 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 31 NUMBER 2 APRIL 1992
Articles and Notes
Changes in Male-Female Interactions after Introduction of a New Adult Male in Vervet Monkey (Cercopithecus aethiops) Groups, by H. S. Morland, M. A. Suleman, & E. B. Tarara...... 1
Nonhuman Primate Socialization and Environmental Enrichment Using a Transfer Tunnel, by K. J. Field, J. Denny, & G. Kubicz...... 5
Environmental Enrichment Branches That Do Not Clog Drains, by V. Reinhardt...... 8
Protection of a Disabled Group Member in Hamadryas Baboons, by R. C. Kyes...... 9
Foraging for Commercial Chow, by V. Reinhardt...... 10
Alternative Forage Types for Captive Chimpanzees, by L. Grief, J. Fritz, & S. Maki...... 11
Object Rubbing in Balinese Macaques (Macaca fascicularis), by A. Fuentes...... 14
News, Information, and Announcements
Editors' Notes: Correction; Cayo Santiago Rhesus Macaques...... 4
Travellers' Health Notes: Cholera Update...... 4
Fyssen Foundation 1992 International Prize...... 7
Environmental Enrichment Education and Training Resource...... 13
News Briefs...... 15
. .
South African Research Group Disbands; ABS Recruiting;
Oops, Wrong Primates
Grants Available...... 15
. .
NRC Associateship Programs; AmFAR Grants Available;
Biomedical Research Support Program; Restitution of
Neurological Motor Control
Information Requested and Available...... 16
. .
International Directory: Progress Report;
Potto Studbook; Self-injurious Behavior in Callitrichids;
Primate Surgical Anatomy; AAALAC Animal Well-Being Poster;
A.P.A. E-mail List
Meeting Announcements...... 18
. .
NABR Fall Conference; SCAW Conference;
Symposium on Models for AIDS; NY Regional Group Meetings
Directory of Graduate Programs: Addendum...... 18
Research and Education Opportunities...... 19
. .
Wildlife Preservation Trust; ASP Summer Internship List;
Fogarty/Japan Postdoctoral Program; Summer Pathology
Program; Volunteer for Field Study; School for Field Studies
Ben, My Ten-Year-Old Son, Discovers Henry Purcell on the Evolutionary Chain of Being, by K. T. Wolman...... 28
Departments
Address Changes...... 4
Positions Available: Ankara, Turkey; Ciba-Geigy, Switzerland...... 7
Recent Books and Articles ...... 20
* * *
Hilary Simons Morland, Mbarak A. Suleman, and Erna B. Tarara
Institute of Primate Research
Introduction
Captive macaque and vervet monkeys often attack, wound, or kill conspecifics during formation of new groups and during introduction of unfamiliar animals to stable groups. In macaque groups, adult males usually inhibit aggression among females (e.g., Erwin, 1978), but females occasionally attack and wound males during group formation (e.g., MacArthur et al., 1978). Aggression can be even more intense in captive vervet groups (e.g., Kaplan, 1987). Female vervets form coali- tions to chase, attack, and wound adult males (see Bloomstrand & Maple, 1987). Serious fighting between females was observed during group formation in the colony of vervet monkeys at the Institute of Primate Research (IPR) in Nairobi, Kenya (Else, 1985).
In our study at the IPR to develop the vervet monkey as a model for heterosexual transmission of African green monkey simian AIDS (SIVagm), the protocol required removing and replacing the adult male in one-male groups. This paper compares social interactions between males and females in these groups before and after adult males were replaced.
Methods
The study involved 84 vervet monkeys (Cercopithecus aethiops) housed in 7 one-male groups, each including 4 to 7 adult females from one to 3 matrilines, and 1 to 7 colony-born juveniles and infants. Adult females were trapped in Kenya or colony-born, and had been in the same cages for more than 1 yr. Adult males were wild-born and had been resident in the same cage between 2.6 and 6 yr. Vervets were dye-marked with picric acid for identification. Groups were housed in outdoor cages (3 x 6 x 2.5 m) made of one-inch weld mesh on a wooden frame, with gravel floor and partial overhead cover (Else, 1985). Commercial monkey diet (Unga Feeds Ltd., Nairobi) and supplements of fresh fruit or vegetables were provided daily. Water was available ad lib.
Behavior sampling (Martin & Bateson, 1986) was conducted on the seven vervet groups for a total of 292.5 hr. Each group was observed for 6 to 8 hr/month during six months distributed between 25 August 1989 and 24 May 1990. During 30-min morning and afternoon observation sessions, all occurrences of the following behaviors were recorded: 1) Sexual activity: presenting, ano-genital inspection, grabbing hips, soliciting of copulation by female, mounting, copulation with ejaculation; 2) Aggression: staring, chasing, grabbing, biting; 3) Grooming (Struhsaker, 1967). Identities of both initiator and recipient were recorded.
On 9 February 1990 (month 5 of the study), the resident adult males were removed from five "experimental cages" and replaced with new adult males. Three of the novel males were transferred from adjacent cages in the colony, while the other two had been housed in solitary cages for 8 to 9 months. In two "control cages", where no male exchange was made, one male had been present for 5 yr and one for 2.5 yr. Mean rates of grooming, sexual activity, and aggression per hour per monkey were computed for two periods: 1) Months 1 to 4: all seven groups contained a "resident" male who had been present for at least 2.5 yr, and 2) Months 5 and 6: the resident male was removed and replaced with a new male in the five experimental cages.
Results
Behavioral changes: In all of the experimental cages, females groomed newly introduced males less often than they had groomed resident males in the previous month (mean rate[month 4] = 2.35; mean rate[month 5] = 0.96), while females in the control cages continued to groom resident males frequently (mean rate[4] = 3.13; mean rate[5] = 4.09).
Figure 1: Mean rates of sexual activity for (a) control cages with resident males, months 1-6; (b) experimental cages with resident males, months 1-4; (c) experimental cages with new males, months 5-6.
Females initially engaged in little sexual activity with newly introduced males (Figure 1). No newly introduced males mounted or copulated in month 5, although one resident male mounted twice in this period. Two new males each mounted females once in month 6. It is not possible to say whether new males failed to copulate because females refused them, or because females simply were not cycling. Vervet monkeys in the IPR colony mate during most months of the year, but show a distinct mating peak in June to August (Else, 1985). In our study more sexual activity was observed in September-November (months 1 to 3) than between January and May. In the two control cages, resident males performed 41 mounts and copulations in September-November, but only 5 in January-May.
Aggression increased dramatically in experimental groups after new males were introduced (Figure 2). Females, both individually and in coalitions, repeatedly harassed, chased, and attacked newly introduced males. Females directed more aggression toward new males in month 5 than toward resident males in the previous months (sign test n(+) = 0, n = 7, p = 0.025). Newly introduced males were involved in about twice as much of the total aggression in their cages as their predecessors in the previous month (mean[4] = 31.3%; mean[5] = 65.6%).
Figure 2: Mean rates of aggression for (a), (b), and (c), as above.
The high rates of aggression observed in months 1 to 3 probably were related to frequent sexual activity in those months. In month 1, the two cages with the most sexual activity also had the highest rates of aggression.
Female aggression to newly introduced males varied considerably among cages. Two variables appeared to influence this. First, newly introduced males received more aggression when put into cages containing females belonging to one matriline (mean rate = 0.63) than when put into cages with two or three unrelated adult females (mean rate = 0.17). Second, dependent infants under 6 months were present when new males were introduced in four of the five cages. Comparing two experimental cages illustrates the importance of these variables. Two cages had 8 or 9 females and juveniles, and received new males who came directly from solitary housing. The male (#1227) introduced to the cage containing one matriline and four young infants received more aggressive acts than any other new male (mean rate = 2.08), and later died (see below). In contrast, the male introduced to a cage containing 3 matrilines and no young infants received the least aggression among new males (mean rate = 0.13).
Wounds and deaths: Within 6 weeks after introduction, 4 of 5 new males had been treated for bite wounds at least once. Within 12 weeks after introduction, all new males had been treated for wounds 1 to 4 times. No resident males were treated for wounds in this period.
Two of the five males died within 16 weeks after being introduced to new cages. One, known to be antibody positive for SIVagm, died on 26 April with nephritis, pneumonia, and pericarditis. The other (#1227) had been housed with females who were antibody- and virus-positive for SIVagm before he died on 7 June with multiple skin wounds, lymphadenopathy, gastric ulcers, and trauma-related septicemia (Emau, personal communication). We do not know whether these deaths were SIV-related. The degree to which SIVagm is immune-suppressive in vervets, and its role, if any, in contributing to trauma or disease related mortalities must be investigated.
In summary, all new males initially were harassed and attacked by female and juvenile vervets. Novel males were socially isolated and initially engaged in few or no affiliative interactions, but most eventually integrated into their new groups. Female aggression toward new animals was not restricted to males. Two attempts to move adult females between groups during our study failed because newcomers were immediately chased, attacked, and wounded by resident females.
Discussion
As reported in other colonies of vervet monkeys and in wild groups (Fairbanks & Maguire, 1987), females in the IPR colony formed coalitions against new adult males and were able to attack and seriously wound them. In contrast, a study on one macaque group that, like ours, involved replacing the resident male with a new male, found that females groomed and mated with the novel male more frequently than they had with his predecessor (Jones et al., 1982). Group size, female density, prior male social experience, and enclosure size influence levels of aggression in some macaque groups (e.g., Bernstein et al, 1974), but none of these variables were important in the IPR vervet monkey groups.
In the vervets, differences in female tolerance of new males probably were related to two variables: 1) the number of female matrilines in the cage, and 2) the presence of dependent infants under 6 months old. Females from the same matriline may have been more likely to form coalitions against males. Young infants seemed to precipitate female attacks on males because they tended to give distress calls when males approached. Fairbanks and McGuire (1987) found vervet mothers in their colony were more protective of dependent infants in the presence of newly introduced males. Our limited results support their view that vervet females form coalitions to counter a potential threat of infanticide from new males.
Our results were surprising since Else (1985) had no difficulty
exchanging adult males between groups or introducing new males in the
IPR colony during its early history. Our study demonstrates the risks
associated with introducing new adult males to small vervet monkey
groups. When adult males must be exchanged between one-male groups to
avoid father-daughter matings, our experiences suggest two precautions
that may help reduce aggression and facilitate acceptance of new
males: :
*Don't introduce new males into groups containing dependent
infants (Fairbanks & McGuire, 1987).
*Don't introduce males into
groups containing only one matriline.
References
Bernstein, I. S., Gordon, T. P., & Rose, R. M. (1974). Factors influencing the expression of aggression during introductions to rhesus monkey groups. In R. E. Holloway (Ed.), Primate Aggression, Territoriality, and Xenophobia (pp. 211-240). New York: Academic Press.
Bloomstrand, A. & Maple, T. (1987). Management and husbandry of African monkeys in captivity. In E. L. Zucker (Ed.), Comparative Behavior of African Monkeys (pp. 197-234). New York: Alan R. Liss, Inc.
Else, J. G. (1985). Captive propagation of vervet monkeys (Cercopithecus aethiops) in harems. Laboratory Animal Science, 35, 373-375.
Erwin, J. (1978). Factors contributing to intragroup aggression in captive pigtail monkey groups. In D. J. Chivers & J. Herbert (Eds.), Recent Advances in Primatology (pp. 581-583). New York: Academic Press.
Fairbanks, L. A. & McGuire, M. T. (1987). Mother-infant relationships in vervet monkeys: Response to new adult males. International Journal of Primatology, 8, 351-366.
Jones, E., Byrne, B., & Chance, M. R. A. (1982). Influence of a novel male on the social behavior of a captive group of mature female long-tailed macaques (Macaca fascicularis). Laboratory Animals, 16, 208-214.
Kaplan, J. (1987). Dominance and affiliation in the cercopithecini and papionine: A comparative perspective. In E. L. Zucker (Ed.), Comparative Behavior of African Monkeys (pp. 127-150). New York: Alan R. Liss, Inc.
MacArthur, J. A., Seamer, J. H., & Veall, D. (1978). Establishment of a small breeding colony of rhesus monkeys. Laboratory Animal, 12, 151-156.
Martin, P. & Bateson, P. (1986). Measuring Behaviour. Cambridge: Cambridge University Press.
Struhsaker, T. (1967). Behavior of Vervet Monkeys (Cercopithecus aethiops). Berkeley: University of California Press.
-------------------------------------------------------------------
First author's address: 4301 N. Ocean Blvd., #606A, Boca Raton, FL
33431.
This research was supported by the Contraceptive Research and
Development Program (CONRAD #CSA-89-049). We thank M. A. Isahakia, P.
A. Marx, P. Emau, and M. Otsyula for discussions and guidance during
this project. O. Odhiambo continued behavioral observations on the
vervet groups after May, 1990, and gave us insights about behavioral
problems. J. Martin compiled records on medical histories. D. Chai
provided veterinary services, and E. Muthoni and A. Gettie supplied
technical support.
* * *
Karl J. Field, Jack Denny, & Genowefa Kubicz
Hoffmann-La Roche Inc.
Introduction
A variety of novel cage designs or devices have been used to enrich the environments of captive nonhuman primates (NHP). Some of the more common strategies include releasing the animal into an exercise pen, using a puzzle feeder or forage feeding area to satisfy food gathering instincts, co- or gang-housing, and providing access to toys (Bryant et al., 1988; Line, 1987; O'Neill, 1988). At Hoffmann-La Roche, we use a vertical stainless steel transfer tunnel to provide an enlarged and novel cage design, to promote socialization, and to enrich the individually housed NHP's environment.
In the present report we describe our experiences and observations while using the vertical transfer tunnel to house cynomolgus monkeys (Macaca fascicularis).
Materials and Methods
Subjects: The subjects for this study were three adult male (M1, M2, M3) and one adult female (F1) cynomolgus monkeys. Body weights ranged from 4.0 (female) to 8.6 (male) kg. Animals were routinely housed individually in cages on a four-unit rack (stainless-steel, wire bottom). Each cage was 30 inches high and provided 4.3 sq. ft. of floor space.
Apparatus: The transfer tunnels (Figure 1) were in use for approximately six months prior to this study. The tunnels (Allentown Caging Equipment, Inc.) were attached to a top and bottom cage so that each animal was provided with free access to the tunnel and either cage. Tunnels were attached to the cages for a minimum of 24 consecutive hours, and were removed only for cleaning and animal handling.
The animals' use of the tunnels was recorded with a video cassette camera that was programmed to simultaneously record two animals for two continuous hours. Animals were acclimated to the presence of the videotape recorder in the animal room for one week. Feeding, cleaning, and animal facility activities were kept routine throughout the recording period. Recordings were made in the morning prior to feeding and in the afternoon prior to total darkness (lights on at 0600 and off at 1800). Approximately 4 hours of video tape time per animal were obtained and evaluated.
Tunnel Use Scoring: The following parameters were determined for
each animal:
* percent of time in the tunnel (top vs bottom);
* percent of time in a cage (top vs bottom);
* number of trips up and down the tunnel;
* other significant observations.
Results
+------+----------------------+---------------------+------+
| |Percent time in tunnel| Percent time in cage|Trips/|
|Animal| Top Bottom Total | Top Bottom Total | Hour |
+------+----------------------+---------------------+------+
| M1 | 71 11 82 | 18 0 18 | 6 |
| M2 | 81 10 91 | 4 5 9 | 2 |
| M3 | 74 17 91 | 4 5 9 | 15 |
| F1 | 76 15 91 | 6 3 9 | 17 |
+------+----------------------+---------------------+------+
| Mean | 76 13 89 | 8 3 11 | 10 |
+------+----------------------+---------------------+------+
Table 1. Percent of time at specific locations, and and trips per hour up and down the transfer tunnel.
Results are summarized in Table 1. The monkeys spent an average of 89% of their time in the tunnel (range 82-91%) and 11% of their time (range 9-18%) in a cage. Animals averaged 76% of their time at the top of the tunnel and 13% at the bottom. Similarly, 8% of their time was spent in the top cage and 3% in the bottom.
The monkeys averaged 10 trips/hour (range 2-17) up and down the tunnel. The male and female housed in adjacent cages (M3 and F1) used the tunnel in unison: when one climbed up or down the other would follow. In contrast, the two males housed in adjacent cages (M1 and M2) tended to ignore one another. However, on occasion M1 (the alpha male in the room) would taunt and threaten M2 and others in the room. In addition, all of the animals in this study socialized through gestures and vocalizations with animals on either side of their cages, and with those across the room.
A majority of the time in the tunnel was spent self-grooming or as inactive time, hanging horizontally from the top of the tunnel, playing with toys (dropping toys down the tunnel and retrieving them), or climbing up and down the tunnel. The animals also sat on the upper ledge of the tunnel, facing into the cage to self-groom or rest. They generally only entered the cage to self-groom or to eat. On occasion they would close their cage doors after entering or leaving the cage. In addition, the animals would play with the tunnel latch or the cage door.
Discussion
We believe that the transfer tunnel enriches the NHP's environment immensely when compared to standard single cage housing methods. The tunnel provides the NHP's with an expanded novel vertical space, the ability to socialize with animals on either side of their cage, and their choice of a top or bottom cage. In addition, animals were free to access the tunnel at any time, thus exerting a control over their environment that is not available when the transfer tunnel is not in use. This factor alone is considered to be a key element of an environmental enrichment program, and is postulated to be crucial for improving the animals' well-being (Line, 1987).
Since conducting this study, we have used the tunnel to provide the
following changes in the animals' environment:
* To provide a forage feeding area in a bottom cage, and routine housing in a top cage.
* To introduce, acclimate, and co-house animals. Introduction is
accomplished by pushing cages close together and allowing animals to
touch one another between cages. Compatible animals are then
cohoused. When cohoused, animals tend to remain in one cage
together.
* Puzzle feeders and cage toys have been attached to the outside of
the tunnel at the top and bottom.
Conclusions
* Even after extended acclimation, the monkeys do not become bored or
complacent with the vertical space. In our study, animals continued
to use the tunnel 89% of the time.
* The tunnel provides an expanded cage size and novel cage design that
the NHPs can access as they please. The additional space is both
vertical and horizontal, allowing the animals to choose the location
and type of space they wish to use. In our study, the majority of
their time (76%) was spent at the top of the tunnel self-grooming or
resting quietly.
* The monkeys obtain exercise climbing up and down the tunnel (10
trips/hour in our study). The tunnel can be used to encourage exercise by feeding animals in the top and bottom cages, requiring them
to collect their food from different locations.
* Animals in the tunnel extend further out into the room, and thus are
able to see and socialize more easily with neighboring animals.
References
Bryant, C. E., Rupniak, N. M., & Iversen, S. D. (1988). Effects of different environmental enrichment devices on cage stereotypies and autoaggression in captive cynomolgus monkeys. Journal of Medical Primatology, 17, 257-269.
Line, S. (1987). Behavioral and physiologic response of the rhesus macaque to an environmental enrichment device. Laboratory Animal Science, 37, 509.
O'Neill, P. (1988). Developing effective social and environmental enrichment strategies for macaques in captive groups. Laboratory Animal, 17, 23-36.
-------------------------------------------------------------------
Authors' address: Hoffmann-La Roche Inc., Dept. of Laboratory Animal Resources, 340 Kingsland St., Nutley, NJ 07110.
* * *
Viktor Reinhardt
Wisconsin Regional Primate Research Center
Deciduous tree branches offer inexpensive, effective environmental enrichment for macaques (Reinhardt et al., 1987; Champoux & Reinhardt, 1987; Reinhardt, 1990a). Properly installed, branches increase usable cage space by enabling the animals to perch on an elevated dry loca tion with optimal visual control of the environment outside of the cage (Reinhardt, 1991). If loose segments are offered, the animals manipulate and gnaw them (Reinhardt, 1990b).
Deciduous tree branches permit caged macaques to express noninjuri-ous species-typical behavior in accordance with Federal Rules (USDA, 1991, sec 3.81,b), but they may at the same time create maintenance problems by clogging sewer drains. Box elder (Acer negundo), silver maple (A. saccharinum) black cherry (Prunus serotina), white oak (Quercus alba), black locust (Robinia pseudoacacia), white elm (Ulmus americana), weeping willow (Salix babylonica), and other deciduous tree species have been tested here for several years with the consistent observation that animals do not merely gnaw but actively tear off strips of bark and wood. Inevitably, this clogs the drains in the animal rooms if many monkeys are simultaneously engrossed in shredding branch material.
Recent testing has shown that red oak (Quercus rubra) branches, unlike other deciduous tree branches, are not shredded by macaques though they are equally attractive to the animals for gnawing and perching. When gnawed, the branches gradually wear into flakes of bark and wood, so small that even large quantities pass through drains without clogging (Figure 1). More than 700 caged rhesus (Macaca mulatta) and stump-tailed (M. arctoides) macaques housed in 29 rooms have been exposed to red oak perches and/or loose branch segments for a period of six months. Drains did not clog in any of the 29 rooms during this time although the animals gnawed the wood extensively (Figure 1).
Figure 1: Different stages of wear of red oak branch segments.
References
Champoux, M., Hempel, M. & Reinhardt, V. (1987). Environmental enrichment with sticks for singly-caged adult rhesus monkeys. Laboratory Primate Newsletter, 26[4], 5-7.
Reinhardt, V., Houser, W. D., Cowley, D. & Champoux, M. (1987). Preliminary comments on environmental enrichment with branches for individually caged rhesus monkeys. Laboratory Primate Newsletter, 26[1], 1-3.
Reinhardt, V. (1990a). Comparing the effectiveness of PVC perches versus wooden perches as environmental enrichment objects for singly caged rhesus monkeys. Laboratory Primate Newsletter, 29[1], 13-14.
Reinhardt, V. (1990b). Environmental enrichment program for caged stump-tailed macacques (Macaca arctoides). Laboratory Primate Newsletter, 29[2], 10-11.
Reinhardt, V. (1991). An environmental enrichment program for caged rhesus monkeys at the Wisconsin Regional Primate Research Center. In M.A. Novak & A.J. Petto (Eds.), Through the Looking Glass. Issues of Psychological Well-being in Captive Nonhuman Primates (pp. 149- 159). Washington, DC: American Psychological Association.
U.S. Department of Agriculture (1991). Animal Welfare, Standards, Final Rule. Federal Register, 56, 6499-6500.
-------------------------------------------------------------------
Author's address: Wisconsin Regional Primate Research Center, 1223 Capitol Court, Madison, WI 53715.
* * *
Randall C. Kyes
Bowman Gray School of Medicine
Reports of nonhuman primates helping or protecting injured or disabled group members are infrequent and generally anecdotal. Although difficult to assess systematically, protective behavior in nonhuman primates may have important social significance and should be documented when possible. A recent observation of protective behavior in a langur colony was reported by Avallone and Johnson (1992). A similar incident was observed 18 June 1982, involving the protection of a temporarily disabled group member in a small group of baboons located at Bucknell University.
The group was a family unit of six hamadryas baboons (Papio hamadryas), composed of two parents: M(male)1 (17 yrs) and F(female)1 (17 yrs), and four offspring: F2 (6 yrs), M2 (5 yrs), F3 (4 yrs), and M3 (3 yrs, a paternal half-sibling). The group was housed in a 12x12x4.5 m outdoor field cage. At about 14:00 hours, I observed M3 having a grand mal type epileptic seizure (although M3 was never clinically diagnosed as having epilepsy). I was observing the group from a window in a nearby lab building and they were unaware of my presence. M3 was about 1.5 m up the chain link wall at the front of the cage when the seizure began. Within several seconds, M3 fell to the ground on his back, 1 m from the front of the cage. About 10 seconds later, M2 moved across the cage to within .5 m of M3 and directed most of his attention toward M3. The remaining group members also directed their attention toward M3 but did not change their original positions (all were at least 3 m from M3). The convulsions continued for about 1.5 minutes.
When the convulsions ended, I left the lab to check M3's condition. My presence was first noticed by the group as I exited the lab (about 10 m from the front of the cage). I approached at a normal pace. M3 was lying on his back showing no movement and apparently unconscious. When I was about 4 m from the front of the cage (5 m from M3), M2 moved toward M3, placed his hand on M3's chest, and directed several threats toward me (eyebrows raised and mouth open). F2 and F3 also threatened me, but did not change position. M1 and F1 observed my approach but showed no response. At that point, I slowed my pace but continued to approach; M2 increased his threats. When I was within one meter of the front of the cage (and about 2 m from M3), M2 lunged at me, threatened both visually and vocally, and grabbed at me through the chain link. F2 and F3 joined M2 at this point -- threatening me from a distance of about 2 m. This situation continued for about 10 seconds, until I stepped back about 2 m. M2 then moved back to M3 (who was still unconscious), took him by the left forearm, and dragged him away from the front of the cage about 1 m (about 5 m from me). For the next 3.5 minutes, M2 remained next to M3, occasionally threatening me. I remained standing about 3 m from the front of the cage (5 m from M3). M3 then regained consciousness, staggered (alone) toward the back of the cage, and sat down. The entire incident, from onset of the seizure until M3 regained consciousness, lasted about 7 minutes. Throughout the episode, M1 and F1 showed little interest or concern.
The behavior directed toward me during this incident was uncharacteristic of my interactions with the baboons. I had worked closely with the group for several months prior to this incident and had never experienced an aggressive display of this nature.
Although I cannot confirm that this was the first seizure experienced by M3, I am not aware of any previously reported incidents. M3 did experience several subsequent seizures which I observed. In each case, M2, F2, and F3 approached M3 and occasionally touched him. None of these seizures, however, resulted in the level of protective behavior described above. It is important to note that during each of the subsequent seizures, M3 was near the back of the cage and partially hidden by large rocks. Perhaps the degree to which a disabled group member is aided or protected is proportional to the degree of danger that individual is perceived to be in by the other group members.
Reference
Avallone, M. A. & Johnson, S. (1992). "Paternalistic" behavior in a langur colony. Laboratory Primate Newsletter, 31[1], 18.
-------------------------------------------------------------------
Author's address: Dept. of Comparative Medicine, Comparative Medicine Clinical Research Center, Bowman Gray School of Medicine, Medical Center Blvd., Winston-Salem, NC 27157.
* * *
Viktor Reinhardt
Wisconsin Regional Primate Research Center
There is general agreement that offering laboratory nonhuman primates freely accessible food may meet the animals' physiological need for nutrients but not their behavioral need for foraging. This issue has been addressed by federal rules, listing "foraging or task-oriented feeding methods" as recommended for promoting the expression of "noninjurious species-typical activities" in nonhuman primates (USDA, 1991). The following "food puzzle" has been implemented in compliance with these rules since January 1991 for two breeding troops living in different pens at the Wisconsin Regional Primate Research Center.
Both enclosures are covered with chain-link fencing material. The mesh is square, measuring 33 x 33 mm. Purina monkey chow (# 5037) is distributed, not on the floor or in feeder boxes, but at several locations (to avoid possible competition over access to food) on the mesh ceiling of the pens. The dry biscuit has a star-like shape with four spikes, about 15 mm long and 15 mm wide, protruding from a sphere measuring approximately 30 mm in diameter. These dimensions ensure that the chow cannot fall through the mesh, though spikes of it protrude (Figure 1). This creates an ideal foraging and task-oriented feeding method: While sitting on swings, platforms or other elevated structures, or clinging to the mesh, individual animals seize a piece of chow, press part of it through a mesh and finally break and retrieve a piece (Figure 1). Supplemental food such as fruit, vegetables and bread are distributed in the same way, to encourage further foraging activities.
This simple "food puzzle" not only promotes non-injurious species-typical foraging skills but also keeps the floor relatively clean by avoiding undue spoilage of food. The animals only work for food that they actually eat. Pieces falling on the floor are not wasted but picked up by animals that are too lazy to work for food themselves.
Reference
U.S. Department of Agriculture (1991). Animal Welfare; Standards; Final Rule. Federal Register, 56, 6495-6505.
-------------------------------------------------------------------
Author's address: Wisconsin Regional Primate Research Center, 1223
Capitol Court, Madison, WI 53715.
The environmental enhancement plan is supported by NIH grant
RR-00167 to the WRPRC. Publication number 31-042 of the WRPRC.
* * *
Laurie Grief, Jo Fritz, and Susan Maki
Primate Foundation of Arizona
Introduction
Psychological health is very important for primates in captivity. One way to define psychological wellness is to compare the behavioral repertoire of animals in captivity with those in the wild (Maki & Bloomsmith, 1989). To replicate species-typical environments and behaviors in captivity, environmental complexity is necessary. Chimpanzees' natural habitats are tropical forest, tropical grassland, and the transitional area between the two (Napier & Napier, 1985). The greatest portion of a chimpanzee's day is spent foraging for food peculiar to its ecological niche (Goodall, 1986). For captive chimpanzees, food is usually provided on a timed schedule to facilitate husbandry operations. Foraging activities are thus eliminated from the behavioral repertoire of the captive chimpanzee.
According to Spinelli and Markowitz (1985), we must return some of the power over its environment to the chimpanzee if we are to ensure behaviorally healthy animals. If we provide foods that make it necessary for the animals to forage throughout the day, then the temporal patterns of feeding bouts may resemble more closely the manner in which wild chimpanzees spend their days. Research has shown that longer periods of time spent foraging do increase general activity levels (Anderson & Chamove, 1984; Maki et al., 1989) and decrease boredom (Clarke et al., 1982; Maple, 1979; Maki & Bloomsmith, 1989) in captive chimpanzees.
At the Primate Foundation of Arizona we provide forage material daily for the chimpanzees. The chimpanzee's bedding, straw or shredded paper, mixed with sawdust, is continuously available to them and effectively hides the forage material. The purpose of this study was to test for a particular food or group of food items that would increase foraging time and also be cost effective. Size of the food item, its ability to be widely scattered and hidden in the bedding, nutritional value, and taste were all considered when we chose the forage types to be tested. The three foods that most appropriately fit these criteria are: 1) chicken scratch, for its hard consistency and small size; 2) sweet feed, for its intermediate size and softer consistency; and 3) popcorn, for its larger size and medium consistency.
Methods
Subjects: Subjects were 9 adult male and 13 adult female chimpanzees. Three males and two females were housed together in a social group and the remainder were housed singly, but with tactile, visual, and auditory contact with adjacent cagemates. Six of the females had an infant present with them, but the infants were not included in the analysis.
Procedure: Three types of forage material were tested: chicken scratch, sweet feed, and popcorn. The chicken scratch consisted of milo, cracked corn, and wheat. "Sweet feed" was rolled corn and barley with a small amount of molasses. The popcorn was unsalted and hot air popped. Each subject received equal amounts of forage whether in a social setting or housed singly. This did not guarantee that each subject housed in the social group actually received its allotted amount. Each type of forage material was presented on a weekly rotation basis with a different forage introduced each week. Forage material was distributed throughout the enclosures at 8 am daily.
Sampling and Data Analysis: This study took place from November 1989 through March of 1990. Observations were conducted between 0815 and 1330 hours, with subjects observed in randomized order. The scan sampling technique at one minute intervals was used to record the behavior of each of the subjects. Subjects were observed for three 15-minute sessions per day, two in the morning and one in the afternoon, three times a week for a total of nine hours of data on each subject. Foraging was the only behavior analyzed for this study and was defined as using the hand and/or mouth for searching through straw, sawdust, and paper for food items, transfering the forage items to the mouth, or chewing the forage material.
The Wilcoxon test was used to test for significant differences between subjects' data point scores for foraging of the three different forage types and to examine the pattern of foraging behavior during the day. We tested whether foraging declined faster for one type of food than for another, and whether one sex foraged longer in the day than the other. The Mann-Whitney U test was used to compare amounts of forage behavior between males and females and between the two different housing conditions.
Results
All of the subjects foraged for all three types of foods. Interestingly, one of the females, blind since birth, had one of the highest foraging scores of the 22 subjects. Of all the possible data points, chicken scratch had the highest percentage at 53.8%, followed by sweet feed at 30.8%, and popcorn at 15.4%.
+------+-----------+-------------+-----------+
| | Scratch 1)|Sweet Feed 2)| Popcorn |
+------+-----------+-------------+-----------+
|Range | 0.58-31.9 | 0.16-25.7 | 0.16-15.6 |
|Median| 17.45 | 8.62 | 5.25 |
|Mean | 17.17 | 10.44 | 6.33 |
+------+-----------+-------------+-----------+Table 1: Forage behavior of subjects in three experimental conditions (frequencies of scan sample data points).
Foraging occurred significantly more with chicken scratch than with either sweet feed (T =4, p < 0.001) or popcorn (T =1, p < 0.01). There was also a significant difference between sweet feed and popcorn (T =55, p < 0.01). Males (x=24) foraged significantly more than females (x=13) for chicken scratch (Mann-Whitney U=11, p < 0.001), but this sex difference was not observed for either sweet feed or popcorn.
There were no significant differences in time spent foraging between group- and singly-housed animals, for any food type. All subjects foraged for longer periods when chicken scratch was present. Examining the afternoon scores alone, there was a 40% reduction in the mean between chicken scratch and sweet feed, significant by the Wilcoxon test (T =20, p < .001). A 30% reduction in the mean (T =118, p < .001) between chicken scratch and popcorn was also observed. There were no significant differences in forage behavior in the afternoons between sweet feed and popcorn. The Wilcoxon test showed a significant difference (T =4, p < .01) between males (x=4.7) and females (x=2.35) for foraging behavior in the afternoons, but there were no significant gender differences in the mornings.
Discussion
Wild chimpanzees live in an extremely complex environment, constantly making decisions about where and when to feed. Distributing food materials for foraging throughout a captive primate's cage gives back to the animal some power over its environment: control over the time and manner of eating.
All of the animals engaged in foraging to some extent, no matter what food was available. Males spent more time foraging, and foraged until later in the day, suggesting that foraging is a particularly good way to stimulate captive male chimpanzees. Although chicken scratch proved to be the favorite (and least expensive) forage, all three had their benefits. Sweet feed was consistently searched for and consumed: its best use might be as an alternative, keeping the novelty effect higher. Popcorn also proved successful, but, because it was more visible than the other two feeds, was consumed more rapidly. It might be particularly useful as a short-term distraction, such as during cage cleaning.
Chicken scratch, because it is small and harder to find, elicited the most foraging of the three foods. It is very encouraging to see the chimpanzees still foraging late in the day for these small kernels. In addition, for subjects such as our blind animal, who had one of the highest foraging scores in our study, this enrichment cannot be overemphasized. Chimpanzees are social animals and group housing is preferable (Fritz, 1989), but when single caging is necessary, forage materials can provide a more complex environment without expensive manipulative devices.
References
Anderson, J. R., & Chamove, A. S. (1984). Allowing captive primates to forage. In Standards in Laboratory Animal Management (Part 2, pp. 253-256). Potters Bar, UK: Universities Federation for Animal Welfare.
Clark, A. S., Juno, C. M., & Maple, T. L. (1982). Behavioral effects of a change in the physical environment: A pilot study of captive chimpanzees. Zoo Biology, 1, 371-380.
Fritz, J. (1989). Resocialization of captive chimpanzees: An amelioration procedure. American Journal of Primatology, Supplement 1, 79-86.
Goodall, J. (1986). The Chimpanzees of Gombe. London: Belknap Press of Harvard University.
Maki, S., Alford, P., Bloomsmith, M., & Franklin, J. (1989). Food puzzle device simulating termite fishing for captive chimpanzees (Pan troglodytes). American Journal of Primatology, Supplement 1, 71-78.
Maki, S. & Bloomsmith, M. (1989). Uprooted trees facilitate the psychological wellbeing of captive chimpanzees. Zoo Biology, 8, 79-87.
Maple, T. (1979). Great apes in captivity: The good, the bad, and the ugly. In J. Erwin, T. L. Maple, & G. Mitchell (Eds.), Captivity and Behavior (pp. 239-272). New York: Van Nostrand Reinhold.
Napier, J. & Napier, P. H. (1985). Natural History of the Primates. Cambridge, MA: MIT Press.
Spinelli, J. & Markowitz, H. (1985). Prevention of cage-associated distress. Lab Animal, 14[8], 19-28.
-------------------------------------------------------------------
Authors' address: Primate Foundation of Arizona, P.O. Box 86,
Tempe, AZ 85280.
Laurie Grief was supported by a fellowship grant to the Primate
Foundation of Arizona from the E. Blois du Bois Foundation. The
authors would like to thank the Care Staff at the Primate Foundation
of Arizona for their assistance in completing this project. This study
was partially supported by the National Institutes of Health, Division
of Research Resources, grants R01-RR03578 and U42-RR03602.
* * *
Agustin Fuentes
University of California, Berkeley
Object rubbing, holding a leaf, paper, plastic, or cloth in both hands and moving it in a stereotyped back-and-forth motion on an object, has been seen by two observers (Chiang, 1967, and Wheatley, 1988) in Macaca fascicularis, but is not mentioned in major field studies of this species (e.g., Van Noordwijk, 1985). During three months of observation at Ubud, in Bali, Indonesia, in 1986, Wheatley (1988) reported 17 instances of object rubbing behavior in a group of 26 animals. Chiang (1967) reported a similar behavior pattern for four groups in the Singapore Botanical Gardens and termed it "leaf washing". He suggested the motor pattern was an isolated, local behavior pattern.
From May 1 to June 5, 1990, I observed a group of 55 Macaca fasicularis near Ubud for 40 hours. The group lived in a 4 hectare patch of forest surrounded by cultivated land. A local official provisioned the monkeys with about 10 kg of sweet potatoes daily. During this time I recorded rubbing of objects including food, rocks, and coconut shells. Food items were the objects of 40% of the rubbing observed. Brief observations were made on four other macaque groups on Bali. No object rubbing was seen at these other sites.
Thus, object rubbing behavior has been reported at two sites by three observers. Older infant, juvenile, sub-adult, and adult animals displayed this behavior.
Object rubbing by Ubud and Singapore long-tailed macaques is most likely not a "food-washing" behavior, as reported for Japanese macaques (Kawai, 1965). Of the 58 objects rubbed, a significant portion, 23 (Chi-square= 9.1, p < .05), were non-food items. Most of the rubbing occurred in dirt, and most of the materials used to rub (leaves, paper, etc.) were themselves covered with dirt, suggesting that rubbing was not performed to clean or prepare food items for eating.
Both food-washing and generalized rubbing behaviors are common to macaques (see, e.g., Visalberghi & Fragaszy, 1990). However, the incorporation of items such as leaves, paper, plastic, and cloth for rubbing, and the rubbing of many non-food items suggests that object rubbing as described here may be a distinct behavior. The motivation for performing this behavior is not apparent as the objects rubbed are often discarded, or are of no food value.
At Ubud, villagers from the surrounding area come daily to the small river in the forest to wash clothes and bathe. The monkey group is often in the same area, and many juveniles play and swim 15-20 m from the villagers. The Balinese women generally wash clothes with a two-handed back-and-forth rubbing motion, a pattern similar to the macaques' object rubbing behavior.
Both the Singapore groups observed by Chiang (1967) and the Ubud group were surrounded by human populations, and it is possible that the Singapore groups were exposed to conditions similar to those encountered by the Ubud group, including exposure to human clothes washing and bathing activity. This may have been a factor in the appearance of the object rubbing behavior at these two sites.
References
Chiang, M. (1967). Use of tools by wild macaque monkeys in Singapore. Nature, 214, 1258-1259.
Kawai, M. (1965). Newly-acquired pre-cultural behavior of the natural troop of Japanese monkeys on Koshima Islet. Primates, 6, 1-30.
Van Noordwijk, M. A. (1985). The socio-ecology of Sumatran long-tailed macaques (Macaca fasicularis): II. The behavior of individuals. Proefschrift. Utrecht: Drukkerij Elinkwijk.
Visalberghi, E. & Fragaszy, D. (1990). Food-washing behaviour in tufted capuchin monkeys, Cebus apella, and crab-eating macaques, Macaca fasicularis. Animal Behaviour, 40, 829-836.
Wheatley, B. P. (1988). Cultural behavior and extractive foraging in Macaca fasicularis. Current Anthropology, 29, 516-519.
-------------------------------------------------------------------
Author's address: Department of Anthropology, University of California, Berkeley, CA 94720.
* * *
Correction
On page 17 of the last issue (January, 1992), the listing of the original seven Directors of the Regional Primate Research Centers should have included Dr. Pickering, of the Oregon RPRC.
Cayo Santiago Rhesus Macaques
Jean Baulu has asked us to remind readers again about the avail- ability of Cayo Santiago rhesus macaques (this Newsletter, 1991, 30[4], 10). The colony has been maintained since 1938, and is free of tuberculosis, simian retroviruses, SV40, and measles. Contact Jean at the Caribbean Primate Research Center, Univ. of Puerto Rico, Medical Sciences Campus, P.O. Box 1053, Sabana Seca, PR 00749 [809-422-8826; FAX: 809-432-5339].
* * *
The epidemic of cholera that began in Peru in January 1991 continues to spread. Most recently, Bolivia, El Salvador, Honduras, Nicaragua, and Panama were added to the list of countries reporting cholera cases. Travelers who develop severe watery diarrhea or diarrhea and vomiting during or within 1 week after travel to an area with known cholera should seek medical attention immediately. We should remember that cholera continues to be endemic in Africa as well; there were more than 100,000 cases there in 1991, with about 15,000 deaths.
* * *
American Psychological Association, 750 First Street N.E., Washington, D.C. 20002-4242.
Aleksandar L. Knezevich, 24 Manners Rd., Ringoes, NJ 08551.
Judith Masters, 60 Second Ave., Westdene, 2092, South Africa.
Thomas K. Shehan, 1824 Hyman Lane, Crofton, MD 21114.
Donna A. Steiner, 5941 E. Vermillion St., Mesa, AZ 85205.
Charles Weisbard, 31-31 29th St., Apt. 3F, Astoria, NY 11106.
* * *
Ankara, Turkey
Middle East Technical University, Ankara, Turkey, Department of Biology, has a position open for an Animal Physiologist and an Animal or Marine Ecologist at the Assistant Professor level. Candidates should have the Ph.D. in Animal Biology, Ecology or Marine Ecology. All candidates should be able to teach courses in the English language, and are expected to cooperate with other department members. For more information please contact: Prof. Dr. Aykut Kence (Chairman), Dept. of Biology, Metu 06531, Ankara, Turkey [90(4) 223 71 00 Ext.3103; FAX: 90(4) 223 69 45; E-Mail: [email protected].
Ciba-Geigy, Switzerland
Applications are invited from ethologists or psychologists for an appointment as Head of the Primate Station within the Research Department of the Pharmaceuticals Division of Ciba-Geigy, Basel, Switzerland. Candidates should have extensive practical experience with direct observation of behavior in primates. The successful candidate will be in charge of the overall operation of this facility housing a colony of some 40 rhesus monkeys and will lead a team composed of one postdoctoral research associate and two technicians. The group is principally engaged in the pursuit of experimental projects designed to characterize the activity profiles of new psycho-pharmacological agents, e.g. anxiolytics and antidepressants, with respect to their influence on the behavioral pattern of monkeys in groups. Preference will be given to persons whose past performance shows evidence of unusual resourcefulness, intellectual agility and originality.
Qualified applicants should submit their curriculum vitae and a list of publications to: Ciba-Geigy Ltd, Personnel Department, Attn: Dr. H. Schmid, Postfach, CH-4002 Basel, Switzerland. For further information, please contact Dr. C. Mondadori, Head of the Behavioural Neurosciences Section, Ciba-Geigy Ltd [41-61-6966454].
* * *
The Fyssen Foundation's general aim is "to encourage all forms of scientific enquiry into cognitive mechanisms, including thought and reasoning, underlying animal and human behavior, into their biological and cultural bases, and into their phylogenetic and ontogenetic development." For this purpose, a substantial International Scientific Prize is awarded annually to a scientist who has conducted distinguished research in the areas supported by the Foundation. The subject under consideration for the 1992 Prize is Elementary Mechanisms of Cognitive Development. Nominations should include a curriculum vitae of the nominee; a list of his publications; a summary (4 pages maximum) of the research upon which the nomination is based. Deadline for receipt of nominations is September 1, 1992. For more information contact Fondation Fyssen, 194, rue de Rivoli, 75001 Paris, France [33 (1) 42 97 53 16; FAX: 33 (1) 42 60 17 95].
* * *
Under new federal regulations research institutions are required to take two steps toward assuring the psychological well-being of nonhuman primates in their care. The first step is to develop a general institutional plan to address psychological well-being issues for all animals. The second step is for the Institutional Animal Care and Use Committee (IACUC) to review proposed protocols to determine whether the psychological well-being of primates used in the proposed project has been adequately addressed.
With funding from the Edna H. Tompkins Trust, a regional resource has been established, based at Harvard Medical School's New England Regional Primate Research Center (HMS/NERPRC), to help individual reseachers and IACUCs meet these requirements. The chief objective of this program is to support and facilitate efforts to enhance the psychological well-being of captive nonhuman primates by advising individual researchers and IACUCs. The secondary objective is to extend the awareness of and attention to environmental enrichment for nonhuman primates in biomedical research projects by training and educational activities.
Program activities will include:
* Consultation -- a) Experienced,
expert consultation to individual researchers on laboratory environments for nonhuman primates and help in fashioning research protocols
that meet both regulatory and scientific requirements; b) advice to
animal research review committees and IACUCs on research protocols
that are affected by the federal psychological well-being regulations.
* Information -- The project staff will maintain and make available a
unique and wide-ranging bibliographic resource providing annotated
guides to current literature on laboratory primate environments.
*
Training -- The project staff will produce and present materials for
education and training to health care professionals, researchers,
technicians, and laboratory assistants.
For more information, contact Andrew Petto at NERPRC, P.O. Box 9102, Southborough MA 01772 [508-624-8089; FAX: 508-624-8190; e-mail: [email protected]].
* * *
South African Research Group Disbands
Professor G. A. Doyle, of the University of the Witswatersrand, Johannesburg, South Africa, reports that "the Primate Behaviour Research Group ceased to exist on 31 December, 1991. Research groups, such as mine was, go out of existence when the persons around which they are built retire. I retire at the end of this year, but I'm on sabbatical for the entire 1992. In anticipation of my retirement we've been winding down for the last few years, i.e., not taking on any new students, etc."
ABS Recruiting
The Animal Behavior Society is encouraging zoologists, psychologists, zoo keepers, behavioral geneticists, etc, to consider joining A.B.S. For more information, contact Ira Perelle, Membership Chairman, A.B.S., Mercy College, Dobbs Ferry, NY 10522.
Oops, Wrong Primates
The University of Wisconsin's Regional Primate Research Center sent Archbishop George Cram, of the Anglican Church of Canada, a questionnaire while preparing an international directory of primatology. The envelope was addressed to "George Cram, Primates World Relief and Development Fund, Toronto, Canada."
Rev. Michael Ingham, secretary for the senior archbishop, suggested in a letter of reply that "primates in your study are perhaps of a different species. While it is true that our primate occasionally enjoys bananas, I have never seen him walk with his knuckles on the ground or scratch himself publicly under the armpits," Ingham said. "There are a mere 28 Anglican primates in the whole world," he added. "They are all males, of course, but so far we have had no problems of reproduction."
* * *
NRC Associateship Programs
The National Research Council Associateship Program announces three annual competitions for scholarly awards for basic research in, among others, biological, health, and behavioral sciences and biotechnology. The Council in cooperation with participating federal laboratories will offer approximately 450 awards in 1992 for independent scientific research. Recipients will be free to work on problems of their own choice as guest investigators at the participating federal laboratories. Many of the programs are open to experienced senior scientists as well as to recent Ph.D's. Annual stipends for recent graduates range from $27,150 to $38,000 and are appropriately higher for seniors. Relocation reimbursement and funds for professional travel are available. A health insurance program is offered. Deadlines are January 15th, April 15, and August 15. For further information on participating laboratories and application materials contact: Associateship Programs, National Research Council, 2101 Constitution Avenue, NW, Washington, DC 20418 [FAX: 202-334-2759].
AmFAR Grants Available
The American Foundation for AIDS Research (AmFAR) awards Research Grants for biomedical and humanistic investigations, short-term Travel Grants, three year Scholar Awards and Pediatric AIDS Foundation grants. For further information and applications, call the AmFAR Grants Department at 213-857-5900, or write to AmFAR Research, Grants Department, 5900 Wilshire Boulevard, Second Floor, East Satellite, Los Angeles, CA 90036-5032.
Biomedical Research Support Program
The National Center for Research Resources announces the Biomedical Research Support Grant (BRSG) Program for Fiscal Year (FY) 1992. Because the FY 1992 appropriation of $5,204,000 for the BRSG Program is a significant reduction in the amount of funds available, awards will be made on a competitive basis and fewer awards will be made.
The objectives of the BRSG Program for FY 1992 have been narrowed to focus on support for biomedical research projects in ways that are not adequately or efficiently provided for by other funding mechanisms such as the traditional investigator-initiated research project grant. The allowable uses of BRSG funds for FY 1992 are: * Pilot research. * New investigators. * Unexpected requirements and opportunities for projects supported by other NIH funding mechanisms. This includes funding to enable research programs to continue during temporary lapses in project grant support.
Institutions eligible to apply must have received a minimum of three allowable Public Health Service (PHS) research grants and/or cooperative agreements, totalling $200,000 (including direct and indirect costs), awarded by the PHS during FY 1991. Application receipt date is June 15, 1992. Direct inquiries to: BRS Program Office, Westwood Building, Room 10A11, Bethesda, MD 20892 [301-496-6743].
Restitution of Neurological Motor Control
The National Institute of Neurological Disorders and Stroke (NINDS), a component of NIH, invites research grant applications for support of research on neurological motor control, motor control disorders, and the restitution of function. Applications covering a broad range of activities in the neurological sciences from basic research to clinical research, focused on normal and abnormal performance of the nervous system in regard to motor control, are encouraged. Examples of research objectives appropriate for inclusion in applications responsive to this PA include: * Studies of the neurophysiological foundation of motor control; * Studies of the biomechanics and neuromuscular physiology of motor control including conscious and automatic control in humans or animals; * Studies of the role of sensory systems in motor control; * Studies of the electrophysiological, neuroendocrine, and biochemical foundation of motor control.
Applications will be accepted June 1, October 1, and February 1. Application kits are available at most business and grants/contracts offices and from the Office of Grants Inquiries, Division of Research Grants, National Institutes of Health, Westwood Building, Room 449, Bethesda, MD 20892, telephone (301) 496-7441. For fiscal and administrative matters contact: Kathleen Howe, Grants Management Specialist, NINDS, Federal Building, Room 1004, 7550 Wisconsin Ave., Bethesda, MD 20892 [301-496-9231].
* * *
International Directory: Progress Report
The Wisconsin Regional Primate Research Center is compiling a directory of organizations associated with the field of primatology. This International Directory of Primatology will be broadly distributed to libraries and other pertinent organizations. It includes primatological programs in the U.S.A. and in other countries. The expected completion date is early June. To date, over 150 organizations have returned questionnaires.
Any organization which has not received a questionnaire to complete for this directory, should contact Larry Jacobsen, WRPRC Library, 1220 Capitol Court, Madison, WI 53715 or (608) 263-3512 or [email protected].
Potto Studbook
Cheryl Frederick, keeper at the Franklin Park Zoo, is trying to establish a studbook on Peridicticus potto. She is also trying to coordinate husbandry information on either the potto or the golden potto, Arctocebus calabarensis. If you are keeping or have kept either of these species, please contact Cheryl: Franklin Park Zoo, c/o African Tropical Forest, Boston, MA 02121 [617-442-4872; FAX: 617-442-2815].
Self-injurious Behavior in Callitrichids
As part of a graduate research project, Tami Guy is attempting to gather information concerning self-injurious behavior (SIB) of marmosets and tamarins. Although any information about all types of SIB would be appreciated, she is especially interested in self-inflicted wound aggravation. If your institution houses callitrichids, please complete and return this survey to Tami Guy, Psychology Department, Bucknell University, Lewisburg, PA 17837 [717-524-3021 or 717-524-1200].
*1. Do you presently or have you ever housed any callitrichids that
have exhibited self-injurious behavior? YES. NO.
*2. List the species, sex, age, and SIB exhibited.
*3. Describe the environment in which the animal is/was housed (dimen-
sions, objects in enclosure, size of group, other species).
*4. Describe how/if the situation was resolved (treatments attempted,
effective treatment, if behavior resolved itself).
*5. Please contribute any other pertinent information.
Primate Surgical Anatomy
Mike Worrell, Indiana University School of Medicine, is conducting dissections on primate carcasses to demonstrate and document areas of surgical, veterinary, and laboratory interest. In the summer of 1990, he began a series of dissections to investigate the anatomy of the great apes (gorillas, orangs, and chimps). He quickly found that the anatomy as documented in the literature is often incomplete and/or inaccurate (Gorilla Gazette, 1991, 5[1], 9). He hopes to publish more accurate and helpful documentation of great ape morphology, and specifically to demonstrate anatomy important to veterinarians and keepers who care for these animals. He is asking for suggestions of areas of interest, which might include surgical approaches, injection sites, etc.
Current work concentrates on surgical anatomy in great apes, but he hopes to include gibbons and langurs soon, and would like to continue to expand the project with other species. Carcasses of most nonhuman primates are also needed for class dissections and research. If you have occasion to dispose of carcasses, please consider this project. Two institutions have already entrusted him with the remains of their deceased apes. To make this project a significant improvement over those of the past, more specimens are required, with a variety of animal ages. While whole, intact carcasses are most desirable, biopsied or even necropsied animals are useful.
Suggestions for species or regions of anatomical interest, and notification of availability of carcasses can be made by contacting Dr. Worrell, Educational Resources Coordinator, Dept. of Pathology, Indiana University School of Medicine, Medical Sciences Bldg., Room 157, 635 Barnhill Drive, Indianapolis, IN 46202-5120 [317-274-1598; FAX: 317-274-3223].
AAALAC Animal Well-Being Poster
The American Association for Accreditation of Laboratory Animal Care (AAALAC) has produced a poster promoting laboratory animal well-being as essential to life sciences research, teaching, and testing. One copy of the 18 x 22 inch poster is free, while additional copies cost $5 each. They can be ordered from AAALAC, 11300 Rockville Pike, Suite 1211, Rockville, MD 20852-3035 [301-231-5353].
A.P.A. E-mail List
The APA Science Directorate has developed an electronic mailing list, called APA Scientific Psychology Action Network (APA-SPAN), intended to be a partnership between the Directorate and scientific/academician psychologists. Each month, members of APA-SPAN receive a mailing via BITNET, the electronic mail system. Mailings include information on activities and opportunities such as legislative developments, science conference programs, and student funding programs and awards. The Science Directorate also assists SPAN members in conducting local/district legislative and political activities.
When constituent action is called for on a particular legislative issue, or when the information about an important event needs to be communicated, the Science Directorate contacts SPAN members with instructions and background information. To get involved in current policy issues and become a member of the APA-SPAN, contact Stephanie Holaday via BITNET at APASDSDH@GWUVM.
* * *
NABR Fall Conference
The National Association for Biomedical Research has tentatively scheduled its fall conference, focussing on education issues, for October 4-6, in Washington DC. For information, contact NABR, 818 Connecticut Ave. N. W., Suite 303, Washington, DC 20006.
SCAW Conference
The Scientists' Center for Animal Welfare (SCAW) will sponsor a conference on "Refinement Strategies in Animal Testing," September 17-18, 1992, in the Trenton/New Brunswick/Philadelphia area. The purpose of this conference is to review advances in refinement and reduction of animal use in the testing of medical, industrial, and consumer products. For more information, contact: Conferences, SCAW, 4805 St. Elmo Ave., Bethesda, MD 20814 [301-654-6390; FAX: 301-907-3993].
Symposium on Models for AIDS
The Caribbean Primate Research Center and the University of Puerto Rico Medical Sciences Campus will be hosting the Tenth Annual Symposium on Nonhuman Primate Models for AIDS, from 17-20 November, 1992, at the Condado Plaza Hotel and Casino, in San Juan, PR. Oral presentations and posters will focus on virology and molecular biology, pathogenesis, immunology, prophylaxis and therapy, opportunistic infections and diseases, epizootiology and transmission studies, and human studies relevant to simian AIDS models. Selected papers presented at the symposium will be published in the Journal of Medical Primatology as a special issue. For additional information, contact Matt J. Kessler, Caribbean Primate Research Center, Univ. of Puerto Rico, Medical Sciences Campus, P.O. Box 1053, Sabana Seca, PR 00952-1053 [809-784-0322; FAX: 809-795-6700].
NY Regional Group Meetings
The New York Regional Primatology Group will hold meetings at 7:30 pm on Thursday, April 23 (speaker: Dr. Carey Yeager of the Louis Calder Center, Fordham University, on "The Proboscis monkey of Borneo: Conservation & ecology") and on Thursday May 7 (speaker: Dr. Mark Weiss of NSF and Wayne State University, on "DNA Fingerprinting in Primates"). All meetings are held at the CUNY Graduate Center, 33 W. 42 Street, eleventh floor, room 1100, New York. The cafeteria on the 18th floor is open for dinner prior to the seminar. Parking becomes available on 42nd Street at 7:00 p.m. For further information contact Warren Kinzey [212-650-7363; bitnet: anthwk@ccnyvme]. -- From the electronic bulletin board, PRIMATE-TALK.
* * *
Pennsylvania
*University of Pittsburgh, Department of Anthropology
PROGRAM NAME: Biological Anthropology Graduate Program
FACULTY AND THEIR SPECIALTIES: Steven J. C. Gaulin (evolution of
behavioral adaptations, particularly those that differ between the
sexes; use of evolutionary theory, behavioral ecology, and comparative psychology to model the evolution of human behavior); Mark P.
Mooney (craniofacial and development biology, comparative anatomy,
experimental morphology, physiological adaptations to extreme environments, development of animal models for facial clefts); Jeffrey H.
Schwartz (method, theory, and philosophy in evolutionary biology;
origin and diversification of primates; human and faunal skeletal
analysis; dentofacial growth and development); Michael I. Siegel
(craniofacial biology, with a clinical speciality in cleft palate;
functional anatomy; animal models; physiological adaptation).
FOR FURTHER INFORMATION: Nancy J. Stugan, Graduate Admissions Coordinator, Department of Anthropology, University of Pittsburgh, Pittsburgh, PA 15260.
* * *
Wildlife Preservation Trust
The Wildlife Preservation Trust develops and maintains captive breeding programs worldwide, as well as conducting research, educational programs, and professional training. Their Training Program is a 10 or 16 week internship designed for postgraduates and zoo or animal center personnel to give intensive training in propagation techniques with a variety of species. For application and further information, contact Training Program, Wildlife Preservation Trust International, 34th St. & Girard Ave., Philadelphia, PA 19104 [215-222-3636; FAX: 215-222-2191]. Deadline for application for 1993 positions is July 1, 1992. The Summer Course is designed for advanced undergraduates and graduate students. For details on the 1993 course, write to John McNicholas, Assistant Training Officer, Summer School Coordinator, Jersey Wildlife Preservation Trust, Les Augres Manor, Trinity, Jersey, Channel Islands, GB.
ASP Summer Internship List
The Education Committee of the American Society of Primatologists (ASP) has again published a listing of opportunities for students in primate research and husbandry, in the March 1992 issue of the ASP Bulletin. For copies of this listing, or to add information to it, contact the Chair of the Committee, Dr. Chris Duggleby, Dept. of Anthropology, 380 MRAC, SUNY, Buffalo, NY 14221 [716-636-2356; FAX: 716-636-3808].
Fogarty/Japan Postdoctoral Program
The Japan Society for Promotion of Science and the Japan Science and Technology Agency, in collaboration with the Fogarty International Center, are sponsoring two fellowship programs for American biomedical and behavioral scientists to conduct postdoctoral studies in Japan. American scientists with a doctorate in clinical, behavioral or biomedical science who are under 35 years old are eligible to apply. The programs provide salary, family, housing and relocation allowances, round-trip air-fare and travel allowance from six months to two years. Applications must be received by May 10. For applications and further information, contact: the International Research and Awards Branch, Fogarty International Center, NIH, Bethesda, MD 20892 [301-496-1653]. Summer Pathology Program
The Registry of Comparative Pathology, located at the Armed Forces Institute of Pathology (AFIP), announces a summer student program in comparative pathobiology from about June 1 to August 15. This externship offers research experience in a stimulating environment, as well as the opportunity to participate in various educational programs at the AFIP. The student, closely supervised by a senior pathologist, will be integrated into a pathology project applying new technologies such as quantitative morphometry, molecular biology, etc. Applicants must have six years of higher education, be familiar with basic laboratory techniques, and have completed core pathology coursework in a veterinary or medical curriculum. A stipend of $419 per week is provided. For additional information and applications, contact Dr. Linda Johnson, Registry of Comparative Pathology, AFIP, Washington, DC 20306-6000 [202-576-2452].
Volunteer for Field Study
A responsible person with common sense, a safe driving record, and course work in animal behavior or primate behavioral ecology is sought to assist in developing a field study of vervets and patas monkeys in Kenya, beginning August, 1992. Field experience would be a great asset; graduate credit is a possibility. Room and board under spartan conditions will be provided. Please send a cover letter detailing your reasons for applying, and a resume with addresses and phone numbers of three references to Dr. Lynne A. Isbell, Dept. of Anthropology, Univ. of California, Davis, CA 95616. -- From the ASP Bulletin, 1992, 16[1].
School for Field Studies
The School for Field Studies offers 13-week semester programs and month-long summer courses in ecology and natural resource management. Over 200 colleges and universities accept credit for these courses. Courses include Wildlife Ecology and Management (in Kenya), Studies in Sustainable Development (Costa Rica), and Ecology and Conservation of the Golden-headed Lion Tamarin (Brazil). For information, contact the School for Field Studies, 16 Broadway, Beverly, MA 01915 [508-927-7777].
* * *
He got it from the movies, of course: discovered
Virgil the chimpanzee in Project X, and
--because the creature acted so damned human,
like an inappropriately hairy child
still wearing a diaper--assumed his character:
began to gibber, made uncanny hooting sounds,
played grotesque games with citrus fruit, and loped
around the house, arms dangling, until we said
Okay, Ben! time! enough!
And he would tilt his head
and look at us, purse his lips, and bare-teethed hiss and spray
one last time before--as though Virgil were a
Darwinian Pinocchio becoming
a real boy--he'd go to his room to blow
Purcell fanfares for his next day's trumpet lesson.
What I think was fearsome was how close he came:
for I suspected not that he was a boy
with an overactive imagination,
aping an ape, but that Nature had reversed
the impersonation, that it was dropping
the wall chart to teach us Evolution as
the Grand Masque of Deception, reminding us
how close Ben, child of our loins, carrier of our genes,
was to the being he tributed in mockery:
that Henry Purcell wrote trumpet voluntaries
to keep from rubbing his crotch with a banana,
not because he had forgotten how; and that Ben
blows them through a hundred feet of copper tubing
because he has seen Nature as a third parent
that he has learned, necessarily, to deceive.
Kenneth Wolman, December 15, 1991
* * *
Books
* Primate Behavior: An Exercise Workbook. J. D. Paterson. Prospect
Heights, IL: Waveland Press, 1992. 105 pp. [Price: $9.95]
. .
Detailed descriptions, instructions, and data sheets for students
to perform eight observation projects, plus chapters on data analysis
and computer aided data collection. The projects are described in
terms of primate species living in zoos, but could be carried out with
other species in other settings.
* Through the Looking Glass: Issues of Psychological Well-being in Captive Nonhuman Primates. M. A. Novak & A. J. Petto (Eds.). Washington,
DC: American Psychological Association, 1991. 285 + xiii pp. [Price:
$40; $32 to A.P.A. members and affiliates]
. .
Papers presented at a conference held in 1988 at Harvard Medical
School. The subjects addressed are Historical perspectives on psychological well-being legislation; Defining psychological well-being;
Potential parameters of psychological well-being; Implications of the
new amendments from an administrative and a veterinary perspective;
Procedures for promoting psychological well-being in nonhuman primates; and Public reaction to primate research. There is also a
52-page bibliography, Welfare of Nonhuman Primates in Research Settings, with over 600 entries.
Magazines, Newsletters, and Reports
* 1991 Highlights. National Association for Biomedical Research. (NABR, 818 Connecticut Ave., N.W., Suite 303, Washington, DC 20006).
* Humane Innovations and Alternatives, 1990, Volume 5. (Published by
PsyETA, Box 1297, Washington Grove, MD 20880-1297). [Subscriptions:
$20 for non-PsyETA members].
. .
Articles include: Primate enrichment, by H. Shaw; Enrichment toys
and tools in recent trials, by P. Hamilton; Captivity as torture? by
D. K. Candland; Primate preference for outdoors, by M. E. Pereira;
and articles on the ethics of animal experimentation.
* Primate Conservation: The Newsletter and Journal of the IUCN/SSC Primate Specialist Group, Number 9, December 1988. [Subscription: $15,
payable to Primate Conservation. Order from Conservation International, 1015 18th St., NW, Suite 1000, Washington, DC 20036]
. .
Announcements and articles from September 1987 to December 1988.
Issue Number 10, with material to December 1989, is to be published
soon.
Special Journal Issues
* Monkey behavior and laboratory issues. Laboratory Animal Science, 1991, 41[4].
. .
Articles, based on a January, 1990, NIH-sponsored workshop, begin
with an evaluation of the premise of "need" for an enriched environment. Subsequent articles discuss methods adaptable to the research
setting which promote behavioral well-being. Guest editors were K.
Bayne and M. Novak.
Anatomy
* Comparative anatomy of the tracheobronchial circulation. Magno, M.
(Dept. of Surgery, Thomas Jefferson Univ., 605 College Bldg., 1025
Walnut St., Philadelphia, PA 19107). European Respiratory Journal,
1990, 3, 447s-563s.
. .
The monkey, dog, and cat lung appear more similar to each other
than to the human lung.
Animal Models
* Visual cortex development in rhesus monkeys deprived of dietary taurine. Neuringer, M., Palackal, T., Kujawa, M., Moretz, R. C., & Sturman, J. A. (Oregon RPRC, Beaverton, OR 97006). In Taurine: Functional
Neurochemistry, Physiology, and Cardiology (pp. 415-422). New York:
Wiley-Liss, 1990.
. .
The morphology of the primary visual cortex (area 17) in rhesus
monkeys was significantly affected by dietary taurine deprivation from
birth to three months of age. The monkeys also showed changes in cortical pattern evoked potentials, which suggest an abnormality in the
processing of spatial information.
* In utero bone marrow transplantation of fetal baboons with mismatched
adult baboon marrow. Roodman, G. D., Kuehl, T. J., Vandeberg, J. L., &
Muirhead, D. Y. (Research Service (151), V.A. Hospital, 7400 Merton
Minter Blvd., San Antonio, TX 78284). Blood Cells, 1991, 17, 367-375.
. .
Thirty-one fetuses received marrow. Of the six which demonstrated
engraftment, three died, and three rejected the grafts before birth.
* Use of serum lipid and apolipoprotein concentrations to predict
extent of diet-induced atherosclerotic lesions in aortas and coronary
arteries and to demonstrate regression of lesions in individual rhesus
monkeys. Eggen, D. A., Bhattacharyya, A. K., Strong, J. P., Newman, W.
P. III, Guzman, M. A., & Restropo, C. (A. K. B., Dept. of Pathology,
LSU Medical Center, 1901 Perdido St., New Orleans, LA 70112). Arteriosclerosis and Thrombosis, 1991, 11, 467-475.
. .
A statistical method is developed for assessing regression of
experimentally induced atherosclerotic lesions in individual animals,
which might be a useful alternative to the usual comparison by group
and an additional tool for studies of regression of experimental atherosclerosis.
* The behavioral neurobiology of self-injurious behavior in rhesus monkeys. Kraemer, G. W. & Clarke, A. S. (Harlow Primate Lab., 22 N.
Charter St., Madison, WI 53715). Progress in Neuro-Psychopharmacology
& Biological Psychiatry, 1990, 14, S141-S168.
. .
A review of the causes and behavioral and physiological effects of
self-injurious behavior (SIB). In rhesus monkeys, SIB probably
reflects the operation of a system that promotes and maintains the
mechanisms necessary for dominance and sexual behavior in the absence
of social factors that would normally bring these behaviors under control.
* Dose-response effects of oral yohimbine in unrestrained primates.
Rosenblum, L. A., Coplan, J. D., Friedman, S., & Bassoff, T. (Box 120,
SUNY Health Science Center, 450 Clarkson Ave., Brooklyn, NY 11203).
Biological Psychiatry, 1991, 29, 647-657.
. .
Yohimbine increased episodes of motoric activation and affective
response interspersed with intervals of behavioral enervation, but
there was no dose-response relationship. The cyclic enervative episodes suggest limitations to primate models of panic disorder utilizing oral yohimbine.
* Retinal toxicity of intravitreal gentamicin. Brown, G. C., Eagle, R.
C., Shakin, E. P., Gruber, M., & Arbizio, V. V. (910 E. Willow Grove
Ave., Wyndmoor, PA 19118). Archives of Ophthalmology, 1990, 108,
1740-1744.
. .
A gentamicin dose equivalent to that routinely administered for
subconjunctival injections by many opthalmic surgeons after ocular
surgery caused generalized vascular incompetence, diffuse retinal
necrosis, thrombosis of the large retinal blood vessels, widespread
loss of the retinal capillary pericytes and endothelial cells, and a
rapid extinction of the electroretinogram, when injected intravitreally in pigtail macaques.
* Endogenous bilirubin excretion in Bolivian squirrel monkeys with a
Gilbert's-like syndrome. Myers, B. A., Bruss, M. L., George, J. W., &
Cornelius, C. E. (Hazelton Labs, Rockville, MD). Journal of Medical
Primatology, 1991, 20, 97-103.
. .
Fasting Bolivian squirrel monkeys, a model for humans with Gilbert's syndrome, show high bilirubin excretion, which may be due to a variety of factors, which are discussed.
Animal Welfare
* Enrichment devices for nonhuman primates. Schapiro, S. J., Brent, L.,
Bloomsmith, M. A., & Satterfield, W. C. (M. D. Anderson Cancer Center,
Science Park, Dept. of Veterinary Resources, Route 2, Box 151-B1, Bastrop, TX 78602). Lab Animal, 1991, 20, 22-28.
. .
Descriptions and discussion of different types of enrichment
devices, with suggestions for acquisition and maintenance.
* Customizing an enrichment program: Rhesus monkeys. O'Neill, P. L. &
Price, C. S. (NIH Animal Center, 22765 Club Hollow Rd., Dickerson, MD
20842). Lab Animal, 1991, 20, 29-42.
. .
Normative activity schedules for four age groups and both genders
of rhesus monkeys in two distinct settings: a free-ranging outdoor
enclosure, and a confined indoor housing unit.
Behavior
* Rank distance as a central feature of rhesus monkey social organization: A sociometric analysis. De Waal, F. B. M. (Dept. of Psychology,
Emory Univ., Atlanta, GA 30322). Animal Behaviour, 1991, 41, 383-395.
. .
Data from 3 studies on two captive groups confirm that rank distance correlates with a range of social behavior independently of the effects of kinship.
* Infant carrying in a polygynous group of common marmosets (Callithrix jacchus). Koenig, A. & Rothe, H. (Inst. of Anthropology, Univ.
of Gottingen, Burgerstr. 50, 3400 Gottingen, Germany). American Journal of Primatology, 1991, 25, 185-190.
. .
In a captive group of six animals plus the three infants of two
breeding females, carrying and suckling were analyzed. During the
first two weeks of the infants' lives, the breeding females carried
their own infant(s) almost exclusively. At the beginning of the third
week, nearly all group members began to carry all infants, and both
mothers suckled all infants, indicating that the greater energetic
demands in rearing three infants simultaneously are compensated by an
extended division of labor among group members.
* Mirror-induced social facilitation in stumptailed macaques (Macaca
arctoides). Straumann, C. & Anderson, J. R. (J.R.A., Lab. de Psychophysiologie, Univ. Louis Pasteur, 7 rue de l'Universite, 67000 Strasbourg, France). American Journal of Primatology, 1991, 25, 125-132.
. .
Possible explanations for the ability of a mirror to induce social
facilitation of drinking but not of object manipulation are discussed,
along with possible underlying mechanisms.
* Object manipulation in hand-reared and mother-reared infant chimpanzees. Mattison, A., Howell, S. M., & Fritz, J. (Primate Foundation of
Arizona, P.O. Box 86, Tempe, AZ 85280). The Newsletter, 1991,
3[2], 4-5.
. .
135 hours of observation of four hand-reared and five mother-reared
infants, ranging in age from 13 to 39 months at study onset, showed
little difference in amount of time playing, motor coordination, and
manipulative abilities. However, the hand-reared infants prefered to
handle non-food objects, while the others preferred food objects.
* Dominance and feeding competition in captive rhesus monkeys.
Deutsch, J. C. & Lee, P. C. (Dept. of Biological Anthropology, Downing
St., Cambridge CB2 3DZ, England). International Journal of Primatology, 1991, 12, 615-628.
. .
The feeding behavior of 16 adult female rhesus monkeys living in 3
captive social groups was observed. Higher-ranking females had greater
access to feeding sites, but no consistent differences in estimates of
total intake were found between females of high rank and those of low
rank. The effect of dominance on reproductive performance (higher-ranking females had more surviving offspring) appeared to be less
related to food intake than to competitive and aggressive interactions, potentially resulting in higher levels of stress for subordinates.
* Social behavior of the emperor tamarin in captivity: Components of
agonistic display and the agonistic network. Knox, K. L. & Sade, D. S.
(Dept. of Anthropology, Northwestern Univ., Evanston, IL 60208).
International Journal of Primatology, 1991, 12, 439-480.
. .
Agonistic behavior was studied longitudinally for 16 months in an
intact family group, using methods from quantitative ethology and
social network analysis. The frequency with which individual tamarins
long called and scent marked was not closely correlated with their
dominance rank, but individuals undergoing change in status long
called and scent marked most frequently, regardless of rank.
* The reconciliation behavior of golden monkeys (Rhinopithecus roxellanae roxellanae) in small breeding groups. Ren, R., Yan, K., Su,
Y., Qi, H., Liang, B., Bao, W., & de Waal, F. B. M. (Dept. of Psychology, Beijing Univ., Beijing, People's Republic of China). Primates, 1991, 32, 321-327.
. .
Observations following 130 spontaneous aggressive incidents in two
groups showed the dominance hierarchy is "indulgent" for the dominant
and "relaxed" for the subordinant. Adult males mediated in agonistic
disputes among females.
Care
* Normalizing laboratory-reared rhesus macaque (Macaca mulatta) behavior with exposure to complex outdoor enclosures. O'Neill, P. L.,
Novak, M. A., & Suomi, S. J. (NIH Animal Center, 22765 Club Hollow
Rd., Dickerson, MD 20842). Zoo Biology, 191, 10, 237-245.
. .
Laboratory reared animals showed more locomotion and exploration
after 9 weeks in an outdoor enclosure than similar animals kept
indoors. The experimental animals also showed a decline in self-oral
behavior which persisted even upon return to the laboratory, suggesting that juvenile monkeys can benefit from even brief exposure to enriched outdoor settings.
* Rearing infant monkeys (Macaca nemestrina) in pairs produces deficient social development compared with rearing in single cages. Ruppenthal, G. C., Walker, C. G., & Sackett, G. P. (G.P.S., Primate Center SJ-50, Univ. of Washington, Seattle, WA 98195). American Journal
of Primatology, 1991, 25, 103-113.
. .
Infants paired from postnatal week 3 through month 4, especially
females, developed a playroom behavioral repetoire consisting largely
of mutual clinging, fear, and social withdrawal.. Unlike singly caged
infants, pair-reared monkeys did not successfully adapt to living in a
large social group at 8-10 months of age.
* Use of a grooming and foraging substrate to reduce cage stereotypies
in macaques. Lam, K., Rupniak, N. M. J., & Iversen, S. D. (N.M.J.R.,
Merck Sharp & Dohme Research Labs., Neuroscience Research Centre, Terlings Park, Eastwick Rd., Harlow, Essex, CM20 2QR, UK). Journal of
Medical Primatology, 1991, 20, 104-109.
. .
Stereotyped behaviors were reduced by up to 73% by use of a fleece
pad, either alone or sprinkled with morsels of food.
* Mother-infant cannibalism in thick-tailed bushbabies (Galago crassicaudatus umbrosus). Tartabini, A. (Dip. Scienze dell'Educazione, Univ. di Calabria, 87030 Arcavacata di Rende, Cosenza, Italy). Primates, 1991, 32, 379-383.
. .
Three cases of cannibalism, not associated with starvation or
infanticide, seemed to be due to stressful laboratory conditions.
* Hand-rearing and reintroduction of a golden-crowned sifaka, Propithecus tattersalli, at the Duke University Primate Center. Coffman, B.
S. (Duke Univ. Primate Center, 3705 Erwin Rd., Durham, NC 27705).
International Zoo Yearbook, 1990, 29, 137-143.
. .
Detailed description of a success story.
Conservation
* Financial considerations of reserve design in countries with high
primate diversity. Ayres, J. M., Bodmer, R. E., Mittermeier, R. A. (R.
E. B., Dept. de Zoologia, Museu Paraense E. Goeldi, Caixa Postal 399,
66.040, Belem, Para, Brazil). Conservation Biology, 1991, 5, 109-114.
. .
Large protected areas have substantially less perimeter to patrol
than many small reserves, minimizing maintenance costs, but some key
primate-containing countries, such as Madagascar, Indonesia, and S.E.
Brazil, require many small reserves to insure preservation of endemic
taxa.
Development
* Behavioural changes at adolescence in chimpanzees. Pusey, A. E.
(Dept. of Ecology, Evolution, and Behavior, Univ. of Minnesota, Minneapolis, MN 55455). Behaviour, 1990, 115, 203-246.
. .
A report on a partly cross-sectional and partly longitudinal study
of the free-living but habituated juvenile and adolescent chimpanzees
at Gombe National Park, Tanzania.
* Deciduous dentition eruption sequence of the laboratory-reared chimpanzee (Pan troglodytes). Mooney, M. P., Siegel, M. I., Eichberg, J.
W., Lee, D. R., & Swan, J. (Dept. of Anthropology, Univ. of Pittsburgh, Pittsburgh, PA 15260). Journal of Medical Primatology, 1991, 20, 138-139.
. .
Eruption sequence and age ranges for the maxillary and mandibular
deciduous dentition were generated from serial dental study models
from 28 laboratory-reared chimpanzees.
* Postnatal change in lipid composition and nerve conduction of peripheral nerves of young rhesus monkeys. Rana, S. V., Mehta, S., & Chopra,
J. S. (Dept. of Gastroenterology, Postgrad. Inst. of Medical Education
and Research, Chandigarh, India). Journal of Medical Primatology,
1991, 20, 318-322.
. .
Measurements on 15 subjects, over a 5 month period, indicate that
myelination continues for at least 11 to 13 months after birth.
Disease
* Biology of B virus in macaque and human hosts: A review. Weigler, B.
J. (Dept. of Companion Animal & Special Species Medicine, College of
Veterinary Medicine, North Carolina State Univ., 4700 Hillsborough
St., Raleigh, NC 27606). Clinical Infectious Diseases, 1992, 14,
555-567.
. .
B virus is a zoonotic alphaherpesvirus enzootic in Asian monkeys of
the genus Macaca. At least 25 cases of human disease caused by B
virus have occurred to date, leading to death in 16 instances. Advances
in the technology available for the diagnosis of B virus infection and
in the agents for its treatment are improving the prognosis for cases
in human beings. Efforts are underway at several institutions in the
U.S. to establish B virus-free colonies of rhesus macaques for use in
biomedical research. Unfortunately, the epidemiology of B virus in
group-housed macaques is poorly understood. The elucidation of factors
important in the transmission of B virus between monkeys will greatly
enhance efforts to eradicate this virus and may help to minimize further human exposure to the agent.
* A coprological survey of parasites of wild mantled howling monkeys,
Alouatta palliata palliata. Stuart, M. D., Greenspan, L. L., Glander,
K. E., & Clarke, M. R. (Dept. of Biology, Univ. of North Carolina,
Asheville, NC 28804). Journal of Wildlife Diseases, 1990, 26,
547-549.
. .
Fecal samples from 155 animals revealed that 48% had parasitic
infections: Controchis biliophilus, Trypanoxyuris minutus, unidentified strongylid eggs, and Isospora sp. oocysts. Three monkeys died,
and were discovered to also be infected with Ascaris lumbricoides.
* Species-specific diversity among simian immunodeficiency viruses from
African green monkeys. Allan, J. S., Short, M., Taylor, M. E., Su, S.,
Hirsch, V. M., Johnson, P. R., Shaw, G. M., & Hahn, B. H. (Dept. of
Virology & Immunology, Southwest Foundation for Biomedical Research,
7620 NW Loop 410, San Antonio, TX 78228-0147). Journal of Virology,
1991, 65, 2816-2828.
. .
West African green monkeys are naturally infected with SIVs which
are closely related to East African SIVagm isolates. However, structural, antigenic, and genetic differences strongly suggest that the
West African green monkey viruses comprise a phylogenetically distinct
subgroup of SIVagm. SIVagm viruses may have evolved and diverged coincident with the evolution and divergence of their African green monkey host.
* Evaluation of a chimpanzee colony for antibodies to hepatitis C
virus. Lanford, R. E., Notvall, L., Barbosa, L. H., & Eichberg, J. W.
(Address same as above). Journal of Medical Virology, 1991, 34,
148-153.
. .
The recently developed enzyme immunoassay (EIA) for antibodies to
hepatitis C virus (HCV) was applied to chimpanzees of known history.
Only 31.3% of the animals that had been inoculated with a non-A, non-B
hepatitis agent were currently positive, and a retrospective analysis
suggested that an additional 20% of the animals had been positive at
some time following infection. Due to the low seroconversion rate and
the uncertainties surrounding many positive reactions in uninoculated
animals, this assay cannot be used to determine the initial source or
extent of spread of HCV infections in uninoculated animals.
* Gastric stromal tumors in two rhesus macaques (Macaca mulatta). Banerjee, M., Lowenstine, L. J., & Munn, R. J. (Animal Health Diagnostic Lab., Michigan State Univ., P.O. Box 30076, Lansing, MI 48909). Veterinary Pathology, 1991, 28, 30-36.
. .
Two fatal tumors in two animals histologically resembled smooth
muscle tumors but were of neuroectodermal and primitive mesenchymal
origin.
* Experimental infection of pig-tailed macaques (Macaca nemestrina)
with Campylobacter cinaedi and Campylobacter fennelliae. Flores, B.
M., Fennell, C. L., Kuller, L., Bronsdon, M. A., Morton, W. R., &
Stamm, W. E. (W.E.S., Dept. of Medicine, Div. of Infectious Diseases,
Harborview Med. Center, Seattle, WA 98104). Infection and Immunity,
1990, 58, 3947-3953.
. .
C. cinaedi and C. fennelliae consistently produce a diarrheal
illness accompanied by bacteremia and followed by prolonged gastrointestinal colonization in M. nemestrina.
* Antiviral effects of 3'-fluorothymidine and 3'-azidothymidine in
cynomolgus monkeys infected with simian immunodeficiency virus. Lundgren, B., Bottinger, D., Ljungdahl-Stahle, E., Norrby, E., Stahle, L.,
Wahren, B., & Oberg, B. (B.O., Dept. of Virology, Karolinska Inst.,
SBL, S-105 21 Stockholm, Sweden). Journal of Acquired Immune Deficiency Syndromes, 1991, 4, 489-498.
. .
Neither compound tested prevented infection despite dosing prior to
virus inoculation. FLT was about 10 times more potent than ZDV in
delaying the appearance of SIVsm antigen in monkeys. The serum half-life of FLT was longer than that of ZDV. ZDV was bound to plasma proteins to about 60% while FLT was virtually unbound.
* Lymphocyte and neutrophil dysfunction associated with hepatitis B
virus and hepatitis non-A, non-B virus infection in the chimpanzee.
Lafrado, L. J., Javadian, M. A., Marr, J. M., Wright, K. A., Kelliher,
J. C., Dezzutti, C. S., Cummins, L., & Olsen, R. G. (Ohio State Univ.,
Center for Retrovirus Research, 1925 Coffey Rd., Columbus, OH 43210).
Journal of Medical Primatology, 1991, 20, 302-307.
. .
Mitogen-induced lymphocyte proliferation was lower in virus-infected chimps than in uninfected animals. Neutrophils from infected
animals exhibited decreased or altered chemiluminescence kinetics.
* Gastro-intestinal helminths in a natural population of Macaca sinica
and Presbytis spp. at Polonnaruwa, Sri Lanka. Dewit, I., Dittus, W.
P. J., Vercruysse, J., Harris, E. A., & Gibson, D. I. (Inst. of Fundamental Studies, Hantana Rd., Kandy, Sri Lanka). Primates, 1991, 32, 391-395.
. .
Nematodes and one cestode, found in eight fatally wounded or
recently deceased monkeys, and in fresh fecal samples.
Facilities
* Battelle Primate Facility. Weller, R. E., Wierman, E. L., Malaga, C.
A., Baer, J. F., & LeMieux, T. P. (Developmental Toxicology Section,
Battelle, Pacific Northwest Labs, Richland, WA 99352). Journal of
Medical Primatology, 1991, 20, 133-137.
. .
Description of the facility, which houses one of the largest collections of neotropical primates in the U.S., and its procedures and programs.
General
* A curious kinship: Apes and humans. Linden, E. Photos by M. Nichols.
National Geographic, 1992, 181[3], 2-45.
* Bonobos, chimpanzees with a difference. Linden, E. Photos by F.
Lanting. National Geographic, 1992, 181[3], 46-53.
. .
Photo-essays on the great apes.
Genetics
* Genetic polymorphism of the vitamin D-binding protein (DBP) in crab-eating macaques (Macaca fascicularis). Tanaka,H., Kawamoto, Y., &
Terao, K. (Lab. of Animal Genetics, Fac. of Agriculture, Nagoya
Univ., Nagoya, Aichi 464-01, Japan). Journal of Medical Primatology,
1991, 20, 126-132.
. .
Plasma samples were examined from 529 animals imported from Malaysia, Indonesia, and the Philippines. A total of 17 phenotypes governed
by at least 11 alleles were observed. Genetic variability was high in
Malaysian and Indonesian populations, but low in the Philippine population.
Instruments & Techniques
* The use of nonmetal electrodes in electroejaculation of restrained
but unanesthetized macaques. Sarason, R. L., VandeVoort, C. A., Mader,
D. R., & Overstreet, J. W. (C.A.V., California PRC, Univ. of California, Davis, CA 95617-8542). Journal of Medical Primatology, 1991,
20, 122-125.
. .
A technique for electroejaculation with nonmetallic electrodes cut
from defibrillation pads was used on cynomolgus and rhesus macaques.
119 semen specimens were obtained in 123 attempts.
Physiology
* The clearance of immune complexes from the circulation of man and
other primates. Hebert, L. A. (Dept. of Medicine, Ohio State Univ.,
Columbus, OH 43210). American Journal of Kidney Diseases, 1991, 17,
352-361.
. .
In mammals the liver and spleen are the major sites of immune complex (IC) uptake. Primates differ from nonprimates in that erythrocytes are important in clearing IC from their circulation. Primate
erythrocytes express complement receptor type 1 (CR1), which permits
IC opsonized with C3b to bind to the circulating erythrocytes. Once
bound, the IC appear to be prevented from depositing in vulnerable
organs such as the kidney. Instead the IC remain bound to the erythrocyte until the erythrocyte traverses liver or spleen, where the IC is
quickly stripped from the erythrocyte and taken up by the mononuclean
phagocyte system of liver or spleen. The erythrocyte then returns to
the circulation apparently able, once again, to participate in this
clearing mechanism.
* Development of a circadian variation of plasma oxytocin concentration
in the late gestation rhesus monkey. Seron-Ferre, M., Taylor, N. F.,
Martin, M. C., & Leake, R. D. (R.D.L., Harbor/UCLA Medical Center,
1124 W. Carson St., RB-1, Torrance, CA 90502-2064). Journal of Clinical Endocrinology and Metabolism, 1991, 72, 1323-1327.
. .
A clear circadian pattern of oxytocin (OT) was detected about 8
days before delivery in 4 pregnant monkeys, suggesting that there is a
relationship between the pattern of OT secretion and advancing pregnancy.
* Taste and olfactory modulation of feeding related neurons in behaving
monkey. Oomura, Y., Nishino, H., Karadi, Z., Aou, S., & Scott, T. R.
(Dept. Biological Control Systems, Nat. Inst. Physiological Sciences,
38, Myoudaji, Okazaki, 444, Japan). Physiology & Behavior, 1991,
49, 943-950.
. .
Glucose-sensitive (GS) neurons responded predominantly to two or
more taste and odor stimuli, while glucose-insensitive (GIS) neurons
responded to only one stimulant. Apparently GS neurons process endogenous chemical information and integrated chemical sensations, while
GIS neurons process external information, motor control, and discriminative chemical sensations.
* Effects of growth hormone on neonatal growth in nursing rhesus monkeys. Wilson, M. E., Gordon, T. P., Chikazawa, K., Gust, D., Tanner,
J. M., & Rudman, C. G. (Yerkes PRC, Field Station, Emory Univ., 2409
Taylor Rd., Lawrenceville, GA 30243). Journal of Clinical Endocrinology and Metabolism, 1991, 72, 1302-1307.
. .
Neonatal body weight gain can be accelerated in nursing infants
whose mothers have received GH from at least the second trimester of
prenancy through the lactational period. Infants of mothers receiving
GH during lactation only were not different from controls.
* ACTH and glucocorticoid responses under two conditions of stress in
macaques. Clarke, A. S. (Harlow Primate Lab., Univ. of Wisconsin, 22
N. Charter St., Madison, WI 53715). American Journal of Primatology,
1991, 25, 115-124.
. .
Blood was drawn from captured and restrained (high stress), and
from trained (low stress) animals. While both ACTH and glucocorticoid
values were increased by the high stress procedure, only ACTH was
increased by the other, with glucocorticoid values decreased.
* CSF monoamine metabolite concentrations vary according to age, rearing, and sex, and are influenced by the stressor of social separation in rhesus monkeys. Higley, J. D., Suomi, S. J., & Linnoila, M. (NIH Animal Center, NICHD, P.O. Box 289, Poolesville, MD 20892). Psychopharmacology, 1991, 103, 551-556.
. .
Twenty-eight subjects, reared either in peer-only (PO) groups or in
mother-infant (MI) dyads, were studied at 6, 18, or 50 months of age.
The results reveal a number of developmental changes, sex differences,
and rearing condition effects on CSF monoamine metabolite concentrations. Serotonin was most affected by age, rearing, and sex. There
were significant age-related declines in 5-HIAA across all three ages.
PO reared subjects had higher levels of 5-HIAA and MHPG than MI reared
subjects.
* Hematology, serum chemistry values, and rectal temperatures of adult
greater galagos (Galago garnetti and G. crassicaudatus). Izard, M.
K., Heath, S. J., Hayes, Y., & Simons, E. L. (Research Triangle Inst.,
P.O. Box 12194, Research Triangle Park, NC 27709). Journal of Medical
Primatology, 1991, 20, 117-121.
. .
No species or sex differences were found for 4 hematological parameters and 15 serum chemistry parameters. Species differences or sex differences were seen in 12 other parameters, while species and sex differences were seen in chloride and erythrocyte number.
* Time-dependent effects of peer separation on lymphocyte proliferation
responses in juvenile squirrel monkeys. Friedman, E. M., Coe, C. L., &
Ershler, W. B. (Dept. of Psychology, Univ. of Wisconsin, 1202 W. Johnson St., Madison, WI 53706). Developmental Psychobiology, 1991, 24,
159-173.
. .
Immunological, hormonal, and behavioral responses of juvenile
squirrel monkeys to repeated social separations of varying length.
Following a 3-hr separation, lymphocyte responses to stimulation with
the mitogen concanavalin A (Con A) declined significantly; these
alterations were sustained after a 24-hr separation. At the end of a
7-day separation, immune responses did not differ significantly from
basal values. Plasma cortisol levels were elevated above basal levels
in all animals after the 3-hr, 24-hr, and 2-day separations, but not
after the 7-day separation.
* The effects of four types of restraint on serum alanine aminotransferase and aspartate aminotransferase in the Macaca fascicularis.
Landi, M. S., Kissinger, J. T., Campbell, S. A., Kenney, C. A., &
Jenkins, E. L. Jr. (Dept. of Lab. Animal Science, SmithKline Beecham,
King of Prussia, PA 19406-0939). Journal of the American College of
Toxicology, 1990, 9, 517-523.
. .
Animals were continuously restrained for 60 and 120 min in a
restraining box, a restraint chair, and on a restraining board, and
also hand caught for manual restraint. Significant differences between
the enzyme levels of different restraint methods were not seen during
the 60-min studies, but were present during the 120-min ones. Levels
of alanine aminotransferase for board restraint were significantly
higher than for the other methods.
* A pilot study of the social correlates of levels of urinary cortisol,
prolactin, and testosterone in wild long-tailed macaques (Macaca fascicularis). van Schaik, C. P., van Noordwijk, M. A., van Bragt, T.,
& Blankenstein, M. A. (Ethology and Socio-ecology, Utrecht Univ., P.O.
Box 80086, 3508 TB Utrecht, Netherlands). Primates, 1991, 32,
345-356.
. .
Urine samples and extensive activity data were collected from identified individuals. Urinary cortisol showed a marked decrease during
the day, testosterone was higher in adult than in immature males, and
prolactin levels were higher in lactating than in non-lactating
females. Hence, the method of assessing urinary hormones collected in
the wild is sufficiently reliable to undertake a study of the endocrine effects of social events and social status in an undisturbed social and ecological setting.
* Clustering of ipsilateral cortico-cortical projection neurons to area
7 in the rhesus monkey. Hiorns, R. W., Neal, J. W., Pearson, R. C.
A., & Powell, T. P. S. (Dept. of Statistics, South Parks Rd., Oxford
OX1 3TG, UK). Proceedings of the Royal Society of London, B, 1991,
246, 1-9.
. .
The intra-area distribution of cortico-cortical projection neurons
is comparable to the distribution of neurofibrillary tangles in the
cortex in Alzheimer's disease.
Reproduction
* Photoperiod regulation of uterine activity and melatonin rhythms in
the pregnant rhesus macaque. Ducsay, C. A. & Yellon, S. M. (Div. of
Perinatal Biology, School of Medicine, Loma Linda, Univ., Loma Linda,
CA 92350). Biology of Reproduction, 1991, 44, 967-974.
. .
Measurements during a 12L:12D cycle with lights on 0700-1900h, were
compared with those during a 12L:12D cycle with lights on 2000-0800h,
in six pregnant monkeys. After 7 days of reversed photoperiod, peak
uterine activity shifted by 12 hours, as did melatonin concentration.
Labor began in all 6 animals during periods of darkness.
* "Environmental stress" and reproductive success in the common marmoset (Callithrix jacchus jacchus). Johnson, E. O., Kamilaris, T. C.,
Carter, S., Gold, P. W., & Chrousos, G. P. (Developmental Endocrinology Branch, NICHHD, NIH, Bldg. 10, Rm. 10N262, Bethesda, MD 20892).
American Journal of Primatology, 1991, 25, 191-201.
. .
Reproductive records of a marmoset colony through: 1) establishment; 2) stability; 3) disruption by nearby construction; 4) return to
stability. The colony exhibited severe reproductive suppression during
the time of environmental disruption. The number of live births
decreased, and the number of spontaneous abortions increased during
this period.
* Harassment of sexual interactions among stumptail macaques, Macaca
arctoides. Drukker, B., Nieuwenhuijsen, K., van der Werff ten Bosch,
J. J., van Hooff, J. A. R. A. M., & Slob, A. K. (A.K.S., Dept. of
Endocrinology & Reproduction, Fac. of Medicine & Health Sciences,
Erasmus Univ., P.O.B. 1738, 3000 DR Rotterdam, Netherlands). Animal
Behaviour, 1991, 42, 171-182.
. .
Harassment, defined here as active behavior directed at copulants
by individuals of lower rank, is triggered by ejaculation and directed
at the mating male. Social and motivational factors are considered,
and it is suggested that harassment reinforces power structures in the
group.
* The role of males in the stimulation of reproductive function in
female cotton-top tamarins, Saguinus o. oedipus. Widowski, T. M.,
Ziegler, T. E., Elowson, A. M., & Snowdon, C. T. (Psychology Dept.,
Univ. of Wisconsin, Madison, WI 53706). Animal Behaviour, 1990, 40,
731-741.
. .
Results of this and previous studies indicate that the onset of
sexual behavior and ovarian cyclicity may require release from suppression not only by the mother, but also by other family members, as well as direct contact or stimulation from an unfamiliar male.
* The macaque model for in vitro fertilization: Superovulation techniques and ultrasound-guided follicular aspiration. VandeVoort, C. A.
& Tarantal, A. F. (California RPRC, Univ. of California, Davis, CA
95616-8542). Journal of Medical Primatology, 1991, 20, 110-116.
. .
Pergonal (hMG) was administered for 7 or 8 days beginning on cycle
day 1 or 2 or for 6 days beginning on cycle day 3, and was followed by
Pregnyl (hCG) prior to follicular aspiration. The quality of oocytes
recovered from the 6-day treatment group was better than those treated
for �7 days. Ultrasound can provide a reliable means for documenting
the response to ovarian stimulation and the successful transabdominal
aspiration of multiple follicles.
* Presence of adult or juvenile relatives increases livebirth chances
in captive stumptail macaques (Macaca arctoides). Santillan-Doherty,
A. M., Mayagoitia, L., & Mondragon-Ceballos, R. (R.M.-C., Inst. Mexicano de Psiquiatria, Lab. de Etologia y Psicobiologia, Camino a Xochimilco 101, Tlalpan, Mexico, DF 14370 Mexico). Journal of Medical Primatology, 1991, 20, 140-142.
. .
Data from 35 deliveries in the 9 fertile adult females in a small colony.
* Hormones and sexual behavior associated with postconception perineal
swelling in the sooty mangabey (Cercocebus torquatus atys). Gordon,
T. P., Gust, D. A., Busse, C. D., & Wilson, M. E. (Yerkes RPRC, Emory
Univ., Atlanta, GA 30322). International Journal of Primatology,
1991, 12, 585-597.
. .
Vaginal bleeding and perineal swelling were charted in 29 subjects,
showing that maximum perineal tumescence corresponded with the periovulatory phase of the cycle, but a second tumescence was recorded during pregnancy, with a peak at 49 days postconception. Behavioral data
showed that, although there was no overall difference in male mount
rate to females during the ovulatory and postconception maximum swelling period, when only the alpha male was considered, significantly
fewer mounts were directed to females exhibiting a postconception versus an ovulatory maximum tumescence.
* Reproductive biology of the vervet monkey (Cercopithecus aethiops):
A review. Eley, R. M. (Inst. of Primate Research, National Museums of
Kenya, Box 24481, Karen, Nairobi, Kenya). Utafiti, 1992, 4[1], 1-33.
. .
A summary of existing information on reproductive anatomy, physiology, and endocrinology of the vervet monkey.
* Exogenous progesterone recycling and early detection of pregnancy in
rhesus monkeys. Harrison, R. M. (Tulane Primate Center 18703 Three
Rivers Rd., Covington, LA 70433). Journal of Medical Primatology,
1991, 20, 323-324.
. .
A method to control ovulation so that conception would occur near a
preselected date, using progesterone injections.
* Induction of multiple follicular development and superovulation in
the olive baboon, Papio anubis. McCarthy, T. J., Fortman, J. D.,
Boice, M. L., Fazleabas, A. T., & Verhage, H. G. (H.G.V., Dept. of
Ob-Gyn, Univ. of Illinois, 840 S. Wood St., Chicago, IL 60612). Journal of Medical Primatology, 1991, 20, 308-314.
. .
Beginning at menses, 19 adult female baboons were treated with hMG
for 10 days, followed by hCG on day 11. Multiple follicular development was seen in all animals, with superovulation in eleven. Ova produced were fertilizable, but development of antibodies against the
human hormones used precludes restimulation.
-------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. (Phone: 401-863-2511)
Judith_Schrier@brown.edu
ACKNOWLEDGMENTS
The Newsletter is supported by U. S. Public Health
Service Grant RR-00419 from the Animal Resources Program,
Division of Research Resources, N.I.H.
Cover photo of a 33-year-old male rhesus (Macaca mulatta)
with a companion juvenile, by Viktor Reinhardt,
University of Wisconsin, Madison
Copyright @1992 by Brown University
Editor: Judith E. Schrier, M. Sc.
Associate Editor: James S. Harper, D.V.M.
Consulting Editor: Morris L. Povar, D.V.M.
Copy Editor: Elva Mathiesen, B. A.
Founding Editor: Allan M. Schrier, Ph.D.
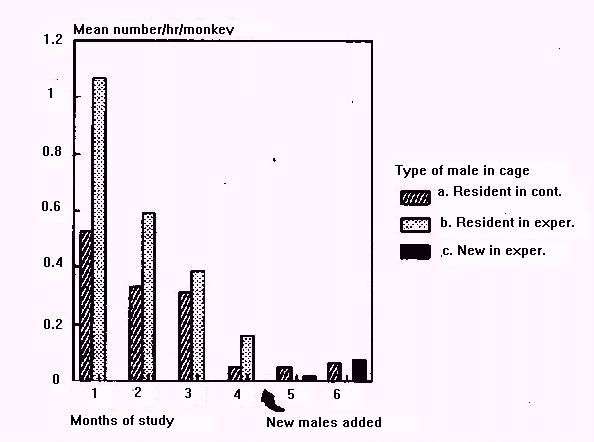{kind=link}
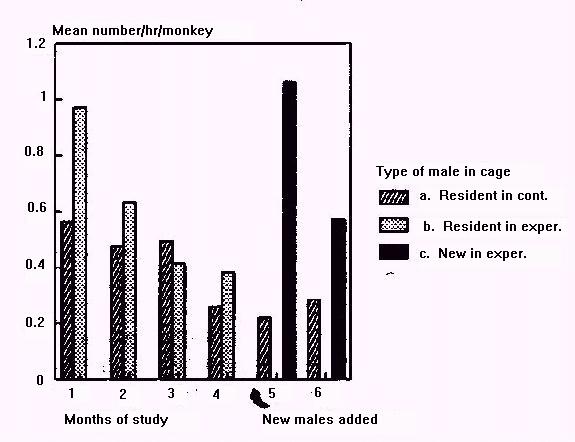{kind=link}
{kind=link}
{kind=link}
{kind=link}