 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 28 NUMBER 4 OCTOBER 1989
Articles and Notes
Rehousing Nonreproductive Rhesus Macaques with Weanlings: I. Behavior of Adults toward Weanlings, by M. Champoux, B. Metz, & S. Suomi ...... 1
Rehousing Nonreproductive Rhesus Macaques with Weanlings: II. Attachment Reformation by Weanlings, by M. Champoux, J. D. Higley, & S. Suomi ...... 4
Apparent Health, Hidden Disease: Vitamin B12 Deficiency and the Nervous System, by C. Oxnard ...... 7
Technique for Non-Invasive Marking of Infant Primates, by E. Halloren, E. C. Price, & W. C. McGrew ...... 13
A New Monkey Swing, by K. Bayne, S. Suomi, & B. Brown ...... 16
Re-pairing Caged Rhesus Monkeys, by V. Reinhardt ...... 19
News, Information, and Announcements
Travel to IPS Congress in Japan ...... 12
. . Travel Funds Available; Travel Agencies Offer Services
Confiscated Apes in Sarawak ...... 12
Tips from the Metro Toronto Zoo ...... 15
Research Opportunities ...... 1a5
. . Lake Naivasha, Kenya; Foundation for Field Research
Grants Available ...... 17
. . Facility Improvements for Small Programs; L. S. B. Leakey Foundation;
Office of Naval Research; Research Training Groups Program
News Briefs ...... 20
. . Animal Welfare Regulations; New Antimalarial Drugs; Tamarins & Cocoa
Extending Your Lecture's Life ...... 22
Meeting Announcements ...... 22
. . Lion-Tailed Macaques; Australian Primate Society
Revision of Directory of Graduate Programs in Primatology and Primate Research Planned ...... 22
Information Available ...... 33
. . CAUZ, Lion-Tailed Macaque Newsletter; Animal Transportation;
Japanese Macaque Book; Diagnon Corporation
Birth Announcement ...... 36
Departments
Position Available ...... 21
Recent Books and Articles ...... 24
Address Changes ...... 36
* * *
Maribeth Champoux, Becky Metz, and Stephen J. Suomi
University of Wisconsin and NICHD
Introduction
In primate colonies, a key concern is the maintenance of optimal psychological well-being of all animals. One management issue concerns the disposition of breeders as age renders them nonreproductive. These animals may be relegated to single-cage housing, with its marked restriction of social interaction with conspecifics. Provision of adequate socialization of weaned infants is another concern. In our laboratory, in order to allow lactating females to resume sexual cycling for the next breeding season, infants are removed from their mothers at 6 months of age. Continued and adequate socialization is imperative for these weanlings, both for their own well-being and for practical colony management as well. It has been demonstrated that rhesus infants reared only with mothers but no peer experience are hyperaggressive when socialized as adults (Alexander, 1966). Conversely, those monkeys which are reared only with peers from birth (together-together monkeys: Chamove, Rosenblum, & Harlow, 1973) are delayed in the development of sexual and play patterns and are deficient in social skills as adults.
For the past several years at the Harlow Primate Laboratory we have been housing nonreproductive adults, primarily aged animals of feral origin, with weanlings in large social groups. The original purpose was to provide social companions for the aged adults, as well as to provide the weanlings with disciplinary figures and models for appropriate social behavior. In this and the following report we describe the behavior of the nonreproductive adults to their weanling companions, and the redirection of the weanlings' attachment bond from mother to their adult cagemates.
The behavior of aged or nonreproductive monkeys, both in the laboratory and in the field, has received little study. Aged animals spend more time resting, and less time engaged in social interaction and locomotion, than their non-aged counterparts (Boer & Davis, 1968; Hauser & Tyrrell, 1984; Nakamichi, 1984). The reasons that have been proposed for the decline in sociability in aged animals include physical disability associated with aging, loss of dominance status, and deliberate choice of other aged animals as preferred social companions. Armed with this knowledge, we investigated what form the interactions of aged nonreproductive adults with infants would take under conditions of forced proximity in the laboratory.
Methods
Subjects were nine postreproductive rhesus macaques (Macaca mulatta), five females and four males. All animals except one female and one male were of feral origin. The feral-born monkeys were postreproductive due to advanced age, whereas the two non-aged laboratory-born monkeys were nonreproductive for medical reasons. The subjects ranged from 13 to 29 years of age (ages estimated for feral-born animals).
Each animal was placed into a social group which consisted of 6 to 12 weanling rhesus macaques. In all but one case, subjects were placed into the group of weanlings with an aged partner of the opposite sex, resulting in a total of five social groups altogether. Five subjects were placed with groups of 6-month-old infants; four were placed with groups of 18-month-olds. Each social group occupied one-half (2 units) of a nuclear-family apparatus cage (Harlow, 1971). Each group of weanlings consisted of a mixture of mother-reared and nursery/peer-reared animals. Mother-reared animals had lived with their mothers in mother-infant dyads until 5-6 months of age. Nursery/peer-reared monkeys had been reared in the Harlow Primate Laboratory nursery facility until 30 days of age using the procedures described by Ruppenthal (1979), after which they were placed into peer-only social groups. All weanlings had lived together for at least 2 weeks prior to being rehoused with adult pairs.
Behavioral observations were conducted by the second author between 1000-1200 or 1300-1500. The total frequencies and durations of 14 predefined behaviors (see Table 1 for a description of behaviors) were recorded on a battery-operated laptop computer during 5-min focal animal testing sessions. All animals were tested 4 times/week for a 10-14 month testing period, resulting in a total of 155-219 testing sessions, or approximately 13-18 hours total testing time per subject. For purposes of analysis, data were combined into three 3-month blocks for all subjects; only the last 9 months of obtained data were analyzed. Data were analyzed using three-way analyses of variance, with sex of subject and age of weanling (6 mo. vs 18 mo.) as between-subject factors, and three month blocks as a within-subjects factor.
Self-directed: Any self-contact such as self-mouthing, self-
grooming, self-scratching, etc.
Groom infant: Discrete picking through fur of infant
cagemate.
Groom adult: Discrete picking through fur of adult
cagemate.
Sex infant: Any sexual behavior (e.g. mounts, presents)
directed toward infant cagemate.
Sex adult: Any sexual behavior (e.g. mounts, presents)
directed toward adult cagemate.
Environmental exploration: Visual, manual, oral, or pedal
manipulation or examination of
Stereotypy: Any repetitive locomotor or
nonlocomotor activity,
repeated for at least 3 cycles.
Discipline/aggression: Acts such as threat faces, biting, slapping,
pushing.
Passive: Sitting/standing quietly in the absence of
any social or locomotor behaviors.
Locomotion: Self-induced change in location.
Proximity: Sitting/standing/lying within arm's reach of
another animal.
Mutual ventral: Ventral/ventral contact, with or without
nipple contact.
Contact cling: Close body contact which is not ventral
oriented.
Vocalization: Any sound made by the subject (except
sneezes, coughs, etc.).
Table 1: Behavioral Definitions
Results
The behavioral profiles of the nonreproductive adults are presented in Table 2. For both male and female adults, the majority of time was devoted to proximity to infant and adult cagemates, and passive behavior (i.e., sitting alone). Affiliative behaviors directed toward infants such as grooming, ventral clinging, and physical contact, made up the next largest block of time. Sexual behavior, either with other adults or infants, was extremely rare, averaging less than 0.2 sec per 5-min testing session. Aggressive or disciplinary acts toward the infants were also infrequent, averaging less than 0.5 incidences per 5-min testing session. Play with infants was exhibited by one female only.
Age of Infant
6-month old 18-month old
Sex of Subject Female Male Female Male
------------------------------------------------
Behavior
Self-directed 10.1 15.7 13.0 10.9
Groom infant 7.6 1.2 18.3 15.5
Groom adult 12.5 11.3 8.0 7.9
Environ. explore 6.7 10.2 1.7 1.6
Stereotypy 0.0 1.0 19.1 3.7
Passive 20.4 32.3 29.8 20.6
Locomotion 2.9 7.0 5.3 3.2
Proximity 36.3 26.7 18.5 36.6
Mutual ventral 12.3 1.1 0.5 1.9
Contact cling 10.1 12.5 6.1 12.3
Discipline/aggress.** 0.6 0.3 0.2 0.5
Vocalization** 0.7 0.3 0.9 0.3
* Values add up to > 100% because not all categories are mutually exclusive.
** Values indicate mean frequency in 300-sec testing session.
Table 2: Percentages of time spent in each behavior by subject category*
Adult females were more likely than males to exhibit ventral clinging with the weanlings (F [1,5] = 12.34, p < .05; 19.1 vs. 4.5 sec). Females also spent more time in stereotyped behavior than males, (F [1,5] = 13.80, p < .05; 28.7 sec vs. 7.1 sec). There was a nonsignificant trend (p < .06) for females to spend more time grooming infants (38.9 sec vs. 25.0 sec). No sex differences were observed in any other behaviors.
Six-month-old weanlings received more ventral clinging than 18-month-olds (F [1,5] = 9.25, p < .05; 20.1 vs. 3.6 sec), whereas 18-month- olds were the recipients of more grooming (F [1,5] = 45.25, p < .01; 50.7 sec vs. 13.2 sec). The adults housed with 18-month-olds engaged in more stereotyped behaviors (F [1,5] = 31.91, p < .005; 34.3 sec vs. 1.5 sec). There were borderline significant (p < .07) trends for adults with 18-month-olds to exhibit less grooming of adult cagemates (23.7 vs. ,sk 20 35.6 sec) and less environmental exploration (4.9 vs. 25.3 sec).
Most behaviors were stable across the 9-month testing period, with a few exceptions. Disciplinary and aggressive acts toward wean- lings decreased across time (F [2,10] = 6.88, p < .05), as did locomotion (F [2,10] = 12.44, p < .005). Contact cling increased during the study (F [2,10] = 5.72, p < .05).
Discussion
These data indicate that retired rhesus macaque breeders can serve as compatible social companions for large groups of weanlings. The predominant interactions between adults and weanlings were affiliative in nature, with little aggression expressed toward weanlings by the adult cagemates. Females were particularly responsive to the weanlings, engaging in more ventral cling and grooming than their male counterparts. This sex difference in ventral clinging was not simply a result of males displaying less ventral clinging than females overall, however. Whereas all females in the study engaged in ventral clinging with their young companions, not all males did so. However, those males which did express ventral cling as part of their behavioral repertoire did so as frequently as did females. Although the propensity to exhibit ventral clinging appears to be an individual characteristic for adult males, all males in the study engaged in other types of positive interaction with the infants, which mainly took the form of nonventral physical contact. We conclude that adults of either sex can be placed into infant groups with excellent results.
In our laboratory we have been routinely using this housing procedure as a method of socialization for both mother-reared and nursery/peer-reared animals from 6 to 24 months of age. Although this has not been tested directly, anecdotal observations indicate that this housing situation reduces aggression within the group of weanlings. The impact on the adults housed with large groups of infants is less clear. Studies in which adult macaques have been paired with conspecifics (Reinhardt et al., 1987a; Reinhardt et al., 1987b; Reinhardt, 1989) indicate that providing social companions is an effective method of environmental enrichment for animals that would otherwise be singly caged. Our results show that adult macaques placed with large groups of infants form affiliative social relationships with the infants, indicating that this housing procedure constitutes an efficient and practical method of environmental enrichment as well.
References
Alexander, B. K. (1966). The effects of early peer deprivation on juvenile behavior of rhesus monkeys. Doctoral dissertation, University of Wisconsin, Madison.
Boer, A. P. & Davis, R. T. (1968). Age changes in the behavior of monkeys induced by ionizing radiations. Journal of Gerontology, 23, 331-342.
Chamove, A. S., Rosenblum, L. A., & Harlow, H. F. (1973). Monkeys (Macaca mulatta) reared only with peers: A pilot study. Animal Behaviour, 21, 316-325.
Harlow, M. K. (1971). Nuclear family apparatus. Behavior Research Methods and Instrumentation, 3, 301-304.
Hauser, M. D. & Tyrrell, G. (1984). Old age and its behavioral manifestations: A study on two species of macaque. Folia Primatologica, 43, 24-35.
Nakamichi, M. (1984). Behavioral characteristics of old female Japanese monkeys in a free-ranging group. Primates, 25, 192-203.
Reinhardt, V. (1989). Behavioral responses of unrelated adult male rhesus monkeys familiarized and paired for the purpose of environmental enrichment. American Journal of Primatology, 17, 243-248.
Reinhardt, V., Cowley, D., Eisele, S., Vertein, R., & Houser, W. D. (1987a). Preliminary comments on pairing unfamiliar adult female rhesus monkeys for the purpose of environmental enrichment. Laboratory Primate Newsletter, 26[2], 5-8.
Reinhardt, V., Houser, W. D., Eisele, S. G., & Champoux, M. (1987b). Social enrichment of the environment with infants for singly caged adult rhesus monkeys. Zoo Biology, 6, 365-371.
Ruppenthal, G. C. (1979). Survey of protocols for nursery-rearing infant macaques. In G. C. Ruppenthal (Ed.), Nursery Care of Nonhuman Primates (pp. 165-185). New York: Plenum Press.
-------------------------------------------------------------------
First author's current address: Primate Facility 7-930T,
Department of Psychiatry and Behavioral Sciences, Stanford University
School of Medicine, Stanford, CA 94305-5095.
The authors would like to thank Chris Ripp, Jr. This experiment
was conducted as part of the Intramural Research Program of the
Laboratory of Comparative Ethology, NICHD.
* * *
Maribeth Champoux, James D. Higley, and Stephen J. Suomi
University of Wisconsin and NICHD
Introduction
Rhesus macaque infants typically form strong attachments to their mothers or mother substitutes within the first weeks of life (Harlow & Harlow, 1965). The work of Mason and his colleagues (Mason & Kenney, 1974; Mason & Capitanio, 1988) has demonstrated that rhesus monkeys reared with dogs or inanimate surrogates are capable of redirecting their attachment following separation from the initial attachment figure. However, the possibility of transferral of the attachment bond has not been empirically tested in mother-reared monkey infants. In free-ranging rhesus, individuals other than the mother may provide infant care (Berman, 1982). Whereas redirection of attachment may occur under these conditions, it is unclear whether the bond with the original filial attachment source remains intact, or if the attachment to the new caregiver supercedes the previous filial bond. The present study investigated whether yearling rhesus monkeys which had been living with their mothers for several months would form attachments to other adult females following prolonged separation from their mothers and constant access to the new females. In addition, the study addressed whether these new attachments would supercede those initially formed to the mother. Attachment was assessed by the tendency of the juveniles to seek proximity to the attachment figure under mildly stressful conditions. This measure has been aptly used to study attachment in primates (Harlow, 1958; Mason & Capitanio, 1988).
Methods
Subjects were five laboratory-born rhesus macaques (Macaca mulatta) ranging from 8 to 14 months of age at the time of testing. Subjects were raised in mother-infant dyads until 3 to 6.5 months of age. Following maternal separation, infants were placed in peer groups for 2 to 7 months until they were 6 to 12 months of age. Two months prior to the beginning of the study, animals were transferred as a unit into a social group containing three unrelated aged adults of feral origin and five age-matched peers. One adult was a 32-year old male; the others were postreproductive multiparous females, 25 and 32 years old.
Each monkey was tested in the Sackett Self-Selection Circus (Sackett, 1970). The experiment consisted of 3 preference tests. During each test, the monkey had 4 stimulus animals among which it could choose. For Test 1 the choice was between aged adult female cagemate A, an unfamiliar aged female, an unfamiliar multiparous female similar in age to the animal's mother, and the animal's own mother. Test 2 was identical to Test 1 except the other aged adult female cagemate (female B) was used. Test 3 pitted aged adult female cagemates A and B against an unfamilar adult female and the monkey's mother. Two trials were run on each preference test; therefore, all animals received 6 trials in the Circus, with the exception of one animal whose data for Test 2 were incomplete and thus were not included in the analysis. All six trials were completed in a 3-week period.
Each test trial consisted of a habituation period, during which no data were collected, followed by a choice period. During habituation, monkeys were placed in the central start compartment with the Plexiglas doors leading to the choice compartments lowered. After 5 min the doors were opened, allowing unrestricted access to all 4 chambers for 10 min. Animals were scored in an adjacent room from a video monitor. During each trial, one of five observers trained to a reliability criterion of .95 (Pearson product-moment correlation) scored the amount of time that the subjects spent in each choice compartment.
The data from each preference test were analyzed using within-subject two-way analyses of variance, with the factors being trial number (Trial 1 versus Trial 2), and stimulus animal choice. Duncan New Multiple-Range analyses (Duncan, 1955) were conducted on significant main effects. Further analyses were performed correlating age of subject at maternal separation, length of time in peer group, and age at time of test, with duration of chamber entries for each test session.
Figure 1: Social choices in Tests 1 and 2. Values represent time spent next to each stimulus animal's chamber. FFG = familiar aged female from group; UFG = unfamiliar aged female; UF = unfamiliar young adult female; MA = subject's mother.
Results
There were no significant correlations between age of separation from mother, length of time between maternal separation and placement into the social group, or age at time of testing, with choices in the preference tests. There was a significant main effect of stimulus animal in all three preference tests. In both Test 1 (F [3,12] = 11.17, p < .001) and Test 2 (F [3,9] = 34.98, p < .001), Duncan analyses revealed that the monkeys spent significantly more time next to the familiar aged adult female than next to any other stimulus animal (p < .01 for all comparisons). The social choices in Tests 1 and 2 are depicted in Figure 1.
For Test 3 (F [3,12] = 7.73, p < .005), Duncan tests revealed that the means for the two familiar aged female adult cagemates, while not differing significantly from each other, differed significantly from the means for the other two stimulus animal choices (p < .05 for female A, and p < .01 for female B). Figure 2 depicts the monkeys' social choices in Test 3.
Figure 2: Social choices in Test 3. Values represent time spent next to each stimulus animal's chamber. FFGA = familiar cagemate aged adult female A; FFGB = familiar cagemate aged adult female B; UF = unfamiliar young adult female; MA = subject's mother.
Discussion
Results of this study indicate that, under mildly stressful conditions, yearling rhesus macaques preferred an adult female with whom they were currently living to their own mothers, who had been their original sources of attachment. These findings extend the findings of Mason and Kenney (1974) by indicating that attachment in mother-reared rhesus monkeys, as well as those raised by animate and inanimate surrogates, can be redirected following separation from the original attachment object. The data also suggest that an infant's attachment to its mother may not be maintained following a lengthy period of separation.
One possible explanation of these results involves a conditioning model of attachment. This model proposes that an infant will become attached to any object to which it is in proximity, especially if the object is associated with primary or secondary reinforcers (Hoffman & Ratner, 1973; Gewirtz, 1976). According to this model, infant-mother attachment develops because mothers provide many important primary reinforcers for infants. One prediction based on this model is that, following separation from mother, an infant can form an attachment to another animal, if this animal either constitutes a source of reinforcement directly or possesses stimulus attributes of the mother.
The finding that the infants showed no preference for their mothers above even an unfamiliar adult female was unexpected. One possible explanation, consistent with the conditioning model, is that the attachment bond underwent an extinction process in the absence of the attachment object. This may account for the ease with which the infants "forgot" their mothers. These results suggest that continual interaction may be necessary for the maintenance of the mother-infant attachment bond.
An alternate explanation for the results is that the juveniles were simply expressing a preference to approach a familiar, rather than an unfamiliar, animal. This is a less likely explanation for two reasons. First, when allowed a choice between the two familiar adult female cagemates, a borderline significant (p < .10) preference arose for one of them. The preferred female was found to show more positive social interactions with the subjects than the less-preferred female. In addition, the subjects did not prefer their mothers, which were presumably familiar animals, to the unfamiliar stimulus females.
These results indicate that weanling infants can form strong attachment relationships with new, adult cagemates following rehousing into a social group containing adults and age-matched peers. The results of this study suggest that this housing procedure can provide for the psychological needs of the weanling monkeys. Housing nonreproductive adults with large groups of weanlings appears to be an effective method of environmental enrichment, proving beneficial for all animals involved.
References
Berman, C. M. (1982). The social development of an orphaned rhesus infant on Cayo Santiago: Male care, foster mother-orphan interaction and peer interaction. American Journal of Primatology, 3, 131-141.
Duncan, D. B. (1955). Multiple range and multiple F tests. Biometrika, 11, 1-41.
Gewirtz, J. L. (1976). The attachment acquisition process as evidenced in the maternal conditioning of cued infant responding (particularly crying). Human Development, 19, 143-155.
Harlow, H. F. (1958). The nature of love. American Psychologist, 13, 637-685.
Harlow, H. F. & Harlow, M. K. (1965). The affectional systems. In A. M. Schrier, H. F. Harlow, & F. Stollnitz (Eds.), Behavior of Nonhuman Primates (pp. 287-355). New York: Academic Press.
Hoffman, H. S. & Ratner, A. M. (1973). A reinforcement model of imprinting: Implications for socialization in monkeys and men. Psychological Review, 80, 527-544.
Mason, W. A. & Capitanio, J. P. (1988) Formation and expression of filial attachment in rhesus monkeys raised with living and inanimate mother substitutes. Developmental Psychobiology, 21, 401-430.
Mason, W. A. & Kenney, M. D. (1974). Redirection of filial attachments in rhesus monkeys: Dogs as mother surrogates. Science, 183, 1209-1211.
Sackett, G. P. (1970). Unlearned responses, differential rearing experiences and the development of social attachments by rhesus monkeys. In L. Rosenblum (Ed.), Primate Behavior: Developments in Field and Laboratory Research (pp. 112-140, Vol. 1). New York: Academic Press.
-------------------------------------------------------------------
First author's current address: Primate Facility 7-930T,
Department of Psychiatry and Behavioral Sciences, Stanford University
School of Medicine, Stanford, CA 94305-5095.
The authors wish to thank the following: R. H. Beinert, W.
Hopkins, M. Kreisler, H. Leroy, L. Marra, and C. Ripp, Jr. This study
was supported in part by NIMH grant MH-11894.
* * *
Charles Oxnard University of Western Australia
Everyone knows today that captive primates should receive a diet containing adequate vitamin B12 or else vitamin B12 supplements. It is possible, however, that we may have forgotten why this is so, which primates it involves, and what happens if we fail to remember (Oxnard, 1987).
First, we must remind ourselves that vitamin B12 is not a regular "B" vitamin. Dietary supplements that contain B vitamins do not contain B12 unless it has been specially added. The B vitamins are generally available from extracts such as yeast. Vitamin B12 is not found in yeast and other vegetable products but in foodstuffs containing animal products (and also, as it happens, in earth, in feces, and in sewage sludge as a result of the activity of microorganisms).
Vitamin B12 enters the animal body through absorption in the jejunum and upper ileum. As a result it is necessary that it 1) be present in the diet as a result of ingesting animal products as in omnivores and carnivores, (2) be present as a result of being manufactured in the stomach through the activities of vitamin B&s'12 producing microorganisms in those creatures such as cows that possess the appropriate flora in a multilobed stomach, or (3) be present through ingestion of vitamin B12 produced by microorganisms in the colon when coprophagy allows colonic contents (which contain large quantities of microorganismally produced vitamin B12) to reach the jejunum and upper ileum in creatures such as rabbits.
Vitamin B12 deficiency can occur in many ways, but one of the commonest is through a dietary deficiency in those species that are mainly (but not totally) herbivorous, and that have no special mechanisms for getting microorganismally manufactured vitamin B12to the absorptive site in the small intestine.
Implications for Primates
This has implications for primates (Oxnard, 1966, 1969). Thus, colobines probably never develop vitamin B12 deficiency because of the production of the vitamin in their multilobed stomachs. Primates that display coprophagy as a behavioral disturbance (e.g., a chimpanzee at the London Zoological Society Gardens many years ago) also never become deficient whatever the diet. And certain prosimians probably never develop deficiency because their dietary requirements for insects or other animal foodstuffs (which contain large amounts of vitamin B12) are so great that they do not thrive in their absence.
The remaining nonhuman primates, those that are primarily herbivorous, are at risk of developing the deficiency. Of course, it is known that most eat small amounts of animal products. It is now well documented that many monkeys and apes ingest insects, birds' eggs, even birds and small animals when they can catch them. It is even recorded that some primates (e.g., baboons) catch and eat larger animals on occasion. Nevertheless, in contrast with humans, animal items form only a small proportion of the diet. Frequently only some of the animals (e.g., large males and favored females) ever actually receive the animal products. At certain times of year all animals may be confined to vegetarian foodstuffs. As a result, it is these primarily herbivorous primates that are at risk of developing hypovitaminosis B12.
on entry 6 months 12 months 18 months 24 months mean range mean range mean range mean range mean range 210 110-480 120 80-240 85 60-150 80 50-130 35 <20-70
Table 1. Vitamin B12 levels in captive rhesus monkeys (milli-milli-gm/ml serum).
If such species are placed upon a solely vegetarian diet in captivity, deficiency of the vitamin develops within 9 to 18 months (Oxnard, 1964). This is well documented for rhesus monkeys (Macaca mulatta, Table 1) but there is also evidence that it occurs in various other primates, but not all (Table 2).
Species Recently Species Long
Captive Captive
----------------------------------------------------
Erythrocebus 180 Erythrocebus 100
Erythrocebus 410 Erythrocebus 64
Papio 424 Papio 50
Cercopithecus 420 Papio 100
Cercopithecus 700 Cercopithecus *310
Mandrillus 228 Cercopithecus *280
Presbytis 980 Cercocebus 40
Callithrix 450 Aotus 100
Tupaia glis <4000 Macaca nemestrina 116
Galago demidovi 1680 Macaca nemestrina 62
Galago crassicaudatus <4000 Macaca maura 156
Perodicticus potto 2000 Pan 120
Loris tardigradus 1840 Pan **1100
Nycticebus coucang 1210 Hylobates 34
. . .* fed meal worms . . .** developed coprophagous habits
Table 2. Vitamin B12 levels in other primates (milli-milli-gm/ml serum).
How common is this deficiency? And what are its effects?
The deficiency occurs in every animal that receives a vegetarian diet. This was so even in a colony so large as to average around 300 rhesus monkeys. It was recorded as serum levels of vitamin B12 below 200 picograms/ml. The deficiency does not occur in animals that are fed "monkey chows" because most of these contain animal by-products stuffed full of vitamin B12, and also most are supplemented with additional vitamin B12.
We cannot easily see the effects of the deficiency unless we are aware of the possibility. Overt signs of long-standing and severe deficiency include the development of paralysis of the hind limbs and tail leading to death (Oxnard & Smith, 1966). But this is rare, occurring in fewer than half-a-dozen rhesus monkeys in a colony of 300 over a period of 5 years.
Careful clinical observation and examination of apparently normal animals, however, can reveal signs of deficiency (e.g., weakness and reduced reflexes prior to overt paralysis). Histopathological examinations of otherwise normal animals demonstrate peripheral and central neurological lesions. The study of other systems (e.g., the mouth, the eye, growth, reproduction) also shows abnormalities. But these were only obvious when they were sought, and the most surprising feature of the condition is that the fact that pathological lesions were widespread and quite severe in many animals that appeared, superficially, to be quite normal.
There are sufficient differences from the human condition of hypovitaminosis B12 that reliance upon the human analogy is misleading. For example, because stores of vitamin B12 in humans are so large, a dietary deficiency may not produce damage in humans for many, many years; indeed, it may never produce overt signs and symptoms. In contrast, in the particular nonhuman primates mentioned above, a dietary deficiency produces the hypovitaminosis within a few months.
The human condition is most often revealed through a very obvious macrocytic and megaloblastic anemia. This cannot be recognized in deficient nonhuman primates, though careful examination of a sample of animals before and after treatment does show a statistically significant increase in hemoglobin and reduction in mean corpuscular volume. But the changes are so slight that they are only revealed in before-and-after comparison of a group of animals that are deficient and then treated. They are not revealed through study of the differences between separate deficient and nondeficient groups (Oxnard, Smith, & Torres, 1970).
The human condition is often noticed through the development of oral mucous membrane lesions (e.g., angular cheilosis and glossitis). These are also present in deficient rhesus monkeys but are difficult to distinguish without careful comparative examination (Rose & Oxnard, unpublished results). A smooth, shiny tongue (glossitis) can be recognized, but angular cheilosis [fissuring at the corners of the mouth -- Ed.] is hard to separate from the trauma of handling.
Nervous System Lesions
The principal findings in nonhuman primates (Oxnard & Smith, 1966; Torres, Smith & Torres, 1971) are the nervous system changes (and these are among the rarest of the lesions in humans). Undoubtedly, could the animals complain, they would speak of "pins and needles" in their hindlimbs and tails. And certainly, severely affected animals often sit with their feet in inverted positions so that the plantar surfaces are not in contact with the cage floor. Again, severely affected animals sometimes have self-inflicted wounds on the feet and tail, possibly evidence of irritation there. And neurological examination in severely affected animals reveals evidence of disturbed sensory and motor function and changed reflexes in the hindlimbs and tail. But these are not readily elicited; it is just difficult to perform neurological examinations on captive monkeys. Certainly, most of the apparently normal animals (the great majority) display no obvious signs. Nothwithstanding this, lesions may be found throughout the entire nervous system, even in apparently normal animals.
Figure 1: Low (x17) and high (x120) power micrographs of demeylinating lesions in the spinal cord showing spongiform degeneration. Luxol fast blue - cresyl violet.
Figure 2: Low (x8.5) and high (x120) power micrographs of demyelinating lesions in the cerebrum showing spongiform degeneration. Luxol fast blue - cresyl violet.
In the peripheral nerves there is marked demyelination. Though this was first thought to be due to dying back of fibers (compare cross sections of distal and proximal sciatic nerves, e.g., Oxnard & Smith, 1966), further study of individual teased fibers showed that the initial lesions were due to segmental demyelination (e.g., Torres, Smith, & Oxnard, 1971).
In the spinal cord, patchy demyelination is readily observed in the white matter, especially the dorsal and lateral columns. When very severe it results in a spongiform degeneration because of the loss of large numbers of myelinated fibers. Similar lesions occur throughout the rest of the central nervous system, especially in the white matter of the cerebrum.
Optic lesions are especially interesting (Hind, 1966), the more so as the experimental use of so many primates is bound up with abilities of the animals to see well. Thus there are lesions that are obvious upon opthalmoscopy (e.g., flattening of the disc) and these are mirrored in histopathological changes in the retina. The demyelination found in the rest of the central nervous system is especially evident in the optic nerve, the optic chiasma, and various other parts of the optic system.
Other effects of deficiency
In addition to the neurological pathology, there are a series of other long term changes that are especially important in keeping normal healthy animals in captivity.
Problems with fertility are referable to the deficiency (Oxnard, 1964). Many of the animals that were deficient, though they did sometimes have infants, had them far less frequently than expected given the breeding schedule. They did not readily become pregnant. Their cycles seemed at first sight irregular, but when the cycles were inspected carefully over a period of several years, it became evident that the irregularity actually consisted of double and triple length cycles interspersed among normal length cycles. This may mean that the animals were becoming pregnant but were subsequently undergoing resorption of the conceptus products at a fairly early stage of pregnancy. It is certainly known that the presence of vitamin B12in the blastocyst is necessary for continuation of normal pregnancy in the rabbit.
Attempts were also made to obtain information about the vitamin B12 content of semen. It is known that adequate vitamin B12 is necessary for normal semen effectiveness in cattle (vitamin B12 must be added to the medium for artificial insemination). However, we never managed to determine semen vitamin B12 levels satisfactorily in rhesus monkeys, though we did make attempts.
It does seem clear, however, that hypovitaminosis B12is implicated in reduced fertility in deficient captive monkeys. In addition, it is apparent that even when the animals do succeed in becoming pregnant, the ratios of serum levels of vitamin B12in deficient mothers and their infants was rather different from those in non-deficient mothers and infants. Evidence suggests that this may occur in the field situation if the mothers are low enough on the totem pole that they do not receive a large enough share of animal foodstuffs, or if the time of year is such that animal foodstuffs are not available for any animal. Certainly the fetus "parasitizes" the mother for vitamin B12 so that deficiency during pregnancy is rather readily produced even under field conditions. To some degree this latter is self-correcting because many primate mothers eat the placenta (which is loaded with intra-cellular vitamin B12).
Again, after such births did occur in the colony, it was evident that, once the infants were weaned, a growth deficit gradually occurred. This was rather readily corrected by treatment with the vitamin. Indeed, the degree of interference with growth rate was so great that treatment increased it by between between 40 and 200 percent (Flinn & Oxnard, 1966)!
Finally, some primates have the reputation of being, alternately, morose and apathetic in captivity. It is one of the features that makes them difficult to handle. Some of this may be a manifestation of vitamin B12 deficiency. Deficient humans feel much better within a very short time of receiving therapy; undoubtedly this must be an immediately reversible condition of brain metabolism. It is highly likely that something similar occurs with captive monkeys. Their apathy and moroseness, the difficulties in handling them, disappear very quickly after treatment if the original deficiency has not been of long duration (Oxnard & Smith, 1967).
Incidence of the Condition
Given the widespread use of monkey "chows" which contain more than adequate amounts of vitamin B12, it is unlikely today that most primate colonies will see major evidence of the deficiency. Nevertheless, when it does occur, it may affect many animals yet be hidden for many months, indeed, for years, to casual observation. This is best seen from the incidence of lesions in the animals that we examined (Table 3).
Group Total Number Number Number with lesions
number of paralyzed without Peripheral Spinal Post. Cerebral
of animals lesions nerve cord root & lesions
ganglion
deficient 12 1 0 10 12 3 3
deficient,
then
treated 14 4 2 11 11 6 2
recently
captive 17 0 13 3 2 1 0
Table 3. Incidence of neurohistopathology.
Thus, 100% of the very small number of paralyzed animals showed severe lesions throughout the nervous system. In addition, however, 100% of apparently normal (though proven deficient through blood tests) animals also showed neuropathological lesions. And even a significant proportion (20%) of animals that were recently received from the field situation and which, therefore, would not have been thought deficient, showed some lesions. These last were most common in pregnant animals whose fetuses had, presumably, been taking most of what little vitamin B12was available from the mother. But lesions are also found in a few other animals, which suggests that this may be a risk to any nonhuman primate in the field, depending upon what foods are available at particular seasons of the year.
Conclusions
One conclusion from these studies relates to the importance of continually assessing apparently healthy animals. They may not be as healthy as they appear.
A second conclusion relates to the continuous need to examine the habits of husbandry that may have become ingrained over the years. These habits may not be as good as they appear; they may actually hide the problem.
A third requires that we critically reexamine the older research results that stemmed from primate colonies of yesteryear. Many of them were made upon apparently normal animals deficient in vitamin B12. And as many of them relate to broad matters like growth, reproduction, behavior, and the nervous system, the possible existence of the deficiency may be critical in their reevaluation.
But a final conclusion is of a different order. It is entirely possible that many nonhuman primates have been neurologically constrained (in an evolutionary sense) by a basic herbivorous diet that provided a barely adequate vitamin B12 milieu for the nervous system. Such creatures may have been meandering along for millenia on the verge of neurological deficiency, as it were. It may well be that it was the development of more frequent meat eating in early prehuman ancestors who took up first scavenging and later hunting that produced a new metabolic environment for neurological evolution. Is it possible that this was at least one of the factors that allowed the progenitor of the human brain to realize its special potential for growth in size, complexity, and function? This is a wild speculation. It just might be true.
References
Flinn, R. M. & Oxnard, C. E. (1966). The relationship between growth and the administration of cyanocobalamia in the rhesus monkey. Folia Primatologica, 4, 432-437.
Hind, V. M. D. (1969). Degeneration of the peripheral visual pathway of captive monkeys. Unpublished Ph.D. thesis, University of Birmingham.
Oxnard, C. E. (1964). Some variations in the amount of vitamin B12 in the serum of the rhesus monkey. Nature, 201, 1188-1191.
Oxnard, C. E. (1966). Vitamin B12 nutrition in some primates in captivity. Folia Primatologica, 4, 424-431.
Oxnard, C. E. (1969). A note on the ruminant-like digestion of langurs. Laboratory Primate Newsletter, 8[2], 24-25.
Oxnard, C. E. (1987). Apparent health, hidden disease: Vitamin B12 in captive primates. Australian Primatologist, 3[1], 28.
Oxnard, C. E. & Smith, W. T. (1966). Neurological degeneration and reduced serum vitamin B12 levels in captive monkeys. Nature, 210, 507-509.
Oxnard, C. E. & Smith, W. T. (1967). Vitamin-B12 deficiency in psychiatry. Lancet, 1, 161.
Oxnard, C. E., Smith, W. T., & Torres, N. (1970). Vitamin B12 deficiency in captive monkeys and its effect on the nervous system and the blood. Laboratory Animals, 4, 1-12.
Torres, N., Smith, W. T., & Oxnard, C. E. (1971). Peripheral neuropathy associated with vitamin-B12 deficiency in captive monkeys. Journal of Pathology, 105, 125-146.
-------------------------------------------------------------------
Author's address: Department of Anatomy and Human
Biology, University of Western Australia, Nedlands, WA 6009, Australia.
Reprinted with permission from Australian Primatology, 1989,
4[1], 2-5, with additions by the author.
* * *
Travel Funds Available
The International Primatological Society has applied for funds from the National Science Foundation, through its International Travel Grant Program, to help support travel expenses of U.S. participants in the IPS's Congress scheduled for July 18-24, 1990, in Japan. If received by IPS, the grant will be $500 per person. Use of U.S. carriers is required.
Individuals wishing to be considered for these travel awards should prepare an application that contains (a) their name, Social Security number, title, institutional affiliation, and address; (b) a brief outline, NOT TO EXCEED ONE PAGE, of their proposed participation in the Congress (and/or in Pre- and Post-Congress activities that are part of the Congress's overall scheduled program) and of their qualifications for such participation; and (c) when an NSF travel grant was last received. These applications should be sent to: Dr. W. Richard Dukelow, President IPS, Endocrine Research Center, Michigan State University, East Lansing, MI 48824. The deadline for receipt of application is December 1, 1989.
All applications will be reviewed by an Ad Hoc Committee of IPS, and all awards will be made primarily on the basis of proposed participation in the Congress's overall scheduled program. Decisions regarding awards will be made in accordance with the Civil Rights Act of 1964 and the implementing regulations prohibiting discrimination against any person on the grounds of race, color, religion, or national origin.
Travel Agencies Offer Services
The following travel agencies have contacted the IPS executive and expressed a desire to deal with members wishing to attend the Japanese meetings in an effort to arrange the lowest cost transportation.
As was the practice for past meetings, IPS makes no formal appointment of an "official" travel agent, but just makes these names and numbers available to you for your convenience.
1. E. N. Tours, Columbus, OH [800-843-4978].
2. Getaway Travel Tours, Lansing, MI (group department)
[800-541-7679].
* * *
Last year Dr. Jane Bennett, of the National Parks and Wildlife Office of Sarawak, East Malaysia, sent a letter to eighteen zoos, appealing for help in finding homes for surplus gibbons and orang-utans. This letter was reprinted this year in Australian Primatology (4 [1], 6-8). Dr. Bennett's office accepts confiscated animals of protected species which are being kept illegally, and attempts to rehabilitate them at its Wildlife Rehabilitation Centre at Semengok Forest Reserve. Many of the animals received are Bornean gibbons (Hylobates muelleri) and orang-utans (Pongo pygmaeus). Release of the rehabilitated animals has not been successful for the most part, for various reasons, including crowding in the available reserves, the difficulty of controlling hunting, and the inability to sufficiently rehabilitate animals which have been in captivity for a long time. Dr. Bennett is appealing to the primatology community in general for advice and help in finding appropriate homes for some of these animals. Her address is: Dr. Jane Bennett, National Parks and Wildlife Office, First Floor, STIDC Building, 93050 Kuching, Sarawak, East Malaysia.
* * *
Evelyn Halloren, Eluned C. Price, and W. C. McGrew
University of Stirling
Introduction
Primatologists seeking longitudinal data on marmosets and tamarins (Callitrichidae) face a special problem: offspring are born in litters. Whereas other simians typically produce only one infant at a time, callitrichids produce twins. Thus, any data collection on individuals requires some kind of marking to differentiate them, preferably from birth onwards.
Standard methods of marking incur costs for the subject, which may suffer stress (e.g., being handled, temporarily isolated), pain (e.g., tattooing, freeze-branding), or mutilation (e.g., ear-notching, toe-clipping) from the marking process. The least distressing technique seems to be dye-marking of the subject's hair, although we know of no prior research on this topic (see Follow-up section). But even dye-marking usually involves capture (by net or gloved hand) and handling, which may be traumatic.
In this note, we describe a recently developed technique for the non-invasive marking of neonatal cotton-top tamarins (Saguinus oedipus). It should generalize to other callitrichids, who carry dependent young dorsally, making them more accessible than offspring carried ventrally. It is minimally disturbing for the infants and for their family members, as well as for the human beings involved in husbandry and research.
The Stirling colony of cotton-top tamarins dates from May, 1982, and comprises 8 breeding families, numbering overall about 65 individuals on average (Price & McGrew, 1989). Most families are nuclear, with several sets of offspring present, including full adults, who cooperate in the rearing of their younger siblings. All research is non-invasive and most data collection is observational. The monkeys live in spacious indoor-outdoor housing that is as naturalistic as possible. Monkeys are rarely touched by humans and move about the colony through elevated "tunnels" of plastic ducting up to 30m in length.
Old Marking Techniques
Until August, 1986, infants were caught about 6 - 8 weeks (range: 4 - 13) after birth when they were moving independently. They were marked by dyeing the crest of one of the twins with picric acid; the resulting bright yellow mark easily distinguished that individual from its unmarked co-twin. (Singleton offspring were left unmarked. In no case did all 3 members of any triplet litter survive for more than 2 weeks, so the resulting pair were marked as twins.)
From August, 1986 to December, 1988 a similar technique was used when infants were about 4 days old. The infants were caught in a nylon fishing net while being carried by another family member; this took about 15 minutes. Sometimes 2 or 3 persons were needed for this, depending on the family's housing arrangement and make-up. For a mature family in a large walk-in room, one technician sometimes had to fend off the family's mobbing while one or two other persons pursued and netted the carrier and infant. In other cases, the ducting was used to isolate the carrier and infant from the rest of the family.
After capture, the infant had to be separated from its carrier, which had to be restrained. This required 3 persons. After dyeing, weighing, and sexing the infant, it and its caretaker were returned to the family's home-cage. This was done in two ways: One was to offer the infant to its caretaker in a small transport cage. Once it was being carried again, both were released into the home-cage. The other was first to return the caretaker to the family, then place the transport cage, containing the infant, next to the home-cage. When the former was opened, a family member rushed in to retrieve the infant and both were returned to the family.
The technique had several drawbacks: (1) During capture the collective anti-predator response of the monkeys included biting the humans. (2) When netted, the carrier often attacked the infant being carried on its back, and in 4 of 16 cases, this led to injury. Once a panicky carrier's bite caused permanent facial damage to an infant. (We interpret this apparently paradoxical behavior as a desperate, self-saving attempt to divest itself of the encumbrance in order to escape a predator. All 4 cases were from the only 2 families with wild-born parents). (3) Finally one fatal incident and several illnesses caused us to question the procedure. During re-introduction, an infant fell from its carrier to the floor, and 5 agitated family members tried to retrieve it at once. In the confusion, the caretakers fought each other for possession of the infant, and it was injured so badly that it had to be euthanitized. Of the 46 infants marked at 6-8 weeks, none had a fatal neonatal illness; of the 46 neonates marked at about 4 days, 5 died of respiratory illness within a few days, while none of the unmarked neonates died during this period.
New Marking Technique
From January, 1989, we have used a non-invasive method of marking which entails 4 stages of training of about 15 minutes per day. (1) 2 - 4 days. The marker enters the cage and feeds desirable tidbits by hand to as many family members as possible. We use Farley's Rusk (Farley's Health Products, Nottingham, England), but presumably any treat will do. In the breast pocket of the marker's lab coat is a 15.5-cm long plastic vial containing a 13.5-cm long cotton-tipped swab. .br;(2) 2 - 4 days. While feeding the tidbits one by one from one hand, the marker removes the swab with the other, and holds it progressively closer until it is about 5 cm from the monkey taking the treat. .br;(3) 7 - 10+ days. While holding in one hand the tidbit being nibbled by the monkey (for about 5 sec), the marker uses the other hand gently to touch and then to stroke the monkey's head and shoulders with the swab. This is repeated for each potential carrier in the group. .br;(4) On the morning after the birth of infants (thus as soon as 14 hours post-partum; see Price, 1989), the procedure in (3) is repeated, except that now the vial contains dye, and the swab is used delicately to "paint" the sagittal hair of the neonate's head. This may be repeated until the mark is clearly visible.
Family Days of Site Comments
Training
----------------------------------------------------------
Roxanne 11 Homecage 2 of 3 triplets marked
on eldest brother
Elsa 40 Homecage Each twin carried by
co-eldest brother
Delaware 17 Homecage Both twins on father
Erica 0 Test cage Both twins on father
Jille 1 Outdoor area 1 of 2 triplets marked on
primiparous mother, but she
resisted marking of other;
3rd triplet on father, who
also disallowed marking
Table. Details of cases of new marking technique.
So far, the new method has enabled us to mark infants within 24 hours of birth in 5 families (See Table). Surprisingly, the length of pre-training seems to be highly variable, so that in one case, none was needed. We suspect that this will prove to be a multi-variate problem, e.g., when hand-feeding is already a normal practice in a colony, then Stage 1 is largely superfluous; when parents are captive-born and multiparous, they may be tolerant of close interaction with humans, thus accelerating Stages 2 and 3, etc. The whole process can be done by one person, so it is economical of time and effort, but mainly it is free of stress for monkey and human.
Follow-up
Unnatural marking may disadvantage an infant, so we have sought to test this. In all 48 pairs of infants marked, one was randomly assigned to be dyed. Three measures of developmental progress were noted: survivorship, weight-gain, and age of emigration. Two of the unmarked infants have died before the age of 12 months versus 5 of the marked infants. At about 12 months of age, each subject is given an identity necklace and weighed. There was no difference in body-weights between unmarked and marked individuals: N = 36, mean = 482 +/- 38 grams versus N = 35, mean = 478 +/- 36 grams. Monkeys leave their natal families by eviction, removal, or death. When all such ways were pooled, there was no difference in age at departure between unmarked and marked subjects: N = 21, mean = 30 +/- 11 months, versus N = 19, mean = 31 +/- 13 months. Thus, dye-marking seems to make no difference to an infant's prospects.
References
Price, E. C. (1989). Parturition and perinatal behaviour in captive cotton-top tamarins (Saguinus (o.) oedipus). Manuscript submitted for publication.
Price, E. C. & McGrew, W. C. (1989). Cotton-top tamarins (Saguinus (o.) oedipus) in a semi-naturalistic captive colony. Manuscript submitted for publication.
-------------------------------------------------------------------
Authors' address: Scottish Primate Research Group,
Department of Psychology, University of Stirling, Stirling FK9 4LA,
Scotland.
We thank Ms. E. Moodie for helpful advice.
* * *
OJ in the Morning
What keeper hasn't thought how clever he or she is to hide a pill inside a gorilla's favorite food, only to find that the gorilla daintily eats the food -- and then spits out the pill! Even crushing the pill and smearing it over the food often will not work, as the exasperating animal will carefully rub it off. Quite by accident, I came across a solution to this problem that has worked very well with our seven gorillas [and may work with other primates -- Ed.].
We all know how suspicious gorillas are of new food items, so as an everyday part of their diet, I offer the gorillas, by spoon, a mixture of cottage cheese and orange juice. It sounds awful, but they love it. Then, when it is necessary to administer medication, whether it be liquid or solid, I simply mix it in with a small amount of the cottage cheese/OJ mixture. The texture of the cottage cheese hides any powder, and the color and taste of the orange juice masks the presence of the "foreign object." So far, none of our highly suspicious gorillas has caught on to the fact that there is sometimes something extra in this mixture. [Some medicines are not compatible with acids or milk products -- Ed.]
Fecal Sample Exchange
Once a week we collect a fecal sample for a reproductive study, and we have taught our gorillas to hand over a piece of their feces in exchange for peanuts, thus eliminating the necessity to shift an animal to obtain a sample. -- Marilyn Cole, Metro Toronto Zoo, Box 280, West Hill, Ontario M1E 4R5 Canada. Reprinted with permission from the Gorilla Gazette, 1989, 3[2], 11.
* * *
Lake Naivasha, Kenya
For investigators or study groups in animal behavior, biology, and conservation, the Elsamere Conservation Centre offers comfortable accomodations at a modest cost. Elsamere is located on the lakefront, a short drive from the new Hell's Gate National Park as well as several ranches that support wildlife including monkeys. Elsamere guest house accomodations with full board are approximately $14/day. For information, contact Ben or Lynette Hart [914-756-5345], or The Warden, Elsamere Conservation Centre, P.O. Box 4, Naivasha, Kenya.
Foundation for Field Research
Financial support for researchers is available for projects that can be funded and assisted by volunteer teams. The project must consist of basic field research. Proposals must be submitted at least a year in advance of the project dates, and should follow the requirements available upon request from the Foundation. For information contact the Foundation for Field Research, 787 South Grade Road, Alpine, CA 92001 [619-445-9264].
* * *
K. Bayne, S. Suomi, and B. Brown
National Institutes of Health
As standards for environmental enrichment become better defined, creative means of meeting the mandate for providing "an environment adequate to promote the psychological well-being" of laboratory nonhuman primate populations must be designed. One of the many forms of environmental enrichment the USDA is proposing for nonhuman primates involves installing a swing in the animal's primary enclosure (Federal Register, March 15, 1989). A variety of materials have been used to construct swings, including polyvinyl chloride (PVC) piping, bent metal rods, suspended milk crates (O'Neill, 1988), suspended platforms, and chains (although several cases of animal injuries involving chains have been reported to the authors).
Figure 1: Use of PVC tubing by rhesus monkeys in an indoor-outdoor run.
A swing for primates would ideally be sanitizable (i.e., remain unaffected by 180deg. F temperature of a cagewash); able to withstand potential damage from the primates themselves, and inexpensive. In our laboratory garden hoses have been used for swings in indoor/outdoor runs and in corn cribs (O'Neill, 1988). This type of swing, when attached at one or both ends to the top of the housing unit (see Figure 1), is readily utilized by rhesus monkeys (Macaca mulatta). Garden hose swings are quite inexpensive, but they are degraded by high temperatures and by the monkeys playing on them. We have subsequently utilized transparent polyvinyl chloride (PVC) flexible tubing (3/4" internal diameter) for swings in our indoor/outdoor runs and in single cages. This tubing is available in most hardware stores at a relatively low cost ($1.10/ft.; each single cage requires approximately 3 ft.); the tubing can go through a standard cage wash while still in the cage without being damaged, and appears to be impervious to nonhuman primate canine teeth. We have utilized this tubing in some of our cages for over a year without need for replacement. The monkeys use the tubing in their leaps across the runs, they hang from it, and some even sleep on it, when it is suspended by both ends. PVC tubing can easily be suspended by pushing an end between two bars at the top of the cage and then down between the next pair. No additional hardware is required to keep it firmly in place, since the spaces between the bars is slightly narrower than the tubing.
Figure 2: Schematic of method of attaching and filling PVC tube with banana pellets.
We have started placing PVC tubing in the cages of those monkeys that engage in self-abusive behaviors. Although our sample size of self-abusing monkeys is small to date (n = 3), we have had some success in arresting this behavior (two successes, with rhesus monkeys, and one failure, a cynomolgus monkey). We have also increased the attractiveness of this tubing in single cages by filling a large, loose knot tied in it with banana pellets (see Figure 2). Individually housed monkeys, using one finger, can reach up from the dangling end of the tubing and work the pellets down. This is a time-consuming activity for monkeys, and one that is also quite safe for facility personnel because the tubing may be filled from the top of the cage by cutting a small hole in the end projecting above the cage. We have not noted any significant increase in body weight, nor reduction of the animal's appetite with the provision of these food treats.
New materials, such as PVC flexible tubing, which are durable and sanitizable should be considered when choosing enrichment techniques. They enhance the animal's environment while still maintaining current sanitation standards.
Reference
O'Neill, P. (1988). Developing effective social and environment enrichment strategies for macaques in captive groups. Lab Animal, 17 [4], 23-36.
United States Department of Agriculture (1989). Animal Welfare proposed rules. Federal Register, 54, 10822-10954.
-------------------------------------------------------------------
First author's address: Department of Health & Human Services, NIH, Building 14D, Room 315, Bethesda, MD 20892.
* * *
Animal Facility Improvements for Small Research Programs
The Division of Research Resources (DRR) announces a competitive grant program to help institutions with small research programs using animals upgrade and develop their animal facilities. DRR anticipates that six to eight awards will be made Fiscal Year 1990. Institutional animal resource improvement grants are awarded to assist biomedical research and educational institutions in upgrading their animal facilities and developing a centralized animal care program. A major objective is to enable institutions to comply with the USDA Animal Welfare Act and DHHS policies on the care and treatment of animals. These awards are limited to Alterations and Renovations (A&R) to improve laboratory animal facilities and related major resource equipment, such as animal cages and washers.
To gain approval and support, the need for resource improvement, as well as a sound plan to meet the requirements of the Public Health Service Policy on Humane Care and Use of Laboratory Animals, must be presented and described in the context of the biomedical research training program of the institution.
Any domestic public, or private institution, organization or association with one or more research projects supported by the Public Health Service (PHS) and involving the use of animals is eligible to apply. The total PHS support in the last fiscal year for projects using animals must have been less than $500,000. Applicants are expected to develop a single proposal for campus-wide service. There will be a single receipt date of December 4, 1989. Applications received after this date will be returned.
Institutions may request major equipment items for their animal resources as well as funds for A&R. Support for new construction is not authorized. The award is limited to $100,000 for A&R and $150,000 for equipment. Matching funds are not required. Funds awarded for A&R may not be obligated until final architectural drawings, specifications, and updated cost estimates are received and approved by the DRR.
A copy of the complete RFA, which describes the research goals and scope, terms and conditions, review procedures and criteria, and method of applying, may be obtained by contacting: Director, Laboratory Animal Sciences Program, Division of Research Resources, 5333 Westbard Avenue, Room 853, Bethesda, Maryland 20892 [301-496-5175].
L. S. B. Leakey Foundation
The L. S. B. Leakey Foundation supports research related to human origins, behavior, and survival. Priority is given to studies in the areas of human paleontology, archaeology, and environmental studies of the Miocene, Pliocene, and Pleistocene; the behavior of the Great Apes and other Old World primate species; and the ecology and adaptation of living hunter-gatherers. Eligibility is limited to applicants who are doctoral candidates or engaged in post-doctoral research. Potential applicants are encouraged to submit a Petition for Grant Application. On invitation of the Science and Grants Committee, projects falling within the range of priorities of the Foundation will be asked to submit a formal application. Deadline for submission of the formal application is December 1. For application forms contact: Kathleen Galvin, Grants Officer, L. S. B. Leakey Foundation Center 1-7, Pasadena, CA 91125.
Office of Naval Research
The Office of Naval Research (ONR) has recently described new opportunities for psychological research support through its Life Sciences Directorate. Research in the Perceptual Science Program emphasizes issues of perceptual elements, their representations and transformation in the domains of vision, audition, touch and manipulation, multi-modal integration and the control of motor function. The research goal of the Biological Intelligence Program is to identify the organization, structural bases, and operational algorithms that characterize information-processing networks within neural systems. An Accelerated Research Initiative in Biological Intelligence includes research to understand processes by which life stresses come to influence the functions of the immune system and susceptibility to illness and the development of animal models of visual processing.
The first step in approaching ONR for research support is to make personal contact with the program manager in order to verify mutual interests. The program manager for Perceptual Science is Dr. Harold Hawkins [202-696-4323], and for Biological Intelligence, Dr. Joel Davis [202-696-4744]. Assuming there is mutual interest, a pre-proposal and, eventually, a proposal will be required.
Research Training Groups Program
The Directorate for Biological, Behavioral and Social Sciences (BBS) of the National Science Foundation (NSF), in recognition of the need to encourage multidisciplinary training and enhance diversity in the next generation of scientists, will offer Research Training Group (RTG) awards, to provide groups of investigators with funds to foster multidisciplinary, research-based training and education. It will encourage thematic, multidisciplinary training opportunities in excellent research environments; and provide support for training activities and for the acquisition or development of state-of-the-art shared instruments and special purpose materials to be used in research.
An RTG should focus on a major research theme for which multidisciplinary efforts hold promise. An important component of the effort must include education and training in areas of research funded by BBS. However, disciplines funded by other NSF directorates may be involved. All RTGs must: 1) Be based in a U.S. academic institution that grants the Ph.D. 2) Create new or enhanced multidisciplinary educational and research opportunities at the institution. 3) Be directed by a principal investigator, with an appropriate number of other faculty members who will assist in management and in the educational and research programs of the RTG. 4) Have secured resource commitments and financial contributions from the home institution and any institutional collaborators. 5) Have well-formulated plans for recruitment of students and fellows, with provisions that will increase participation of members of groups underrepresented in science.
Awards will be made for five years. Annual budgets are expected to average $250,000 (including direct and indirect costs), and up to $250,000 may be requested for specific items of special purpose research materials and equipment. BBS expects that $4 Million of FY90 funds will be available for the first year of this program.
The deadline for receipt of proposals is March 1, 1990. Inquiries regarding the RTG program should be directed to Division of Instrumentation and Resources, Room 312, NSF, Washington, DC 20550 [202-357-9880; EMAIL: [email protected]]
* * *
Viktor Reinhardt
University of Wisconsin
There is growing consensus that pair-housing of compatible conspecifics is an effective way to enrich the environment of caged primates. One concern, however, has been raised repeatedly: If one companion dies or is permanently assigned to an investigation prohibiting social housing, it may be problematic to house the remaining partner with a new conspecific. The present report addresses this question.
At the Wisconsin Regional Primate Research Center the majority of caged adult (>5 years old) rhesus monkeys are pair-housed. It is the Center's policy to ensure continual social enrichment for these animals whenever possible. If an adult individual loses its cage-companion we have tested 2 options to quickly provide new companionship: 1) Pairing the animal without preliminaries with a naturally weaned 1- to 1.5-year-old surplus infant from the breeding colony; and 2) Pairing the animal without preliminaries with another adult monkey which has also lost its adult companion.
During the last 2 years, 31 adult rhesus monkeys lost their compatible cage-mates. All these animals were successfully housed with new partners. Seven males and females were paired following option 1. Twenty-four females were paired following option 2. None of the new pairings were associated with serious injuries. This was expected when animals were paired according to option 1, as this procedure has proven to be safe on many other occasions. Following option 2, however, was an experiment that implied a certain risk as the new adult partners were strangers to each other. To the author's surprise, the 24 unfamiliarized females established clear dominance-subordination relationships (unidirectional yielding and/or fear-grinning) in each of the 12 pairs within the first 3 minutes after pairing without any serious fighting. The establishment of rank relationships was associated with no apparent aggressive behavior at all in 5 pairs; it was associated with threatening (intended aggression leading to no physical contact) in 4 pairs, with biting in only 3 pairs (Table I). While biting, the animals inflicted no visible wounds on each other. Previous exposure to another adult companion apparently provided the animals with important social experiences necessary to adequately cope with the potentially dangerous situation inherent in option 2.
New Pair Aggressive acts New Pair Aggressive acts
performed during performed during
first hour after first hour after
pair formation pair formation
-----------------------------------------------------------
Jolly threatening Silk biting
Tina threatening Jessy none
Grabby threatening Nice none
Claudia none Dashi none
Nurse threatening Susi none
Mila none Ninni none
Floppy threatening Trude none
Mimmi none Horse biting
Berta biting Addy none
Wanda biting Clair none
Bald biting Gusti none
Pi none Tilla none
Table I: Aggressive encounters between previously paired strange adult female rhesus monkeys during the first hour after new pair formation.
Our attempts to re-pair adult caged rhesus monkeys that have lost their companions have consistently been successful and compatibility between the new companions (food sharing, no serious injury) was ascertained throughout follow-up periods of 3 to 24 months. This may encourage others to provide their animals continued social enrichment even in those cases of pairs which may have to be separated later.
-------------------------------------------------------------------
Author's address: Wisconsin Regional Primate Research Center, 1223 Capitol Court, Madison, WI 53715.
* * *
Animal Welfare Regulations
Parts 1 and 2 of the U.S. Department of Agriculture Animal Welfare Act (AWA) were published in final form on 31 August in the Federal Register (54, 36112-36163). The final rule shall become effective on October 30. Portions of the regulations which relate to information collection and record keeping will also become effective on that date, but must be approved by the Office of Management and Budget (OMB). If any portion of the paperwork requirements is disapproved by OMB, notice will be published in the Register prior to October 30.
The Animal Welfare regulations (Parts 1 and 2) have been significantly rewritten and reorganized. The tone of stated requirements is much improved in the final version, but major provisions are really unchanged. Although basic requirements for compliance remain the same, a number of very onerous reporting and record keeping requirements have been dropped. Language very similar to that of the U.S. Public Health Service Policy is used to describe requirements for research facilities which are common to both the Policy and the AWA regulations.
The Register indicates that Part 3, the standards by animal species, is also final, but this refers to existing Part 3 standards with only a minor change. The "veterinary care" section for each species is removed in favor of an overall statement of veterinary care requirements contained in Part 2. USDA/APHIS indicates Parts 1 and 2 can be fully implemented with the existing Part 3 Standards. The Department is continuing to review public comments and consider alternatives to the new standards proposed on 31 March 1989. The anticipated date for republication of the Part 3 proposals to implement exercise of dogs and psychological well-being of primates requirements is not known at this time.
The National Association for Biomedical Research (NABR) is carefully reviewing the final definitions and regulations. Their first reading indicates that the following provisions are still of concern:
Institutional Animal Care and Use Committees (IACUCS) will "review and approve, require modifications in (to secure approval), or withhold approval of those components of proposed activities related to the care and use of animals" as well as "proposed significant changes regarding the care and use of animals in ongoing activities."
Proposals to conduct an activity involving animals must contain the same five elements required by the PHS Policy: identification of species and number of animals, complete description of proposed use of animals, description of procedures to limit discomfort and pain to animals and any euthanasia method to be used. However the USDA will also require the following not mentioned in the PHS Policy: 1) Principal investigators must consider "alternatives to procedures that may cause more than momentary or slight pain or distress to the animals," and provide "a written, narrative description of the methods and sources used to determine that alternatives were not available." 2) Principal investigators must provide "written assurance that the activities do not unnecessarily duplicate previous experiments."
Research facility records and IACUC records including those related to review of proposed activities involving animals must be made available to APHIS inspectors for copying; therefore, this information would be available under the federal Freedom of Information Act. APHIS also claims authority to take photographs in facilities.
Other than the concerns stated above, many changes suggested by the research community have been made. Interested parties are encouraged to review the regulations thoroughly in anticipation of being in full compliance by 30 October. NABR would appreciate hearing about any questions and concerns. -- From the NABR Update, 1989, 10[22]. New Antimalarial Drugs
Countries in the developing world now have a new weapon against malaria. It is a drug called halofantrine, produced by Smith, Kline & French (SK&F). Some scientists fear, however, that when the manufacturer makes halofantrine available worldwide next year, indiscriminate use of it could induce resistance in the malarial parasites. The launch of the drug could also jeopardize an agreement between the World Health Organization (WHO) and Hoffmann-La Roche to limit the marketing of another drug for malaria, called mefloquine, to developed countries.
Chloroquine, the standard treatment for malaria since 1946, is now ineffective in many areas of the world because of the spread of chloroquine-resistant parasites that cause malaria. Mefloquine was first marketed in 1984 under the trade name Lariam, along with a combined drug, Fansidar, containing mefloquine and two other antimalarial drugs. Before launching mefloquine, Hoffmann-La Roche made an agreement with WHO to limit marketing of the single drug to developed countries. The aim of the agreement was to prevent resistance to the drug from developing through overuse. SK&F's plans for rapid worldwide distribution of halofantrine may now persuade Hoffmann-La Roche to reconsider its agreement with WHO rather than lose out on a massive market for their drug. -- From New Scientist, 19 November 1988, 23.
Tamarins and Cocoa
Maria Cristina Alves, a graduate student at the University of Florida's Center for Latin American Studies, described her survey of wildlife on cacao farms in Bahia, Brazil, in the Fall, 1988, issue of On the Edge, a publication of the Wildlife Preservation Trust. In southern Bahia cacao is traditionally grown by a system called cabrucagem, in which the subcanopy and understory from primary forests are replaced with young cacao shrubs. Since a certain number of canopy trees are left to provide shade essential to the cacao, a relative diversity from the original habitat is maintained. Some species were more abundant in the disturbed habitat, but the golden-headed lion tamarin, although sometimes present, seems to require the presence of a neighboring forest, even a small-sized one, for its survival. This holds true as well for the southern Bahian marmoset (Callithrix kuhii). Maintenance of intact forest patches within cacao plantations is accepted and already practiced by a few farmers. Reinforcing this practice would greatly benefit species like these callitrichids.
* * *
Behavioral Research
A Research Associate position is available, requiring a Ph.D (A.B.D. considered) in Psychology, Zoology, or Anthropology, and experience in behavioral research. The Associate will work directly under the Director of a breeding colony of chimpanzees (Pan troglodytes), and will be responsible for assistance with study design, analysis, and write-up; training and daily supervision of Staff Observers and Interns; maintenance of research records; and assisting with the collection and computer entry of behavioral data. The position requires a solid background in statistics; good organization, managerial, and communication skills; and an ability to work well with students and other investigators. Salary is negotiable; benefits are excellent. We are an equal opportunity employer. Send a letter of interest with requested salary, resumé, and three letters of reference to: Jo Fritz, Primate Foundation of Arizona, P.O. Box 86, Tempe, AZ 85280. Applications will be accepted until the position is filled. Note: All personnel must have a negative T.B. skin test and a negative Hepatitis B surface antigen test prior to employment.
Veterinarian
A position is open for a full-time staff veterinarian for our breeding facility, to coordinate preventative medicine programs, manage clinical cases, assist in complying with USDA and NIH regulations, develop staff health policies and training programs, write SOP's, and compile and publish valuable chimpanzee information. We have an excellent consulting staff in human and primate medicine, and offer good benefits. Candidates must have the D.V.M. degree, clinical experience, and a strong interest in primate medicine. Salary will be commensurate with experience. Applications will be accepted until the position is filled. Send a letter of interest, including requested salary, resumé, and three letters of recommendation to Jo Fritz at the address above.
* * *
The Library at the Wisconsin Regional Primate Research Center is encouraging primatologists to consider videotaping or audiotaping slide presentations about nonhuman primate topics. A talk, with all the work that goes into it, can have a life beyond its live presentation as part of the WRPRC visual lecture series, says Librarian Larry Jacobsen. To date the series includes tapes and informal slide-talks by WRPRC staff, affiliates and visiting scientists, but others would be welcome. For advice on how to tape your lecture for inclusion in the collection, please call Ray Hamel, Special Collections Librarian, at 608-263-3512. The library publishes a biannual audiovisual acquisitions newsletter and makes collection materials available for loan. For information on how to borrow materials, call the number listed above or write: Larry Jacobsen, Primate Center Library, 1220 Capitol Court, Madison, WI 53715-1299.
* * *
Lion-Tailed Macaque Symposium
The third international symposium on lion-tailed macaques will be held in San Diego during May, 1990. The program will feature workshops, paper sessions, and invited lectures, including a keynote speaker. The Zoological Society of San Diego, the hosts, invite those having valuable information on any aspect of captive management, reproduction, health, behavior, and exhibit design to consider presenting the results of their work at this meeting. For information, contact Don Lindburg, Center for the Reproduction of Endangered Species, San Diego Zoo, San Diego, CA 92112.
Australian Primate Society
The Eighth Annual Conference of the Australian Primate Society will be held in Sydney. Tentative dates are December 9-10, with registration on Friday, December 8. For more information, contact Andrew Phippard, Department of Renal Medicine, Royal Prince Alfred Hospital, Camperdown, NSW 2050, Australia.
* * *
An updated Directory will be published in the January, 1990, issue of the Laboratory Primate Newsletter. If you wish your program to be represented in this Directory, or to revise your present entry, please send us the necessary information, following the format shown here as closely as possible. Return the information as soon as possible, but not later than December 10, 1989, to the Laboratory Primate Newsletter, Psychology Department, Brown University, Providence, RI 02912. Please note that the Directory is not intended for post-doctoral programs, though any such sent to us will be listed separately.
* * *
CAUZ
The Consortium of Aquariums, Universities and Zoos (CAUZ) is a communication network of people interested in sharing ideas and collaborating on research and educational projects. CAUZ publishes an annual Directory, listing the people in the Network, with information on their interests and on-going projects, which is available for $10 (payable in U.S. funds to C.S.U.N. Trust Fund) from Donna F. Hardy, Department of Psychology, California State University, Northridge, CA 91330. Dr. Hardy is the person to contact to be included in future Directories. CAUZ also collects, maintains, and distributes specialized bibliographies, in printed and electronic form. Those currently available which may be of interest to primatologists are on the golden lion tamarin, the mangabeys, Pongo pygmaeus, and the reintroduction of captive-bred animals into their native habitats. Printed bibliographies, and information on how to receive them in electronic form, are also available from Dr. Hardy. Bibliographies to be included in this collection may be submitted to Suzanne K. Braun, Indianapolis Zoo Library, 1200 West Washington Street, Indianapolis, IN 46222.
Lion-Tailed Macaque Newsletter
Lion-Tales: Lion-Tailed Macaque Newsletter, is published bi-annually by the Lion-Tailed Macaque SSP Propagation Group. It contains articles on care, reproduction, and exhibiting, as well as bibliographies. For information, write to the Editor, Helena Fitch-Snyder, Zoological Society of San Diego, Center for the Reproduction of Endangered Species, Box 551, San Diego, CA 92112.
Animal Transportation
The Animal Air Transportation Association, Inc. is an international organization made up of government representatives, airlines, animal shippers, and others involved with the shipment of live animals, no matter what mode of transport. The Association publishes a quarterly newsletter and holds an annual conference. For information, contact AATA, P.O. Box 863, Lemoore, CA 93245-0863 [209-935-2654].
Japanese Macaque Book
Several pioneering studies on Japanese macaques, primarily from Volume 1 of Primates, have been translated from Japanese to English and published in a volume, "Japanese Monkeys: A Collection of Translations," edited by Stuart Altmann (published 1965 by the Editor). A few copies are still available, at the cost of printing and mailing. To receive one, send US$4 to Stuart Altmann, University of Chicago, 940 East 57th St., Chicago, IL 60637.
Diagnon Corporation
Diagnon Corporation of Rockville, MD, has recently established a Department of Primate Ecology to identify and implement effective techniques to promote the well-being of nonhuman primates in breeding colonies, laboratories, and zoological parks. The Department brings together behavioral primatologists and primate veterinarians to provide the following and related services: 1) objective evaluation of environmental influences on indicators of psychological well-being; 2) reviews of relevant scientific literature; 3) consultation on facilities design and environmental enrichment programs; 4) seminars on primate environmental requirements; and 5) preparation of Institutional Primate Environmental Plans.
Diagnon's Department of Primate Ecology is devoted to increased understanding of the relationships between nonhuman primates and their environments. A major emphasis is on practical studies of the behavior and physiology of primates confined in artificial settings. The department is also active in field research on behavioral ecology, evolution, health, and conservation. Current research activities include the following: 1) the impact of foraging opportunities; 2) the consequences of changes in feeding schedules; 3) evaluation of differential effects of manipulable objects; 4) assessment of cage size effects; and 5) primate population field studies. For more information, please contact: Dr. J. Irwin, Director, Department of Primate Ecology, Diagnon Corporation, 15215 Shady Grove Road, Suite 306, Rockville, MD 20850 [301-258-9477].
* * *
(Addresses are those of first authors)
Books
*From Learning to Love: The Selected Papers of H. F. Harlow.
Clara Mears Harlow (Ed.). New York: Praeger, 1986. 340 pp. [Price:
$56.95]
. . For many years, H. F. Harlow had intended to find time and a
collaborator to write The Book, which would summarize his research and
his career. For the Centennial Psychology Series, he selected the
papers in this volume, which were edited, after his death, by Clara
Mears Harlow, who also wrote the introductory chapter.
*Primate Vocal Communication. D. Todt, P. Goedeking, & D. Symmes
(Eds.). New York: Springer-Verlag, 1988. 222 pp. [Price: $82.50]
. . Most of the chapters in this volume are based on papers presented
at a symposium entitled "New approaches towards an understanding of
primate vocal communication," held at the Freie Universität Berlin in
August, 1986. Contents:
Comparative and Field Studies. Vocal communication in primates and
birds: Parallels and contrasts, by P. Marler & J. Mitani. Vocal
quavering: A basis for recognition in forest guenons, by J.-P. Gautier &
A. Gautier-Hion. An analysis of toque macaque cohesion calls from an
ecological perspective, by W. Dittus. Environmental influences on the
structure of primate vocalizations, by C. H. Brown & P. M. Waser.
Social Interactions. A new approach to finding components essential
for intraspecific communication, by M. Maurus, K. M. Streit, D. Barclay,
E. Wiesner, & B. Kühlmorgen. Serial calling as a mediator of
interaction processes: Crying in primates, by D. Todt. Communications as
social interaction: Its importance in ontogeny and adult behavior, by C.
T. Snowdon. Conversational vocal exchanges in squirrel monkeys, by D.
Symmes & M. Biben. Vocal play behavior in cotton-top tamarins, by P.
Goedeking.
Biological Substrates. Ethopharmacology of vocal behavior in
primates, by J. D. Newman. Neural basis of signal detection, by P.
Müller-Preuss. Central control of monkey calls, by U. Jürgens.
Conceptual Frameworks. Primate vocal expression of affective state,
by K. R. Scherer & A. Kappas. Neurobiology and pathology of subhuman
vocal communication and human speech, by D. Ploog.
*A Primate Radiation: Evolutionary Biology of the African
Guenons. Annie Gautier-Hion, François Bourliére, & Jean-Pierre
Gautier (Eds.). Cambridge: Cambridge University Press, 1988. 567 pp.
[Price: $120.00]
. .Contents: I. Guenons and the African Environment: Past and
Present. Fossil evidence for the evolution of the guenons, by
M. Leakey. Guenon evolution and forest history, by A. C. Hamilton.
Habitat and locomotion in Miocene cercopithecoids, by M. Pickford & B.
Senut. Classification and geographical distribution of guenons: a
review, by J.-M. Lernould. The distribution of
Cercopithecus monkeys in West African forests, by J. F. Oates.
Distribution of guenons in the Zaïre-Lualaba-Lomani river system, by
M. M. Colyn.
II. Genetic and Phenetic Characteristics: Their Use in Phylogenetic
Reconstruction. Genetic evolution in the African guenons, by M.
Ruvolo. Population differentiation in
Cercopithecus monkeys, by T. T. Turner, J. E. Maiers, & C. S. Mott.
Chromosomal evolution of Cercopithecinae, by B. Dutrillaux, M. Muleris,
& J. Couturier. Quantitative comparisons of the skull and teeth in
guenons, by R. D. Martin & A. M. MacLarnon. Comparative morphology of
hands and feet in the genus
Cercopithecus. J. Kingdon. Interspecific affinities among guenons
as deduced from vocalizations, by J.-P. Gautier. What are face patterns
and do they contribute to reproductive isolation in guenons? by J.
Kingdon. Olfactory marking behaviour in guenons and its implications, by
J.-N. Loireau & A. Gautier-Hion.
III. Ecology and Social Behaviour. The diet and dietary habits of
forest guenons, by A. Gautier-Hion. Guenon birth seasons and correlates
with rainfall and food, by T. M. Butynski. Mating systems of forest
guenons: a preliminary review, by M. Cords. Male tenure, multi-male
influxes, and reproductive success in redtail monkeys
(Cercopithecus ascanius) , by T. T. Struhsaker. Group fission in
redtail monkeys
(Cercopithecus ascanius) in the Kibale Forest, Uganda, by T. T.
Struhsaker & L. Leland.
Cercopithecus aethiops: a review of field studies, by L. Fedigan &
L. M. Fedigan. The natural history of patas monkeys, by J. Chism & T. E.
Rowell. The social system of guenons, compared with baboons, macaques
and mangabeys, by T. E. Rowell. Polyspecific associations among forest
guenons: ecological, behavioural and evolutionary aspects, by A.
Gautier-Hion. Hybridization between redtail
(Cercopithecus ascanius schmidti) and blue (C. mitis stuhlmanni)
monkeys in the Kibale Forest, Uganda, by T. T. Struhsaker, T. M.
Butynski, & J. S. Lwanga. Concluding comments: problems, old and new, by
The Editors.
*Housing, Care and Psychological Wellbeing of Captive and
Laboratory Primates. Evalyn F. Segal (Ed.). Park Ridge, NJ: Noyes,
1989. 544 pp. [Price: $64]
. . Contents: Mental well-being in anthropoids, by C. Bramblett. What
is psychological well-being? Can we know if primates have it? by R. K.
Thomas & R. B. Lorden. Resolving issues of psychological well-being and
management of laboratory nonhuman primates, by K. Bayne. Maintenance of
primates in captivity for research: The need for naturalistic
environments, by M. E. Pereira, J. M. Macedonia, D. M. Haring, & E. L.
Simons. Psychological well-being of nocturnal primates in captivity, by
P. C. Wright, D. M. Haring, M. K. Izard, & E. L. Simons. Psychological
well-being of captive primates: General considerations and examples from
callitrichids, by C. T. Snowdon & A. Savage. The problem of foraging in
captive callitrichid primates: Behavioral time budgets and foraging
skills, by E. M. Molzen & J. A. French. Free-environment rooms as
alternative housing for squirrel monkeys, by J. E. King & V. R. Norwood.
A closed colony of squirrel monkeys for laboratory studies, by E. A.
Salzen. A room with a view for captive primates: Issues, goals, related
research and strategies, by P. O'Neill. Enriching the lives of captive
primates: Issues and problems, by M. A. Novak & K. H. Drewsen. Examining
environmental enrichment, by A. S. Chamove & J. R. Anderson. Primate
research models and environmental enrichment, by H. Markowitz & S. Line.
Long term animal studies, by C. L. Bennett & R. T. Davis. Solution to
psychological enhancement of the environment for the nonhuman primate,
by W. M. Blackmore. Environmental enrichment for monkeys used in
behavioral toxicology studies, by S. G. Gilbert & E. Wrenshall.
How not to enlarge a stable group of stumptailed macaques
(Macaca arctoides), by R. J. Rhine & R. L. Cox. Influence of people
on the behavior of display primates, by J. E. Fa. Langur monkeys
(Presbytis entellus) in captivity, by M. A. Taff & P. Dolhinow.
Beyond chronic catheterization in laboratory primates, by J. S. Turkkan,
N. A. Ator, J. V. Brady, & K. A. Craven. An inexpensive,
climate-controlled enclosure for gibbons utilizing appropriate
technology, by J. F. Dahl. Feeding enrichment for captive great apes, by
M. A. Bloomsmith. On the care of captive chimpanzees: Methods of
enrichment, by D. M. Rumbaugh, D. Washburn, & E. S. Savage-Rumbaugh.
Signs of enrichment: Toward the psychological well-being of chimpanzees,
by R. S. Fouts, M. L. Abshire, M. Bodamer, & D. H. Fouts. Environmental
influences on reproduction and maternal behavior in captive gorillas:
Results of a survey, by P. Miller-Schroeder & J. D. Paterson. The
psychological well-being of captive gorillas, by R. D. Nadler.
*Chimpanzee Politics: Power and Sex among Apes. Frans de Waal.
Baltimore: Johns Hopkins University Press, 1989. 227 pp. [Price: $12.95
paperback]
.br;*Peacemaking among Primates. Frans de Waal. Cambridge, MA:
Harvard University Press, 1989. 294 pp. [Price: $29.95]
. . Descriptions and discussions of the "political" behavior of
nonhuman primates.
Bibliographies
*Behavioral Observations of Feral African and Asian Colobine Monkeys: A Bibliography, March 1982-June 1989. J. B. Williams. Seattle: Primate Information Center, 1989. (211 citations, species & subject indexes) [Price: $6.50. Send order to Primate Information Center, Regional Primate Research Ctr., SJ-50, Univ. of Washington, Seattle, WA 98195.]
*Behavioral Observations of Feral and Free-Ranging Chimpanzees and Bonobos (Pan): A Bibliography, December 1982-May 1989. J. B. Williams. Seattle: Primate Information Center, 1989. (183 citations, species & subject indexes) [Price: $6.50. Ordering information same as above.]
Handbooks
*Handbook: Animal Models of Human Disease (Seventeenth
Fascicle). C. C. Capen, T. C. Jones, & G. Migaki (Eds.). Washington, DC:
Registry of Comparative Pathology, 1989. [Price: $10 in a binder, or $7
unbound. Send check or money order payable to UAREP, to Registry of
Comparative Pathology, Armed Forces Inst. of Pathology, Washington, DC
20306.]
. . Sixteen new studies and 8 supplemental updates from the
American Journal of Pathology and the Comparative Pathology
Bulletin. Two of the studies (strabismus and Legionnaires' disease)
and two of the supplements (baldness and NANB-hepatitis) deal with
primates.
Magazines, Newsletters, and Reports
*Primate Report, No. 22, December, 1988. [Published in
cooperation with the German Primate Center (DPZ). Price: $8]
. . This issue includes some of the Proceedings of the XIIth Congress
of the International Primate Society. Congress papers are: Normal and
abnormal behaviour in captive primates, by T. B. Poole; Atypical
behavior associated with prolonged postpartum amenorrhea in chimpanzees,
by C. E. Graham & E. J. Struthers; Morphological adaptations for loud
phonations in the vocal organ of howling monkeys, by M. A. Schön
Ybarra; Effects of the dry season on gorilla diet in Gabon, by M. E.
Rogers, E. A. Williamson, C. E. G. Tutin, & M. Fernandez; The
application and evaluation of behavioural enrichment in zoos, by D. J.
Shepherdson. Other papers are: Pattern of spatial distribution in
free-ranging monkeys of Tughlaqabad, by I. Malik; and Colonic
adenocarcinomas in
Callitrichidae (Saguinus oedipus, S. fuscicollis), by M. Brack.
Proceedings
*The Well-being of Nonhuman Primates in Research. Bethesda:
Scientists Center for Animal Welfare, 1989. [Price: $30 (discounts for
volume orders). Order from SCAW, 4805 St. Elmo Ave., Bethesda, MD
20814-4805]
. . A conference held on June 23, 1989, chaired by Jeanne Altmann and
Steven M. Niemi.
Special Journal Issues
*Proceedings of the Meeting to Celebrate the 50th Anniversary of the
Cayo Santiago Rhesus Monkey Colony.
Puerto Rico Health Sciences Journal, 1989, 8 [1]. [Price: $20.
Send check or money order payable to the University of Puerto Rico to:
Puerto Rico Health Sciences Journal, Univ. of Puerto Rico, Medical
Sciences Campus, G.P.O. Box 5067, San Juan, PR 00936.]
. . Thirty-six papers, grouped under the following subjects: Cayo
Santiago: Historical perspective, future directions; Recent behavioral
and biomedical research on Cayo Santiago; The Cayo Santiago-derived
colonies; The Caribbean Primate Research Center skeletal collection:
Status and current research.
*Nonhuman Primate Models for AIDS. J. W. Eichberg, Guest Editor.
Journal of Medical Primatology, 1989, 18 [3/4].
. . Twenty-two of the 95 presentations at the "Symposium on Nonhuman
Primate Models for AIDS," sponsored by Southwest Foundation for
Biomedical Research, in San Antonio, TX, November 16-18, 1988. Contents:
Nonhuman primate models for acquired immunodeficiency syndrome, by J. W.
Eichberg. Guidelines for the prevention of simian immunodeficiency virus
infection in laboratory workers and animal handlers, by M. D. Lairmore
et al. Monoclonal antibodies to type D retrovirus (SRV-2), by C. C.
Tsai, C.-C. Tsai, J. M. Sligh, M. Yarnall, K. E. Follis, & S. T.
Roodman. Serologic and virologic analysis of type D simian retrovirus
infection in a colony of Celebes black macaques
(Macaca nigra), by S. Shiigi et al. Mucosal epithelial cells and
Langerhans cells are targets for infection by the immunosuppressive type
D retrovirus simian AIDS retrovirus serotype 1, by A. A. Lackner, M.
Schiødt, G. C. Armitage, P. F. Moore, R. J. Munn, P. A. Marx,
M. B. Gardner, & L. J. Lowenstine. Detection of simian immunodeficiency
in macaque lymph nodes with a SIV&s'MAC envelope probe, by M. S. Wyand,
D. J. Ringler, M. Mattmuller, M. D. Daniel, R. C. Desrosiers, & N. W.
King. The productive infection of alveolar macrophages by simian
immunodeficiency virus, by D. J. Ringler, M. S. Wyand, D. G. Walsh, J.
J. MacKey, P. K. Sehgal, M. D. Daniel, R. C. Desrosiers, & N. W. King.
Use of Old World monkeys for acquired immunodeficiency syndrome
research, by I. Nicol et al. Transmission of the simian immunodeficiency
virus SIV&s'mne in macaques and baboons, by W. R. Morton et al.
Infection of macaque monkeys with simian immunodeficiency virus from
African green monkeys: Virulence and activation of latent infection, by
M. Gravell, W. T. London, R. S. Hamilton, G. Stone, & M. Monzon. Genetic
analysis and infection of SIV&s'AGM and SIV&s'MND., by T. Miura, H.
Tsujimoto, M. Fukasawa, Y. Ohta, S. Honjo, & M. Hayami. Genetic
diversity of simian immunodeficiency virus, by Y. Li, Y. Naidu, P.
Fultz, M. D. Daniel, & R. C. Desrosiers. Genetic diversity among simian
immunodeficiency virus isolates from African green monkeys, by P. R.
Johnson et al.
Molecular cloning of SIV from sooty mangabey monkeys, by V. M. Hirsch,
G. Dapolito, C. McGann, R. A. Olmsted, R. H. Purcell, & P. R. Johnson.
Molecular characterization and comparison of simian immunodeficiency
virus isolates from macaques, mangabeys, and African green monkeys, by
R. E. Benveniste et al. Use of infectious molecular clones of simian
immunodeficiency virus for pathogenesis studies, by H. W. Kestler III,
Y. N. Naidu, T. Komada, N. W. King, M. D. Daniel, Y. Li, & R. C.
Desrosiers. Rhesus macaques inoculated with molecularly cloned simian
immunodeficiency virus, by M. L. Marthas, B. Banapour, S. Sutjipto, M.
E. Siegel, P. A. Marx, M. B. Gardner, N. C. Pedersen, & P. A. Luciw.
Postexposure immunotherapy of simian immunodeficiency virus (SIV)
infected rhesus with an SIV immunogen, by M. B. Gardner, M. Jennings, J.
R. Carlson, N. Lerche, T. McGraw, P. Luciw, P. Marx, & N. Pedersen.
Inoculation of New World primates with the human immunodeficiency virus,
by M. O. McClure, T. F. Schulz, R. S. Tedder, J. Gow, J. A. McKeating, R.
A. Weiss, & A. Baskerville. Human immunodeficiency virus (HIV) from
experimentally infected chimpanzees: Isolation and characterization, by
B. A. Castro, C. M. Walker, M. Tateno, C. Cheng-Mayer, R. Heberling, J.
W. Eichberg, & J. A. Levy. The biology of human immunodeficiency virus-1
IIIB infection in the chimpanzee: in vivo and in vitro correlations, by
P. Nara, W. Hatch, J. Kessler, J. Kelliher, & S. Carter. Specificity of
chimpanzee antibodies binding a strain-specific HIV-1 neutralization
epitope of the external envelope, by J. Goudsmit, M. Bakker, L. Smit, &
R. H. Meloen. Proliferative T-cell response to HIV envelope glycoprotein
in immunized and infected primates and human beings, by K. Krohn, A.
Ranki, P. Lusso, B. Moss, P. Ashorn, L. O. Arthur, S. D. Putney, & R. C.
Gallo.
Videotapes
*Research Methods for Studying Animal Behavior in a Zoo Setting.
Washington Park Zoo & Minnesota Zoological Garden. 80 minutes. [Price:
$200, as 3/4-inch videocassette or 1/2-inch VHS tape. Order from Steven
Hage, Minnesota Zoological Garden, Apple Valley, MN 55124]
. . Formulating a research question, sampling methods, data
collection, statistical analysis, and interpretation of results are
outlined. For each of 5 sampling methods, the viewer observes and may
practice scoring taped animal behavior. Ten copies of a text with
examples of ethograms, bibliography, and checksheets are included.
Animal Models
*Spontaneous diabetes mellitus in a rhesus monkey:
Neurophysiological studies. Cornblath, D. R., Dellon, A. L., &
MacKinnon, S. E. (Meyer 2-147, Johns Hopkins Hospital, Baltimore, MD
21205).
Muscle & Nerve, 1989, 12, 233-235.
. . Four months after developing diabetes, median and ulnar sensory
action potential amplitudes and the median motor distal latency of a
22-year-old female rhesus monkey differed from controls by more than 2
standard deviations. These electrophysiologic abnormalities are similiar
to those described in human diabetes and suggest that the spontaneously
diabetic monkey may be a suitable model of experimental diabetic
neuropathy.
*Cell-mediated immune response toward viral envelope and core
antigens in gibbon apes
(Hylobates lar) chronically infected with human immunodeficiency
virus-1. Lusso, P., Markham, P. D., Ranki, A., Earl, P., Moss, B.,
Dorner, F., Gallo, R. C., & Krohn, K. J. E. (R. C. Gallo, Lab. of Tumor
Cell Biology, NCI, Bldg. 37, Room 6A09, Bethesda, MD 20892).
Journal of Immunology, 1988, 141, 2467-2473.
. . The specific T cell reactivity toward HIV-1 proteins observed in
infected gibbons contrasts with findings in HIV-1 infected humans, and
may help to explain the apparent discrepancy in the natural history of
the infection between the two species. The ability to isolate infectious
virus from the peripheral blood of these gibbons has now persisted for
more than a year. The clinical and hematologic picture of the infected
animals included lymphadenopathy, eosinophilia, and neutrophilia.
Lymphopenia, reduction in CD4-positive cells or other features
characteristic of HIV-1 infection in humans have not occurred to date.
*Characterization of infectious molecular clones of simian
immunodeficiency virus (SIV&s'mac.) and human immunodeficiency virus
type 2: Persistent infection of rhesus monkeys with molecularly cloned
SIV&s'mac.. Naidu, Y. M., Kestlerr, H. W. III, Li, Y., Butler, C. V.,
Silva, D. P., Schmidt, D. K., Troup, C. D., Sehgal, P. K., Sonigo, P.,
Daniel, M. D., & Desrosiers, R. C. (R. C. Desrosiers, New England
Regional Primate Research Center, 1 Pine Hill Dr., Southborough, MA
01772).
Journal of Virology, 1988, 62, 4691-4696.
. . This report describes 3 infectious molecular clones of SIV&s'mac
and one of HIV-2 and their use in the study of cell and species
specificity, animal infection, and the releationship of gene sequence to
function. These clones will be very useful for future studies directed
at the molecular basis for persistence, pathogenicity, tropism, and cell
and species specificity.
*Cellular localization of simian immunodeficiency virus in lymphoid
tissues. I. Immunohistochemistry and electron microscopy. Ringler, D.
J., Wyand, M. S., Walsh, D. G., MacKey, J. J., Chalifoux, L. V.,
Popovic, M., Minassian, A. A., Sehgal, P. K., Daniel, M. D., Desrosiers,
R. C., & King, N. W. (Address same as above).
American Journal of Pathology, 1989, 134, 373-383.
. . Immunohistochemical procedures were used to localize the p27
gag protein to specific cellular immunophenotypes. In rhesus
monkeys with follicular hyperplasia, viral protein was found associated
predominantly with follicular dendritic cells. In lyphoid tissues with
other histopathologic changes, macrophages and multinucleate giant cells
were the predominant cell types containing detectable quantities of
viral protein, and viral particles were found within the extracellular
space adjacent to tissue macrophages. Certain histologic patterns seen
during the course of infection correlated with the localization of viral
antigen to specific cellular immunophenotypes, and viral protein is
preferentially localized in sections of lymph node and spleen to cells
of the macrophage and dendritic cell lineages.
*Serial passage of hepatitis delta virus in chronic hepatitis B
virus carrier chimpanzees. Ponzetto, A., Negro, F., Popper, H., Bonino,
F., Engle, R., Rizzetto, M., Purcell, R. H., & Gerin, J. L. (J. L.
Gerin, Div. of Molecular Virology & Immunology, Georgetown Univ. Medical
Center, 5640 Fishers Lane, Rockville, MD 20852).
Hepatology, 1988, 1655-1661.
. . HDV consistently infected HBV carrier chimpanzees; HDV infection
was associated with severe liver disease; the adaptation of HDV to
chimpanzees was associated with increased HDV virulence; and the
duration of delta hepatitis was linked to individual host response. The
chimpanzee reproduces the human disease, thereby providing a model in
which to study the pathogenesis of acute delta hepatitis and to evaluate
therapeutic strategies.
Animal Welfare
*Methods to evaluate the wellbeing of laboratory primates:
Comparisons of macaques and tamarins. Evans, H. L., Taylor, J. D.,
Ernst, J., & Graefe, J. F. (Inst. of Environmental Medicine, New York
Univ. Medical Center, New York, NY 10016).
Laboratory Animal Science, 1989, 39, 318-323.
. . Three objective behavioral tests are described for assessment of
wellbeing and possible enrichment: foraging in a pickup tray, a puzzle
feeder, and measurement of diurnal rhythm of homecage activity. Such
definitive measurements, coupled with data on long-term health outcomes,
are essential before regulations for primate care can be revised with
confidence.
*Adult female rhesus macaque responses to novel objects. Line, S.
W., Clarke, A. S., & Markowitz, H. (California Primate Research Center,
Univ. of California, Davis, CA 95616).
Lab Animal, 1989, 18[4], 33-35, 37, 39-40.
. . The novelty of introducing a plastic teething ring apparently was
not sufficiently stressful to significantly elevate cortisol levels. A
Nylaball was, unexpectedly, manipulated more than a set of colored
plastic blocks. Large behavioral variations both within and between
monkeys suggests individual preferences for objects, and indicates a
need for long-term evaluation on many subjects.
Behavior
*Adult male replacement and group formation in
Presbytis rubicunda. Davies, G. (Dept. of Anthropology, University
College London, Gower St., London, WCIE 6BD, UK).
Folia Primatologica, 1987, 49, 111-114.
. . Overall, the pattern of group formation in
P. rubicunda conforms well with features described for other species
in the genus
Presbytis, despite differences in their taxonomy and ecology.
*Cathemeral activity in primates: A definition. Tattersall, I.
(Dept. of Anthropology, American Museum of Natural History, New York, NY
10024).
Folia Primatologica, 1987, 49, 200-202.
. . The term "cathemeral" is proposed for activity during both night
and day, as seen in
Alouatta, Aotus, and several lemurs and hapalemurs.
*Activity patterns in a captive group of Celebes black apes
(Macaca nigra). Bernstein, I. S. & Baker, S. C. (Dept. of
Psychology, Univ. of Georgia, Athens, GA 30602).
Folia Primatologica, 1988, 51, 61-75.
. . Activity patterns of
M. nigra are compared to those of M. arctoides, M. nemestrina,
sooty mangabeys, and geladas. The consistency of activity profiles
across the sample of cercopithecine species living under nearly
identical environmental conditions suggests a broad ecological
adaptation for this subfamily.
*Orangutan activity budgets: Monthly variations and the effects of
body size, parturition, and sociality. Mitani, J. C. (Dept. of Zoology,
Univ. of California, Davis, CA 95616).
American Journal of Primatology, 1989, 18, 87-100.
. . Adult orangutans are primarily solitary, with animals associating
for only temporary periods. Adult females and subadult males travelled
and fed significantly longer than an adult male. Associations with
females forced a male orangutan to travel significantly more compared
with periods in which he was solitary, although he did not lose an
appreciable amount of time feeding when accompanying a female.
*Social relationships in a cooperatively polyandrous group of
tamarins
(Saguinus fuscicollis) . Goldizen, A. W. (Dept. of Ecosystem
Management, Univ. of New England, Armidale, N.S.W. 2351, Australia).
Behavioral Ecology and Sociobiology, 1989, 24, 79-89.
. . Detailed data on the social relationships among the adults, and
between the adults and young, of a cooperatively polyandrous saddle-back
tamarin group. The two polyandrous males in the main study group were
very rarely aggressive to each other, rarely tried to disrupt each
others' copulations, groomed each other, and occasionally shared food.
It is hypothesized that the mating pattern of a saddle-back tamarin
group is determined primarily by the number of older offspring still
present in the group, because pairs need help in raising infants. This
help can be provided either by older offspring or by additional adult
males.
*Female-female aggression and male indifference in response to
unfamiliar intruders in lion tamarins. French, J. A. & Inglett, B. J.
(Dept. of Psychology, Univ. of Nebraska, Omaha, NE 68182).
Animal Behaviour, 1989, 37, 487-497.
. . Female intolerance of conspecifics may be important in the
regulation of sociosexual relationships in breeding pairs, and
female-female reproductive competition may be particularly extreme in
golden lion tamarins.
*Mirror reactions in a zoo colony of
Cebus monkeys. Collinge, N. E. (Dept. of Anthropology, Univ. of
Alberta, Edmonton, Alta, T6G 2H4 Canada).
Zoo Biology, 1989, 8, 89-98.
. . Intermittent exposure to a mirror over a 5-week period produced
species-specific social reactions to the mirror, supporting the
suggestion that at least in some instances the monkeys were viewing
their mirror image as a conspecific.
*Effects of social environment on play in squirrel monkeys:
Resolving Harlequin's dilemma. Biben, M. (Bldg. T-18, NIHAC, NIH,
Bethesda, MD 20892).
Ethology, 1989, 81, 72-82.
. . Yearling-male squirrel monkeys were paired with play partners
either matched or different from them in age and/or sex. Frequency of
play was maintained at equally high levels, but play between mismatched
partners suggest that youngsters had considerable flexibility in the use
of alternate strategies to sustain play activity with partners who were
atypical for them. The technique of restricting youngsters to atypical
play companions is suggested as an effective means of manipulating play,
with minimal disruption of other social experiences, for long-term
studies of the developmental impact of social play.
*Styles of dominance and their endocrine correlates among wild olive
baboons
(Papio anubis). Sapolsky, R. M. & Ray, J. C. (Dept. of Biological
Sciences, Stanford Univ., Stanford, CA 94305-5020).
American Journal of Primatology, 1989, 18, 1-13.
. . Low basal cortisol concentration is only found among dominant
males with any of the following behaviors: the most marked ability to
distinguish between threatening and merely neutral interactions with
rivals and, if the former, the greatest likelihood of initiating a
fight; the most skill at distinguishing between winning and losing a
fight and, if the latter, the greatest likelihood of displacing
aggression onto a third party. Collectively, these behaviors suggest
high degrees of social skillfulness, control, and predictability over
social contingencies. Dominant males lacking these behavioral features,
in contrast, had as high cortisol concentrations as did subordinate
males.
*Defensive behaviors in infant rhesus monkeys: Environmental cues
and neurochemical regulation. Kalin, N. H. & Shelton, S. E. (Dept. of
Psychiatry, Univ. of Wisconsin, Madison, WI 53705).
Science, 1989, 243, 1718-1721.
. . Infants separated from their mothers were active and emitted
frequent distress vocalizations. When a human entered the room but did
not look at the infant, it became silent and froze in one position. If
the human stared at the infant, it responded with aggressive barking.
Alterations of the opiate system affected the frequency of the infant's
distress calls without affecting barking and freezing, whereas
benzodiazepine administration selectively reduced barking and freezing.
This suggests that opiate and benzodiazepine systems regulate specific
defensive behaviors in primates and that these systems work together to
mediate behavioral responses important for survival.
*Rank relations of juvenile and subadult natal males of Barbary
macaques
(Macaca sylvanus) at Affenberg Salem. Kuester, J. & Paul, A. (Inst.
für Anthropologie der Univ. Göttingen, Bürgerstrasse 50, D-3400
Göttingen, DFR).
Folia Primatologica, 1988, 51, 33-44.
. . An analysis of rank relations of more than 100 juvenile and
subadult natal Barbary macaque males showed heirarchical relations among
individuals of the same age early in the first year of life. Rank
position of a male among his male peers was influenced by birth order,
maternal rank, and the presence of juvenile brothers. There was no
cohort where ranking among males was determined by maternal rank alone,
as is the case in rhesus monkeys and Japanese macaques.
*The behavioral repertoire of the black-and-white ruffed
lemur, Varecia variegata variegata (Primates: Lemuridae). Pereira,
M. E., Seeligson, M. L., & Macedonia, J. M. (Duke Univ. Primate Center,
3705 Erwin Rd., Durham, NC 27705).
Folia Primatologica, 1988, 51, 1-32.
. . A stable social group of 7 semifree-ranging black-and-white ruffed
lemurs was studied for 4 months, resulting in a catalog of 11 major
behavioral categories: postures, terrestrial locomotion, arboreal
locomotion, feeding behavior, vocalizations, scent-marking, affinitive
social behavior, agonistic social behavior, play, sexual behavior, and
parental behavior.
*Feeding strategies of Japanese monkeys against deterioration of
habitat quality. Nakagawa, N. (Primate Research Inst., Kyoto Univ.,
Inuyama, Aichi, 484 Japan).
Primates, 1989, 30, 1-16.
. . As monkeys or their competitors, such as mice, utilized food
resources, the monkeys controlled the decrease in food intake by
exploiting new food patches, extending the time spent on feeding, and
changing their food.
*Activity budget and diet of patas monkeys in Kala Maloue National
Park, Cameroon: A preliminary report. Nakagawa, N. (Address same as
above).
Primates, 1989, 30, 27-34.
. . Patas monkeys spent 30% of waking time feeding, mostly on flowers
and buds of herbaceous plants and the larvae of insects. They spent more
time in feeding and travelled for longer distances per day than the
sympatric tantalus monkey.
*Longitudinal changes of dominance rank among the females of the
Koshima group of Japanese monkeys. Mori, A., Watanabe, K., & Yamaguchi,
N. (Address same as above).
Primates, 1989, 30, 147-173.
. . Changes over a period of 29 years from 1957 were studied. The most
important factor which determined the females' rank was their dominance
relations under the influence of their mothers in early life.
Care
*The effects of single caging on chimpanzee behavior. Brent, L.,
Lee, D. R., & Eichberg, J. W. (Dept. of Virology & Immunology, Southwest
Foundation for Biomedical Research, San Antonio, TX 78284).
Laboratory Animal Science, 1989, 39, 345-346.
. . Ten chimpanzees, housed in groups of 3 to 5 animals, were studied
for 1 week before and 5 weeks after being moved to single cages. Self
manipulation, self-directed behavioral pathologies, and nonspecific
vocalizations did not change significantly after the move. Environmental
manipulation and specific vocalization decreased significantly, while
stereotyped behaviors were increased significantly.
*Effects of environmental conditions on the psychological well-being of primates: A review of the literature. Woolverton, W. L., Ator, N. A., Beardsley, P. M., & Carroll, M. E. (Dept. of Pharmacological & Physiological Sciences, University of Chicago, 947 E. 58th St., Chicago, IL 60637). Life Sciences, 1989, 44, 901-917.
*Uprooted trees facilitate the psychological well-being of captive
chimpanzees. Maki, S. & Bloosmith, M. A. (Univ. of Texas Cancer Center,
Route 2, Box 151-B1, Bastrop, TX 78602).
Zoo Biology, 1989, 8, 79-87.
. . After the introduction of uprooted trees to their environment, the
behavior of 28 socially housed, laboratory chimpanzees was studied for 5
months. Subjects used the tree during 41.9% of the data points collected
during the first day of introduction. Thereafter, the mean for tree use
dropped to 3.5% and remained fairly consistent.
*Effect of food preparation on feeding behavior of lion-tailed
macaques. Smith, A., Lindburg, D. G., & Vehrencamp, S. (CRES, San Diego
Zoo, P.O. Box 551, San Diego, CA 92112).
Zoo Biology, 1989, 8, 57-65.
. . Ten group-living
Macaca silenus, accustomed to receiving fruits and vegetables in
chopped form, were given the same foods in whole form to compare
reactions. Contrary to common belief, the access of individuals to the
different food items was not equalized through chopping into bite-sized
morsels. Mean dietary diversity actually increased with presentation of
whole foods, as did time spent feeding and total amount of food
consumed. The widespread practice of chopping food for captive primates
has little to commend it in light of these findings.
Conservation
*Some observations of two kinds of lemuridae
(Varecia variegata variegata and Lemur fulvus albifrons)
in the reserve of Nosy Mangabe. Iwano, T. (Dept. of Physical
Anthropology, Fac. of Science, Univ. of Tokyo, Hongo 7-3-1, Bunkyo-ku,
Tokyo, Japan).
Primates, 1989, 30, 241-248.
. . Results of a short term survey conducted shortly before the
population declined from alleged poaching.
Development
*Twins in free-ranging Hanuman langurs
(Presbytis entellus). Winkler, P., Wrogemann, D., & Prestel, H.
(Inst. Anthropologie der Univ. Göttingen, Bürgerstrasse 50, D-3400
Göttingen, DFR).
Primates, 1989, 30, 255-259.
. . Two pairs of twins were observed at Jodhpur, India. In more than
60% of all activity bouts both showed the same behavior, with "nipple
contact" and "playing" prevailing. Differences in physical strength and
development were apparent within each pair.
Disease
*Intestinal trichomonads
(Tritrichomonas mobilensis) in the natural host Saimiri sciureus
and Saimiri boliviensis. Scimeca, J. M., Culberson, D. E., Abee, C.
R., & Gardner, W. A. Jr. (Dept. of Comparative Medicine, Univ. of South
Alabama College of Medicine, Mobile, AL 36688).
Veterinary Pathology, 1989, 26, 144-147.
. . A retrospective study of cecal and colonic tissues from 28 squirrel
monkeys demonstrated enteric trichomonads within luminal crypts.
Histologic changes included mucosal ulceration, which raises the
possibility that trichomonads are pathogens in the squirrel monkey,
multifocal cryptitis, and focal epithelial necrosis. Most areas
containing trichomonads did not have an associated inflammatory
response.
*Studies of GB hepatitis agent in tamarins. Karayiannis, P.,
Petrovic, L. M., Fry, M., Moore, D., Enticott, M., McGarvey, M. J.,
Scheuer, P. J., & Thomas, H. C. (H. C. Thomas, Dept. of Medicine, St.
Mary's Hospital Medical School, South Wharf Rd., London W2 1NY,
England).
Hepatology, 1989, 9, 186-192.
. . Serum and liver extracts from tamarins which had been infected
with serum from a human GB hepatitis patient transmitted the disease to
other tamarins. Fecal specimens were noninfectious. GB hepatitis virus
is distinct from the viruses causing type A and blood-borne NANB
hepatitis.
*Human-rhesus reassortant rotavirus vaccines: Safety and
immunogenicity in adults, infants, and children. Halsey, N. A.,
Anderson, E. L., Sears, S. D., Steinhoff, M., Wilson, M., Belshe, R. B.,
Midthun, K., Kapikian, A. Z., Chanock, R. M., Samorodin, R., Burns, B.,
& Clements, M. L. (Center for Immunization Research, Dept. of
International Health, Johns Hopkins Univ. School of Hygiene & Public
Health, 615 N. Wolfe St., Baltimore, MD 21205).
Journal of Infectious Diseases, 1988, 158, 1261-1267.
. . Human-rhesus rotavirus vaccines appeared to be safe and
immunogenic in the small numbers of adult and pediatric subjects in
these studies. Approximately 80% of young infants and older participants
with serum neutralization antibody titers </= 1:160 responded by developing
a fourfold rise in titer of serum antibody.
Lack of transmission of the vaccine virus
from vaccinees to placebo recipients in a day care-like setting is
evidence that the reassortant vaccines will not be associated with a high
risk of transmission to close contacts.
Facilities
*The Yunnan National Laboratory Primate Center of China (YNLPC): An
outline. The Editors (Japan Monkey Centre, Inuyama, Aichi, 484, Japan).
Primates, 1989, 30, 143-145.
. . A description of the facility, which holds over 2000 nonhuman
primates of 8 species, including macaques, gibbons, and the slow loris.
General
*Interface of field and laboratory-based research in primatology.
Rosenblum, L. A., Kummer, H., Nadler, R. D., Robinson, J., & Suomi, S.
J. (S. J. Suomi, Lab. of Comparative Ethology, NICHD, Building 31, Room
B2B-15, 9000 Rockville Pike, NIH, Bethesda, MD 20892).
American Journal of Primatology, 1989, 18, 61-64.
. . A statement of the consensus and conclusions of a conference,
February 26-29, 1988, convened by the National Institute of Child Health
and Human Development through its Laboratory of Comparative Ethology,
IRP.
Instruments & Techniques
*A technique for catheterization and cystoscopic evaluation of
cynomolgus monkey urinary bladders. Bahnson, R. R., Ballou, B. T.,
Ernstoff, M. S., Schwentker, F. N., & Hakala, T. R. (Div. of Urological
Surgery, Univ. of Pittsburgh School of Medicine, Pittsburgh, PA 15213).
Laboratory Animal Science, 1988, 38, 731-733.
. . A catheter was trimmed so the end of a pediatric otoscope just
protruded, and a camera was attached, allowing cystoscopic examination
and photography.
Pharmacology & Anesthesia
*Ketamine enhances Phase I and Phase II neuromuscular block of
succinylcholine. Tsai, S. K., Lee, C., & Tran, B. (C. Lee, Dept. of
Anesthesiology, Harbor-UCLA Medical Center, 1000 W. Carson St.,
Torrance, CA 90509).
Canadian Journal of Anaesthesia, 1989, 36, 120-123.
. . In Macaca cyclopis, in the absence of succinylcholine,
ketamine had no consistent neuromuscular effect of its own. In the
presence of succinylcholine, ketamine in a dose-dependent manner
potentiated both the Phase I and the Phase II neuromuscular blocking
effect of succinylcholine.
*Experimental drug-induced immune thrombocytopenia in monkeys.
Petersen, B. H., Heim, M. C., & White, J. F. (Lilly Lab. for Clinical
Research, Wishard Memorial Hospital, 1001 W. 10th St., Indianapolis, IN
46202).
Diagnostic and Clinical Immunology, 1988, 5, 349-354.
. . Three of 8 rhesus monkeys developed thrombocytopenia following
administration of high doses of 7,8-dimethoxy-1H-3-benzaepin-2-amine
HCl. Platelet surface-associated immunoglobulin could be detected by
flow cytometry as early as 21 days before the onset of thrombocytopenia.
Evidence suggests that drug interaction with the platelet surface
produced a site for antibody binding, possibly a neoantigen, which
required the continued presence of the drug.
*Immobilization with a single dose of ketamine hydrochloride and a
combination of xylazine hydrochloride-ketamine hydrochloride and
antagonism by Yohimbine hydrochloride in the Japanese monkey
(Macaca fuscata). Hayama, S.-I., Terazawa, F., Suzuki, M., Nigi, H.,
Orima, H., Tagawa, M., & Inagaki, H. (Dept. of Wild Animal Medicine,
Nippon Veterinary & Zootechnical College, Musashino, Tokyo, 180 Japan).
Primates, 1989, 30, 75-79.
. . Mean and standard deviations of time of induction,
immmobilization, sitting, and walking for different dosages and
treatments of 49 free-ranging Japanese monkeys.
Physiology
*Reference values of hematological, serum biochemical and
hormonal examinations in female cynomolgus monkeys
(Macaca fascicularis) of feral origin. Yoshida, T., Katsuka, A.,
& Cho, F. (Tsukuba Primate Center for Medical Science, National Inst.
of Health, Hachimandi, Tsukuba-shi, Ibaraki 305, Japan).
Experimental Animals, 1989, 38, 259-262.
. . Blood samples from clinically healthy female cynomolgus
monkeys, maintained under uniform environmental conditions for
4 to 5 years, were examined for their hematological, serum
biochemical, and hormonal values. Ranges of values, as well as
their arithmetic means and standard deviations have been
tabulated with respect to each examination item.
*Plasma alpha-tocopherol, retinol, cholesterol, and mineral
concentrations in captive gorillas. McGuire, J. T., Dierenfeld, E. S.,
Poppenga, R. H., & Brazelton, W. E. (Animal Health Center, New York
Zoological Park, Bronx, NY 10460).
Journal of Medical Primatology, 1989, 18, 155-161.
. . Vitamins A, E, cholesterol, and mineral concentrations were
examined in the plasma of 74 captive lowland gorillas aged newborn to 41
years. Effects of age or sex were not significant. Since comparative
gorilla values for certain blood components (vitamins E and A, copper,
and selenium) are lacking, normal human values may provide the best
available indicators for evaluating the plasma levels of these
components in gorillas.
*The intrinsic rate of natural increase and reproductive effort in
primates. Ross, C. (Anthropologigiches Inst. & Museum der Univ.
Zürich-Irchel, Winterthurerstrasse 190, CH-8057 Zürich,
Switzerland).
Journal of Zoology, London, 1988, 214, 199-219.
. . The intrinsic rate of natural increase is calculated for 58
primate species. Relationships are sought between that rate, body
weight, environmental predictability, prenatal maternal investment, and
diet.
*Color vision variations in Old and New World primates. Jacobs, G.
H. & Harwerth, R. S. (Dept. of Psychology, Univ. of California, Santa
Barbara, CA 93106).
. . Widespread individual variation in color vision has been found to
occur in members of several genera of New World monkeys. Behavioral
measurements of increment-threshold spectral sensitivity was compared
for large samples of squirrel monkeys and macaque monkeys. If they occur
at all, individual variations in color vision among macaque monkeys must
be rare.
Reproduction
*1988 Regional Studbook Black Howler Monkey Alouatta caraya. B. Baker. 67 pp. [Riverbanks Zoological Park, P.O. Box 1060, Columbia, SC 29202-1060].
*Environment, competition and reproductive performance of female
monkeys. Harcourt, A. H. (L.A.R.G., Dept. of Zoology, Univ. of
Cambridge, 34A Storey's Way, Cambridge CB3 0DT, UK).
Trends in Ecology and Evolution, 1989, 4, 101-105.
. . A review, which indicates that dominant females may have higher
reproductive performance than subordinates if environmental conditions
permit them to monopolize resources sufficiently to disadvantage
subordinates.
*Generational bias in mating in captive groups of rhesus monkeys
(Macaca mulatta). Smith, D. G. (Dept. of Anthropology, University
of California, Davis, CA 95616).
. . Paternity exclusion analysis was
conducted for 431 offspring born over 7 years in 3 closed,
corral-housed groups. it is concluded that natal rhesus monkeys prefer
to reproduce with their generational peers when they are available.
*Births following the transfer of cultured embryos obtained by in
vitro and in vivo fertilization in the marmoset monkey
(Callithrix jacchus). Lopata, A., Summers, P. M., Hearn, J. P.
(Dept. of Obstetrics & Gynaecology, Univ. of Melbourne, Royal Women's
Hospital, 132 Grattan St., Carlton, Victoria 3053, Australia).
Fertility and Sterility, 1988, 50, 503-509.
. . Five embryos obtained by in vitro fertilization were transferred
at early stages to the uterus of 3 recipients, and 2 gave birth to live
infants. In vivo fertilized oocytes were also cultured and transferred
to 2 recipients, and 1 gave birth.
*Maternal factors affecting reproduction in a breeding colony of
cynomolgus macaques
(Macaca fascicularis). Gardin, J. F., Jerome, C. P., Jayo, M. J., &
Weaver, D. S. (Dept. of Comparative Medicine, Bowman Gray School of
Medicine, Wake Forest Univ., Winston-Salem, NC 27103).
Laboratory Animal Science, 1989, 39, 205-212.
. . The effects of age, gravidity, parity, origin, previous
stillbirths, clinical incidents, and type of housing on pregnancy rate,
stillbirth rate, infant mortality rate, and pregnancy success were
evaluated for a breeding colony over 9 years.
*Lactation and care for unrelated infants in forest-living
ringtailed lemurs. Pereira, M. E. & Izard, M. K. (Duke Univ. Primate
Center, 3705 Erwin Rd., Durham, NC 27705).
American Journal of Primatology, 1989, 18, 101-108.
. . An adult female Lemur catta known not to have been pregnant
showed spontaneous lactation in response to twin infants born to an
unrelated female. Another adult female, who never carried either infant
and had no milk, like the two "maternal" females frequently attacked
unfamiliar immigrating adult males when the males approached the
infants. Potential implications of these observations concerning the
social organization of ringtailed lemurs are discussed.
*Gestation and birth-related behaviors in Temminck's red colobus.
Starin, E. D. (CUNY, Dept of Anthropology, 33 W. 42nd St., New York, NY
10036).
Folia Primatologica, 1988, 51, 161-164.
. . Observations of various aspects of behavior related to gestation
and parturition made during a 5.5-year field study in the Abuko Nature
Reserve, The Gambia.
*Semen characteristics of vervet monkeys. Seier, J. V., Fincham, J.
E., Menkveld, R., & Venter, F. S. (Research Inst. for Nutritional
Diseases, South African Medical Research Council, P.O. Box 70, Tygerberg
7505, South Africa).
Laboratory Animals, 1989, 23, 43-47.
. . Semen characteristics of 37 singly caged males of unknown
fertility are compared to those of 10 breeding males of known fertility.
*Changes of plasma estradiol, progesterone and LH level in the
menstrual cycle of the adult female Japanese monkey during the mating
season. Hamada, K., Suginai, H., & Matsuura, S. (Dept. of Obstetrics &
Gynecology, School of Medicine, Ehime Univ., 791-02 Shitsukawa,
Shigenobu-chou, Onsen-gun, Ehime, Japan).
Experimental Animals, 1989, 38, 187-194 (Japanese, with English
abstract).
. . Seventeen graphs of results from 15 monkeys show a variety of
cycle patterns.
*The reproductive status of nonbreeding group members in captive
golden lion tamarin social groups. French, J. A., Inglett, B. J., &
Dethlefs, T. M. (Dept. of Psychology, Univ. of Nebraska, Omaha, NE
68182).
American Journal of Primatology, 1989, 18, 73-86.
. . A suppression of ovulatory cyclicity is responsible for the lack
of reproductive activity in daughters and subordinate females in 3
species of callitrichids studied to date. In golden lion tamarins
studied here, mean levels of estrogen excretion in these females were
similar to those of nonpregnant, breeding adult females. While younger
daughters showed lower levels of estrogen excretion, older daughters and
subordinate females may be capable of expressing normal ovarian function
in the presence of a breeding adult female. This may account for the
high level of female-female aggression, and the presence of groups in
the wild with more than one actively breeding female.
*Serum hormone levels in pregnant cynomolgus monkeys. Hein, P. R.,
Schatorjé, J. S. J. O., Frencken, H. J. A. A. M., Segers, M. F. G., &
Thomas, C. M. G. (Dept. of Obstetrics & Gynecology, St. Radboud
Hospital, 6500 HB Nijmegen, The Netherlands).
Journal of Medical Primatology, 1989, 18, 133-142.
. . Serum levels of estradiol (E2.), progesterone (P), prolactin
(Prl), monkey chorionic gonadotropin (mCG), dehydroepiandrosterone
sulfate (DHEAS), and cortisol (C) in pregnant cynomolgus monkeys are
described. mCG, E2., and PRL patterns resembled those of pregnant
rhesus monkeys. P patterns differed from data from the literature. DHEAS
patterns did not follow E2 patterns. C levels did not change during
pregnancy.
*Serum chorionic gonadotropin, progesterone, and estradiol-17-beta
levels during pregnancy in the common marmoset,
(Callithrix jacchus). Torii, R., Nigi, H., Koizumi, H., & Tanioka,
Y. (Inst. for Experimental Animals, Shiga Univ. of Medical Science,
Seta, Ohtsu, Shiga, 520-21, Japan). Primates, 1989, 30, 207-215.
. . The time course of the serum progesterone and estradiol-17-beta during
pregnancy and the role of the placenta which synthesized and secreted
these hormones in the common marmoset showed a similarity to those of
humans and anthropoid apes rather than those of
Macaca species. The common marmoset represents a valuable animal
model for investigating the feto-placental unit in humans.
*Seasonal food shortage, weight loss, and the timing of births in
saddle-back tamarins
(Saguinus fuscicollis) . Goldizen, A. W., Terborgh, J., Cornejo, F.,
Porras, D. T., & Evans, R. (Dept. of Anthropology, Univ. of Michigan, Ann
Arbor, MI 48109-1382).
Journal of Animal Ecology, 1988, 57, 893-901.
. . It is suggested that tamarin births are timed such that lactation
and weaning occur when food is abundant, because during the period of
low food availablility, there would be insufficient food to meet the
demands of lactation and to serve as easily obtainable weaning foods.
*Season determines timing of first ovulation in rhesus monkeys
(Macaca mulatta) housed outdoors. Wilson, M. E. & Gordon, T. P.
(Yerkes Regional Primate Research Center, Emory Univ., Field Station,
2409 Taylor Rd., Lawrenceville, GA 30243).
Journal of Reproduction & Fertility, 1989, 85, 583-591.
. . The timing of puberty in spring-born females was compared to that
of fall and winter-born ones. Results suggest that exposure to an
outdoor environment restricts the occurrence of first ovulation to the
fall or winter in female rhesus monkeys, regardless of season of birth,
such that seasonal cues may override the endogenous control of puberty
onset.
*Female dominance rank and behavior during artificial menstrual
cycles in social groups of rhesus monkeys
(Macaca mulatta). Zumpe, D. & Michael, R. P. (R. P. Michael, Dept.
of Psychiatry, Emory Univ. School of Medicine, 1256 Briarcliff Rd.,
N.E., Atlanta, GA 30306).
American Journal of Primatology, 1989, 17, 287-304).
. . To assess the role of dominance, artificial menstrual cycles were
induced with hormone injections in 2 of 4 ovariectomized females in 8
social groups. The amount of male sexual activity received by dominant
and subordinate females differed little, but dominant females initiated
significantly more sexual activity
(P < .0001) than did subordinate females. The findings provide new
evidence for both female mate competition and mate choice by males for
higher-ranking females.
*Male careers in Sumatran long-tailed macaques
(Macaca fascicularis) . van Noordwijk, M. A. & van Schaik, C. P.
(Lab. voor Vergelijkende Fysiologie, Rijksuniversiteit Utrecht, Jan van
Galenstraat 40, 3572 LA Utrecht, The Netherlands).
Behaviour, 1988, 107, 24-43.
. . Focal animal samples were collected over 4 years in 2 groups of
long-tailed macaques to study their behavior. A generalized career
profile of males is constructed in order to facilitate comparison with
other promiscuous species.
Supply
*Monkeys as a renewable resource. Aronson, R. (MRC NHB163,
Smithsonian Inst., Washington, DC 20560).
New Scientist, 1988, 123 [1679], 60-61.
. . A discussion of Jean Baulu's methods of harvesting African green
monkeys, which are agricultural pests in Barbados.
Taxonomy
*Variations in vocal patterns of Senegal and South African lesser
bushbabies and their implications for taxonomic relationships.
Zimmermann, E., Bearder, S. K., Doyle, G. A., & Andersson, A. B. (Inst.
of Zoology, Univ. of Stuttgart-Hohenhei, D-7000 Stuttgart, 70 DFR).
Folia Primatologica, 1988, 51, 87-105.
. . Striking divergences in almost all of the tonal or harmonic call
types given in association with contact or contact-seeking, defensive or
alarm behavior provide strong support for the separation of the two
forms into distinct species
Galago senegalensis and G. moholi.
*Taxonomy and distribution of the François' langur
(Presbytis francoisi). Ma, S., Wang, Y., & Poirier, F. E. (Dept. of
Vertebrate Zoology, Kunming Inst. of Zoology, Academia Sinica, Kunming,
Yunnan, PRC).
Primates, 1989, 30, 233-240.
. . It is suggested that the white-headed langur is a subspecies of
Presbytis francoisi and should be referred to as
P. francoisi leucocephalus.
*Origin, speciation, and distribution of South American titi
monkeys, genus
Callicebus (family Cebidae, Platyrrhini). Hershkovitz, P. (Field
Museum of Natural History, Chicago, IL 60605).
Proceedings of the Academy of Natural Sciences of Philadelphia,
1988, 140, 240-272.
. . A summary of the biogeographic data of the 13 species and 16
subspecies of Callicebus, based on nearly 1200 specimens.
Initial differentiation of the subspecies and species of
Callicebus was in coat display or recognition colors. The guide for
and measure of evolutionary change of mammalian tegumentary coloration
is described as metachromism. Three modes of metachromatic
differentiation are described and illustrated cladistically in 3 color
plates. The concept of forest refuges as centers of differentiation is
discussed and shown to be inapplicable to Callicebus. This paper
was a contribution to the International Symposium on Speciation, held
5-8 November 1987, in celebration of the 175th anniversary of the
Academy of Natural Sciences of Philadelphia.
-------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
* * *
LTC Larry D. Brown, Div. of Experimental Therapeutics, Walter Reed Army Inst. of Research, Washington, DC 20307-5100.
Charles Bruce, Dept. of Neuroanatomy, Yale Medical School, 333 Cedar St., New Haven, CT 06510.
Col. David E. Davidson, Jr., Div. of Experimental Therapeutics, Walter Reed Army Inst. of Research, Washington, DC 20307-5100.
Capt. Judith A. Davis, 4119 Sampson Rd., Wheaton, MD 20906.
John Duktig, Smith, Kline & French Labs., Box 1539 L-70, King of Prussia, PA 19406
J. G. Else, Yerkes Regional Primate Center, Emory University, Atlanta, GA 30322.
John E. Fa, 77 Irishtown, Gibraltar.
William I. Gay, 5200 Chandler St., Bethesda, MD 20814.
Maj. Roger C. Harvey, USAFSAM/VSR, Brooks AFB, TX 78235-5301.
Dick R. Haskin, Foundation for Primate Research and Conservation, P.O. Box 26, Royal, NE 68773.
Mack Holt, USAS/SAMUSP, Brooks Air Force Base, San Antonio, TX 78235.
Martin Kriete, 4879 Battery Lane, Apt. 32, Bethesda, MD 20814.
Charles A. Montgomery, Director, Baylor Animal Program, Baylor College of Medicine, Houston, TX 77030. Michele A. Papero, 333 Cedar St., Yale School of Medicine, New Haven, CT 06511.
SANOfi Recherche, 195 Route d'Espagne, 31036 Toulouse, France.
George S. Ward, Lab. Animal Breeders & Services, P.O. Box 557, Yemassee, SC 29945.
Sarah Williams-Blangero, Dept. of Genetics, Southwest Foundation for Biomedical Research, P.O. Box 28147, San Antonio, TX 78284.
* * *
Birth Announcement
Portrait: of Associate Editor James S. Harper and his daughter.
Lillian Beth Harper
9 September l989
7 pounds, 15 ounces
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. (Phone: 401-863-2511)
Judith_Schrier@brown.edu
ACKNOWLEDGMENTS
The Newsletter is supported by U. S. Public Health
Service Grant RR-00419 from the Animal Resources Program,
Division of Research Resources, N. I. H.
Cover drawing of an infant marmoset (Callithrix jacchus) by Penny Lapham.
Copyright © 1989 by Brown University
Editor: Judith E. Schrier, M. Sc.
Associate Editor: James S. Harper, D.V.M.
Consulting Editor: Morris L. Povar, D.V.M.
Founding Editor: Allan M. Schrier, Ph.D.
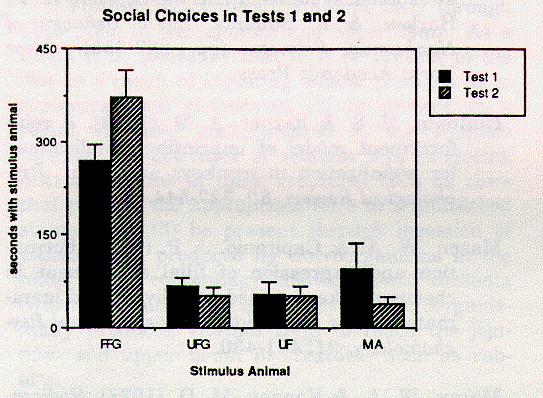{kind=link}
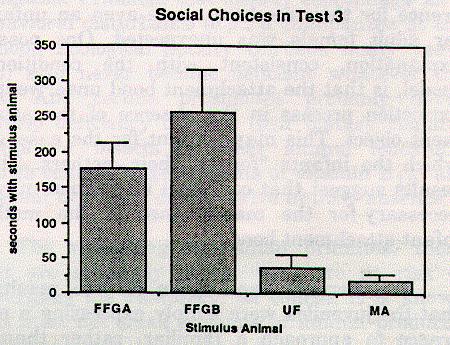{kind=link}
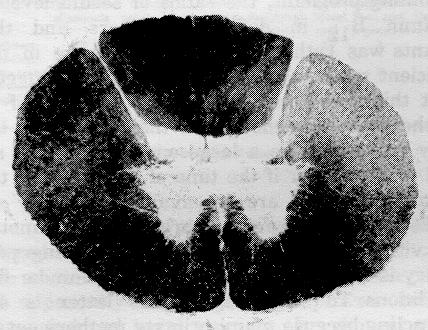{kind=link}
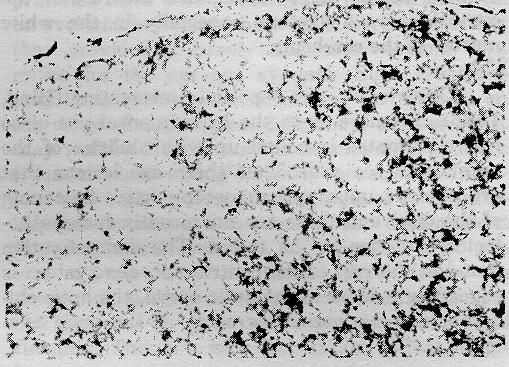{kind=link}
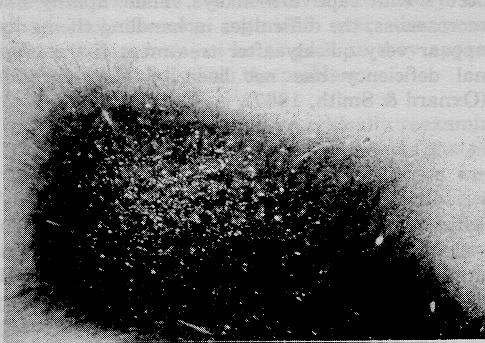{kind=link}
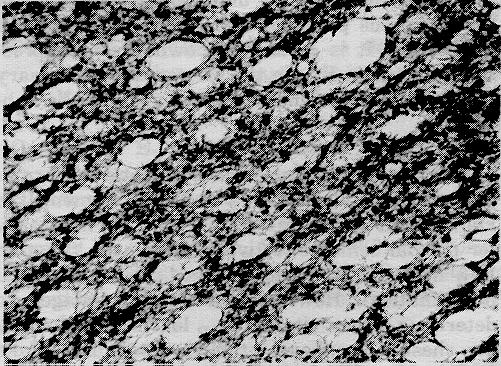{kind=link}
{kind=link}
{kind=link}
{kind=link}