 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 32 NUMBER 4 OCTOBER 1993
Articles and Notes
Foraging Enrichment for Caged Macaques: A Review, by V. Reinhardt...... 1
Responses of Singly-Housed White-Crowned Mangabeys (Cercocebus torquatus lunulatus) to Different Enrichment Devices, by K. Phillippi-Falkenstein...... 5
Initial Response to Introduction of a PVC Perch by Singly-Caged Macaca fascicularis, by M. Shimoji, C. L. Bowers, & C. M. Crockett...... 8
Tuberculosis in Imported Nonhuman Primates -- United States, June 1990-May 1993...... 11
Interim Guidelines for Tuberculin Skin Testing of Nonhuman Primates During Quarantine...... 13
Adoption of a Neonate in a Captive Group of Red-Capped Mangabeys(Cercocebus torquatus torquatus) and the Use of DNA Fingerprinting to Assess Maternity, by E. Watts, D. G. Smith, & E. L. Zucker...... 18
Relative Strength of Influence of Relatedness and Familiarity on Affiliation in Captive Rhesus, by D. R. Rasmussen...... 21
Time to Ban Imports? A Response, by J. Baulu...... 26
Kingston Replies...... 27
News, Information, and Announcements
Planned 1994 Revision of Directory of Graduate Programs in Primatology and Primate Research...... 4
Travellers' Health Notes...... 15
. .
Malaria Reminder; Cholera in Africa;
Schistosomiasis in Malawi, 1992
Report on Working Conference on Ethics in Primatology...... 17
Primate Material Requested or Available...... 20
. .
Tissue for Odorant Binding Protein Research;
Resources for Conducting Research on Aging
Grants Available...... 25
. .
Human Frontier Science Program; Dissertation
Research Funds; Helicobacter pylori Pathogenesis;
Hastings Center; Zoo Research Grants; American Cancer Society
Meeting Report: Zoo Conference on Environmental Enrichment...... 28
News Briefs...... 29
. .
Squirrel Monkey Twins Reported; Digit Fund
Changes Its Name; Malaria-AIDS Link? VandeBerg to Lead ILAR;
New SCAW Board Members; Birute Galdikas Honored; Tonkin Snub-
nosed Langurs
Meeting Announcements...... 30
. .
Wisconsin RPRC Seminars; Behavioral Ecology Congress;
Anesthesia; Analgesia and Surgery; Human/Research Animal
Relationship
Information Requested and Available...... 31
. .
Import Permits; Nonhuman Primate Audiovisual
Resources; Yerkes in Florida; Pathology Slide Set
Letter from Russia -- Boris Lapin......insert
Departments
Address Changes...... 24
Position Available: Primate Veterinarian...... 28
Recent Books and Articles...... 32
* * *
Viktor Reinhardt
Wisconsin Regional Primate Research Center
The 1985 amendment to the USA Animal Welfare Act stipulates that the barren cage environment of laboratory nonhuman primates must be enriched in order to ameliorate the adverse effects attendant upon chronic understimulation. Most attempts to provide enrichment focus on foraging behavior, an activity predominant in wild but virtually absent in caged animals. Macaques voluntarily work for food when given the opportunity to do so (Line et al., 1989; Evans et al., 1989; Reinhardt, 1994a). This indicates that foraging behavior may serve as its own reward (Reinhardt, 1994a). Foraging enrichment aims at fostering the expression of food retrieval behaviors by offering the diet in less accessible ways. The present review evaluates published for- aging enrichment options for caged macaques.
Foraging is defined as the gathering of food (The Random House Dictionary of the English Language, 1987). It includes collecting freely accessible food (e.g., presented in a feeder-box) or retrieving food items that are not directly accessible (e.g., presented in a food dispenser). Unlike feeding, foraging does not include eating behavior such as taking food into the mouth, storing food in cheek pouches, retrieving food from cheek pouches, or chewing and swallowing food.
The majority of enrichment options provide food items in such a way that manipulative and cognitive skills are required to retrieve them.
* Line & Houghton (1987) developed a plexiglass puzzle feeder, measuring 30 x 15 x 5 cm. The apparatus is attached on the outside of the animal's home cage and loaded with primary (ordinary monkey biscuits) or supplemental (e.g., peanuts) food. It has five internal levels, which can be arranged to form various maze patterns. The animal inserts a finger into small holes in the front of the feeder and pushes the food to the bottom level, from which it is removed through a large, central hole. Line et al. (1989) tested five adult rhesus macaques (Macaca mulatta) that had been exposed daily to the biscuit-filled puzzle for four weeks. The animals removed an average of 3.8 biscuits per day despite the fact that other biscuits were freely available. Bloom & Cook (1989) presented the puzzle to two adult single-housed rhesus macaques on several days. The subjects spent an average of 12.5 min to eat 10 biscuits. Their feeding times were extended from 10 min to approximately 25 min when the standard ration of biscuits was presented in the puzzle instead of in the ordinary feeder-box.
* Murchison (1991) developed a puzzle specifically designed for shelled peanuts. The device is made from three 30 cm long sections of PVC pipe, 2.5 cm in diameter. The tubes are stacked parallel and fastened together by a bolt on each end. Interconnecting holes in the bottom of one pipe and the top of the adjoining pipe allow peanuts to be pushed via finger-holes from the upper to the lowest pipe where they can be extracted through a hole. The gadget is attached on the ouside of the animal's cage. Eight subadult cynomolgus macaques (M. fascicularis) were exposed to this puzzle loaded with six peanuts daily for 21 days. The animals removed on average 3.5 peanuts each day. Heath et al. (1992) modified the feeder for unshelled peanuts.
* The Boomer ball is a further variation of the peanut puzzle. It consists of a 10.2 cm diameter hard plastic ball, attached on the ouside of the animal's cage with a chain (Murchison, 1992). Shelled peanuts are loaded into the ball through an access hole. An animal manipulates the device by reaching through a small opening on the front of the cage. Holding the ball upside down with one hand and using the fingers of the other hand, it pulls peanuts from the access hole. Four adult cynomolgus macaques were presented with the puzzle for 32 days. Each morning the balls were loaded with twelve whole peanuts. The subjects spent on average 17.4 min retrieving 6.6 peanuts during the first two hours (Murchison, 1992).
* The plastic tube food puzzle is made of two 5 mm thick, 20 cm long, and 6 cm in diameter clear flexible plastic tubes held together in the shape of an X by an adjustable steel bolt and suspended from a chain attached to the front of the animal's cage (Murchison & Nolte, 1992). Four shelled peanuts are placed inside the tubes near the center and held in place by tension from the compressed tubes. The animal reaches through a small cage opening, holds the device with one hand and uses fingers of the other hand to pull or push peanuts out of the tubes. Twenty-four adult pig-tailed macaques (M. nemestrina), exposed to the daily-loaded puzzle for 20 days, removed on average 3 peanuts each day (Murchison & Nolte, 1992).
* Reinhardt (1993a) created food puzzles by remounting ordinary feeder-boxes a few centimeters away from their original position. Skillful manipulation with the fingers through the 22 mm mesh was then required to retrieve food items. Offering the standard ration of 33 small biscuits (Purina Monkey Chow No. 5038) in "food puzzles" instead of in feeder-boxes during a 1-month habituation period resulted in a 69-fold increase in foraging time (1589 sec vs 23 sec) in 12 singly-caged adult stumptail macaques (M. arctoides ; Reinhardt, 1994b). Eight pair-housed rhesus macaques showed a 141-fold increase in foraging time (2532 sec vs 180 sec) under the same conditions (Reinhardt, 1993a). Subjects spent on average 60 sec retrieving one biscuit from the puzzle (Reinhardt, 1994a).
* Placing the daily standard ration of small biscuits directly on the 22 mm mesh ceiling of the cages instead of in feed-boxes or food puzzles induced a 80-fold (1363 sec vs 17 sec) increase in foraging time in eight pair-housed rhesus macaques (Reinhardt, 1993b).
* Moazed & Wolff (1988) described a portable polypropylene pick-up board measuring 9 x 30.5 x 2.6 cm. The device has 19 equally spaced 8-mm-diameter holes into which raisins are inserted from either side. Evans et al. (1989) used a similar board, consisting of * an ice cube tray (23 cm x 10 cm) with 90 compartments, each holding a raisin or marshmallow. Twenty-eight singly-caged cynomolgus macaques required on average 8.7 sec to retrieve one raisin and 2.5 sec for one marshmallow (Evans et al., 1989).
Some foraging enrichment options promote food gathering activity without requiring special manipulative skills.
* Harris (1988) replaced the mesh floor of 10 singly-caged cynomolgus macaques with a removable solid metal tray filled with a woodchips/food mix every other day. Average feeding time increased from 2.7 min to 3.3 min per 30 min observation session when food [type not indicated] was offered in woodchips rather than in feeder-boxes (Harris, 1988). Bryant et al. (1988) exposed six singly-caged cynomolgus macaques to * woodchip litter scattered with sunflower seeds and peanuts. The animals engaged in foraging for approximately 10 min during 30-min observation sessions (cf. Rupniak & Iverson, 1989).
* Bayne et al. (1991) developed a "foraging/grooming" board that is bolted to the outside of the animal's cage. The board is a 0.1 m-square piece of plexiglass covered with artificial shearling. Tiny bits of flavored food are rubbed into the shearling daily. The authors exposed eight singly caged adult rhesus macaques to the board for six months. It induced the subjects to show a combination of foraging/eating/grooming behaviors on average 12.1 min during the first 30 min after the board was baited with food.
* The foraging board consists of plexiglass covered with artificial turf. It occupies approximately one third of the depth of the cage, and is replenished with food particles each day (Bayne et al., 1992). Eight adult individually caged rhesus macaques received the board for six months. They spent an average of 15.7 min feeding from the board during twenty 30-min sessions immediately following the distribution of food crumbles.
* Lam et al. (1991) created a portable fleece cush- ion (20 x 20 x 60 cm). Like Bayne's foraging/grooming board, the cushion is sprinkled with tidbits of flavored food. Three adult individually caged cynomolgus macaques received the cushion daily for one hour on six consecutive days. The average amount of time spent feeding during the first 60 min immediately after introduction of the fleece was 6.7 min when unfamiliar flavors, 24.0 min when familiar flavors were employed. Food dispensers indirectly promote foraging by making the animal manipulate a mechanical device to get food delivered.
* Bramblett & Bramblett (1988) described a liquid dispenser. The body of the pump feeder is constructed from a PVC pipe approximately 5 cm in diameter, permanently capped at one end. A hole drilled in the center of the threaded cap at the other end accommo- dates a 1 cm diameter pipe that serves as plunger. Small notches cut in the side of the hole allow fluid to adhere to the plunger when it is raised. The pump is filled with a sticky fluid, such as frozen juice concentrate, and mounted on a vertical part of the cage.
* Markowitz & Line's (1989) dispenser consists of a metal battery-operated box (measuring approximately 25 x 30 x 10 cm) containing a radio and a banana-flavored pellet dispenser. Contact detectors, extending approximately 2 cm into the cage, provide independent controls for turning the radio on and off and for activating the food dispenser. Five single-caged adult rhesus macaques were continuously exposed to this device for 20 weeks. The animals retrieved an average of 562 banana pellets per day (Line et al. 1991). The radio was played an average of 76 min/day.
* Gullekson et al. (1991) described a seed dispenser for singly housed cynomolgus macaques. The apparatus is also attached to the outside of the animal's cage front. It consists of a polyethylene ring (approximately 13.5 cm in diameter, 1.5 cm wide, 1.5 cm thick) with a small depression. The ring turns through a PVC tube containing sunflower seeds. By turning the ring, the subject can remove seeds that are caught in the depression.
* Crockett et al., (1989) converted hollow rubber Kong toys into foraging devices by filling them with frozen apple juice. Eight macaques (three different species, different age classes) received a rubber toy filled with 36 ml of frozen juice daily for 7 weeks. The animals engaged in foraging-related activities approximately 3 min per 1-hour observation session.
A total of 15 different foraging enrichment options have been described for caged macaques. Eleven of them have been specifically designed as secondary feeders (for peanuts, raisins, marshmallows, flavored tidbits, banana pellets, liquid food, frozen juice). Because of the relatively high labor (baiting, cleaning, maintenance) and material investment, their practical use will be restricted to facilities with small numbers of animals. Only four foraging enrichment options (plexiglass puzzle, food puzzle, mesh ceiling, woodchips) are designed in such a way that they can be used both as secondary and primary feeders (for standard biscuit ration). They will be useful for facilities housing high numbers of animals because of their relative labor-inexpensiveness. The most inexpensive option is the ordinary food box converted into a food puzzle; it requires no extra time for baiting, cleaning and maintenance. The foraging-promoting effectiveness of the various options described cannot readily be compared because of the differences in food items used (e.g. supplemental raisins vs supplemental peanuts vs ordinary biscuits), and the heterogeneity of data collected (e.g., assessment of feeding time vs assessment of foraging time vs assessment of number of food items removed/time unit).
References
Bayne, K., Mainzer, H., Dexter, S., Campbell, G., Yamada, F., & Suomi, S. (1991). The reduction of abnormal behavior in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology, 23, 23-35.
Bayne, K., Dexter, S., Mainzer, H., McCully, C., Campbell, G., & Yamada, F. (1992). The use of artificial turf as a foraging substrate for individually housed rhesus monkeys (Macaca mulatta). Animal Welfare, 1, 39-53.
Bloom, K. R. & Cook, M. (1989). Environmental enrichment: Behavioral responses of rhesus to puzzle feeders. Lab Animal, 18 [5], 25-31.
Bryant, C. E., Rupniak, N. M. J., & Iversen, S. D. (1988). Effects of different environmental enrichment devices on cage stereotypies and autoaggression in captive cynomolgus monkeys. Journal of Medical Primatology, 17, 257-269.
Crockett, C., Bielitzki, J., Carey, A., & Velez, A. (1989). Kong toys as enrichment devices for singly-caged macaques. Laboratory Primate Newsletter, 28 [2], 21-22.
Evans, H. L., Taylor, J. D., Ernst, J., & Graefe, J. F. (1989). Methods to evaluate the well-being of laboratory primates. Comparison of macaques and tamarins. Laboratory Animal Science, 39, 318-323.
Gullekson, R., Bench, L., Harrigan, K., & Pyle, K. (1991). Seed-feeder as a foraging device for singly-housed cynomolgus monkeys (Macaca fascicularis). Lab Animal, 20 [6], 44-46.
Harris, D. (1988). Welfare and Housing of Old World Nonhuman Primates (Macaca fascicularia and Papio sp.). Potters Bar, U.K.: Universities Federation for Animal Welfare.
Heath, S., Shimoji, M., Tumanguil, J., & Crockett, C. (1992). Peanut puzzle solvers quickly demonstrate aptitude. Laboratory Primate Newsletter, 31 [1], 12-13.
Lam, K., Rupniak, N. M. J., & Iversen, S. D. (1991). Use of a grooming and foraging sub- strate to reduce cage stereotypies in macaques. Journal of Medical Primatology, 20, 104-109.
Line, S. W. & Houghton, P. (1987). Influence of an environmental enrichment device on general behavior and appetite in rhesus macaques. Laboratory Animal Science, 37, 508.
Line, S. W., Markowitz, H., Morgan, K. N., & Strong, S. (1989). Evaluation of attempts to enrich the environment of single-caged nonhuman primates. In J. W. Driscoll (Ed.), Animal Care and Use in Behavioral Research: Regulations, Issues and Applications (pp. 103-117). Beltsville, MD: National Agricultural Library.
Line, S. W., Markowitz, H., Morgan, K. N., & Strong, S. (1991). Effects of cage size and environmental enrichment on behavioral and physiological responses of rhesus macaques to the stress of daily events. In M. A. Novak & A. J. Petto (Eds.), Through the Looking Glass; Issues of Psychological Well-Being in Captive and Laboratory Primates (pp. 160-179). Washington, DC: American Psychological Association.
Markowitz, H. & Line, S. (1989). Primate research models and environmental enrichment. In E. F. Segal (Ed.), Housing, Care and Psychological Well-Being in Captive and Laboratory Primates (pp. 202-212). Park Ridge, NJ: Noyes Publications.
Moazed, T. C. & Wolff, A. V. (1988). The raisin board as an environmental enrichment tool for laboratory primates. Laboratory Primate Newsletter, 27[1], 16.
Murchison, M. A. (1991). PVC-pipe food puzzle for singly caged primates. Laboratory Primate Newsletter, 30[3], 12-14.
Murchison, M. A. (1992). Task-oriented feeding device for singly caged primates. Laboratory Primate Newsletter, 31[1], 9-11.
Murchison, M. A. & Nolte, R. E. (1992). Food puzzle for singly caged primates. American Journal of Primatology, 27, 285-292.
Reinhardt, V. (1993a). Enticing nonhuman primates to forage for their standard biscuit ration. Zoo Biology, 12, 307-312.
Reinhardt, V. (1993b). Using the mesh ceiling as a food puzzle to encourage foraging behaviour in caged rhesus macaques (Macaca mulatta). Animal Welfare, 2, 165-172.
Reinhardt, V. (1994a). Caged rhesus macaques voluntarily work for ordinary food. Primates, 35, in press.
Reinhardt, V. (1994b). Promoting increased foraging behavior in caged stumptail macaques. Folia Primatologica, in press.
Rupniak, N. M. J. & Iverson, S. D. (1989). Psychological welfare, behaviour, and housing conditions for laboratory primates: A discussion. Laboratory Animal Welfare Research Primates (pp. 31-38). Potters Bar: Universities Federation for Animal Welfare.
-------------------------------------------------------------------
Author's address: Wisconsin Regional Primate Research Center, 1223 Capitol Court, Madison, WI 53715. Supported by NIH grant RR-00167. Publication No. 33-016 WRPRC.
* * *
Kathrine Phillippi-Falkenstein
Tulane Regional Primate Research Center
Introduction
Many types of device, as well as various housing styles and configurations, have been studied to find the most effective forms of enrichment for captive nonhuman primates (see Fajzi et al., 1989 for a review). Two enrichment devices (foraging/grooming boards and peanut puzzle feeders) were in use at Tulane Regional Primate Research Center when five new devices (turf foraging feeder, finger feeder, shaker puzzle, perch/swing, and rubber pull) were received from Bio-Environmental Modifiers (Olsburg, KS) to test on monkeys for a limited time. While most studies have been conducted on macaques and chimpanzees (Boysen et al., 1989; N.A.L. et al., 1992), we decided to test these new devices on mangabeys for a four-week period. The following paper describes the responses of singly-housed white-crowned mangabeys (Cercocebus torquatus lunulatus) to the various enrichment devices.
Methods
Subjects: Subjects were 8 wild-born, female white-crowned mangabeys, 2.8 to 4.8 years of age. All subjects were singly-housed and were unfamiliar with all enrichment devices used.
Procedure: Animals were matched for age. Each pair received two enrichment devices presented on an "ABBA" schedule over the 4-week period. A fleece-covered foraging/grooming board (see Bayne et al., 1991) was alternated with a turf foraging feeder (similar to that described in Bayne et al., 1992, except horizontally mounted outside a cage). A peanut puzzle feeder (see Murchison, 1991) was alternated with either a finger feeder or a shaker puzzle. A finger feeder is a clear acrylic cylinder (with a hole for extracting contents) hung on the outside of the cage, while a shaker puzzle has a dispensing tube and is placed inside the cage. A perch/swing or a rubber pull was alternated with periods of no enrichment. The perch/swing is a clear acrylic tube attached to the top of a cage by chains and clips at both ends of the tube. The pull is made of yellow rubber in a triangular shape with a chain and clip at one apex for attachment to the cage top. Prima-foraging crumbles@ were used with the fleece and turf boards; a combination of crumbles and shelled peanuts with the finger feeder and shaker puzzle; and unshelled peanuts with the peanut puzzle feeder.
Behavioral observations were done three times a week between 3:00 and 4:30 PM after the animal caretakers had left the area. After forage was placed in the devices, ten-minute samples of all occurences of behaviors (including frequency and duration of stereotypic behavior and apparatus-directed behavior) were taken for each animal according to a pre-determined, randomized order (Altmann, 1974). Data were recorded using Observer 2.0 (Noldus, 1991) on a portable computer. A total of 16 hours (2 hours/animal) of data were collected.
Analysis: Data were analyzed by individual device, by device combination, and by week using one-way analysis of variance. To determine if there were order effects, differences between the first device presented (A) and the second one (B) were tested with paired t-tests for frequency of stereotypic behavior, duration of stereotypic behavior, and duration of device use.
Results
Individual Device: Both the total frequency and duration of stereotypic behavior were lower when food-related devices (especially the fleece or shaker) were present than when the perch or pull was present (see Figure 1a-b). Food-related devices (especially the turf and shaker) also were used for longer periods than the perch or pull ( F6,7 = 6.74, p = .012; Figure 1c). No differences in stereotypic or device-directed behavior were found between the food-related devices.
Device Combination: Animals with the turf/fleece combination exhibited fewest stereotypic behaviors ( F 3,12 = 4.11, p = .032). The puzzle/finger combination and the puzzle/shaker combination were associated with shorter bouts of stereotypic behavior ( F 3,12 = 5.08, p = .017).
By Week: For animals with enrichment throughout the study, the frequency of stereotypic behavior occurring during week 3 was lower than during week 1 ( F 5,3 = 5.29, p = .011; Figure 2a-c). For animals with periods of no enrichment, the duration of stereotypic behavior dropped significantly after week 1 ( F 1,3 = 18.42, p = .020; Figure 2b).
Order of Presentation: For animals with enrichment throughout the study, there were no significant differences in the frequency or duration of stereotypic behaviors or the duration of device use between the first (A) and the second (B) device presented, regardless of device type. For animals with periods of no enrichment, the duration of stereotypic behavior significantly declined with enrichment ( t2 = 4.58, p = .045).
Discussion
While the effects of novelty cannot be eliminated in a short-term study, it appears that providing mangabeys with enrichment devices both increases species-typical behavior and reduces stereotypic behavior (following Novak & Suomi, 1988). Alternating food-related devices was associated with the maintenance of animals' interest and reduced levels of stereotypic behavior. For animals with constant enrichment, time spent using devices increased throughout the study. Thus, the benefits of providing foraging opportunities, which has been documented in other species (e.g. Lam et al., 1991; Bayne et al., 1991, 1992), applies also to white-crowned mangabeys.
It can be argued that the observation periods were times of increased use due to the recent placement of forage (e.g., Watson, 1992), but these devices were often found taken apart or hanging askew at other times of day, indicating that they were used even when no forage was present. Even the perch and pull received ad lib use and lower occurrences of stereotypic behavior were associated with its presence in the cage. Stereotypic behavior did increase slightly following the removal of the perch or pull, but did not return to preenrichment rates (week 1). It is possible that the lower rates of stereotypic behavior seen were due to the presence of observers (see Bayne et al., 1993).
Enrichment devices should be durable, easy to present, remove, and clean, and should not compromise personnel safety. The wing nuts on the bottom of the turf feeder were easily disassembled by our mangabeys, allowing them to pull it up and chew on the turf. The turf feeder was easier to load with forage than the fleece board, and it did not have to be removed during daily room cleaning. The shaker puzzle was easily and often disassembled by the mangabeys and the edges became jagged. The shaker puzzle also had to be placed inside the cage. Both these devices received considerable use and had a positive effect on the animals, but modifications are necessary to prevent hazards to animals or caretakers.
Providing enrichment devices reduced, but did not eliminate, stereotypic behaviors in our mangabey subjects. The order of device presentation was not important, but food-related enrichment proved most effective. All of the enrichment devices appeared to have positive effects on the animals.
References
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 48, 227-2267.
Bayne, K., Mainzer, H., Dexter, S., Campbell, G., Yamada, F., & Suomi, S. (1991). The reduction of abnormal behavior in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology, 23, 23-35.
Bayne, K., Dexter, S., Mainzer, H., McCully, C., Campbell, G., & Yamada, F. (1992). The use of artificial turf as a foraging substrate for individually housed rhesus monkeys (Macaca mulatta). Animal Welfare, 1, 39-53.
Bayne, K. A. L., Dexter, S. L., & Strange, G. M. (1993). The effects of food treat provisioning and human interaction on the behavioral well-being of rhesus monkeys (Macaca mulatta). Contemporary Topics in Laboratory Animal Science, 32, 6-9.
Bloom, K. R. & Cook, M. (1989). Environmental enrichment: Behavioral responses of rhesus to puzzle feeders. Lab Animal, 18, 26-31.
Boysen, S. T., Quigley, K. S., & Woods, V. R. (1989). Primate enrichment: An annotated bibliography. The Ohio State University and Columbus Zoological Garden Cooperative Research Program.
Fajzi, K., Reinhardt, V., & Smith, M. D. (1989). A review of environmental enrichment strategies for singly caged nonhuman primates. Lab Animal, 18, 23-35.
Lam, K., Rupniak, N. M. J., & Iversen, S. D. (1991). Use of a grooming and foraging sub- strate to reduce cage stereotypies in macaques. Journal of Medical Primatology, 20, 104-109.
Murchison, M. A. (1991). PVC-pipe food puzzle for singly caged primates. Laboratory Primate Newsletter, 30[3], 12-14.
Nat. Agricultural Lib., Nat. Lib. of Med., & Primate Information Center. (1992). Environmental enrichment information resources for nonhuman primates: 1987-1992. Beltsville, MD: N.A.L.
Noldus, L. P. J. J. (1991). The Observer: A software system for collection and analyses of observational data. Behavior Research Methods, Instruments, and Computers, 23, 415-429.
Novak, M. A. & Suomi, S. J. (1988). Psychological well-being of primates in captivity. American Psychologist, 43, 765-773.
Watson, L. M. (1992). Effect of an enrichment device on stereotypic and self-aggressive behaviors in singly-caged macaques: A pilot study. Laboratory Primate Newsletter, 31[3], 8-10.
-------------------------------------------------------------------
Author's address: Dept. of Veterinary Sciences, Tulane Regional Primate Research Center, 18703 Three Rivers Rd, Covington, LA 70433. This research was supported by a NIH grant to Tulane RPRC (R00164). Animals used in this study were supported by a grant to B. J. Gormus (NIAID #19302). The author wishes to thank Dr. Gormus for his support, Dr. Margaret R. Clarke for her research assistance, support, and critical evaluation of this manuscript, and Dr. Evan L. Zucker for critical evaluation of this manuscript.
* * *
Mika Shimoji, Charles L. Bowers, and Carolyn M. Crockett
Regional Primate Research Center, University of Washington
Introduction
Enrichment of the captive environment through cage furnishings is one way to promote the psychological well-being of research animals. Longtailed macaques (Macaca fascicularis), one of the most commonly used laboratory primates, exhibit a vertical flight response when alarmed (Crockett & Wilson, 1980). Therefore, the height of a cage is important for allowing the animal to withdraw from potentially stressful or alarming situations in the laboratory setting. Perches can provide more usable vertical space. In this study, we investigated the use of a PVC perch by individually-housed adult longtailed macaques during the first 3 days after the perch was installed. Since our subjects were wild-caught, and given the arboreal nature of this species, we anticipated immediate use of the PVC perch. Based on other studies, we expected monkeys in lower level cages to use the perch more than those in upper level cages (Reinhardt, 1989; Woodbeck & Reinhardt, 1991).
Method
Subjects and Housing: Subjects were 10 female and 10 male wild-caught adult longtailed macaques that had been used in previous studies (Crockett et al., 1993a, b). None had previous exposure to the PVC perch. Prior to this study, they had occupied cages without perches for 3 years, housed individually except for limited pair housing for about 6 weeks (Crockett et al., 1993a). Housing and maintenance are described elsewhere (Crockett et al., 1993a, b).
Procedure: No Perch Phase: Videotaped behavioral data were collected for 4 days about 2 months before the perch was installed. Perch Phase : On day 1, a PVC perch (four 2.6 x 36 cm tubes connected in parallel) was attached with snap hooks to the back of the cage 27 cm from the floor, after routine cage washing. The perch remained in the cage for 3 days and nights, after which the monkeys were moved to another room where video monitoring was not possible.
Data Collection: Behavior was coded from videotapes recorded remotely during the day (0600h - 1730h, 1 min every 30 min/subject, continuous and point sampling) and during the night (1800h - 0530h, 0.5 min every 30 min/subject, point sampling). No human was present except during room cleaning and feeding. Behaviors were coded with a 3-digit number system: the 1st digit coded location, the 2nd digit coded behavior category, and the 3rd digit indicated normal vs. abnormal/stereotypic behavior.
Analysis: Daytime behaviors were expressed as mean sec/min or percentage of observation time. Nighttime data were examined only for use of the perch. Statistical tests included nonparametric Mann-Whitney U, Wilcoxon Signed Ranks, Spearman correlation (Wilkinson, 1989), and parametric General Linear Model multiway ANOVA (with log transformation when required to improve normality) (Velleman, 1993). All probability levels are two-tailed unless stated to be one-tailed.
Results
All 20 subjects contacted the perch (sitting on or manipulating it), a mean latency of 14.1 hours following its introduction. Unexpectedly, 6 monkeys (4 females and 2 males) never sat on it during the first 3 days. Overall, monkeys spent an average of 20% of daytime on the perch (29% by the 14 that actually sat on it) and 2% of the time manipulating it. Subsequent informal observations indicated that all subjects eventually sat on the PVC perch on a daily basis, although percentage of time spent on the perch was not quantified.
Perch use by cage level and day since introduction: Monkeys in lower cages tended to sit on the perch more than those in upper cages (lower mean = 15.6 +/- 3.5 SE sec/min, upper mean = 8.5 +/- 3.3 SE sec/min), but overall the cage level factor was not statistically significant (P = 0.16, ANOVA). However, time sitting on the perch increased significantly over the 3 days following its introduction, and lower level monkeys spent significantly more time on the perch on day 2 than did upper level monkeys (Mann-Whitney U, P < 0.05, one-tailed). There were no significant sex differences in use of the perch or latency to first contact with it. By day 3, the monkeys were sitting on the perch 25% of the daytime.
Time spent above floor level with and without perch: Monkeys spent significantly more time clinging to the cage wall ("suspended") in the absence of the perch, 7% of the time vs. 4% when the perch was present (P = 0.02, ANOVA). Regardless of perch presence, females spent significantly more time suspended than males (P = 0.002, ANOVA). The total time spent above the floor area ("up" = "suspended" + "on perch") increased significantly when the perch was present (from 7% to 24%; P = 0.0001, ANOVA; in the no perch condition, "up" = "suspended"). There was a significant interaction in time "up" between sex and perch presence (P = 0.05, ANOVA): when provided with a perch, males increased time spent "up" more than females. When the perch was present, the sexes did not differ in time spent "up" (P = 0.94, Mann-Whitney U).
Other changes in behavior: Time spent in stereotypic behaviors decreased significantly when the perch was present compared with the no-perch baseline (4.6% to 2%; P = 0.02, Wilcoxon Signed Ranks, n = 20). However, the correlation between time in stereotypy and using the perch (on + manipulating it) was not significant (Spearman R = -0.27, P > 0.20, n = 20).
We speculated that the decrease in stereotypy might be related to general behavioral suppression in the presence of a novel object. However, inactivity (a measure of behavioral suppression) instead declined significantly when the perch was present (p < 0.01, ANOVA). Also, inactivity and stereotypy were not significantly correlated (Spearman R = 0.014 when the perch was present and R = -0.082 when it was absent).
Discussion
Although not all subjects used the PVC perch as soon as expected, the percentage of time using it was similar to those reported for rhe- sus macaques (see Table below). Perch use increased significantly over the first 3 days, but 6 monkeys did not sit on the perch until the study was over. Monkeys in lower level cages averaged somewhat more time on the perch than those in upper cages, but, unlike previous studies, the difference was not statistically significant except for day 2. We suspect that the presence of an observer in other studies might make monkeys in lower level cages more likely to retreat to the perch.
+-----------------+-----------------+-------------+----------+-------+ | Reference | Species/sex/age | Housing/ |% of time |Overall| | | | level | on perch |average| +-----------------+-----------------+-------------+----------+-------+ |Shimoji, Bowers, | M. fascicularis | single | | 25.1% | |& Crockett |10 adult females,| Upper | 19.2% | | |(day 3, | 10 adult males | Lower | 31.0% | | |this study) | | | | | +-----------------+-----------------+-------------+----------+-------+ |Reinhardt (1989) | M. mulatta | single | | 28.0% | | | 25 adult | Upper | 16.3% | | | | males | Lower | 48.4% | | +-----------------+-----------------+-------------+----------+-------+ |Woodbeck & | M. mulatta | paired | | 19.2% | |Reinhardt (1991) | 28 adult |(double unit)| | | | | females | Upper | 6.9% | | | | | Lower | 31.6% | | +-----------------+-----------------+-------------+----------+-------+ |Reinhardt (1990b)| M. mulatta | paired | | | | | 60 (42 adult, | (double | | | | | 18 subadult; | unit) | | | | | 36 female, | Upper | | | | | 24 male) | (all) | 10.2% | | +-----------------+-----------------+-------------+----------+-------+ |Reinhardt (1990a)| M. mulatta | single | | | | | 20 males | Lower | | | | | (4 yr old) | (all) | 43.1% | | +-----------------+-----------------+-------------+----------+-------+ |Kopecky & | M. mulatta | paired | |12.2 % | |Reinhardt (1991) | 14 adult females| | | | +-----------------+-----------------+-------------+----------+-------+ |Bayne, Hurst, & | M. mulatta | single | | | |Dexter (1992) | | (double | |16.8 % | | | 8 adult males | cage unit) | | | +-----------------+-----------------+-------------+----------+-------+
Table 1
The perches appeared to fill a behavioral need of the monkeys. This was demonstrated by the fact that clinging to the cage side ("suspended") decreased significantly when the perch was provided. Furthermore, males, who rarely clung to the cage side, increased their time above floor level more than did females in the presence of the perch. The net effect of providing perches was for both sexes to spend a nearly equal amount of time above floor level and thus use more of the vertical cage space.
There was less stereotypy when the perch was present, but we could find no direct effect of perch use (on or manipulating it) to explain its decline. Overall behavior was not suppressed since inactivity actually declined in the perch phase. Since data were collected on only 3 days, we cannot draw conclusions about long-term effects on stereotypic behavior. However, informal observations indicate that perch use probably exceeds the 25% of the time found on day 3 of this study.
Conclusions
*Adult longtailed macaques housed in individual cages spent an average of 25% of the daytime on perches by the third day following perch installation. This average includes 6 monkeys that never sat on the perch during the first 3 days. All 20 monkeys eventually used the perch on a daily basis.
*Perches are a type of cage furnishing that meets the intent of the environmental enrichment requirement of the USDA animal welfare rules and the IPS international guidelines (U. S. Department of Agriculture, 1991; International Primatological Society, 1993).
*We recommend that all laboratory cages for longtailed macaques be equipped with perches except when contraindicated by age (too young or old to climb agilely), health, or research protocol.
References
Bayne, K. A. L., Hurst, J. K., & Dexter, S. L. (1992). Evaluation of the preference to and behavioral effects of an enriched environment on male rhesus monkeys. Laboratory Animal Science, 42[1], 38-45.
Reinhardt, V. (1990a). Comparing the effectiveness of PVC perches versus wooden perches as environmental enrichment objects for singly caged rhesus monkeys. Laboratory Primate Newsletter, 29[1], 13-14.
Reinhardt, V. (1990b). Time budget of caged rhesus monkeys exposed to a companion, a PVC perch, and a piece of wood for an extended time. American Journal of Primatology, 20, 51-56.
U. S. Department of Agriculture. (1991). Animal Welfare, Standards, Final Rule (Part 3, Subpart D: Specifications for the humane handling, care, treatment, and transportation of nonhuman primates). Federal Register, 56, 6495-6505.
Velleman, P. F. (1993). Data Desk: The New Power of Statistical Vision. Ithaca, NY: Data Description Inc.
Wilkinson, L. (1989). SYSTAT: The System for Statistics. Evanston, IL: SYSTAT, Inc.
Woodbeck, T. & Reinhardt, V. (1991). Perch use by Macaca mulatta in relation to cage location. Laboratory Primate Newsletter, 30[4], 11-12.
-------------------------------------------------------------------
Authors' address: Regional Primate Research Center, SJ-50, University of Washington, Seattle, WA 98195.
* * *
Nonhuman primates (NHPs) shipped to the United States must be quarantined and monitored for evidence of infectious diseases transmissible to humans. During May-October 1992, CDC investigated reports from NHP importers of mycobacterial infections in NHPs recently imported from Mauritius. This report describes a review of tuberculosis (TB) in all 249 imported shipments of CDC-permitted NHP species in the United States from June 1990 through May 1993 and updates recommendations regarding the identification and control of TB in imported NHPs and in workers exposed to such animals.
To determine mycobacterial infection rates in recently imported NHPs, in May 1993 CDC reviewed health records of all 249 shipments of imported cynomolgus (Macaca fascicularis), African green (Cercopithicus aethiops), and rhesus (Macaca mulatta) monkeys. These NHPs had completed a minimum 31-day import quarantine during June 1990-May 1993 under special permits issued by CDC (1990a, 1991). The review included 22,913 NHPs (20,580 cynomolgus, 1621 rhesus, and 712 African green monkeys) from nine countries (Barbados, Canada, China, Indonesia, Mauritius, Myanmar [Burma], the Philippines, Saint Kitts, and Tanzania).
Information was obtained from health records maintained for animals at all 18 NHP quarantine facilities that held special permits during this period and from records at five other facilities that received previously quarantined NHPs. All animals had received routine tuberculin skin tests (TSTs) (three tests, with 2-week intervals between tests) by experienced personnel in well-established quarantine facilities using accepted methods and a U.S. Department of Agriculture-licensed skin-test antigen. A diagnosis of TB infection was based on TST reactions and/or histopathologic examination or culture of granulomatous lesions found at necropsy.
Of the 20,580 cynomolgus monkeys, 8910 (43%) originated in the Philippines, 7703 (37%) in Indonesia, and 3967 (19%) in Mauritius. Of the 1621 rhesus monkeys, 1515 (93%) originated in China, 68 (4%) in Myanmar, and 38 (2%) in Canada. Of the 712 African green monkeys, 394 (55%) originated in Saint Kitts, 198 (28%) in Barbados, and 120 (17%) in Tanzania.
Overall, evidence of TB infection was identified in 90 (81 cynomolgus and nine rhesus monkeys) (0.4%) of the 22,913 NHPs, representing 17 (7%) of the 249 shipments. Sixteen of the shipments contained cynomolgus monkeys. Within these shipments, the prevalence of infection ranged from 0.8% (one of 130 cynomolgus monkeys shipped) to 80% (48 of 60 cynomolgus monkeys), with a median rate of 3%.
The prevalence of TB infection in cynomolgus monkeys varied by country of origin and included 2% (76 of 3967) of animals from Mauritius, 0.04% (three of 7703) from Indonesia, and 0.02% (two of 8910) from the Philippines. In addition, of the 81 total infected cynomolgus monkeys, 76 (94%) originated in Mauritius.
TB-infected NHPs were identified from 11 (69%) of the 16 total cynomolgus monkey shipments from Mauritius. Eight of these 11 shipments arrived from August 1991 through February 1992; the most highly infected shipment arrived during November 1991. TB surveillance and eradication efforts were increased by the exporters in Mauritius after U.S. importers notified them of the identification of TB-infected animals in May 1992. Of the seven shipments received since February 1992, two shipments have contained one TST-positive animal each. No evidence of additional secondary transmission has been reported by importers or by facilities receiving animals after quarantine.
Evidence of TB was detected during the initial 31-day quarantine period in 11 (65%) of the 17 shipments. However, most (69 [77%] of 90) infected animals were in the six shipments in which infection was first identified only after the completion of the routine 31-day postimport quarantine period, including one infected animal identified more than 15 months after release from quarantine. When 16 apparently healthy, TST-negative cynomolgus monkeys were euthanized to control the spread of infection at one facility, nine (56%) were found at necropsy to have granulomatous lesions suggestive of TB. Infection was confirmed by the presence of acid-fast bacilli (histopathologic sections) and/or positive mycobacteriologic culture.
During the review process, one NHP facility worker was reported to have developed a positive TST after exposure to infected animals but did not develop active TB. In addition, facility managers reported that TST conversion occasionally occurs among long-time NHP workers.
CDC Editorial Note: Regulation of the importation and quarantine of NHPs was instituted in 1948 to prevent the introduction and transmission of human pathogens. Beginning in 1975, importers of NHPs for permitted scientific, educational, or exhibition purposes were required to register with CDC, and a system was established for monitoring and reporting disease in quarantine facilities(1). Following the identifi- cation of filovirus infection among cynomolgus monkeys imported from the Philippines during 1989-1990, CDC intensified disease-control measures for the handling of NHPs during importation and quarantine, including 1) unannounced inspection of registered importer quarantine facilities; 2) special permit requirements for the handling of imported cynomolgus, African green, and rhesus monkeys during transit and quarantine; 3) monitoring of arriving shipments of NHPs at U.S. ports of entry; and 4) surveillance of postimportation quarantine and distribution procedures (CDC, 1990a, b).
CDC regulations require that newly imported NHPs undergo a 31-day quarantine period in the United States. Many registered importers and recipients of imported NHPs extend quarantine beyond the 31-day minimum. Because TB in captive NHPs is both an animal and a human health problem, surveillance by TST of imported NHPs and workers at NHP facilities is routine (ILAR, 1980, 1985). Any NHP with a positive TST during import quarantine is considered to be infectious and to repre- sent a high risk for disease transmission. When a quarantined TST-positive NHP is identified, the standard practice is to euthanize the animal, attempt laboratory confirmation of TB, and reinstitute quarantine and tuberculin skin testing (at 2-week intervals) of all other exposed NHPs until three consecutive negative TSTs have been completed (ILAR, 1980)
The findings in this report indicate that the risk for TB infection in cynomolgus monkeys recently imported from Mauritius was substantially higher than among those imported from Indonesia or the Philippines. However, all macaques are considered to be highly susceptible to TB, and virtually all are imported from areas of the world with high prevalences of TB in humans and animals. Close confinement of these and other NHPs in holding facilities and shipping crates creates conditions whereby one infected animal could potentially infect many others. Cynomolgus monkeys from Mauritius may have been held longer in identifiable groups than animals from other geographic areas; they are especially desirable for breeding colonies in the United States because they are free of herpes B, a pathogen fatal to humans and for which macaques are the natural hosts. The longer quarantine may have increased both the likelihood of transmission and detection of TB.
In the investigation described in this report, nine TST-negative animals had evidence of TB. The TST is less reliable for NHPs than for humans because of timing of the TST relative to individual exposure, lack of uniformity in testing procedures, and differences in host responses to the skin-test antigen. Ancillary TB diagnostic procedures commonly used in humans (e.g., booster or second-strength TST, chest radiographs, and sputum smears) are not standard elements of the NHP quarantine protocols.
The potential for transmission of TB and other pathogens among NHPs and humans underscores the importance of improved surveillance and testing procedures in NHP quarantine and research facility settings. The American Association for Accreditation of Laboratory Animal Care requires that its accredited facilities routinely perform TSTs in NHPs; however, test results are not routinely reported to any federal agency. CDC's NHP permit system is the only national disease-reporting system for imported NHPs, and TB reporting has not been required under this system. Daily contact with groups of potentially infected animals in the quarantine facility work environments may present an increased risk for exposure for humans. Although workers whose skin tests convert are referred to their health-care providers for medical evaluation, reporting of these cases with information on occupational exposure is not required by any federal public health agency, and therefore, background incidence and prevalence data for workers in this industry are not available.
The findings in this report indicate the need for improved estimates of the risks for TB in both NHPs and human contacts. The Division of Quarantine in CDC's National Center for Prevention Services is working with the Association of Primate Veterinarians (APV), the American Association of Zoo Veterinarians, other federal agencies, and industry groups to address these issues. In addition, CDC has developed interim recommendations that update and modify procedures used to identify and control TB in imported NHPs and in workers exposed to such animals.
1. All imported nonhuman primates (NHPs) should be administered a minimum of three tuberculin skin tests (TSTs), with at least 2 weeks between tests (ILAR, 1980) before release from import quarantine. All cohorts containing animals with positive or suspicious reactions should remain in quarantine and be administered at least five additional TSTs following removal of the last affected animal.
2. Records of all TSTs performed during the lifetime of each imported NHP should be maintained at the facility housing the NHP and should accompany the NHP during moves to other facilities.
3. Necropsies of imported NHPs should be performed only by qualified veterinary pathologists or laboratory-animal veterinarians accredited by the American Association for Accreditation of Laboratory Animal Care. Necropsies of tuberculosis (TB)-suspect animals (NHPs with positive or suspicious TST results or NHPs from cohorts within which TB-infected animals have been identified previously) should be performed under animal biosafety level 3 conditions (CDC, 1988). Regardless of gross necropsy findings, fresh and formalin-fixed tissue specimens--to include tracheobronchial lymph node, liver, lung, and spleen--from all such NHPs should be collected for laboratory examination. Granulomatous lesions found in any NHP at necropsy, regardless of whether TB was previously suspected, should be submitted both for laboratory examination for acid-fast bacilli and for mycobacterial culture. Necropsy reports should address all major organ systems and should incorporate clinical history and laboratory findings.
4. All NHPs with positive or suspicious TST results, necropsy findings, or laboratory results should be reported to CDC (telephone: 404-639-8108; FAX: 404-639-2599) within 48 hours, along with a copy or summary of the individual NHP's health records.
5. All facilities that house NHPs should adhere to the Institute for Laboratory Animal Research recommendations regarding baseline and (at a minimum) annual TST screening of employees and routine safe work practices involving NHPs (ILAR, 1980). Results of employee TSTs should be maintained and reviewed by the occupational health professional responsible for the employee health program. Skin-test conversions among employees may suggest transmission of TB in the facility. Workers exposed to NHPs with laboratory-confirmed TB should receive a postexposure TST and, if negative, a TST 3 months after exposure. Workers with a reactive skin test should be referred for medical evaluation.
6. All persons directly involved in the transportation and quarantine of imported NHPs should adhere to the importer's standard operating procedures approved by CDC under the special permit process.
7. In addition to the protective clothing requirements described in previously published guidelines (CDC, 1990b) inspection personnel and other transit workers who handle crates or pallets containing imported NHPs should wear disposable dust/mist respirator masks and be trained in their proper use. Because of the potential for aerosol transmission of certain pathogenic bacteria (e.g., TB) and viruses, face shields or eye protection should be worn by workers whose faces may come within 5 feet of the NHPs.
8. All NHPs should be individually identified with a unique number or alphanumeric code permanently applied to the animal by tattoo. Health certificates, shipping documents, and animal health records should always include this number or code and the age, sex, and species of the NHP (ILAR, 1980).
References
CDC (1988). Biosafety in Microbiological and Biomedical Laboratories. 2nd ed. Atlanta: US Department of Health and Human Services. DHHS publication no. (NIH)88-8395.
CDC (1990a). Requirement for a special permit to import cynomolgus, African green, or rhesus monkeys into the United States. Federal Register, 77, 15210.
CDC (1990b). Update: Ebola-related filovirus infection in nonhuman primates and interim guidelines for handling nonhuman primates during transit and quarantine. Morbidity and Mortality Weekly Report, 39, 22-24, 29-30.
CDC (1991). Update: nonhuman primate importation. Morbidity and Mortality Weekly Report, 40, 684-685, 691.
ILAR (1980). Laboratory Animal Management: Nonhuman Primates (pp. 27-30). Washington, DC: National Academy Press.
ILAR (1985). Guide for the Care and Use of Laboratory Animals. Bethesda, MD: US Department of Health and Human Services. DHHS publication no. (NIH)86-23.
-------------------------------------------------------------------
Reported by the Div. of Quarantine, Nat'l Center for Prevention Services; Scientific Resources Program, Nat'l Center for Infectious Diseases, CDC. From Morbidity and Mortality Weekly Report, (1993), 42, 572-576.
(1) 42 CFR section 71.181 189 (40 FR 33661).
-------------------------------------------------------------------
LPN Associate Editor's Note: This article is reprinted exactly as it appeared in MMWR, but there are some factual errors of which readers should be aware. CDC is aware of them but had not issued any clarifying statements by our press time.
First, and most importantly, in part 3 of the interim guidelines note that AAALAC does not accredit veterinarians. It examines institutional programs, not individuals. Limiting performance of necropsies to those veterinary pathologists and/or clinical veterinarians with appropriate experience is a valid goal, but the published guideline does not necessarily identify those individuals at zoos or other facilities. AAALAC, APV, and the American College of Laboratory Animal Medicine have contacted CDC but have not yet received a response. Second, CDC did not reference the newest edition of their own publication Biosafety in Microbiological and Biomedical Laboratories, 3rd ed., 1993. They referenced the previous edition. Third, ILAR was incorrectly called the Institute for Laboratory Animal "Research" rather than Animal "Resources."
Although the second and third errors are minor, they lead to a document which does little to improve the atmosphere between the zoo and research community and CDC. Given the 26,000 documented human clinical cases of TB in 1992 and the estimated 250,000 skin-test positive humans in the USA, the CDC nonhuman primate "interim guidelines" based on 90 cases over three years is hard to comprehend. This is particularly troublesome since they duplicate many of the existing ILAR guidelines and conflict with pending federal legistlation. Currently, lab animal veterinarians are limited by an antiquated system of TB diagnostics. Duplicative guidelines without more sensitive and spe- cific diagnostic tests will not change our ability to manage the threat of TB. I am disappointed in the document and the lack of interaction that CDC has afforded the regulated community prior to or after the guidelines' publication. -- James Harper.
APV Newsletter Editor's Note (1993, 3 [ 3 ], 2): Does anybody in the regulatory arena really care about tuberculosis diagnosis in nonhuman primates? CDC has shown a passing interest; NIH has made TB a significant disease for investigation, prevention, and diagnosis; USDA has bovine TB under fair control after several intensive decades of work; but the primate is the forgotten animal. An animal that is orphaned in a sea of "no threat to human or agricultural concerns." This is clearly evidenced by our traditional testing method using MOT, by the historical surveys documenting the diagnosis of TB from the 60's and 70's when importation was great and sickness was common in ferals. But we have learned the benefits of conditioning and education for the exporters and importers. We have learned the meaning of quarantine and multiple TB tests. We have come to work with a test system that lacks both sensitivity and specificity. If MOT were submitted today as a diagnostic test for TB would we expect it to clear FDA for efficacy? So does anybody out there really care? And if they do, would they please come forward and involve the APV in finding a solution to the diagnostic nightmare of tuberculosis? Aside from diagnostic methods, how much do we know about the organism itself? Why do we feel there is significant species variability in susceptibility to the agent? Is there really? Why is there variability? Do some species offer better cell-mediated immunity in protecting against the organism? Do some strains of Mycobacterium stimulate such a poor delayed- type hypersensitivity that repeated tuberculin skin tests are negative even when chest lesions are present? Is every strain of TB highly contagious or are there host factors that can reduce transmission? Why do we apparently know so little about tuberculosis, the scourge of every primate colony? Yes, we test for TB repeatedly and often somewhat blindly, asking the same questions and getting the same answers. Our ability to administer the test properly further masks our even more confusing ability to interpret the results meaningfully. Tuberculosis, oh, tuberculosis, will you ever be the same? -- Joe Bielitzki, Yerkes RPRC, Emory University, Atlanta, GA 30322.
* * *
Eve Watts, David Glenn Smith, and Evan L. Zucker
Audubon Park and Zoological Garden, University of California, Davis,
and Loyola University
Adoptions have both theoretical and applied importance. Adoption, described by Thierry and Anderson (1986) as an extreme form of alloparenting, can provide benefits, costs, or some combination to the infants, adoptive parents, and biological parents involved. The distribution of these benefits and costs results in interpretations of adoption based on selfishness, altruism, kin selection, and/or reciprocal altruism (see reviews by Hrdy, 1976, and Thierry & Anderson, 1986).
Reviewing adoption by anthropoid primates, Thierry and Anderson (1986) identified age, sex, kinship, and reproductive status of females as influences on the permanence of adoptions. The dominance relationship of the adults involved also was identified as important if kidnapping were a precursor to adoption. These factors are examined as they apply to an adoption that occurred in a group of red-capped mangabeys at the Audubon Park and Zoological Garden. The conditions of this adoption required DNA fingerprinting to determine maternity, as the successful management of nonhuman primate species in captivity requires accurate knowledge of parentage. The characteristics of the present case are compared to other reports of adoption in this subspecies.
Methods
Subjects and Habitat : A red-capped mangabey (Cercocebus torquatus torquatus) group was formed in July 1989 at the Audubon Park and Zoological Garden, New Orleans, LA. At the time of formation, the group consisted of a multiparous, nine-year-old female (Delilah), her year-old immature male offspring, a nulliparous, five-year-old female (Charlotte), and a 21-year-old adult male (Ivan).
The group was housed in a semi-natural habitat approximately 10 x 5 x 4 m. The exhibit has a grass and dirt terrestrial substrate, numerous branches and ropes for arboreal locomotion and sitting, and rocks for sitting. Indoor areas for the mangabeys are behind the exhibit.
Observations : The mangabeys were observed daily by zoo personnel. In addition, as part of a study on the social interactions of the adult mangabeys, the group was observed on a near-daily basis between March and early May 1990, by a team of student observers.
Chronology of the Adoption
Charlotte gave birth to her first infant on March 7, 1990, and showed adequate maternal behavior. Delilah gave birth on March 15, 1990, and also showed adequate maternal behavior. Later examination revealed one infant (Nikita) to be male and the other (Sasha), female.
On the morning of March 16, Charlotte was carrying both infants. While the other, more dominant female (Delilah) showed interest, she made no attempt to retrieve her infant. Over the next weeks, Charlotte continued to suckle and ventrally carry both infants, and engaged in other maternal behaviors such as sitting with, retrieving, and grooming both infants. As locomotor abilities of the infants developed, both ventured away from Charlotte, but also returned to her. When the infants were away from Charlotte, Delilah made no attempt to retrieve Nikita.
In May, 1990, when the infants were two months old, Nikita began approaching Delilah. She carried him intermittently over a two-week period, then regularly thereafter. Delilah had continued to lactate as her previous offspring frequently was allowed to nurse. Once Delilah regularly carried and nursed Nikita, Charlotte was not seen attempting to retrieve him.
Assessment of Maternity
As the difference in the infants' sexes was not known at the time of the adoption, it was not known whether Nikita had returned to, or left, his biological mother after the two-month period. To assess maternity, blood samples were obtained from all adults and the two infants in December 1991, and DNA fingerprinting was done using methods described by Smith et al. (1992).
Digests of DNA were prepared using three different restriction enzymes: Hinf I, Alu I, and Hae III. All fragments exhibited by either Sasha or Nikita were also exhibited by either Ivan (their father), or by at least one of the two adult females. In no instance did Nikita exhibit a fragment absent in Ivan that was not also exhibited by Delilah, and in no instance did Sasha exhibit a fragment absent in Ivan that was not also exhibited by Charlotte. Moreover, in all instances, such fragments were exhibited by only one of the two adult females; Charlotte could not be the mother of Nikita, and Delilah could not be the mother of Sasha. Thus, the male infant Nikita returned to his biological mother.(1)
Discussion
In describing the typical anthropoid primate adoption, Thierry and Anderson (1986) reported that neonates were more likely to be adopted by adult females, especially lactating females, whereas older infants were more likely to be adopted by juveniles and adult males (see also Capitanio & Taub, 1992). In this mangabey group, and consistent with this typical adoption pattern, the female who adopted the neonate was lactating, although lactation was in conjunction with a viable birth rather than from a reproductive loss (neonatal death or stillbirth).
Thierry and Anderson (1986) also found that adoptions were likely to occur among related individuals, particularly within matrilines. However, in the present case, the adult females were unrelated. The present case also differed from the typical case in that a subordinate female adopted the offspring of a more-dominant female. As Delilah, the dominant female, did not attempt to regain possession of her infant, it is only speculative to assert that she could have regained the infant had she tried.
There are both similarities and differences between the adoption described here and red-capped mangabey adoptions (and alloparental behaviors) that have occurred at other zoos. At the Houston Zoo, a subordinate female has been observed carrying the infant of the alpha female (Lester, personal communication), a situation similar to the adoption reported here. In contrast, at the Los Angeles Zoo, the adopting mother was the older of two females, apparently multiparous, and adopted a newborn the day her own infant died (Bingham, 1990). In our case at the Audubon Zoo, the adopting mother was the younger of the females, primiparous, and raised both infants.
This case of adoption of a neonate by a newly-parturient, primiparous red-capped mangabey can be examined with respect to possible changes in the individual fitnesses of the individuals involved. Whereas long-term changes in the fitness of the adult females cannot be assessed, in the short-term, neither Delilah, her year-old offspring, nor her infant appeared to incur any losses from having Charlotte successfully raise the infant to two months of age. In addition, Charlotte's own infant did not appear to be at any disadvantage from having an adopted sibling, as she appeared to develop normally. In other cases of two infants raised by one female, one infant often fails to survive due to malnourishment (Thierry & Anderson, 1986), but both of these infants survived, indicating sufficient nourishment and care. Charlotte gave birth to a second infant on August 6, 1991, an interbirth interval of 17 months, suggesting that she did not incur any immediate costs from additional lactation. Delilah did not conceive during this period of time, so Charlotte did not lose fitness relative to Delilah. [See Thierry & Anderson (1986) for the theoretical importance of benefits and costs of adoptions.]
These cases of short- and long-term adoptions in three different social groups indicate that red-capped mangabeys might be suitable for the study of alloparenting in captivity and, perhaps, in the wild. The apparent tendency to provide alloparental care also has implications for the captive management of this subspecies. First, adoptions can occur regardless of parity or relatedness of females. Second, if adoption occurs, the infant can be raised successfully by a lactating female, even if she has her own infant. Third, females should not be expected to regain their adopted infants. Fourth, if a newly parturient female fails to show adequate maternal care, a manipulated or induced adoption might succeed, avoiding human-rearing of captive-born infants. More generally, if adoptions occur in social contexts where there are numerous infants and/or possible mothers, DNA fingerprinting, most commonly used to assess paternity (see Martin et al., 1992), can be used to assess maternity.
References
Bingham, V. (1990). Births and hatchings...Los Angeles Zoo. Animal Keepers' Forum, 17, 190.
Capitanio, J. P. & Taub, D. M. (1992). High-level care of a twin infant by a male rhesus macaque. Folia Primatologica, 58, 103-106.
Hrdy, S. B. (1976). Care and exploitation of nonhuman primate infants by conspecifics other than the mother. Advances in the Study of Behavior, 6, 101-158.
Martin, R. D., Dixon, A. F., & Wickings, E. J. (Eds.) (1992). Paternity in Primates: Genetic Tests and Theories. Basel: Karger.
Smith, D. G., Rolfs, B., & Lorenz, J. (1992). A comparison of the success of electrophoretic methods and DNA fingerprinting for paternity testing in captive groups of rhesus macaques. In R. D. Martin, A. F. Dixon, & E. J. Wickings (Eds.), Paternity in Primates: Genetic Tests and Theories (pp. 32-52). Basel: Karger.
Thierry, B. & Anderson, J. R. (1986). Adoption in anthropoid primates. International Journal of Primatology, 7, 191-216.
-------------------------------------------------------------------
Third author's address: Dept. of Psychology, Loyola Univ., New Orleans, LA 70118.
We thank C. Kennedy, M. Fernandez, D. McCarthy, K. Manceaux, and G. Ranglack (all of the Audubon Zoo), B. Lester (Houston Zoo), J. McNary (Los Angeles Zoo), L. Franklin and S. Bastow (Loyola Univ.), and M. R. Clarke (Tulane RPRC) for their assistance in various ways. The Audubon Park and Zoological Garden is fully accredited by AAZPA.
(1) A copy of the DNA fingerprint is available upon request.
* * *
D. R. Rasmussen
Animal Behavior Research Institute and Wisconsin Regional Primate
Research Center
Introduction
Matrilineal associations are major strands in the social fabric of natural and semi-natural groups of macaques (Sackett & Fredrickson, 1987; Crooks & Rasmussen, 1991; Rasmussen et al., in press). Familiarity between offspring and their mothers and between siblings across generations may be a primary proximate mechanism for these associations. Phenotype matching also could be a mechanism for kinship recognition (Fredrickson & Sackett, 1984). Phenotype matching is the recognition of the degree to which another individual has similar characteristics such as coat color, skin color, and behavioral interaction patterns (Holmes & Sherman, 1983).
Pigtail macaques (Macaca nemestrina) are more attracted to familiar but unrelated individuals than to unfamiliar kin (Fredrickson & Sackett, 1984; Sackett & Fredrickson, 1987). Although an earlier study showed a significant preference for unfamiliar kin over unfamiliar nonkin in infants (Wu, Holmes, Medina & Sackett, 1980), Sackett & Fredickson (1987) concluded familiarity is the likely proximate mechanism for visual kin recognition and preference.
Familiarity has recently been shown to be significantly associated with patterns of affiliation in two groups of rhesus Macaca mulatta (Rasmussen, 1993). Here the relative strength of the relationships between relatedness and affiliation and familiarity and affiliation are compared for the individuals in these groups. Partial correlation is used to help determine if relatedness has an influence on affiliation independent from the influence of familiarity.
Methods
These are the identical subjects and data on which analyses of the influence of previous familiarity on affiliation were reported (Rasmussen, 1993). The methods section is therefore abbreviated. Further details are provided in the initial article.
Housing, Group Composition, and Subjects
The groups were housed in identical indoor pens. The experimental group was composed of a 5-year-old vasectomized male, a 4-year-old vasectomized male, an intact 2-year-old male, four 4-year-old females, one 3-year-old female and three 2-year-old females. All males were intact in the sex and age matched control group.
Variable Definitions and Sampling Methods
Data were collected from January 2, 1987 to June 1, 1987, the day before the birth of the first infant in the control group. The analyses are based on 163 hours of 2 min-interval samples.
Relatedness: Sewall Wright's coefficient of relatedness was calculated on the basis of known maternity and paternity. Subjects were assumed to have unrelated male ancestors when paternity was unknown. Only 10 of the 110 possible dyads in the two groups were related. Degree of relatedness varied from .03125 to .2500. Six of the related dyads were heterosexual dyads, three were female-female dyads and one was a male-male dyad (Table 1).
+---------------------------------------------------+----+
| Degree of Relatedness |Sums|
| +----------------+---+------+-----+-----+----+----+
|No. | | 0 |.03125|.0625|.1250|.25 | |
| +----------------+---+------+-----+-----+----+----+
|of |Isosexual 0 |116| 4 | 2 | 2 | 0 |124 |
| +----------------+---+------+-----+-----+----+----+
|dyads |Heterosexual 1 |84 | 2 | 4 | 2 | 4 | 96 |
| +----------------+---+------+-----+-----+----+----+
| |Sums of Columns |200| 6 | 6 | 4 | 4 |220 |
+------+----------------+---+------+-----+-----+----+----+
Table 1: Number of isosexual and heterosexual dyads by their degree of relatedness. There was a slight positive correlation ( r = .16, n = 220) between heterosexual dyads and relatedness: heterosexual dyads therefore had a faint tendency to be more closely related than isosexual dyads.
Previous familiarity was assessed with days individuals spent
together from birth until data collection was initiated. Relatedness
and familiarity were correlated (r =.53).
Neighbor distances: Nearest neighbor distance (NN) was the mean
distance between a given dyad across all samples. Close neighbor distance (CN) was the mean distance between a dyad during those intervals
when members of the dyad were nearest neighbors.
ALF: Approach, leaves and follows combined in a composite variable, ALF.
Grooming was defined as picking through the hair or skin of
another with fingers or teeth.
Grooming Presents: An "A => B" grooming present was defined to
occur when A approached B and exposed a body part at 1/3 m or less of
B's hands or mouth.
Presents: An "A => B" present was scored when A oriented its
ano-genital region towards the face of B when B was within 2 m of A.
Mounts: An "A => B" mount was scored when A climbed on top of B,
placed both hands on B's back and anogenital regions of A and B were
aligned as during copulation.
Descriptive Statistical Analyses
NN and CN distances were calculated for nondirectional dyads since the distance of A -> B is necessarily the same as the distance of B -> A. Rates of directional dyadic interactions were calculated for all the other variables.
The variables were regressed on relatedness and familiarity and the partial correlation calculated between the variables and relatedness with familiarity controlled (Table 2). The transformations used to meet the assumption that the residuals from regression are normally distributed are listed in Table 1 of Rasmussen (1993).
+-----------+--------+-----+----+--------+-------+----+---------+ |Affiliative| r with | r |mean|# of +,-|partial|mean|# of +,- | |Variables |familiar| kin | r | rank r | r kin | r | rank r | +-----------+--------+-----+----+--------+-------+----+---------+ |NN | -.31 @ | -.22|-.26|3, 10 | -.07 |-.08| 9, 10 | |CN | -.39 @ | -.16|-.23|2, 11 * |-.09 |-.00| 10, 9 | |ALF | +.24 * | +.22|+.31|12, 1 @ |+.12 |+.15| 14, 5 | |Grooming | +.27 @ | +.28|+.31|11, 2 * |+.24 |+.26| 17, 2 & | |Groom- | | | | | | | | | Presents | +.14 | +.08|+.22| 11, 2 *|+.05 |+.36| 18,1 & �| |Presents | +.22 | +.04|-.10| 2, 11 *|-.01 |+.32| 17, 2 & | |Mounts | +.22 |+.003|+.25|11, 2 * | -.046 |+.49| 18, 1 & �| +-----------+--------+-----+----+--------+-------+----+---------+
Table 2: The relationships between the affiliative variables, familiarity and relatedness and the significance of those relationships.
Inferential Statistical Analyses
The significance of the relationships was tested using the following method (Rasmussen, 1993): All the dyads involving a given individual were selected. The Spearman rank order correlation was then calculated between interactions in these dyads and their relatedness. This procedure was repeated for all group members. A two-tailed binomial test was used to determine if more of the correlation coeffi- cients were positive than expected by chance.
This procedure was conducted on the residuals from regression of the variables on familiarity to test the partial correlations. The numbers of rank order correlations of each sign are listed in Table 2. Table 2 also lists the mean of the rank order correlations. The correlations between familiarity and affiliation previously reported (Rasmussen, 1993) are also listed in Table 2.
Sex Composition of Dyads:
Previous comparisons of the influences of familiarity and relatedness on affiliation have not examined these influences as a function of the sex of the individuals engaged in affiliative interactions. We therefore conducted a second set of analyses in which we controled for the sex of individuals within dyads. Dyads were coded as either isosexual (male-male or female-female) or heterosexual. We then examined the partial correlations between relatedness and affiliation with familiarity and sex controlled. The significance of these partial correlations was again tested with the previously described binomial test.
Results
Except for grooming all correlations between the affiliative variables and relatedness were of lower magnitude than those between the affiliative variables and familiarity. The partial correlations for all variables except grooming were of lower magnitude than the zero order ones.
For all but one of the affiliative variables the relationships indicated by the correlations with relatedness were significant. Only four of the affiliative variables had significant relationships with familiarity. These results indicate the greater consistency in the direction of the relationships with relatedness within individuals than in the relationships with familiarity. Presents were significantly correlated with relatedness but this was due to a greater number of correlations than expected having a negative sign. There was less presenting between individuals who were more closely related. Four of the partial correlations were also significant: grooming, groom presents, presents, and mounts. The partial correlations were very weak, yet the means of the rank order correlations were substantial.
+-----------+-----------+---------+-----------+-------------+
|Affiliative|Familiarity|Sex Comp.|Relatedness| Relatedness |
| Variables | | of Dyads| | +/- P value |
+-----------+-----------+---------+-----------+-------------+
| NN | -.24 | +.16 | -.09 | 12/10 .83 |
| CN | -.39 | +.16 | -.11 | 9/13 .52 |
| ALF | +.15 | -.10 | +.13 | 14/ 8 .29 |
| Grooming | +.25 | -.18 | +.26 | 15/ 7 .13 |
| Groom- | | | | |
| Presents| +.14 | -.07 | +.05 | 16/ 7 .0525 |
| Presents | +.22 | -.02 | -.01 | 19/ 3 .001* |
| Mounts | +.22 | +.01 | -.05 | 21/ 1 .0001*|
+-----------+-----------+---------+-----------+-------------+Table 3 : Partial correlations between measures of affiliation, familiarity, sex composition of dyads and relatedness.
Heterosexual dyads had a slightly higher likelihood of being related than isosexual dyads (Table 1). The control of the sex composition of dyads had only a faint influence on the magnitude of the partial correlations between the measures of affiliation and relatedness (Table 3). Only two of the tests of the partial correlations remained significant: presents and mounts were significantly correlated with relatedness after control of both previous familiarity and sex composition of dyads. Control of the sex composition of dyads did lead to significantly more of the rank order correlations between presents and relatedness being positive in sign.
Discussion
The results show that relatedness significantly predicts many aspects of rhesus affiliation. However, for all patterns of affiliation except grooming, previous familiarity was more strongly correlated with affiliation than relatedness. When familiarity is controlled for, most of the relationships between relatedness and measures of affiliation are very weak. Familiarity therefore had a stronger influence than kinship on affiliation between the rhesus in the two groups.
These results are consistent with two experimental research studies on pigtail macaques (Fredrickson & Sackett, 1984; Sackett & Fredrickson, 1987) which showed familiarity was the proximate cause of social preference. Thus the literature and the results reported here suggest that familiarity may be the primary proximate mechanism responsible for the strong matrilineal association patterns observed in natural and semi-natural groups of macaques (Chepko-Sade & Sade, 1979; Sackett & Fredrickson, 1987; Crooks & Rasmussen, 1991).
Our results suggest that relatedness was at least as important as familiarity in its influence on grooming. Statistical control of familiarity had only a negligible influence on the relationship between relatedness and grooming. In addition, four of the partial correlations, although weak, were significant. Only experimental studies can prove causation. The results do, however, suggest that a mechanism other than familiarity may contribute to kin recognition in rhesus.
The two experimental studies conducted on pigtail macaques (Fredrickson & Sackett, 1984; Sackett & Fredrickson, 1987) did not exclude mechanisms other than familiarity for kin recognition. Both studies used primarily like-sexed subjects and stimulus monkeys to assess social preference. A previous study in which subjects and stimulus monkeys were predominantly of different sexes did show a preference for kin (Wu, Holmes, Medina & Sackett, 1980).
In this study a majority of the related dyads also were heterosexual. In addition all four of the partial correlations that were significant were for affiliative variables that may be strongly associated with reproduction: grooming, grooming presents, presents and mounts (Rasmussen, 1984; Zumpe & Michael, 1970). Control of the sex composition did, however, result in a reversal of the direction of the majority of rank order correlations between presents and relatedness. Sex composition of dyads can therefore exert a strong influence on the association between relatedness and at least some patterns of affiliation.
Future studies might therefore focus on heterosexual interactions between individuals with a greater variation in age differences. Experimental studies done so far have investigated only a narrow range of affiliative interactions. A more diverse set of variables should be assessed, as in this study. Finally, the strengths of the influences of familiarity and relatedness should be compared for behavioral patterns other than affiliation, i.e., sexual interactions and alliance formation (Aureli et al., 1992).
References
Aureli, F., Cozzolino, R., Cordischi, C., & Scucchi, S. (1992). Kin-oriented redirection among Japanese macaques: An expression of a revenge system? Animal Behaviour, 44, 283-291.
Chepko-Sade, B. D. & Sade, D. S. (1979). Patterns of group splitting within matrilineal kinship groups. Behavioral Ecology and Sociobiology, 5, 67-86.
Crooks, K. R. & Rasmussen, D. R. (1991). The relationship between kinship and grooming in the Tanaxpillo colony of stumptail macaques (Macaca arctoides). Laboratory Primate Newsletter, 30[4], 1-3.
Fredrickson, W. T. & Sackett, G. P. (1984). Kin preferences in primates (Macaca nemestrina): relatedness or familiarity? Journal of Comparative Psychology, 98, 29-34.
Holmes, W. G. & Sherman, P. W. (1983). Kin recognition in animals. American Scientist, 71,46-55.
Rasmussen, D. R. (1984). Functional alterations in the social organization of bonnet macaques (Macaca radiata) induced by ovariectomy: An experimental analysis. Psychoneuroendocrinology, 9, 343-374.
Rasmussen, D. R. (1993). Prediction of affiliation and sexual behavior in rhesus monkeys with previous familiarity. Laboratory Primate Newsletter, 32[2], 12-14.
Rasmussen, D. R., Riordan, E., Farrington, M., Kelly, E., Nachman, J., Fernandez, S., & Churchill, A. (in press). The central-peripheral structure of the Tanaxpillo colony of stumptail macaques (Macaca arctoides). In: R. A. Gardner, A. B. Chiarelli, B. T. Gardner, & F. X. Plooij (Eds.), The Ethological Roots of Human Culture. The Netherlands: Kluwer Academic Publishers.
Sackett, G. P. & Fredrickson, W. T. (1987). Social preferences by pigtailed macaques: familiarity versus degree and type of kinship. Animal Behaviour 35, 603-606.
Wu, H. M. H., Holmes, W. G., Medina, S. R., & Sackett, G. P. (1980). Kin preference in infant Macaca nemestrina. Nature, 285, 225-227.
Zumpe, D., & Michael, R. P. (1970a). Ovarian hormones and female sexual invitations in captive rhesus monkeys (Macaca mulatta). Animal Behaviour, 18, 293-301.
-------------------------------------------------------------------
Author's address: Animal Behavior Research Institute, 314 South Randall Street, Madison, WI 53715, USA.
Dr. V. Reinhardt vasectomized the males and helped select subjects. Dr. S. Sholl and P. DuBois provided programming assistance. S. Eiseley and P. DuBois helped with kinship records. Support was provided by a NIMH National Research Senior Service Award, 1 F32 MH09419-01 RERA and NIH. This is publication number 28 of the Animal Behavior Research Institute.
* * *
Jean Baulu
Barbados Primate Research Center and Wildlife Reserve
This letter is a response to W. Robin Kingston's article "Time to Ban Imports? A European Perspective" in the July 1993 issue of this Newsletter, in which he suggests that commercial trapping in supplier countries be phased out and breeding colonies be established in those countries.
For decades, rich countries have depleted poor countries of one of their renewable natural resources, primates of biomedical importance. In 1977, India put a total ban on the export of rhesus monkeys and, in recent years, both Indonesia and the Philippines have enacted quotas on the capturing of live cynomolgus monkeys for export. On the other hand, the latter countries, as well as China, have also begun large-scale breeding programs to satisfy the needs of the world's biomedical industry. In China, Macaca mulatta are bred on a very large scale and at a cost that is much lower than that of breeding them at the U.S. Regional Primate Centers.
A factor that needs to be taken into consideration is that there
are now several places (mainly islands in the Caribbean) in which primates have been introduced (in some cases over 350 years ago) and have
become serious agricultural pests. These are:
* St. Kitts and Nevis,
where Cercopithecus aethiops sabaeus is an agricultural pest and is
culled whenever there is a demand. Even though there are research
facilities on the island of St. Kitts, there is little or no direct or
even indirect benefit to farmers or the local community from opportunistic culling.
* Barbados, where a very successful Monkey Crop Damage
Control Programme has been on-going for over 12 years with the help of
farmers. The program has involved self-imposed capture quotas; the
development of the Barbados Primate Research Center; a long-term
breeding facility for C. aethiops sabaeus; the Barbados Wildlife
Reserve, one of the region's premier tourist attractions; and the sale
of high-quality monkeys, which are either wild-caught or captive-bred,
at reasonable prices (US $600 to $1600).
* Trinidad, where there are
red howlers and capuchins, considered native since the island is only
six miles from Venezuela. These two species are not in demand, and
therefore are not trapped in significant numbers.
* Grenada, where
C. mona was introduced from Africa. The monkey population is low and
is at present of no biomedical importance. There is some hunting of
the monkeys for food.
* Puerto Rico, where there is a 35-acre off-coast island, as well as a land-based captive-breeding program of
about 2000 Macaca mulatta. The rhesus were imported over 50 years
ago from India. Some escaped (along with patas monkeys) during the
late 1970's and have since become agricultural pests in part of the
island. These wild and purpose-bred colonies are ideal for the supply
of rhesus to user countries as they are priced between US $500 and
$1500 at present.
* Mauritius, where there is an estimated population
of 40,000 introduced wild cynomolgus monkeys as well as some captive
breeding. The monkeys seriously threaten some endemic species of birds.
I understand that the largest purchaser of wild-caught monkeys (Charles
River) is now exporting large numbers of these animals to set up permanent breeding facilities in Texas, rather than helping the local
economy by breeding them on Mauritius. These monkeys, like the green
monkeys in Barbados, are SAIDS, B-virus, filovirus, etc, negative,
which makes them ideal primate models for biomedical research as well
as for vaccine production and testing.
I favor an approach in which the capturing of wild monkeys would continue on a sustainable basis, especially where they have been introduced and become agricultural pests. Breeding centers should also be established in countries of origin, perhaps with the implementation of Dr. Trevor B. Poole's proposal that some kind of accreditation be given to such facilities outside of Europe so that basic standards of husbandry and veterinary care are maintained. This should act as an incentive to suppliers to upgrade their facilities and take control of trapping so that it is done in a humane fashion.
The idea of breeding primates in Europe in order to become self-sufficient for the needs of EEC member countries sounds like a commendable goal. But is it necessary? Surely in countries that have developed sustained-yield culling practices, self-imposed quotas, and captive breeding programs, as well as parks, environmentally sound conservation, and ecological practices, one should encourage these efforts, especially since these countries are often in dire need of job creation and foreign exchange. Also, under proper management, indigenous or even introduced primates are a natural renewable resource, and certainly much cheaper than if bred in user countries. It is also important that developed countries keep good relations with developing nations--especially those harboring primates. Today there are only 13 species of biomedical interest. In the future, we may find that some of the more than 200 other species of primates hold the key to new drugs or treatments.
Finally, I am also of the opinion that certain pharmaceutical companies could very well carry out drug testing, vaccine production, etc, in countries where the resources (monkeys) are available. There are over 90 international pharmaceutical companies in Puerto Rico alone, producing over 75% of all medicines and related health products sold in the continental USA. Their offices and manufacturing plants there enjoy long-term tax write-offs, and also create much-needed employment on the island. There is much room for improving the nature and quality of the demand for nonhuman primates from traditional (and nontraditional) supplier countries.
-------------------------------------------------------------------
Author's address: Barbados Primate Research Center and Wildlife Reserve, Farley Hill, St. Peter, Barbados, West Indies.
* * *
Dr. J. Baulu's response to my article on banning imports of wild-caught primates raises a number of other considerations, particularly in regard to feral, introduced populations in the Caribbean. Certainly the culling of those primates which have become agricultural pests, along with the currency generated by their sale, is an attractive idea. However, they are still wild-caught animals, subject to the stress of capture, and of unknown age, health, and relationships, as are those from their original homeland. I am sure that the methods of trapping, screening, and handling used in Mr. Baulu's operation in Barbados are of the highest standard, but this may not be the case in other areas. Complaints have been heard about the operation in Mauritius.
Two points about captive breeding that are often overlooked are, first, that the stress of capture and handling is very much less in animals accustomed to the proximity of human beings and, second, that there need be little or no change in diet between the breeding center and the laboratory. In addition, the ages, relationships, and veterinary status are all known and properly managed, so there should be no need for culling. The foundation stock of the breeding colony can be rigorously screened for disease and every effort made to exclude potential vectors.
I am in entire agreement with Mr. Baulu that the primates of a country are one of its renewable natural resources and, following that, as much research as possible should be carried out in the countries of origin of the primates. Ideally this should be a national effort with, if necessary, technical assistance from user countries. If the initiative for this is lacking, expatriate agencies should undertake the breeding, employing as much local labor as possible. -- W. Robin Kingston, The Old Smithy, Bishops Frome, Worcester WR6 5BA, U.K.
* * *
An updated Directory will be published in the January, 1994, issue of the Laboratory Primate Newsletter. If you wish your program to be represented in this Directory or to revise your present entry, please send us the necessary information, following the format shown here as closely as possible. Return the information as soon as possible, but not later than December 10, 1993, to the Laboratory Primate Newsletter, Psychology Department, Box 1853, Brown University, Providence, RI 02912 [[email protected] or [email protected]]. Please note that the Directory is not intended for post-doctoral programs, though any such sent to us will be listed separately.
For examples, see the 1992 Directory in the LPN, 1992, 31[1], 25-32.
1. State:
2. Institution:
3. Division, Section, or Department:
4. Program Name and/or Description:
5. Faculty and Their Specialties:
6. Address for Further Information:
* * *
Malaria Reminder
Malaria was diagnosed in 48 U.S. military personnel who were deployed to Somalia from late December 1992 through April 1993. In addition, through late June, malaria was diagnosed in 83 military personnel following their return from Somalia. This substantial number of cases has reinforced concerns regarding malaria prophylaxis, the estimated risk of infection, and the need for prompt recognition and treatment of malaria.
Symptoms included a history of fever and chills (100%), headache (97%), gastrointestinal symptoms (72%), myalgia and/or arthralgia (69%), lumbosacral pain (63%), and upper respiratory symptoms (59%). Malaria must be considered in the differential diagnosis for all persons with fever or a history of fever who have traveled to a malarious area. Initial symptoms may not include the classic pattern of repeated episodes of fever and chills and may have a dominant gastrointestinal or respiratory component. For patients who have contin- ued taking prophylaxis or who have recently discontinued prophylaxis, the clinical presentation often is milder than in patients who have not taken any prophylaxis. Malaria infection can be excluded only after microscopic examination of serial thick and thin blood smears over a 72-hour period. From Morbidity and Mortality Weekly Report, 1993, 42, 524-526.
Cholera in Africa
In 1991, 21 African countries reported 153,367 cholera cases and 13,998 cholera-related deaths to WHO. In 1992-93, outbreaks were reported in Burundi and Zimbabwe. Waterborne transmission of cholera in Africa has been associated with drinking untreated water from rivers and shallow wells. Boiling and chlorination are effective treatments. Cholera also has been transmitted by the foodborne route in Africa. Because Vibrio cholerae O1 is rapidly inactivated at acid pH levels, recipes that acidify food can be protective. Transmission has been documented through consumption of contaminated moist cooked grains that were held for several hours before eating, and through contaminated shellfish. From Morbidity and Mortality Weekly Report, 1993, 42, 407-409, 415-416.
Schistosomiasis in Malawi, 1992
Schistosomiasis ("snail fever" or "bilharzia"), a parasitic infection caused by trematodes (flukes), is endemic in 74 countries in Africa, South America, the Caribbean, and Asia. Workers and travelers in those countries may be at risk for schistosomiasis. During 1992, two U.S. Peace Corps volunteers were evacuated from Africa because of Schistosoma hematobium infection of the central nervous system (CNS). Both were exposed to fresh water while vacationing at Cape Maclear, a resort area on Lake Malawi.
CDC recommends that expatriates and travelers with a history of freshwater exposure in areas with endemic schistosomiasis be serologically screened and, if seropositive, receive treatment with praziquantel following thorough stool and urine examinations for schistosome eggs.
Persons traveling in Africa should be advised of the risk of schistosomiasis associated with freshwater lakes, streams, and rivers throughout the continent. The only completely effective method of prevention is avoiding contact with fresh water in areas with endemic disease. If contact with fresh water is unavoidable, water should be heated to 122~ F (50~ C) for 5 minutes, treated with iodine or chlorine, strained with paper filters, or allowed to stand for 3 days.
Physicians who treat travelers, expatriates, and immigrants should consider the possibility of neuroschistosomiasis in all patients who have a history of freshwater exposure in schistosomiasis-endemic areas and CNS abnormalities, even in the absence of classic symptoms of acute schistosomiasis (e.g., fever, nausea, vomiting, abdominal pain, diarrhea, and hematuria). Neuroschistosomiasis can occur several months after exposure to infested water and in low-intensity infections in which eggs may be undetectable or difficult to identify. Sensitive and specific serologic tests for diagnosing schistosomiasis are available through CDC's Parasitic Diseases Branch, Nat. Center for Infectious Diseases [404-488-4050].
Treatment with a single dose of praziquantel (40-60 mg per kg body weight) is safe and effective therapy against the adult worms of the three major species of schistosomes infecting humans. Corticosteroids are often useful to reduce edema and inflammation. Although CNS schistosomiasis is rare, substantial morbidity from this condition is preventable by early diagnosis and rapid treatment. -- From Morbidity and Mortality Weekly Report, (1993) , 42, 565-570.
* * *
The paradox faced by researchers working with living nonhuman animals, especially nonhuman primates, is that we must see them as sufficiently similar to us to provide interesting answers to compelling issues; yet, we must also regard them as sufficiently different so that we can use them in experiments that we would consider morally or ethically objectionable with human subjects. With this statement Gareth Matthews, a philosopher from the University of Massachusetts in Amherst, opened a special working conference on Ethics in Primatology held August 17-18, 1993 in conjunction with the 16th annual meeting of the American Society of Primatologists.
The conference was organized by Andrew Petto (New England Regional Primate Research Center) and Karla Russell (New Mexico Regional Primate Research Laboratory). Its participants came from the biological and behavioral sciences and from philosophy, history, resource management, and education. Nearly 25% of the attendees were students. The main objective of the conference was to explore the nature of the human-nonhuman relationship from which Matthews' paradox is drawn. The participants considered 1) what specific research results in their own fields could contribute to a better understanding of this relationship and its ethical implications; and 2) what future studies they could propose that would help to clarify remaining questions.
Presentations by invited speakers served as themes for problem-oriented discussion sessions that followed. Matthews explained different philosophical paradigms for exploring the status of nonhuman animals. Elizabeth Adams of Clemson University demonstrated how the language used to characterize nonhuman primate behavior both reflect and shape our concepts of primate societies. Arnold Arluke of North- eastern University described his recent research on researchers' approaches to animals as reflected in the concerns, acculturation, and practical activities of staff in animal laboratories.
Andrew Rowan of Tufts University School of Veterinary Medicine spoke of the application of various ethical constructs in the context of Darwinian models of the biological relationships among taxa. He also called for more scholarship from and collaboration among interested parties in all the fields represented at the conference on ethical issues that confront professional and student primatologists. Finally, Strachan Donnelley, an ethicist from the Hastings Center, discussed "moral ecology" as a metaphor for making ethical decisions about animal use in research. He proposed a multifactorial approach in contrast to a number of competing ethical constructs that are based on a single characteristic shared by humans and nonhumans, such as "sentience" or self-awareness.
Each of these presentations led to vigorous and wide-ranging discussions. For example, after the presentation on language partici- pants explored editorial policy, classroom teaching, undergraduate and graduate mentorships and internships, and taxonomic classifications. Discussions after the presentation on Darwinian models examined the implications of taxonomic categories for shaping our expectations of human-nonhuman relations and the implications of mosaic patterns of similarities and differences among more closely related taxa. The presentation on the culture of animal research stimulated discussions on how the names that we give other animals shape attitudes and expectations of their relationships with and behaviors toward us.
At the end of the conference each of the participants agreed to join in efforts during the coming year to develop these discussions further. These efforts will have two main goals. The first is to engage the membership of the American Society of Primatologists in a more formal and more official inclusion of ethical issues in their annual meeting program. The second is to follow these preliminary discussions with scholarly collaborations that result in a literature base accessible to and appropriate for practicing primatologists and primatologists-in-training. -- Report by Andrew Petto and Karla Russell.
-------------------------------------------------------------------
Acknowledgement: The realization of this conference was underwritten by a major gift from Judith Schrier in memory of her late husband, Allan Schrier, and their parents, Jean Schrier, Bernard Sanow, and Ruth Sanow. Partial support was also provided from the Edna Tompkins Trust and NIH Grant RR00168 to NERPRC.
* * *
Conan Liu, Celtrix Pharmaceuticals, 3055 Patrick Henry Dr., Santa Clara, CA 95054
Preston A. Marx, 1701 10th St, Dept B-8, Alamogordo, NM 88352-5000.
Andrew J. Petto, Center for Biology Education, Univ. of Wisconsin, 660 WARF Bldg, 610 Walnut St., Madison, WI 53705.
Douglas A. Powell, 5 Seville Way, Gaithersburg, MD 20878.
Donna A. Steiner, 4101 NW Alpine Dr., Gainesville, FL 32605.
Creighton J. Trahan, Commander, USAMRICD, Attn: SGRD-UV-V, LTC Trahan, APG, MD 21010-5425.
* * *
Tissue for Odorant Binding Protein Research
Francesco Scalfari is seeking donations of nonhuman primate material for research on odorant binding proteins and related binding proteins (identification, purification, and characterization) with biochemical techniques (electrophoresis, HPLC, amino acid sequencing, molecular cloning). Materials needed are: olfactory/respiratory nasal mucosa, nasal septum, turbinates, vomeronasal organ, tongue with salivary glands, eyes with lacrimal glands, mammary glands, saliva, nasal mucus, lacrimal fluid, urine, vaginal fluid, seminal fluid, milk, and blood. Please contact Francesco Scalfari, Inst. of Anthropology, Univ. of Florence, Via del Proconsolo 12, 50122 Florence, Italy [55-2398065].
Resources for Conducting Research on Aging
The National Institute on Aging (NIA), recognizing that most investigators have neither the facilities nor the fiscal resources needed to develop and maintain colonies of aged animals, has made provision of resources one of its highest priorities. The availability of high quality material for conducting research on aging continues to be a priority area for NIA, with current resources spanning a broad spectrum, including a cell bank, a nematode bank, rodent colonies, and nonhuman primate colonies.
The NIA maintains approximately 300 Macaca mulatta at four regional primate centers for conducting research on aging. These animals, aged approximately 18 to 35 years, are available for both noninvasive and invasive research studies. Some animals are maintained in group housing while others are individually caged. For information contact the Office of Biological Resource Development, National Institute on Aging, Gateway Building, Suite 2C231, Bethesda, MD 20892 [301-496-0181]. -- From the NIH GUIDE, July 23, 1993, 22 [26].
* * *
Human Frontier Science Program
The Human Frontier Science Program aims to promote, through international collaboration, basic research to elucidate the complex mechanisms of living organisms. Applications are invited for the support of research grants, fellowships, and workshops in these areas: (A) Basic research for the elucidation of brain functions: 1. Elementary Processes. 2. Perception and Cognition. 3. Movement and Behavior. 4. Memory and Learning. 5. Language and Thinking; (B) Basic Research for the elucidation of biological functions through molecular level approaches: 1. Expression of Genetic Information. 2. Morphogenesis. 3. Molecular Recognition and Responses. 4. Energy Conversion. Applications for Short-Term Fellowships and Workshops can be submitted throughout the year. Eligible countries are Canada, France, Germany, Italy, Japan, Switzerland, U.K., U.S.A. and nonsummit EC member countries. Short-Term (up to 3 months) Fellowships are for researchers from the eligible countries who wish to do research in foreign countries, or for researchers outside the eligible countries who wish to do research in one of the eligible countries. Grants are available for international workshops organized by researchers from the eligible countries. Contact: H.F.S.P.O., Tour Europe, 20 place des Halles, 67080 Strasbourg, France [(France 33)-88-32-88-33; FAX: (France 33)-88-32-88-97].
Dissertation Research Funds
The National Science Foundation's Biological Science Directorate invites applications for doctoral dissertation research in environmental biology (including ecological studies and systematic, population, and integrative biology) and neuroscience (including animal behavior and functional and physiological ecology). Deadlines are Sept. 1 and Feb. 1. Awards range from $2,000 to $20,000 a year for one or two years, but grants usually are about $10,000. Only students enrolled at U.S. colleges and universities are eligible. Doctoral advisers must submit proposals on behalf of students. Contact: Elizabeth Behrens, Div. of Environmental Biology, NSF, Biological Sciences Directorate, Room 215, 1800 G St. NW, Washington, DC 20550 [202-356-9596] and Div. of Integrative Biology and Neuroscience, Room 321, same address [202-357-7949].
Helicobacter pylori Pathogenesis
The National Institute of Allergy and Infectious Diseases (NIAID) and the National Institute of Diabetes and Digestive and Kidney Diseases (NIDDK) invite submission of research applications for support of basic and clinical studies needed to establish the role of bacterial and host factors in the pathogenesis of the various forms of Helicobacter pylori disease, including infection, gastritis, and duodenal ulceration. The overall goal of this program is to stimulate innovative research to define the host and bacterial factors involved in the various phases of H. pylori infection, excluding gastric carcinoma and lymphoma. This may include animal model studies. Areas of interest include definition of the virulence determinants of H. pylori, and descriptions of the pathophysiology and immunopathology of infection. Multidisciplinary studies involving collaboration among investigators with expertise in appropriate disciplines is encouraged. For information, contact Leslye D. Johnson, Ph.D., Div. of Microbiology & Infectious Diseases, NIAID, Solar Bldg, Rm 3A-06, 6006 Executive Blvd, Bethesda, MD 20892 [301-496-7051; FAX: 301-402-2508] or Frank Hamilton, M.D., M.P.H., Div. of Digestive Diseases, NIDDK, Westwood Bldg, Rm 3A-16, Bethesda, MD 20892 [301-594-7571].
Hastings Center
The Hastings Center has a number of fellowships, internships, and grants available for students; academic, medical, or legal professionals; or journalists who have a serious interest in pursuing directed or independent study of bioethical issues relating to animal welfare or other fields of scientific concern. For information contact: Director of Education, The Hastings Center, 255 Elm Rd, Briarcliff Manor, NY 10510 [914-762-8500].
Zoo Research Grants
The American Association of Zoo Keepers announces two $750 research grants in the field of zoo biology. Interested applicants should direct their inquiries to Sue Barnard, Chairperson, AAZK Research/Grants Committee, Zoo Atlanta, Dept of Herpetology, 800 Cherokee Ave, SE, Atlanta, GA 30315. Deadline for submissions is 1 March 1994.
American Cancer Society
Research and Clinical Investigation Grants to fit a variety of needs in scientific investigations related to cancer are awarded to nonprofit institutions in the U.S. A grant is generally made to cover the cost of such items as salaries for professional and technical assistance, special equipment, animals, consumable supplies and other miscellaneous items required to conduct the proposed research. For information, contact: American Cancer Society, Research Dept, 1599 Clifton Rd NE, Atlanta, GA 30329 [404-329-7558]. Application deadline is October 15, 1993.
* * *
The first Conference on Environmental Enrichment was held at Portland's Washington Park Zoo July 16-20, 1993. Organized by David Shepherdson and Jill Mellen, with ample help from the zoo's volunteers, the conference was certainly a success in terms of the number of people attending--about 180--and the 11 countries represented--U.S., Canada, England, Scotland, Australia, Denmark, Netherlands, Mexico, Switzerland, Sweden, and Finland. A broad diversity of animal groups was discussed, and even plants and their uses and abuses were mentioned. Although the majority of attendees were from zoos, a number of people with particular interest in primates came from various laboratories. The successive sessions over four days included Overview/ Theoretical Background, Well-being of Individuals and Populations, Husbandry Perspectives, Exhibit Design and Planning, Broadening Horizons, The Human Factor, Evaluation Examples and Ideas, and Putting Enrichment into Practice. Enrichment strategies for selected taxonomic groups were developed in working groups one afternoon, with the results being forwarded to the respective Taxon Advisory Group. These strategies were particularly focused on zoo exhibits. A "Training as Enrichment" demonstration was also presented.
Attendees repeatedly mentioned the greater effectiveness of enrichment devices that provided control over the environment. Foraging devices were the most common example. With respect to various toy-type objects, the problem of habituation to novel objects was reported for many species. Several people at the conference mentioned the benefits of varying feeding schedules to reduce stereotypies, and some of the health benefits of feeding more than once a day. It was generally agreed that the reduction or elimination of self-abusive behavior was a major goal of enrichment. Noninjurious stereotypies and other abnormal behaviors are also undesirable in a zoo setting where the public can see them. However, there was considerable disagreement about what these stereotypies indicate about the animals' well-being. Janet Baer gave the veterinarian's point of view and cautioned about the potential hazards of enrichment--disease/bacteria, entanglement, and the potential injuries associated with social housing. Various people emphasized the importance of human/animal interactions, which are obviously necessary for managing animals in zoo exhibits. Positive interactions in the lab environment are equally important. Implementing positive caretaker/primate relationships is one of the major ways to improve the well-being of captive primates. Several representatives from the USDA attended the meeting, including Jo Anne Garbe, who gave an illuminating talk on the regulatory perspective of environmental enrichment. Many attendees from zoological gardens were unaware that their institutions are supposed to have documented environmental enhancement plans to promote the psychological well-being of nonhuman primates. -- A roving unnamed contributor.
* * *
The New England Regional Primate Research Center, Harvard Medical School, is seeking a clinical veterinarian in the Division of Primate Resources. Primary responsibilities include clinical and preventive care of 1,700 primates of seven species. The veterinarian will also work with the veterinary and husbandry staff to administer environmental enrichment programs, assure compliance with Federal and NIH standards, and conduct clinical research. There will be ample opportunities for collaboration and clinical education of veterinarians and veterinary students. AAALAC-accredited facilities include a biocontainment suite and ultrasound and endoscopy equipment. Advanced training in laboratory animal medicine, internal medicine or primatology required.
Please send curriculum vitae and names of three references to: David Lee-Parritz DVM, Chairman, Division of Primate Resources, New England Regional Primate Research Center, PO Box 9102, Southboro, MA 01772-9102.
* * *
Squirrel Monkey Twins Reported
Julio C. Ruiz of the Centro Argentino de Primates (Casilla de Correo 145, 3400 Corrientes, Argentina) reports the first known birth of twins in Saimiri boliviensis. The female was imported from Bolivia as a pregnant adult in 1979; she aborted soon after that. From 1980 to 1991, she was pregnant five times, and bore four infants normally. During the winter of 1991 she gave birth to twins, a stillborn male weighing 75 grams and a female weighing 60 grams at birth. The female infant survived only two days. Her birth weight was 30 grams less than what is considered the minimum weight for survival in this species.
"Of the primates composing the Cebidae family of New World monkeys, only the capuchin (Cebus) and the howler (Alouatta) have been found to twin and then only very rarely." (Quote from D. E. Wildt & W. R. Dukelow (1974), The nonhuman primate as a model for human twinning. Laboratory Primate Newsletter, 13[4], 15-18.]
Digit Fund Changes Its Name
In honor of the 25th anniversary of the founding of Karisoke Research Center by Dr. Dian Fossey, the Board of Trustees announced a name change to the Dian Fossey Gorilla Fund. Reaffirming its mission to protect the gorilla, the Fund urges all interested in helping to preserve the gorilla population to contact them at: 45 Inverness Drive East, Suite B, Englewood, CO 80112-5480.
Malaria-AIDS Link?
Harold McClure, Yerkes Chief of Pathobiology & Immunology, reported at a American Society for Microbiology meeting on a pig-tailed macaque, who previously had shown no evidence of infection by the malaria parasite, developing acute clinical malaria after infection with an SIV. This suggests that scientists who conduct SIV studies with wild-caught pig-tailed macaques, increasingly used in SIV studies, should be alert to the development of clinical malaria in these monkeys. -- From a press release from Yerkes RPRC, Emory Univ., Atlanta, GA 30322.
VandeBerg to Lead ILAR
Dr. John L. VandeBerg, Chairman of the Department of Genetics at the Southwest Foundation for Biomedical Research, has been appointed chairman of the Institute of Laboratory Animal Resources (ILAR) Council.
Consisting of five staff members and a 14-member council, ILAR is an integral advisory panel within the National Research Council, the principal operating agency of the National Academy of Sciences which advises the federal government in science and technology matters.
"We are advocates of the use of alternatives to animals when it's appropriate," said Dr. VandeBerg, a reviewer of ILAR News manuscripts and previous chairman of the ILAR News advisory panel. Alternatives are often referred to as the 3 R's -- refinement, reduction, and replacement of animals in research. Some non-animal methods, Dr. VandeBerg noted, can provide faster results at lower costs; these include bacteria or cell cultures, epidemiological surveys or computer simulations. These must, however, be based on knowledge derived from research on either animals or humans.
ILAR has the important responsibility of updating the "Guide for the Care and Use of Laboratory Animals" for the National Institutes of Health, which sponsors 90 percent of biomedical research in the United States, and for other federal agencies. Compliance with these national guidelines is required by federal law for all Public Health Service grantees. The Guide was last updated in 1985, and one of ILAR's immediate goals under Dr. VandeBerg's leadership will be to revise the Guide to reflect knowledge and perspectives that have evolved since then.
New SCAW Board Members
The Scientists Center for Animal Welfare, a nonprofit educational organization concerned with promoting both good science and the responsible and humane use of animals in testing, research, and education, announces the election of two new additions to its Board of Trustees: Vernon S. Bishop of the Department of Physiology at the University of Texas Health Science Center and Gordon M. Burghardt, Professor of Psychology and Professor of Zoology at the University of Tennessee.
Birute Galdikas Honored
The Sierra Club awarded Birute Galdikas the Chico Mendes award, which is presented to "individuals or nongovernmental organizations, outside of the United States, who have exhibited extraordinary courage and leadership, at the grassroots level, in the universal struggle to protect the environment." Also, on May 13 Dr. Galdikas was given the Chevron Conservation Award, which honors conservationists from the professional, volunteer, and nonprofit realms. Dr. Galdikas studies and protects the orangutan in Borneo.
Tonkin Snub-nosed Langurs Sighted
Recently there has been a confirmed sighting of a "missing" primate in Vietnam. The Tonkin snub-nosed langur, Rhinopithecus avunculus, was thought to be extinct because it had not been sighted in the wild since the late 1960s. (The presumed extinction was partly the result of past military activity in the area.) Following a report that the leaf-eating monkey was still living in a remote region of northern Vietnam, a team from Primate Conservation, Inc. in East Hampton, New York, set out to locate the species in its natural habitat. About half a day's walk from Na Hang village in Tuyen Quang province, a troop of 25 of these langurs was seen traveling through trees growing on a steep limestone ridge. The local people hunt monkeys for traditional medicines, the pet trade, and food, so the Tonkin langur is extremely shy of humans and almost nothing is known about its behavior. Le Xuan Kanh, a Vietnamese scientist who joined the team, estimates the species' population may number only 150, making it one of the most endangered primates in Asia. NYZS/The Wildlife Conservation Society has signed an accord with the Vietnamese government to start a long-term study of the Tonkin snub-nosed langur this fall. From a report by Noel Rowe in Wildlife Conservation, 1993, 94[6 ], 7.
* * *
Wisconsin RPRC Seminars
The Wisconsin Regional Primate Research Center holds weekly seminars in its Conference Room at 1220 Capitol Court, Madison, WI 53715-1299 during the Spring and Fall Semesters. For information contact Philippa Claude [608-263-3525; FAX: 608-263-4031; E-mail: [email protected]].
Behavioral Ecology Congress
The Fifth International Behavioural Ecology Congress will be held 14-20 August, 1994 at the Univ. of Nottingham, England. For information contact: ISBE Congress, Conference Nottingham, Business Information Centre, 309 Hayden Rd, Nottingham NG5 IDC, UK.
Anesthesia, Analgesia and Surgery
Scientists Center for Animal Welfare (SCAW) will sponsor a two-day conference entitled "Research Animal Anesthesia, Analgesia and Surgery" on May 12-13, 1994 in Atlanta, GA. Researchers, regulatory personnel, members of Animal Care and Use Committees, administrators and others interested in these issues are encouraged to attend. For more information, contact: Conferences, SCAW, 4805 St. Elmo Ave, Bethesda, MD 20814 [301-654-6390; FAX: 301-907-3993].
Human/Research Animal Relationship
SCAW will sponsor a seminar on "The Human/Research Animal Relationship in the Laboratory" on November 16, 1993 from 8:00 to 11:00 a.m. at the National AALAS Meeting to be held in Nashville, TN. AALAS attendees are welcome to attend. The seminar will address topics such as: History of the Human/Animal Relationship, Human/Research Animal Relationships: Animal Care Staff Perspectives, Human/Research Animal Relationships: Research Staff Perspectives, How Human/Animal Bonding Affects the Animals, and Personnel Considerations: Hiring, Training, Attitudes. Principal Investigators, animal care technicians, attending veterinarians and others are encouraged to attend and share their experiences. For more information contact SCAW at the address given above.
* * *
Import Permits
According to the NABR Update (1993, 14[11]), about 30% of all animal and plant materials sent internationally to U.S. scientists are delayed or stopped because they don't have an import permit from the U.S. Dept. of Agriculture's Animal and Plant Health Inspection Service. Scientists intending to import animal materials should contact, in advance, the Import-Export Product Staff, Veterinary Services Program, USDA, APHIS, 6505 Belcrest Rd, Hyattsville, MD 20782 [310-436-7885].
Nonhuman Primate Audiovisual Resources
Two new slide/audio/manual programs from the American College of Laboratory Animal Medicine are now available for $120 each, plus shipping. V-9018 (Nonhuman Primates: Biosafety, by J. R. Broderson, 1992) describes the four major classes of health hazards within a nonhuman primate facility and the practices, equipment, and facility design that can help prevent disease or injury from these hazards [60 slides, 17-minute audiocassette, and manual]. V-9019 (Nonhuman Primates: Environmental Enrichment, by K. Bayne, 1992) describes how to promote psychological wellbeing in nonhuman primates by social, nonsocial, contact, and noncontact approaches [61 slides, 27-minute audiocassette, and manual].
These programs are from a long-standing series on laboratory animal
medicine and science, consisting of 42 titles on topics ranging from
various species to more general programs regarding diverse topics such
as alternatives, laws, barrier technology, anesthesia and surgery. The
entire series is currently being updated by the University of Washington Health Sciences Center for Educational Resources. The nonhuman primate titles are co-sponsored by the Association of Primate Veterinarians and the USDA NAL. When finished--in 1994--this part of the
series will include the following eleven titles:
*Available in 1993:
V-9010, Nonhuman Primates: Introduction and Taxonomy, by A. F. Moreland. V-9011, Nonhuman Primates: Use in Research, by A. F. Moreland.
V-9012, Nonhuman Primates: Biology (Old World), by H. J. Klein & G.
Y. McCormick. V-9013, Nonhuman Primates: Biology (New World), by H.
J. Klein & G. Y. McCormick. V-9014, Nonhuman Primates: Care and Management, by W. E. Britz, Jr. & T. M. Butler.
*Available in 1994:
V-9015, Nonhuman Primates: Viral Diseases, by B. M. Flynn. V-9016,
Nonhuman Primates: Bacterial Diseases, by W. C. Cole & J. A. Roberts. V-9017, Nonhuman Primates: Parasitic Diseases, by R. E. Banks
& J. A. Davis. V-9020, Nonhuman Primates: Noninfectious Diseases, by
R. E. Banks & J. A. Davis. For more information, contact Bob J. McGough, Univ. of Washington, T-279 HSB, SB-56, Seattle, WA 98195 [206-685-1161; FAX: 206-543-8051; e-mail: [email protected]]. For a
free catalog of programs, contact Marie Gary, Distribution Manager,
Univ. of Washington, T-252 HSB, SB-56, Seattle, WA 98195 [206-685-1186; FAX: 206-543-8051; e-mail: [email protected]].
Information about Yerkes in Florida
Donald A. Dewsbury of the University of Florida is beginning work on a history of the Yale/Yerkes Laboratories of Primate Biology (YLPB) in Orange Park, FL. He is collecting material and information related to the YLPB and those who worked there. He would like to contact anyone with knowledge of the YLPB and receive any recollections, correspondence, etc., related to both the YLPB and Robert Yerkes himself. Contact him at: Dept. of Psychology, Univ. of Florida, Gainesville, FL 32611-2065 [904-392-0596; FAX: 904-392-7985; e-mail: [email protected]].
Pathology Slide Set
Slide sets illustrating 16 interesting primate cases, complete with clinical histories, results, and discussion, are available through the Registry of Comparitive Pathology. The cases were compiled for the 1993 annual Primate Pathology Workshop, which is held in conjunction with the U.S. and Canadian Academy of Pathology meeting. Cost for the set is $15; checks are to be made payable to UAREP and sent to Registry of Comparative Pathology, AFIP, Washington, DC 20306-6000. For additional information contact the Registry at 202-576-2452 [FAX: 202-576-9161].
* * *
Books
* A Primate Model for the Study of Colitis and Colonic Carcinoma: The Cotton-Top Tamarin (Saguinus oedipus). N. K. Clapp (Ed.). Boca Raton, FL: CRC Press, 1993. 384 pp. [Price: $199.95]
* Prospettive nello Studio dei Primati. Contributi Italiani alla Primatologia Internazionale. [Perspectives in the Study of Primates: An
Italian Contribution to International Primatology] Volume II. A. Bava
(Ed.). 1991. 238 pp.
. .
Papers from the Sixth Congress of the Italian Primatological Association.
Bibliographies
* Longevity in Nonhuman Primates: A Selected Bibliography, 1985-1993. M. McLean. Seattle: Primate Information Center, 1993. 19 pp. (215 citations, primate index) [Price: $6.50. Stock #93-003. Order from PIC, RPRC, SJ-50, Univ. of Washington, Seattle, WA 98195]
* Aging in Nonhuman Primates: Biology and Behavior: A Selected Bibliography, 1985-1993. M. McLean. Seattle: Primate Information Center, 1993. 34 pp. (379 citations, primate and subject indexes) [Price: $6.50. Stock #93-004. Ordering information same as above]
Bulletins
* Comparative Pathology Bulletin, 1993, 25[3]. [Registry of Comp.
Pathology, AFIP, Washington, DC 20306]
. .
Includes an article on the sooty mangabey as a model for leprosy
(Hansen's disease).
Directories
* Directory of the Consortium of Aquariums, Universities and Zoos
1993-1994. D. F. Hardy (Coordinator). Northridge: Cal. State University, 1993. [Price: $15 in U.S. and Canada; $20 elsewhere, from D.
Hardy, Dept of Psychology, Cal. State Univ., Northridge, CA 91330]
. .
Listings of educators, researchers, students, and technical staffs
who are interested in collaborating and communicating; with various
indexes.
Magazines, Newsletters, and Reports
* IPPL News, 1993, 20 [2]. (IPPL, P.O. Box 766, Summerville, SC 29484).
* The Johns Hopkins Center for Alternatives to Animal Testing. 1993, 10[2 and 3]. (Office of Public Affairs, Johns Hopkins Sch. of Public Health, 615 N. Wolfe St, Baltimore, MD 21205-2179).
* SCAW Newsletter, 1993, 15[2]. [4805 St. Elmo Ave, Bethesda, MD
20814]
. .
Includes Part 2 of a summary of new animal welfare regulations, by
R. R. Smeby, and a report on the 1993 AVMA Panel on Euthanasia Report,
by B. T. Bennett.
* Asian Primates: A Newsletter of the IUCN/SSC Primate Specialist
Group. December, 1992/March, 1993, 2[3/4]. [A. A. Eudey, Ed., 164
Dayton St., Upland, CA 91786]
. .
Includes "Evidence for the existence of Trachypithecus francoisi
hatinhensis," by Le X. C., and "Commonly encountered diseases in golden monkeys, and their prevention, under captive conditions," by Fan
P.-Z., Gao Y.-F., & Liu S.-F.
* The Newsletter, 1993, 5[1]. (Primate Foundation of Arizona, P.O.
Box 20027, Mesa, AZ 85277-0027)
. .
Includes "Chimpanzee (Pan troglodytes) infant age-related changes
in the dynamics of the mother-infant relationship," by S. M. Howell,
J. Fritz, & L. T. Nash.
* Neotropical Primates: A Newsletter of the Neotropical Section of the IUCN/SSC Primate Specialist Group, 1993 , 1[2] (Conservation International, 1015 18th St NW, Suite 1000, Washington, DC 20036)
* Baboon Update, Spring/Summer, 1993 (Community Conservation Consultants, Howlers Forever, Inc., RD 1, Box 96, Gays Mills, WI 54631).
* Wisconsin Regional Primate Research Center, Winter 1992-93, 3[1].
Special Journal Issues
*Red and white blood cell groups of chimpanzees: Relationship to their
human counterparts. Journal of Medical Primatology, 1993, 22[1].
. .
Papers presented at the XIVth Congress of the IPS, Strasbourg,
France, August 16-21, 1992, edited by W. W. Socha. Contents: Blood
groups of primates: Historical perspective and synopsis, by J. Moor-Jankowski. The M-N-V-A-B-D blood group system of chimpanzee and other
apes: Serology and genetics, by W. W. Socha & J. Moor-Jankowski. Primate genes for glycophorins carrying MN blood group antigens, by S.
Kudo, M. Onda, A. Rearden, & M. Fukuda. The R-C-E-F blood group system
of chimpanzees: Serology and genetics, by A. Blancher, J. Ruffie, & W.
W. Socha. Relationship between chimpanzee Rh-like genes and the
R-C-E-F blood group system, by I. Salvignol, A. Blancher, P. Calvas,
W. W. Socha, Y. Colin, J.-P. Cartron, & J. Ruffie. Quantitative study
of chimpanzee and gorilla counterparts of the human D antigen, by F.
Roubinet, A. Blancher, W. W. Socha, & J. Ruffie. Recent advances in
molecular and genetic analysis of RH blood group structures, by Y.
Colin, B. Cherif-Zahar, C. Le Van Kim, I. Mouro, & J.-P. Cartron.
Application of maximum likelihood statistics for the population and
family studies of inheritance of the chimpanzee R-C-E-F and V-A-B-D
blood groups, by A. Blancher, J. Clayton, W. W. Socha, & J. Ruffie.
Major histocompatibility complex class II polymorphisms in humans and
chimpanzees, by R. E. Bontrop, M. Kenter, N. Otting, & M. Jonker.
Trans-specific Mhc polymorphism and the origin of species in primates, by J. Klein, Y. Satta, N. Takahata, & C. O'hUigin. Immunoglobulin Gm allotypes in apes: Comparison with man, by J.-M. Dugoujon,
A. Blancher, S. Hazout, & J. Ruffie.
* Program and Abstracts of the Sixteenth Annual Meeting of the American Society of Primatologists, August 18-22, 1993, Sturbridge Massachusetts. American Journal of Primatology, 1993, 30[4].
* Zoonooz, 1993, 66[4]. (Zool. Soc. of San Diego, P.O. Box 551, San
Diego, CA 92112)
. .
Several articles on pygmy chimpanzees.
Anatomy & Physiology
* Urinary and plasma gonadotropin concentrations in golden lion tamarins (Leontopithecus r. rosalia). French, J. A., DeGraw, W. A., Hendricks, S. E., Wegner, F., & Bridson, W. E. (Dept of Psychology, Univ.
of Nebraska, Omaha, NE 68182). American Journal of Primatology,
1992, 26, 53-59.
. .
Development and validation of an immunoassay useful for monitoring
reproductive function in both female and male lion tamarins.
* Differential imaging of ocular dominance and orientation selectivity
in monkey striate cortex. Blasdel, G. G. (Dept of Neurobiology, Harvard Med. School, 220 Longwood Ave, Boston, MA 02115). Journal of
Neuroscience, 1992, 12, 3115-3138.
. .
It is shown that: groups of cells preferring a single orientation
cannot be described as slabs extending farther than 0.5 mm laterally;
even short iso-orientation slabs are not aligned with the bands seen
in single differential images; regions strongly dominated by one eye
and regions strongly selective for orientation are partially segregated in the lateral domain; and orientation selectivity correlates
highly with binocularity.
* Orientation selectivity, preference, and continuity in monkey striate
cortex. Blasdel, G. G. (Address same as above). Journal of Neuroscience, 1992, 12, 3139-3161.
. .
Periodic interspersing of linear zones and singularities suggests
that orientation preferences are organized by at least two competing
schemes. Since upper-layer neurons are likely to have similarly sized
dendritic fields in all regions, those in the linear zones should
receive precise information about narrowly constrained orientations,
while those near singularities should receive coarse information about
all orientations.
* Langerhans cells, macrophages and lymphocyte subsets in the cervix
and vagina of rhesus macaques. Miller, C. J., McChesney, M., & Moore,
P. F. (CRPRC, Univ. of California, Davis, CA 95616). Laboratory
Investigation, 1992, 67, 628-634.
. .
Observations providing a morphological basis for mucosa-associated
lymphoid tissue in the female genital tract.
*Cerebrospinal fluid monoamine and adrenal correlates of aggression in
free-ranging rhesus monkeys. Higley, J. D., Mehlman, P. T., Taub, D.
M., Higley, S. B., Suomi, S. J., Vickers, J. H., & Linnoila, M. (Animal Center, NIH, Lab. of Clinical Studies, P.O. Box 289, Poolesville,
MD 20837). Archives of General Psychiatry, 1992, 49, 436-441.
. .
Based on direct observation of participation or avoidance of
aggressive behavior and apparent fight wounds, 18 monkeys were
selected, along with 10 random controls, from 4200 on Morgan Island
for cerebrospinal fluid taps and blood samples. There was a significant negative correlation between high rankings for aggression and
cerebrospinal fluid 5-hydroxindoleacetic acid concentration. Cerebrospinal fluid, norepinephrine, and plasma corticotropin and cortisol
concentrations were positively correlated with high aggression rankings.
*The striatopallidal projection displays a high degree of anatomical
specificity in the primate. Hazrati, L.-N. & Parent, A. (A. P., Centre
de recherche en neurobiologie, Hopital de l'Enfant-Jesus, 1401, 18e
Rue, Quebec, P.Q., G1J 1Z4, Canada). Brain Research, 1992, 592,
213-227.
. .
In contrast to previous beliefs, axons of striatal neurons from two
small adjacent populations do not converge upon the same pallidal neurons, but instead project to several distinct subsets of pallidal neurons. The striatopallidal projection system in primates is highly
ordered and displays a high degree of specificity with respect to its
target sites in the pallidum.
Animal Models
*Spontaneous lesions in cynomolgus monkeys used in toxicity studies.
Ito, T., Chatani, F., Sasaki, S., Ando, T., & Miyajima, H. (Drug
Safety Res. Labs, Takeda Chemical Industries, Ltd, 6-10-1 Himuro-cho,
Takatsuki-shi, Osaka 569, Japan). Experimental Animals, 1992, 41,
455-469.
. .
Spontaneous lesions in wild-caught, lab-maintained monkeys were
examined histopathologically in an effort to better distinguish toxic
changes from spontaneous lesions and assess the toxicity of drugs more
exactly.
*Aneurysmal enlargement of the aorta during regression of experimental
atherosclerosis. Zarins, C. K., Xu, C., & Glagov, S. (Sect. of Vascular Surgery, Univ. of Chicago, 5841 S. Maryland Ave, Chicago, IL
60637). Journal of Vascular Surgery, 1992, 15, 90-101.
. .
Evidence of early aneurysmal dilation of the abdominal aorta in
monkeys undergoing dietary cholesterol lowering supports the hypothesis that the atherosclerotic process plays a significant role in the
pathogenesis of aneurysm formation. Plaque regression and thinning of
the aortic wall may initiate aneurysmal enlargement.
* Helicobacter infection in animals: A clue to the role of adhesion
in the pathogenesis of gastroduodenal disease. O'Rourke, J. L., Lee,
A., & Fox, J. G. (School of Microbiology & Immunology, Univ. of New
South Wales, P.O. Box 1, Kensington, NSW, Australia 2033). European
Journal of Gastroenterology & Hepatology, 1992, 4, S31-S37.
. .
The authors conclude that adhesion to the gastric mucosa is not an
essential component in Helicobacter-induced gastritis. However, it
is possible that close contact between bacterium and host is required
for ulcerogenesis.
*Vesicoureteral reflux and pyelonephritis in the monkey: A review.
Roberts, J. A. (Dept of Urology, Tulane RPRC, 18703 Three Rivers Rd,
Covington, LA 70433). The Journal of Urology, 1992, 148, 1721-1725.
. .
Early detection and rapid and effective antibacterial treatment of
acute pyelonephritis remain the best hope to prevent chronic atrophic
pyelonephritis and end-stage renal disease.
*Cryopreserved primate bone marrow cells can be used for retroviral-mediated gene transfer. Wieder, R. (Dept of Med., Memorial Sloan-Kettering Cancer Ctr, New York, NY 10021). Human Gene Therapy, 1991, 2, 323-326.
*A novel approach to neural transplantation in Parkinson's disease:
Use of polymer-encap- sulated cell therapy. Emerich, D. F., Winn, S.
R., Christenson, L., Palmatier, M. A., Gentile, F. T., & Sanberg, P.
R. (CytoTherapeutics, Inc., 2 Richmond Sq., Providence, RI 02906).
Neuroscience and Biobehavioral Reviews, 1992, 16, 437-447.
. .
Striatal implants of encapsulated dopamine-producing cells promote
behavioral recovery in rodent and primate models of Parkinson's disease.
*Behavioral effects of adrenal medullary transplants in non-human primates. Dubach, M. (RPRC SJ-50, Univ. of Washington, Seattle, WA
98195). Journal of Neural Transplantation & Plasticity, 1992, 3,
97-114.
. .
Lesions induced by intracerebral injection of 6-hydroxydopamine in
the substantia nigra of 24 long-tailed macaques produced the expected
chronic decrease in percentage of contralateral turning in most. Animals that received the longest viable ribbon grafts showed a reversal
of this effect back toward base line, whereas monkeys whose grafts
left little or no surviving tissue showed no behavioral improvement.
*Changes in the cortical map of the hand following postnatal median
nerve injury in monkeys: Modification of somatotopic aggregates. Wall,
J. T., Huerta, M. F., & Kaas, J. H. (Dept of Anatomy, Med. College of
Ohio, Toledo, OH 43699). Journal of Neuroscience, 1992, 12,
3445-3455.
. .
A hypothesis is proposed, based on a study of 8 marmosets, suggesting that the cortical hand map normally consists of a number of repre-
sentations that are capable of developing and surviving somewhat
autonomously of each other. The features of spatial patterning in the
mosaiclike map of these representations are influenced by postnatal
availability of inputs from intact hand nerves.
*Stress and sodium hypertension in baboons: Neuroendocrine and pharmacotherapeutic assessments. Turkkan, J. S. & Goldstein, D. S. (Johns
Hopkins Univ. School of Med., 617 Traylor Bldg, 720 Rutland Ave, Baltimore, MD 21205). Journal of Hypertension, 1991, 9, 969-975.
. .
Neither a beta-adrenoceptor blocker nor a thiazide diuretic ameliorated the increased systolic blood pressure caused by salt loading and
stress in four baboons.
*One-year monkey heart xenograft survival in cyclosporine-treated
baboons: Suppression of the xenoantibody response with total-lymphoid
irradiation. Roslin, M. S., Tranbaugh, R. E., Panza, A., Coons, M. S.,
Kim, Y.-D., Chang, T., Cunningham, J. N., & Norin, A. J. (A. J. N.,
Box 19, Depts of Med. & Anat. & Cell Bio., SUNY Health Sci. Center,
450 Clarkson Ave, Brooklyn, NY 11203). Transplantation, 1992, 54,
949-955.
. .
Prior to this study it has not been possible to prolong concordant
xenograft survival in primates to a duration that would be useful in
human heart transplantation with standard techniques.
* In vitro and in vivo evaluation of microencapsulated porcine
islets. Sun, A. M., Vacek, I., Sun, Y.-L., Ma, X., & Zhou, D. (Dept of
Physiology, Med. Sciences Bldg, Univ. of Toronto, 1 King's College
Cir., Toronto, M5S 1A8 Canada). ASAIO Journal, 1992, 38, 125-127.
. .
A xenograft of 50,000 microencapsulated porcine islets into a spontaneously diabetic monkey normalized hyperglycemia for more than 150
days.
*Intrauterine xenotransplantation of bone marrow stem cells in nonhuman primates. Michejda, M., Peters, S. M., Bacher, J., Hernandez, L.
F., & Bellanti, J. A. (Georgetown Univ. Med. Center, Washington, DC
20007). Transplantation, 1992, 54, 759-762.
. .
Results suggest that intrauterine xenotrans- plantation of bone
marrow stem cells may be utilized safely for stem cell reconstitution.
*Depletion of natural antibodies in non-human primates--A step towards
successful discordant xenografting in humans. Cooper, D. K. C. (Oklahoma Transplantation Inst., Baptist Med. Center, Oklahoma City, OK
73112). Clinical Transplantation, 1992, 6, 178-183.
. .
A review of methods and techniques for depleting and inhibiting
natural antibodies, and efforts to identify the carbohydrate epitopes
on pig organs (used in baboon allografts) against which human anti-pig
antibodies are directed.
*Monoclonal antibody to tumor necrosis factor-alpha prevents lethal
endotoxin sepsis in adult rhesus monkeys. Fiedler, V. B., Loof, I.,
Sander, E., Voehringer, V., Galanos, C., & Fournel, M. A. (Lehmer
Muhle 46, D-5090 Leverkusen 3, Germany). Journal of Laboratory and
Clinical Medicine, 1992, 120, 574-588.
. .
Short-term experiments with rhesus monkeys indicate protection
against lipopolysacchar- ide-induced endotoxin sepsis by anti-TNF
MoAB, which may have clinical relevance for the treatment of human
septicemia.
*Cobalamin deficiency and the pathogenesis of nervous system disease.
Metz, J. (Dept of Hematology, South African Inst. for Med. Research,
Johannesburg 2000, South Africa). Annual Review of Nutrition, 1992,
12, 59-79.
. .
Review of animal models (monkey, fruit bat, pig) of cobalamin deficiency.
*Effects of in vivo and in vitro administration of morphine sulfate upon rhesus macaque polymorphonuclear cell phagocytosis and chemotaxis. Liu, Y., Blackbourn, D. J., Chuang, L. F., Killam, K. F. Jr., & Chuang, R. Y. (R. Y. C., Dept. of Med. Pharmacology & Toxicology, School of Med., Univ. of California, Davis, CA 95616-8654). Journal of Pharmacology and Experimental Therapeutics, 1992, 263, 533-539. Polymorphonuclear cells prepared from morphine-treated animals showed a marked, transient reduction in their ability to kill ingested yeast blastospores. In vitro studies suggested that the killing activity and the chemotaxis of the cells were reduced by treatment with 50pM, but not 50 fM, morphine sulfate.
*Effects of central administration of opioids on facial scratching in
monkeys. Thomas, D. A., Williams, G. M., Iwata, K., Kenshalo, D. R.,
Jr., & Dubner, R. (NAB, NIDR, NIH, Bldg 30, Rm B20, 9000 Rockville
Pike, Bethesda, MD 20892). Brain Research, 1992, 585, 315-317.
. .
One of three opioid-receptor agonists, DAMGO, produced large,
dose-dependent, naloxone-reversible increases in facial scratching
when injected into the medullary dorsal horn of cynomolgus monkeys.
*Proximal and distal cannulation of the internal jugular vein for ECMO
in a primate. Kitagawa, H., Mccomb, G., Ford, E., & Atkinson, J. B.
(J. B. A., Div. of Pediatric Surgery, Box 100, Childrens Hospital,
4650 Sunset Blvd, Los Angeles, CA 90027). Journal of Pediatric Surgery, 1992, 27, 1189-1191.
. .
Extracorporeal membrane oxygenation (ECMO) is lifesaving for
infants with severe respiratory distress but is complicated by severe
intracranial hemorrhage in 10-30% of patients. Experiments with cynomologus monkeys show that cephalad venous cannulation is not necessary
in all ECMO cases, but should be reserved for patients requiring additional venous drainage.
*Detection of anti-human cell antibodies in sera from macaques immunized with whole inactivated virus. Langlois, A. J., Weinhold, K. J.,
Matthews, T. J., Greenberg, M. L., & Bolognesi, D. P. (Duke Univ.
Med. Center, Center for AIDS Research, Durham, NC 27710). AIDS
Research and Human Retroviruses, 1992, 8, 1641-1652.
. .
Vaccination with whole inactivated SIV grown in human cell lines
leads invariably to the production of serum reactivities that target
surface elements of human cells, but do not protect against challenge
with SIV grown in macaque cells. It would appear that additional studies employing whole inactivated virus preparations originating from
cells of the species in which the vaccine is to be tested would minimize the anti-cell reactivity and enhance the antiviral reactivity.
Such a study would provide a better setting to determine the efficacy
with which virion antigens are able to confer protection in a vaccine
setting of a whole inactivated virus vaccine.
*Long-term acid-base measurements in the fetal and maternal baboon.
Daniel, S. S., James, L. S., MacCarter, G., Morishima, H. O., & Stark,
R. I. (Dept of Anesthesiology, Columbia Univ. College of Physicians &
Surgeons, 630 W. 168th St., New York, NY 10032). American Journal of
Obstetrics & Gynecology, 1992, 166, 707-712.
. .
Results from chronically catheterized baboon fetuses and mothers
(maintained in a tether system) show the baboon fetus has slightly
higher pH and lower PaCO� than the human fetus. PaO�, base deficit,
and maternal-fetal gradients across the placenta in these two primate
species are comparable.
Animal Welfare
*Humans enrich the lives of lab baboons. Choi, G. (Dept. Lab. Animal
Med., Univ. of Cincinnati, Cincinnati, OH 45267-0571). Our Animal
WARDS, Summer, 1993, 3-7, 13.
. .
Description of an unusual program.
Behavior
*Tool-assisted predation on a squirrel by a female chimpanzee in the
Mahale Mountains, Tanzania. Huffman, M. A. & Kalunde, M. S. (Dept of
Zoology, Kyoto Univ., Sakyoku, Kyoto 606, Japan). Primates, 1993,
34, 93-98.
. .
A 12-year-old female chimpanzee used a sturdy tool modified from a
branch of a tree to rouse and capture a squirrel hiding in a hole in
the tree. This appears to be the first reported case of tool use by a
nonhuman primate to capture a mammalian prey.
*Predator mobbing by an all-male band of Hanuman langurs (Presbytis
entellus). Ross, C. (Dept of Anthropology, Univ. of Durham, 43 Old
Elvet, Durham DH1 3HN, UK). Primates, 1993, 34, 105-107.
. .
An all-male band of 15 langurs alarm-barked and mobbed a leopard,
which could be interpreted as altruistic behavior. Unlike the situation in bisexual troops, Hanuman langur males may not be related to
other members of the group, and the evolution of such behavior cannot
necessarily be explained by kin selection.
*Interactions between Lar gibbons and pig-tailed macaques at fruit
sources. Whitington, C. L. (c/o Science Education Ctr, Univ. of Texas,
EDB340, Austin, TX 78712). American Journal of Primatology, 1992,
26, 61-64.
. .
Observations suggest that exploitative and interfering competition
may exist between these species.
*Behavioral asymmetries of psychomotor performance in rhesus monkeys
(Macaca mulatta): A dissociation between hand preference and skill.
Hopkins, W. D., Washburn, D. A., Berke, L., & Williams, M. (D. A. W.,
Dept of Psychology, Language Research Ctr, Georgia State Univ.,
Atlanta, GA 30303). Journal of Comparative Psychology, 1992, 106,
392-397.
. .
Hand preferences were recorded for 35 monkeys, and performance on
joystick manipulation tasks compared, revealing that right-handers
mastered the tasks significantly faster than their left-handed counterparts.
*Object manipulation and the use of tools by infant baboons (Papio
cynocephalus anubis). Westergaard, G. C. (Lab. of Comparative Ethology, NIH Animal Center, Bldg 112, Elmer School Rd, Poolesville, MD
20837). Journal of Comparative Psychology, 1992, 106, 398-403.
. .
Infants in a captive peer group used objects as containers, drinking utensils, and sponges in the context of play. They later used
paper, browse, and other materials as tools to extract sweet liquids
from an apparatus designed to accommodate sponging and probing behavior.
*Parent/offspring conflict and maternal investment in rhesus macaques.
Gomendio, M. (Sub-Dept of Animal Behaviour, Univ. of Cambridge, High
St, Madingley, Cambridge CB3 8AA, UK). Animal Behaviour, 1991, 42,
993-1005.
. .
Differences in amount of suckling between mothers who conceived and
those who did not were not related to differences in demanding behavior by infants, but rather to differences in maternal behavior. Mothers who did conceive were more rejecting. It is suggested that the
less rejecting mothers did not reproduce in an attempt to ensure their
infants' survival.
*Conflict intervention behaviour by adult male macaques: Structural
and functional aspects. Ehardt, C. L. & Bernstein, I. S. (Dept of
Anthropology, Univ. of Georgia, Athens, GA 30602). In A. H. Harcourt &
F. B. M. de Waal (Eds.), Coalitions and Alliances in Humans and Other
Animals (pp. 83-112). New York: Oxford Univ. Press, 1992.
. .
A review of macaque behavior, with particular attention to the
authors' own research with rhesus macaques, shows that female intervention in intragroup conflicts may be more frequent and more effective than intervention by males. For adult females defense of kin may
be particularly important; for adult males, the establishment and
maintenance of relationships and tension reduction may be more
salient.
*Infant-care behavior of non-reproductive helpers in a communal-care
primate, the cotton-top tamarin (Saguinus oedipus). Tardif, S. D.,
Carson, R. L., & Gangaware, B. L. (Dept of Anthropology, Univ. of
Tennessee, So. Stadium Hall, Knoxville, TN 37996). Ethology, 1992,
92, 155-167.
. .
Infant-care behavior patterns of 47 non-reproductive helpers in 18
captive groups were examined. Older helpers carried more than younger
ones, but the difference declined with increasing infant age. Subadults with no previous exposure to infants carried as often as those
with previous experience. Inter-individual variation among helpers was high.
*Sociability in old female Japanese monkeys: Human versus nonhuman
primate aging. Pavelka, M. S. McD. (Dept of Anthropology, Univ. of
Calgary, 2500 University Dr. NW, Calgary, Alberta T2N 1N4 Canada).
American Anthropologist, 1991, 93, 588-598.
. .
Over 20 months, 18 aged and 22 nonaged semi-free ranging female
Japanese monkeys were observed. No relationship was found between age
and sociability.
*Aged monkeys exhibit behavioral deficits indicative of widespread
cerebral dysfunction. Bachevalier, J., Landis, L. S., Walker, L. C.,
Brickson, M., Mishkin, M., Price, D. L., & Cork, L. C. (L. C. C., Neuropathology Lab., 509 Pathology Bldg, Johns Hopkins Univ. School of
Med., 600 N. Wolfe St, Baltimore, MD 21205). Neurobiology of Aging,
1991, 12, 99-111.
. .
Observation of 18 rhesus monkeys aged 3 to 34 years showed behavioral decline beginning in the late teens for certain spatial abilities, while other abilities were not affected until the late 20's.
The finding of correlations between scores of aged animals primarily
within test categories as opposed to across categories suggests that
different animals have different patterns of cerebral involvement.
*Song function in gibbons. Cowlishaw, G. (Dept of Anthropology, Univ.
College London, Gower St, London WC1E 6BT, England). Behaviour,
1992, 121, 131-153.
. .
Results indicate that mated females sing to defend their territories, while mated males sing to repel males in defense of mate and
offspring. Production of coordinated duets by male and female pairs is
associated with encounters and aggression at the territorial boundary,
supporting the hypothesis that duetting advertises pairbond presence
in order to intimidate neighbors.
*Peers as primary attachment sources in yearling rhesus monkeys
(Macaca mulatta). Higley, J. D., Hopkins, W. D., Thompson, W. W.,
Byrne, E. A., Hirsch, R. M., & Suomi, S. J. (NIH Animal Center, P.O.
Box 289, Bldg 112, Poolesville, MD 20837). Developmental Psychology,
1992, 28, 1163-1171.
. .
Both mother-reared (MP) and peer-only reared (PO) monkeys displayed
significantly more intimate contact and significantly less distress
when with their most preferred peer than with other peers, but POs
exhibited more distress than MPs, even when with their most preferred
peer.
*Social responding to mirrors in rhesus monkeys (Macaca mulatta):
Effects of temporary mirror removal. Gallup, G. G., Jr., & Suarez, S.
D. (Dept of Psychology, SUNY, Albany, NY 12222). Journal of Comparative Psychology, 1991, 105, 376-379.
. .
Removing and replacing, after five days, a mirror that rhesus monkeys have become accustomed to produces a short-term reinstatement of
social responding to their own images.
*Behavioral and physiological response of juvenile sooty mangabeys to
reunion with their mothers following a year's absence. Gust, D. A.,
Gordon, T. P., Brodie, A. R., & McClure, H. M. (Yerkes RPRC Field Stn,
2409 Taylor Ln., Lawrenceville, GA 30243). Developmental Psychobiology, 1992, 25, 613-622.
. .
Measurement of cortisol levels and T-cell counts showed that reunion of juvenile mangabeys with their mothers after a year's absence is
an acute stressor for the mothers and a relatively longer-term stressor for the offspring.
*Social separation and reunion affects immune system in juvenile rhesus monkeys. Gordon, T. P., Gust, D. A., Wilson, M. E., Ahmed-Ansari,
A., Brodie, A. R., & McClure, H. M. (Address same as above). Physiology & Behavior, 1992, 51, 467-472.
. .
Removal of juvenile rhesus monkeys from their natal social group to
indoor individual caging resulted in increased basal cortisol secretion and significant decrements in the frequency of lymphoid subpopulations. Significant differences persisted through 11 weeks.
*Immune responses following competitive water tests in two species of
macaques. Boccia, M. L., Laudenslager, M. L., Broussard, C. L., &
Hijazi, A. S. (Dept of Psychiatry, Univ. of Colorado Health Sci. Ctr,
Denver, CO 80262). Brain, Behavior, and Immunity, 1992, 6, 201-
213.
. .
Significant species differences were noted in immune function
between bonnet and pigtail macaques both under unstressed conditions
and following a social stressor. Differences were related to species
differences in agonistic behavior: pigtail monkeys exhibited higher
rates of agonism, competing for access to a water spout, and showed
greatest changes in immune parameters, while bonnet monkeys queued up
by dominance rank, waiting their turns.
Care
*Pickle barrels as enrichment objects for rhesus macaques. Lehman, S.
M. & Lessnau, R. G. (310 1460 West 73rd Ave, Vancouver, BC V6P 3E8,
Canada). Laboratory Animal Science, 1992, 42, 392-397.
. .
Groups of monkeys in outdoor enclosures were compared with and
without barrels for enrichment. Enriched animals accounted for 60.8%
of total affiliative contact, 62.2% of social grooming, and 26% of
agonistic noncontact. Stationary and swiveling barrels were preferred
to unattached ones.
*Opinion: Some thoughts on psychological enrichment. Mahoney, C. J.
(LEMSIP, NYU Med. Center, RR 1, Long Meadow Rd, Tuxedo, NY 10987).
Lab Animal, 1992, 21[5], 27-37.
. .
A review of various aspects of enrichment, emphasizing the importance of the human/animal relationship to well-being.
Development
*Visual habit formation in 3-month-old monkeys (Macaca mulatta):
Reversal of sex difference following neonatal manipulations of androgens. Hagger, C. & Bachevalier, J. (J. B., Univ. of Texas Med. Sch.,
Dept of Neurobiology & Anatomy, 6431 Fannin, Rm 7046, Houston, TX
77225). Behavioural Brain Research, 1991, 45, 57-63.
. .
Ability to perform on a concurrent visual discrimination task with
24-h intertrial intervals develops a few weeks earlier in female than
in male infant monkeys. Orchiectomy hastened performance of males to
the level of intact females. Ovariectomy plus treatment with dihydrotestosterone slowed the learning of females to that of intact males.
*Relative growth of physiques in laboratory-bred cynomolgus monkeys: A longitudinal study during the first 6 years of life. Yoshida, T., Shimizu, T., Cho, F., & Goto, N. (Tsukuba Primate Ctr for Med. Sci., NIH, 1 Hachimandai, Tsukuba-shi, Ibaraki 305, Japan). Experimental Animals, 1993, 42, 435-441. [Japanese, with English abstract, tables, and graphs]
*Neurobehavioral assessment in rhesus monkey neonates (Macaca
mulatta): Developmental changes, behavioral stability, and early
experience. Schneider, M. L. & Suomi, S. J. (2175 Med. Science Center,
1300 University Ave, Madison, WI 53706). Infant Behavior and Development, 1992,
15, 155-177.
. .
Infants reared with a moveable surrogate demonstrated superior
motor maturation in comparison to those reared with only a cloth or
cloth-covered surrogate. Those reared with such a surrogate plus exposure to peers and novel toys had significantly higher orienting
scores, better motor maturity scores, and lower ratings on fearfulness
than all other infants tested. The data illustrate that it is possible
to measure neurobehavioral development in rhesus during the first
month of life using instruments adapted directly from tests used with
human neonates.
*The effect of mild stress during pregnancy on birthweight and neuromotor maturation in rhesus monkey infants (Macaca mulatta). Schneider, M. L. (Address same as above). Infant Behavior and Development,
1992, 15, 389-403.
. .
Infants of mothers exposed to a daily 10-min removal from home cage
and exposure to three unpredictable noise stimuli had lower birth-weights, were delayed in self-feeding, were more distractible, and had
lower scores on a Motor Maturity score compared to offspring from unstressed pregnancies. Low muscle tonus, poor coordination, and slow
response speed characterized the prenatally stressed offspring.
Disease
*Spondyloarthropathy as an Old World phenomenon. Rothschild, B. M. &
Woods, R. J. (Arthritis Ctr of NE Ohio, 5701 Market St, Youngstown, OH
44512). Seminars in Arthritis and Rheumatism, 1992, 21, 306-316.
. .
Limited individual susceptibility to spondyloarthropathy in humans,
Old World monkeys, and lesser apes contrasts with a high frequency of
disease in the great apes.
*Gestational diabetes mellitus in a cynomolgus monkey with group A
streptococcal metritis and hemolytic uremic syndrome. Wagner, J. D.,
Jayo, M. J., Bullock, B. C., & Washburn, S. A. (Comp. Med. Clinical
Res. Center, Bowman Gray Sch. of Med., Medical Center Blvd, Winston-Salem, NC 27157-1040). Journal of Medical Primatology, 1992, 21,
371-374.
. .
Gestational diabetes mellitus was diagnosed retrospectively in a
10-year-old female cynomolgus monkey.
*Human papillomavirus type 13 and pygmy chimpanzee papillomavirus type
1: Comparison of the genome organizations. Van Ranst, M., Fuse, A.,
Fiten, P., Beuken, E., Pfister, H., Burk, R. D., & Opdenakker, G. (G.
O., Rega Inst. for Med. Research, Minderbroedersstr. 10, B-3000 Leuven, Belgium). Virology, 1992, 190, 587-596.
. .
Data indicate that two phylogenetically related papillomaviruses
can elicit similar pathology in different primate host species,
reflecting viral genomic similarities.
*Possible influence of measles virus infection of cynomolgus monkeys
on the outcome of the neurovirulence test for oral poliovirus vaccine.
Contreras, G. & Furesz, J. (J. F., Bureau of Biologics, Virus Bldg,
Tunney's Pasture, Ottawa, P.O. K1A OL2 Canada). Biologicals, 1992,
20, 27-33.
. .
Analysis of 29 tests conducted on type 1 polio vaccines over several years showed a positive correlation between severity of poliomyelitis and the presence of measles serum antibodies in test monkeys.
Tests of over 1500 monkeys used for neurovirulence tests showed a high
proportion of feral animals had measles antibodies, while no animals
bred in a strictly isolated colony did.
*Factors associated with intestinal amyloidosis in pigtailed macaques
(Macaca nemestrina). Ellsworth, L., Farley, S., DiGiacomo, R. F., &
Tsai, C.-C. (R. F. DG., Dept of Comp. Med., School of Med., Univ. of
Washington, Seattle, WA 98195). Laboratory Animal Science, 1992,
42, 352-355.
. .
Records of 74 cases and 74 controls over 5 years indicate that
diarrhea and its various etiologies did not contribute significantly
to intestinal amyloidosis.
*A common-source outbreak of callitrichid hepatitis in captive tamarins and marmosets. Montali, R. J., Scanga, C. A., Pernikoff, D.,
Wessner, D. R., Ward, R., & Holmes, K. V. (Dept of Pathology, Nat.
Zoo, Washington, DC 20008). Journal of Infectious Diseases, 1993,
167, 946-950.
. .
A common-source outbreak of callitrichid hepatitis in golden lion
tamarins and pygmy marmosets at a U.S. zoo resulted from a single
feeding of the primates with newborn mice inapparently infected with
lymphocytic choriomeningitis virus. This virus, prevalent in wild mice
in the U.S., can cause sporadic fatal hepatic disease in primates.
*Immunodeficiency and lymphoproliferative disease in an African green
monkey dually infected with SIV and STLV-I. Traina-Dorge, V., Blanchard, J., Martin, L., & Murphey-Corb, M. (Dept of Microbiology, Tulane
RPRC, 18703 Three Rivers Rd, Covington, LA 70433). AIDS Research and
Human Retroviruses, 1992, 8, 97-100.
. .
Spontaneous dual infections may have contributed to the pathogensis of both agents.
*Arteriopathy in macaques infected with simian immunodeficiency virus.
Chalifoux, L. V., Simon, M. A., Pauley, D. R., MacKey, J. J., Wyand,
M. S., & Ringler, D. J. (D. J. R., NERPRC, Southboro, MA 01772). Laboratory Investigation, 1992, 67, 338-349.
. .
An arteriopathy characterized by intimal and medial thickening and
fibrosis was seen in 19 of 85 rhesus monkeys infected with SIV. Pulmonary arteries were affected in all 19; seven had pulmonary thromboses with varying degrees of organization and recanalization. The
lesion was not uniformly associated with localization of either viral
protein or RNA at the site using immunohistochemistry or in situ
hybridization. Arteriopathy in AIDS may represent a manifestation secondary to primary endothelial injury.
v
*Long-term tolerance and efficacy of 3'-azidothymidine and
3'-fluorothymidine treatment of asymptomatic monkeys infected with
simian immunodeficiency virus. Bottiger, D., Staahle, Li, S.-L., &
Oberg, B. (Dept of Virology, Karolinska Inst., Nat. Bacteriological
Lab., S-105 21 Stockholm, Sweden). Antimicrobial Agents and Chemotherapy, 1992, 36, 1770-1772.
. .
Macaques chronically infected with SIV were treated with zidovudine
or 3'-fluorothymidine. Hematological changes in the treated animals
included macrocytic anemia and leukopenia. At the end of treatment, no
significant changes were observed in general condition, weight, grade
of lymphadenopathy, or T-lymphocyte subset levels between treated and
control animals in either experiment, or in comparison with the condition of the animals at the start of the experiments. Treatment of
asymptomatic SIV-infected macaques could be a useful tool in HIV drug
development, but the model requires more sensitive methods for virus
detection and quantification and more convenient ways of long-term
dosing without restraining animals.
*Penetration of zidovudine and 3'-fluoro-3'- deoxythymidine into the
brain, muscle tissue, and veins in cynomolgus monkeys: Relation to
antiviral action. Ljungdahl-Stahle, E., Guzenda, E., Bottiger, D.,
Wahren, B., Oberg, B., & Stahle, L. (Address same as above) Antimicrobial Agents and Chemotherapy, 1992, 36, 2418-2422.
. .
Concentrations of both drugs in the brain were approximately one-third of that in muscle and veins. The in vivo unbound concentrations
of both drugs in the brain, muscle, and venous blood exceeded those
reported to inhibit HIV replication in vitro. Appearance of p24 antigen in sera of monkeys infected with SIV was significantly delayed by
both compounds.
*Initial therapy for tuberculosis in the era of multidrug resistance: Recommendations of the Advisory Council for the Elimination of Tuberculosis. Morbidity and Mortality Weekly Report, Recommendations and Reports, 1993, 42, No. RR-7.
*Fact or fiction? HIV and polio vaccines. Touchette, N. Journal of
NIH Research, 1992, 4[9], 40-41.
. .
Discussion of the controversial theory that HIV was introduced to
humans by a polio vaccine prepared in African green monkey kidney tissue.
*A carboxyl-terminal fragment of Plasmodium falciparum gp195 expressed by a recombinant baculovirus induces antibodies that completely inhibit parasite growth.
Chang, S. P., Gibson, H. L., Lee-Ng, C. T., Barr, P. J., & Hui, G. S. N. (Dept of Tropical Med. & Med.
Microbiology, 3675 Kilauea Ave, Honolulu, HI 96816). Journal of
Immunology, 1992, 149, 548-555.
. .
Native, conformational epitopes of p42 are critical for the induction of gp195-specific, parasite growth-inhibitory anitbodies, and the BVp42 polypeptide efficiently induces antibodies specific for these native determinants.
*Reversal of Plasmodium falciparum resistance to chloroquine in Panamanian Aotus monkeys. Kyle, D. E., Milhous, W. K., & Rossan, R. N.
(Dept of Immunology & Parasitology, AFRIMS, APO AP 96546). American
Journal of Tropical Medicine and Hygiene, 1993, 48, 126-133.
. .
Combinations of chloroquine plus chlorpromazine or prochlorperazine
confirmed reversal of chloroquine resistance as exhibited by cures
obtained in six monkeys.
*Quantitation of a lentivirus in its natural host: Simian immunodeficiency virus in African green monkeys. Hartung, S., Boller, K., Cichutek, K., Norley, S. G., & Kurth, R. (R. K., Paul-Ehrlich-Inst, Paul-
Ehrlich-Str. 51-59, 6070 Langen 1, Germany). Journal of Virology,
1992, 66, 2143-2149.
. .
One possible explanation of why SIVagm does not induce an AIDS-like
disease in infected African green monkeys could be that infection of
T-cells in vivo does not result in a productive virus infection. Testing this hypothesis, however, showed free infectious virus in the
plasma of infected monkeys.
*Anonymous survey for simian immunodeficiency virus (SIV) seropositivity in SIV-laboratory researchers...United States, 1992. Morbidity
and Mortality Weekly Report, 1992, 41, 814-815.
. .
Three of 472 stored serum samples collected anonymously from lab
workers and animal caretakers were seropositive for HIV-2/SIV; an
additional two samples had antibody to HIV-1. The seropositive persons
cannot be identified or notified.
General
*The raid at Silver Spring. Fraser, C. The New Yorker, April 19, 1993, 66-84.
Instruments & Techniques
*The influence of serum separators on biochemical values in experimental animals. Sunaga, T., Hirata, M., Ichinohe, K., Saito, E., Suzuki,
S., & Tanimoto, Y. (Preclinical Research Labs, Central Inst. for
Experimental Animals, 1433 Nogawa, Miyamae-ku, Kawasaki-shi, Kanagawa
216, Japan). Experimental Animals, 1992, 41, 533-536. (Japanese,
with English abstract).
. .
Detailed tables of results.
*Neuroreceptor imaging with SPECT. Innis, R. B. (V. A. Med. Center,
950 Campbell Ave, West Haven, CT 06516). Journal of Clinical Psychiatry, 1992, 53(suppl.), 29-34.
. .
Single photon emission computed tomography can provide useful measurements of brain receptors and endogenous neurotransmitters.
*Techniques for in utero endoscopic surgery: A new approach for fetal
intervention. Estes, J. M., Szabo, Z., & Harrison, M. R. (Fetal Treatment Program, Univ. of California, P.O. Box 0570, San Francisco, CA
94143-0570). Surgical Endoscopy, 1992, 6, 215-218.
. .
Descriptions of techniques and specialized instruments to facilitate precise work during endoscopic manipulation of a fetus.
*Is preoperative fasting necessary? Popilskis, S., Danilo, P., Acosta,
H., & Kohn, D. (Inst. of Comparative Med., Columbia Univ. College of
Physicans & Surgeons, New York, NY 10032). Journal of Medical Primatology, 1992, 21, 349-352.
. .
Prolonged withholding of oral fluids does not decrease the risk of
aspiration of gastric contents when compared to three hours fasting
time. The addition of an H2 antagonist, cimetidine or ranitidine,
30-40 min before induction provides adequate protection against aspirational pneumonia in baboons.
*A non-invasive technique for the control of intestinal parasites in
rhesus macaques (Macaca mulatta). Bercovitch, F. B., Rodriguez, J.
F., Nieves, P., Ortiz, C. A., Chardon, A., & Kessler, M. J. (Caribbean
PRC, P. O. Box 1053, Sabana Seca, PR 00952). Journal of Medical Primatology, 1992, 21, 363-365.
. .
Percutaneous application of ivermectin was as effective as subcutaneous administration for the control of nematodes in captive macaques.
*Non-surgical uterine flushing for the recovery of preimplantation
embryos in rhesus monkeys: Lack of seasonal infertility. Seshagiri, P.
B., Bridson, W. E., Dierschke, D. J., Eisele, S. G., & Hearn, J. P.
(WRPRC, Univ. of Wisconsin, 1223 Capitol Ct, Madison, WI 53715-1299).
American Journal of Primatology, 1993, 29, 81-91.
. .
Rhesus monkeys, maintained under appropriate environmental conditions, can experience normal fertile menstrual cycles throughout the
summer months, and preimplantation embryos can be recovered year-round. Repeated uterine flushings did not appear to affect reproductive health or the quality or quantity of embryos produced.
*Monitoring of severe head-injured patients with transcranial Doppler
(TCD) ultrasonography. Shigemori, M., Kikuchi, N., Tokutomi, T.,
Ochiai, S., Harada, K., Kikuchi, T., & Kuramoto, S. (Dept of Neurosurgery, Kurume Univ. School of Med., 67, Asahi-machi, Kurume, 830,
Japan). Acta Neurochir, 1992, 55(Suppl), 6-7.
. .
Although many complicated factors influence intracranial blood flow
velocities, a good correlation exists between the flow velocity in the
middle cerebral artery and internal carotid artery flow in Macaca
fuscata with focal compression. Monitoring with TCD provides valuable information.
*A system to acquire and record physiological and behavioral data
remotely from nonhuman primates. Spelman, F. A., Astley, C. A., Golanov, E. V., Cupal, J. J., Henkins, A. R., Fonzo, E., Susor, T. G.,
McMorrow, G., Bowden, D. M., & Smith, O. A. (WRPRC SJ-50, Univ. of
Washington, Seattle, WA 98195). IEEE Transactions on Biomedical Engineering, 1991, 38, 1175-1185.
. .
An integrated system records data simultaneously from two animals
in family groups of five, synchronizing behavioral and physiological
data within 16 ms, either on-line or from recordings. Blood flow,
blood pressure, and heart rate are transmitted from freely behaving
animals by radio telemetry, through a small, light backpack.
*Practice makes predictable: The differential effect of repeated sampling on behavioral and physiological responses in monkeys. Boccia, M.
L., Broussard, C., Scanlan, J., & Laudenslager, M. L. (Dept of Psychiatry, C-268R, Univ. of Colorado Health Sci. Ctr, 4200 East 9th Ave,
Denver, CO 80262). In H. Davis & D. Balfour (Eds.), The Inevitable
Bond: Examining Scientist-Animal Interactions (pp. 153-170). New
York: Cambridge Univ. Press, 1992.
. .
A program to standardize techniques and eliminate the confounding
influences of human contact. However, experience with the researchers'
regime has unmistakable effects on their subjects. The authors ask if
it is possible to minimize the effects of the scientist-animal bond on
research outcome.
Nutrition
*Economic analysis of the effects of caloric alternatives and reinforcer magnitude on "demand" for food in baboons. Foltin, R. W. (Columbia Univ., 722 W. 168th St, Unit 66, New York, NY 10032). Appetite,
1992, 19, 255-271.
. .
Mathematical analysis of demand curves provide a means of comparing
the effects of diverse environmental manipulations on feeding behavior.
*Psyllium husk I: Effect on plasma lipoproteins, cholesterol metabolism, and atherosclerosis in African green monkeys. McCall, M. R.,
Mehta, T., Leathers, C. W., & Foster, D. M. (Lawrence Berkeley Lab., 1
Cyclotron Rd, Bldg 1 #373, Berkeley, CA 94720). American Journal of
Clinical Nutrition, 1992, 56, 376-384.
. .
Animals were fed for 3.5 years on low-cholesterol cellulose (LCC),
high-cholesterol cellulose (HCC), or high-cholesterol psyllium (HCP)
diets. The LCC and HCP groups had significantly lower plasma cholesterol concentrations than did the HCC group. Aortic atherosclerosis
was lowest in the LCC, intermediate in the HCP, and highest in the HCC
group.
*Obesity in free-ranging rhesus macaques. Schwartz, S. M., Kemnitz, J.
W., & Howard, C. F., Jr. (CCM Research, 1230 Park Ave, New York, NY
10128). vInternational Journal of Obesity, 1993, 17, 1-9.
. .
Seven percent of the adult animals surveyed on Cayo Santiago were
defined as obese, based on morphometric indices of adiposity > 2 s.d.
above the mean for adults in the colony. In free-ranging rhesus monkeys, socio-ecological factors and the opportunity for exercise may
mitigate against metabolic and endocrine changes reported for obese
monkeys in laboratory conditions.
*Dietary restriction of adult male rhesus monkeys: Design, methodology, and preliminary findings from the first year of study. Kemnitz,
J. W., Weindruch, R., Roecker, E. B., Crawford, K., Kaufman, P. L., &
Ershler, W. B. (WRPRC, Univ. of Wisconsin, 1223 Capitol Ct, Madison,
WI 53715-1299). Journal of Gerontology, 1993, 48, B17-B26.
. .
Fifteen adult rhesus monkeys were fed 70% of baseline intake levels
previously determined, and compared to 15 controls given free access
to the same food. After one year all animals appear to be in excellent health; experimental animals did not gain weight and have less body fat but show less activity than the controls. These early data indicate that dietary restriction can be safely instituted in adult monkeys, but it is still unknown if it can influence age-sensitive indices in long-lived primates.
Pharmacology & Anesthesia
*Serum concentrations of penicillin, doxycycline, and ciprofloxacin
during prolonged therapy in rhesus monkeys. Kelly, D. J., Chulay, J.
D., Mikesell, P., & Friedlander, A. M. (Rickettsial Diseases Prog.,
Naval Med. Research Inst., Bethesda, MD 20889-5055). Journal of
Infectious Diseases, 1992, 166, 1184-1187.
. .
Concentrations were measured by bioassay in sera of monkeys treated
with these drugs for inhalation anthrax. The antibiotics were well
tolerated. Serum peak and trough concentrations were within the range
usually observed with standard oral doses in humans, and peak concen-
trations in all monkeys exceeded the MICs for 90% of Bacillus anthracis strains.
Reproduction
*Hematological and serum biochemical effects of nursing on the mother
in cynomolgus monkeys. (Macaca fascicularis). Yoshida, T., Ohtoh, K.,
& Cho, F. (Tsukuba Primate Ctr for Med. Science, NIH, 1 Hachimandai,
Tsukuba-shi, Ibaraki 305, Japan). Experimental Animals, 1992, 41,
499-504. [Japanese, with English abstract]
. .
Includes detailed tables in English.
*Convulsions in pregnancy in related gorillas. Thornton, J. G. &
Onwude, J. L. (Dept. of Ob/Gyn, St. James's Univ. Hospital, Leeds, LS9
7TF UK). American Journal of Obstetrics and Gynecology, 1992, 167,
240-241.
. .
Two related gorillas, the second the granddaughter of the first,
had pregnancies complicated by convulsions and edema. They may have
suffered from eclampsia, indicating that this is not an exclusively
human disease and that in gorillas, as in humans, there is a familial
factor.
*Anti-LHRH and anti-pituitary gonadotropin vaccines: Their development
and clinical applications. Thau, R. (Population Council, 1230 York
Ave, New York, NY 10021). Scandinavian Journal of Immunology, 1992,
36, Suppl. 11, 127-130.
. .
Immunizing female primates against LHRH or LH induces infertility,
but also disrupts the menstrual cycle. Immunization against the beta
subunit of chorionic gonadotropin or its fragment prevents pregnancy
without interfering with menstrual cycles or ovulation. Tests of
active immunization against LHRH as a contraceptive method for males
are in progress.
*Primiparous rhesus monkey mothers are more sensitive to the nursing-induced inhibition of LH and ovarian steroid secretion. Wilson, M. E.
(Yerkes PRC of Emory Univ. Field Station, 2409 Taylor Lane, Lawrenceville, GA 30243). Journal of Endocrinology, 1992, 134, 493-503.
. .
Data suggest that parity/age differences in the period of lactational infertility are not due to differences in nursing behavior but
rather to an increased sensitivity to the inhibitory aspects of the
suckling stimulus in adolescent primiparous mothers.
*Effects of management practices on the timing of captive chimpanzee
births. Alford, P. L., Nash, L. T., Fritz, J., & Bowen, J. A. (Univ.
of Texas M. D. Anderson Cancer Ctr, Rt. 2, Box 151-B1, Bastrop, TX
78602). Zoo Biology, 1992, 11, 253-260.
. .
The timing of 153 live, term births of chimpanzees in three institutions with various management practices was examined. In facilities
having marked alterations between weekday and weekend activity, significantly more births occurred Sundays and Mondays. In colonies where
weekend and weekday surveillance and activity were similar, births
were more evenly distributed through the week. Colony managers should
minimize colony activities that are potentially stressful to prepartal
females in order to reduce risks to the fetus that may occur with
delay or disruption of normal parturition.
*Administration of human luteinizing hormone (hLH) to macaques after
follicular development: Further titration of LH surge requirements for
ovulatory changes in primate follicles. Zelinski-Wooten, M. B., Hutchison, J. S., Chandrasekher, Y. A., Wolf, D. P., & Stouffer, R. L.
(R. L. S., ORPRC, 505 NW 185th Ave, Beaverton, OR 97006). Journal of
Clinical Endocrinology and Metabolism, 1992, 75, 502-507.
. .
Novel methods that produce a more nearly normal physiological surge
of LH in nonhuman primates and women during in vitro fertilization
have been attempted recently. This paper reports attempts to further
titrate LH surge requirements for ovulatory changes by administering
monoclonal antibody-purified, urinary hLH to macaques after follicular
development.
*New trends in combined use of gonadotropin-releasing hormone antagonists with gonadotropins or pulsatile gonadotropin-releasing hormone
in ovulation induction and assisted reproductive technologies. Gordon, K., Danforth, D. R., Williams, R. F., & Hodgen, G. D. (D. R. D.,
Ohio State Univ., Dept of Ob/Gyn, Means Hall, Rm 551, 1654 Upham Dr.,
Columbus, OH 43210-1228). Current Opinion in Obstetrics and Gynecology, 1992, 4,
690-696.
. .
The utility and feasibility of employing GnRH antagonists as part
of various ovulation induction regimens has been demonstrated in the
nonhuman primate model.
*Circadian timekeeping during pregnancy: Endogenous phase relationships between maternal plasma hormones and the maternal body temperature rhythm in pregnant rhesus monkeys. Honnebier, M. B. O. M., Jenkins, S. L., & Nathanielsz, P. W. (Dept of Physiology, Cornell Univ.
College of Vet. Med., Ithaca, NY 14853-6401). Endocrinology, 1992,
131, 2051-2058.
. .
Apparently 24-h variations in maternal plasma hormones are endogenously generated and not passively dependent on the light-dark cycle.
The maternal circadian system regulates the 24-h temporal organization
of endogenous plasma hormone variations. It is hypothesized that
rhythms in cortisol, dehydroepiandrosterone sulfate, progesterone, and
estradiol during pregnancy are directly or indirectly governed by the
maternal hypothalamus via the circadian oscillatory output of the
maternal adrenal.
*Maternal rank affects reproductive success of male Barbary macaques
(Macaca sylvanus): Evidence from DNA fingerprinting. Paul, A., Kuester, J., & Arnemann, J. (Inst fur Anthropologie der Univ. Gottingen,
Burgerstr. 50, W-3400 Gottingen, Germany). Behavioral Ecology and
Sociobiology, 1992, 30, 337-341.
. .
High-born males began to reproduce significantly earlier and sired
significantly more infants surviving for at least one year during the
first four years of their reproductive career than low-born males, a
relationship not affected by external conditions such as the level of
intrasexual competition or the number of fertilizable females.
*DNA fingerprinting reveals that infant care by male Barbary macaques
(Macaca sylvanus) is not paternal investment. Paul, A., Kuester, J.,
& Arnemann, J. (Address same as above). Folia Primatologica, 1992,
58, 93-98.
. .
Male Barbary macaques interact extensively and intimately with
infants soon after birth. Although males prefer certain infants over
others in such interactions, DNA fingerprint analysis shows that kinship is neither a primary nor a necessary factor in the "male care"
system.
*Female reproductive cycles and the sexual behavior of moor macaques
(Macaca maurus)in their natural habitat, South Sulawesi, Indonesia.
Matsumura, S. (Primate Res. Inst., Kyoto Univ., Inuyama, Aichi 484,
Japan). Primates, 1993, 34, 99-103.
. .
Observations of a troop of 22 animals indicate an average cycle
length of 36.2 d, with mean duration of swelling 11.9 d. Estimated
duration of gestation in two observed cases was 175 and 176 d, respectively.
Taxonomy
*Identification of chimpanzee subspecies with DNA from hair and
allele-specific probes. Morin, P. A., Moore, J. J., & Woodruff, D. S.
(D. S. W., Dept of Biology, U.C.S.D., La Jolla, CA 92093-0116). Proceedings of the Royal Society of London, B, 1992, 249, 293-297.
. .
A noninvasive method of determining subspecies of Pan troglodytes, which will contribute to colony-level management, recognition of inappropriately matched individuals in comparative studies, and solution of forensic questions involving illegally traded animals.
-------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
* * *
Sochi, August 5, 1993 -- You cannot imagine how turbulent and complicated the life is here. I think you know that we founded a new Institute of Medical Primatology here which belongs to Russian Academy of Medical Sciences. It should be similar to what we had in Sukhumi. Sukhumi (where I am officially still director) is almost dead. Most of the animals died due to severe hunger and cold last winter. Some of the monkeys were killed or stolen. Most of the people left Sukhumi. I think I succeeded in saving the best people and giving them shelter here. Some of them went to Moscow or other cities. Some are still in Sukhumi, being unable to leave it with even a small case. During the last four days it has been rather quiet in Sukhumi and I hope I will be able to go there and help the people, and perhaps collect some scientific material which was left there. Some days ago a ceasefire agreement was signed, but only God knows if it will be honored. There is neither railroad nor air nor highway communications with Sukhumi. Everything is mined. It is also very difficult to run the new Institute because of hyperinflation and shortage of funds. I spend 75% of my time on getting money. We try to carry out research work despite many difficulties. We study tumor viruses (epidemiology, immunology, molecular biology), AIDS, space research, infection, and comparative pathology. The monkey colony has almost 2000 animals--macaques and baboons kept outdoors in open compounds with attached winter houses. Our excellent library is now "abroad" and I do not think it is in order and available to us. Some days ago a lot of shells affected the territory of Sukhumi Institute. Members of the staff were wounded, some severely. But as the Germans say: "Der Mensch strebt so lange er lebt." I wish you every success. Sincerely, as ever, Prof. B. Lapin, Director, Russian Academy of Medical Sciences, Institute of Medical Primatology, 354597 Sochi-Adler, Veseloe 1 [45-12-34-919250; FAX: 862-928730, 8622-920445; e-mail: [email protected]].
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853,
Brown University, Providence, Rhode Island 02912.
Phone: 401-863-2511
FAX: 401-863-1300
Judith_Schrier@brown.edu
ACKNOWLEDGMENTS
The Newsletter is supported by U. S. Public Health
Service Grant RR-00419 from the Comparative Medicine Program,
National Center for Research Resources, N.I.H.
Cover drawing of Japanese "snow" monkeys (Macaca fuscata)
by Anne M. Richardson
Copyright @1993 by Brown University
Editor: Judith E. Schrier, M.Sc.
Associate Editor: James S. Harper, D.V.M.
Consulting Editor: Morris L. Povar, D.V.M.
Copy Editor: Elva Mathiesen, B.A.
Founding Editor: Allan M. Schrier, Ph.D.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
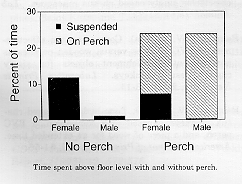{kind=link}