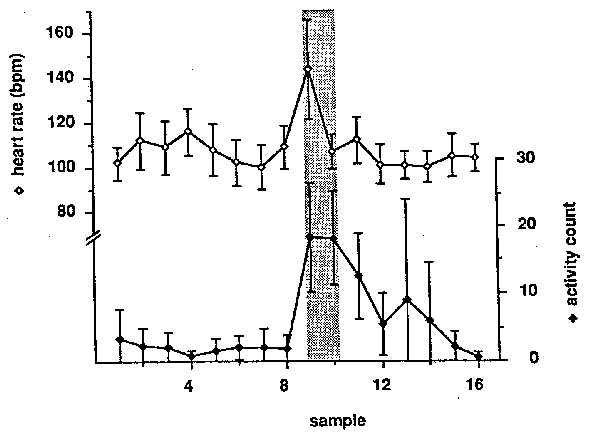{kind=link}
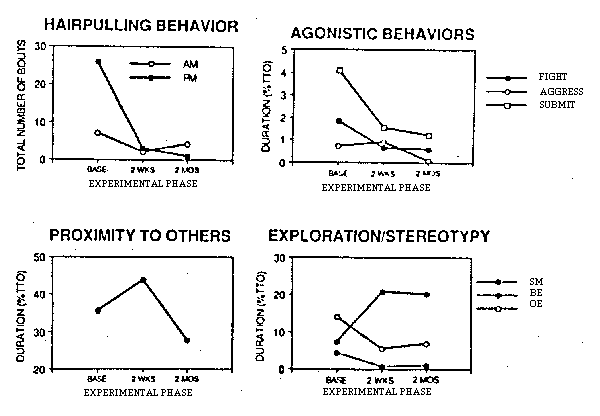{kind=link}
{kind=link}
{kind=link}