 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate NewsletterVOLUME 33 NUMBER 2 APRIL 1994
Articles and Notes
Evidence for Suppression of Ovulation in Singly-Housed Female Common Marmosets (Callithrix jacchus), by S. Tardif, K. Hyde, & L. Digby...... ....1
All-Male Social Group Formation: Does Cutting Canine Teeth Promote Social Integration? by M. R. Clarke & J. L. Blanchard...... ....5
Results of Providing Swings to Individually Housed Rhesus Monkeys (Macaca mulatta), by S. Dexter & K. Bayne...... ....9
Captive Galago and Bushbaby Diet, by J. Ward...... ....12
Time to Ban Imports? Yet Another Response, by J. R. Held...... ....13
News, Information, and Announcements
Primate Trivia...... ....4
Research and Educational Opportunities...... ....8
Pathology of Lab Animals Course; Earthwatch; Volunteers, Venezuela and California
Correction to the Directory of Graduate Programs in Primatology and Primate Research (1994)...... ....14
News Briefs...... ....15
Comments on Guide Revisions; SIV Isolated in Lab Worker; Indonesia Bans Wild-caught Primate Exports
Award Nominations...... ....16
ASP Conservation Committee; HSUS Russell and Burch Award; SCAW Rowsell Award
Information Requested and Available...... ....17
Manuscripts Wanted; STLV and Chimpanzee Collecting Practices; New Genetic Database
Resources...... ....18
Proyecto Peruano de Primatologia; Purpose-bred Monkeys; Time-Labeled Fetal Tissues
Workshop Announcements...... ....19
Training and Enrichment Workshop; Continuing Education and Training; Animal Welfare Training; Animal Welfare Information Requirements
Grants Available...... ....20
Fulbright Scholar Awards; Postdoctoral and Associateship Programs; Neurologic Integrity in Fetus and Neonate; Academic Research Enhancement Award; Whitehall Foundation Grants; Oral Wound Healing, Tissue Regeneration
Travelers' Health Notes...... ....22
Malaria Info Phone Number; Dengue Fever
Solution to January Puzzle...... ....22
Meeting Announcements...... ....23
Animal Behavior Society; Behavioural Brain Research; Meeting Notes
Editor's Notes...... ....24
Thanks to Our Proofreaders; George Ettlinger; James Bowen
Departments
Address Changes...... ....14
Positions Available...... ....24
Postdoc in Behavioral Medicine; Primate Care Technicians
Recent Books and Articles...... ....25
* * *
Evidence for Suppression of Ovulation in Singly-Housed
Female Common Marmosets (Callithrix jacchus)
Suzette Tardif, Karren Hyde, and Leslie Digby
University of Tennessee, Oak Ridge Associated Universities, and University of California, Davis
Reproductive suppression of subordinate females has been documented in marmoset and tamarin monkeys (family Callitrichidae). In common marmosets (Callithrix jacchus) housed in social groups, all but one female failed to ovulate due to failure of gonadotropic releasing hormone stimulation of lutenizing hormone secretion and increased negative feedback sensitivity to 17beta estradiol (Abbott, 1988; Abbott et al., 1988). Both behavioral and olfactory cues from the dominant female affected the establishment and maintenance of suppression (Abbott et al., 1993). Based on these studies, Abbott and George (1991) cautioned that the "effect of social rank should, therefore, always be considered when the reproductive physiology of female primates living in groups is studied." The results we report here suggest that such effects may also be important in the study of singly-housed females.
In the course of examining the ovulatory cycle of common marmosets, we found that only four of ten females singly housed in the same room ovulated as evidenced by cyclic elevations of urinary estrone conjugates (Hodges & Eastman, 1984). We examined whether this lack of ovarian cyclicity was similar to that observed in social groups of members of the family Callitrichidae. Specifically, this paper addresses the following questions regarding suppression of reproductive function in singly-housed common marmosets: 1) Do nonovulatory (i.e., acyclic) females become cyclic when moved away from the presence of cycling females? 2) Do females return to an acyclic state when returned to the presence of cyclic females? 3) Does number of females per room determine or affect the number of females that are cyclic?
Materials and Methods
Ten adult female common marmosets (C. jacchus) were the subjects of this study. These animals were housed in 4.87 x 3.66 m rooms having an air flow rate of 10 exchanges/hour in 60.9 x 45.7 x 53.3 cm hardware cloth cages. Details of colony management have been published previously (Clapp & Tardif, 1985).
Reproductive state was determined by measuring the levels of estrone conjugates (EC) in marmoset urine with creatinine determination for each sample. Morning urine samples were collected three to five mornings per week on plastic sheets placed in the bottom of the cages, centrifuged to remove contaminating solids, aliquotted, and stored at -70~C until assay. Hodges and Eastman (1984) established that the measurement of urinary estrone metabolites is a reliable method of assessing ovarian function in the common marmoset, if the appropriate metabolite is measured. Urinary ECs were measured by a direct method as described by Hodges and Eastman (1984) using samples diluted 1:2000 in buffer. Tritiated estrone-3-sulfate (SA=60 ci/mmol; New England Nuclear Research Products) was used as a tracer and the antibody was a sheep anti-estrone-3-glucuronide antiserum obtained from University College London. All assays were run with pooled adult female common marmoset urine samples for assay validation and calculation of inter- and intra-assay coefficients of variation. Inter- and intra-assay coefficients of variation were 8.90% and 2.52% respectively. Creatinine determinations were made on each sample to correct for urinary concentrations using a creatinine diagnostic kit (Sigma). Urinary !estrone conjugate levels were expressed as !m!g/mg creatinine (Cr). Females were defined as cyclic if EC concentrations rose above 15 milligr/mg Cr and remained elevated for at least one week (see Figure 2). Females displaying an absence of luteal rise in urinary EC > 15 milligr/mg CR/ml urine for more than one cycle length (28 days; see Hearn, 1983) were defined as acyclic.
X: Housed in original room
y: Housed in alternate room
Animal ID Z: Returned to original room
5000 X X X X X X X X X X X X X X X X X
4983 X X X X X X X X X X X X X X X X X
4921 X X X X X X X X X X X X X X X X X
5001 X X X X X X X X X
4332 X X X X X X X y y
5019 X X X X X X X y y
4976 X X X X X X X y y y Z Z Z Z Z Z Z
4918 X X X X X X X y y y y Z Z Z Z Z Z
4926 X X X X X X X y y y y y Z Z Z Z Z
4925 X X X X X X X y y y y y y y Z Z Z
Phase I II III IV
Months 1-6 7-8 13-16 17-18
Figure 1: Housing of the 10 subjects
used from Phase I through Phase III. This study can be broken down into four phases as shown in Figure 1. These phases are defined by the movement of subjects between rooms. Phase I involved the determination of the reproductive status of ten singly-housed adult C. jacchus females housed in one room (the original room). EC concentrations were determined for 30-60 day periods over a total of 80-200 days during this phase. Phase II involved the removal of six acyclic females from the room for placement into other rooms (the alternate rooms) with a maximum density of four adult female marmosets per room. Of these six females, five displayed no EC concentrations above 15 milligr in the 100 days prior to removal. One female (Figure 2g) displayed one daily concentration exceeding 15 milligr; she was also classified as acyclic as she did not maintain elevated concentrations for at least 10 days. Ovarian cyclicity was assessed over a two-month period, following the transfer of these females to alternate rooms. Phase III involved the staggered transfer of three originally acyclic females back to the original room. Transfer occurred at two-week intervals so that each animal introduced had a separate reentry. Ovarian cyclicity was assessed over this entire period for each of the four females reinstated in the original room, as well as the three originally cyclic females that had remained in the room throughout. During Phase IV, three females of unknown reproductive status were moved into the original room 49 days after the introduction of the last originally suppressed female, in order to replicate the original density of females in the room.
fancy graph
Figure 2: Urinary estrone conjugate concentration (expressed as microgr EC/mg Creatinine) versus days through Phases I and II.
Data presented for: (a) a representative
cyclic female who remained in the original
room; (b) - (g) acyclic females who were
removed from the original room during Phase
II; The vertical line in the figure represents the point at which individuals were
moved from the original room (i.e., the shift
from Phase I to Phase II).
Results and Discussion
Figure 2 illustrates the urinary EC concentration over Phase I and Phase II for one female who was continuously cyclic (1) and the six females who were transferred in Phase II (b-g). During Phase II, with removal of the six females, the remaining four females in the original room continued to cycle. Five out of six suppressed females became cyclic within two weeks (Fig. 2a-c, e). The sixth female was found to be cyclic upon examination seven months later (Fig. 2d). Thus, upon removal from a room with an air-flow rate of ten exchanges/hour containing ten singly-housed C. jacchus females, the six suppressed females became cyclic when placed in a room where the final number of adult C. jacchus females was no more than four.
Another Graphic
Figure 3: Urinary estrone conjugate concentration (microgr/mg/Cr) during Phase III for
the two females who became acyclic. The dotted lines represent the points at which individual females were moved back into the original room: . . . 4976; - - - 4918; -- --
4926; --- --- 4925; - ---- three novel
females.
Prior to Phase III, one of the continually cyclic females (5001) had to be removed from the study, leaving three cyclic females in the original room. Each originally acyclic female reintroduced into the room was cyclic upon her re-entry. The first introduction (female 4976) produced no change in any female's reproductive status (that is, four out of four females were cyclic). After the second introduction (female 4918), 4976 became suppressed as evidenced by her lack of an expected luteal rise in EC > 10 milligr/mg Cr/ml (Figure 3). Female 4976 remained acyclic for the duration of Phase III. The total number of cyclic females in the room at this point was four out of five. The third and fourth introductions (females 4926 and 4925 respectively) produced no further change in any animal's reproductive status, so that six out of seven females in the room were cyclic at that point. Upon introduction of three adult female C. jacchus of unknown endocrine status, female 4926 failed to experience a luteal rise in EC > 10 microgr/mg Cr/ml for the duration of Phase IV (Figure 3). The total number of female C. jacchus in the original room at the end of Phase IV was ten. Of the original females from Phases I and II, five out of seven animals were cyclic and two animals were acyclic. Both of the acyclic females, 4976 and 4926, had been previously suppressed.
Suppression of ovulation in social settings is well-established in callitrichid primates. The present study reports on similar suppression in females housed nonsocially. The initiation of ovarian cyclicity in singly-housed common marmosets after transfer to a different room suggests that their initial suppression was similar to the suppression observed in social groups.
The present study suggests that cues leading to reproductive suppression may act upon females that have only visual and limited olfactory contact. Evidence from Abbott et al. (1993) indicates that both behavioral and olfactory cues play a role in the establishment and maintenance of reproductive suppression in marmosets. Our results suggest that olfactory cues related to suppression are probably carried in the volatile portion of scent marks (Epple et al., 1989), since the effects were observed in the absence of tactile contact. The role of visual cues in the establishment or maintenance of relations between singly-housed females is uncertain. It should be noted that cyclic and acyclic females in this study showed no consistent differences in patterns of scent-marking or dominant/subordinate visual displays during the changes from Phase II to Phase IV (Digby & Tardif, unpublished data).
Density is suggested as a factor in the reproductive suppression of singly-housed female C. jacchus as evidenced by the cyclicity of four out of ten animals in Phase I, and five out of seven animals of known endocrine status in Phase III. These results suggest the value of additional experiments in which density, air flow, previous reproductive status and nontactile interactions among singly-housed females should be addressed.
References
Abbott, D. H. (1988). Natural suppression of fertility. Symposium of the Zoological Society of London, 60, 7-28.
Abbott, D. H., Barrett, J., & George, L. M. (1993). Comparative aspects of the social suppression of reproduction in female marmosets and tamarins. In A. B. Rylands (Ed.), Marmosets and Tamarins: Systematics, Behaviour and Ecology (pp. 152-162). Oxford: Oxford Univ. Press.
Abbott, D. H. & George, L. M. (1991). Reproductive consequences of changing social status in female common marmosets. In H. O. Box (Ed.), Primate Responses to Environmental Change (pp. 295-309). London: Chapman & Hall, Ltd.
Abbott, D. H., Hodges, J. K., & George, L. M. (1988). Social status controls LH secretion and ovulation in female marmoset monkeys (Callithrix jacchus). Journal of Endocrinology, 117, 329-339.
Abbott, D. H., McNeilly, A. S., Lunn, S. F., Hulme, M. J., & Burden, F. J. (1981). Inhibition of ovarian function in subordinate female marmoset monkeys (Callithrix jacchus jacchus). Journal of Reproduction and Fertility, 63, 1-11.
Clapp, N. K. & Tardif, S. D. (1985). Marmoset husbandry and nutrition. Digestive Diseases and Sciences, 30, 17S-23S.
Epple, G., Belcher, A., Greenfield, K. L., Kuderling, I., Nordstrom, K., & Smith, A. B. III (1989). Scent mixtures used as social signals in two primate species: Saguinus fuscicollis and Saguinus o. oedipus. In D. G. Laing et al. (Eds.), Perception of Complex Smells and Tastes (pp. 1-25). Sidney, Australia: Academic Press.
Hearn, J. P. (1983). The common marmoset. In J. P. Hearn (Ed.). Reproduction in New World Primates: New Models in Medical Science (pp. 181-215). Lancaster, PA: MTP Press.
Hodges, J. K., & Eastman, S. A. K. (1984) Monitoring ovarian function in marmosets and tamarins by the measurement of urinary estrogen metabolites. American Journal of Primatology, 6, 187-197.
-------------------------------------------------------------------
First author's address: Dept of Anthropology, Univ. of Tennessee, South Stadium Hall, Knoxville, TN 37996.
This work was supported by NIH grant R01-RR02022 to SDT and ORAU Corporation. The animals were kept at the Marmoset Research Ctr, Oak Ridge, an AAALAC-accredited facility.
* * *
All-Male Social Group Formation:
Does Cutting Canine Teeth Promote Social Integration?
Margaret R. Clarke and James L. Blanchard
Tulane Regional Primate Research Center
Introduction
While cutting the canine teeth of adult male rhesus monkeys (Macaca mulatta) reduces risks to caretakers, our experience at Tulane Primate Center indicates that adult males housed in breeding groups need their canine teeth to defend themselves and to use in threats, which help maintain social control in breeding groups. Since housing animals in social groups makes efficient use of housing space, promotes social compatibility, and meets with Animal Welfare Act requirements, we have all-male groups of rhesus monkeys that have been housed together for up to 4 years. Even though most male-male trauma has occurred after introduction to females (see Bernstein et al, 1974a, 1974b), it was suggested that cutting the males' canine teeth would minimize potential damage when males are housed in all-male groups (McCully, pers. comm., Oct., 1992; McCully et al, 1992). To test this, we formed two comparable groups of adult males, one group with canines cut for the purpose of this study (experimental group), the other with canines intact (control group). For 22 weeks following introduction, we assessed social behavior, success at remaining in the group, and the types of injuries that occurred in the two groups.
Methods
Subjects and Housing: Twenty-six adult male rhesus monkeys were the subjects of this study. They had a mean age of 7.4 years (range 5.5-10.6), and an average weight of 11 kg (range 8-13). All of the animals could be considered "social misfits" as each had a history of being injured and removed from social groups (mean 2.8 removals, range 2-6). Seventy-eight percent of these removals were from all-male groups and the remainder from mixed social groups. Just prior to this study, all males were singly housed.
The outdoor corrals in which the animals were housed during the study were 80 x 80 feet (approximately 1/4 acre), with grass/gravel flooring, 49 upright poles, 8 sets of perches, 8 culverts, 6 movable feeders, 6 Lixits, and multiple wind breaks and shady areas. Twice a day animals were fed commercial monkey diet, which was spread throughout the corral in dry weather and presented in feeders in wet weather. Fruit was given twice weekly, and water was available ad libitum.
Procedure: Subjects were paired by age, weight, and number of removals from all-male or mixed social groups. One male from each pair was assigned to the experimental group (canines to be cut), and the other male from the pair was assigned to the control group (canines intact). The 13 males in the experimental group were treated with prophylactic penicillin P.G. for 3 days, and then sedated with Ketamine/Xylazine (0.2 ml/kg). The mouth was thoroughly washed with Nolvasan, and a diamond burr was used to cut all 4 canine teeth at the level of adjacent teeth. The pulp cavity was opened to a depth of 2-3 mm, packed with calcium hydroxide, and filled with amalgam. Animals were then allowed 5 days to recover from the procedure.
The two groups of males were introduced to the two field corrals at the same time (December 14, 1992). Behavioral observations were started on the day of introduction and continued on a weekly basis until May 25, 1993, when the animals were transferred to two smaller catchpen areas (40' by 40') adjacent to corrals housing breeding groups. Thirty-minute behavioral samples were taken, alternating the corral observed first each observation day (22 hours total). Scan samples of the number of affiliative groups (2 or more animals in contact) were recorded at 5-minute intervals, as well as all occurrences of contact and noncontact aggression during the 30-minute period (Altmann, 1974). There were seven categories of noncontact aggression and 12 categories of contact aggression (Bernstein & Ehardt, 1985). Noncontact aggression included open mouth threats, browflashes, stares, supplants, lunges, and chases. Contact aggression included bites, slashes, hits, and grabs, with each behavior receiving a modifier of mild (light, momentary), hard (hard, but less than 5 seconds), or prolonged (longer than 5 seconds).
The cause of animal removal from the group was categorized as "monkey trauma" or "other." A veterinarian noted the type of trauma (laceration, puncture, crushing bite), and the eventual outcome (euthanasia or treatment) was noted.
Analysis: Behavioral data for the day of introduction were adjusted to a standard 30-minute sample length, and then all data were analyzed on a weekly basis. Mean numbers of affiliative groupings for each session were determined, as well as rates for all 19 categories of agonistic behavior per session. Noncontact aggression was combined into mild responses (animal stationary: open mouth threat/browflash/stare) and more intense responses (animal moving: supplant/lunge/chase). Contact aggression was combined into mild, hard and prolonged for bite/hit/grab. Since there were no observations of "slash," it was eliminated from the analysis. Rates for total contact aggression and total noncontact aggression were also calculated for each group.
Sampling "all occurrences" of aggression met the criteria described by Altmann (1974, p.247), and therefore the data were treated like focal sampling data. Individuals in the groups were matched, and observations of each group were conducted sequentially, so data for each day were treated as paired and were compared using paired t-tests. A one-way analysis of variance with repeated measures was used to compare behaviors from the first six weeks and the rest of the study. We used six weeks since all animals removed for trauma were out of the groups by that time.
Results
Success at Remaining in Group: There were no significant differences in success at group membership between the control and the experimental groups. Two animals (one from each group) were removed due to trauma during the first week. During the rest of the study period, two more control animals were removed (one for trauma) and one experimental animal was removed for "other" reasons. For the duration of the study, only three animals were removed for trauma.
After the animals were transferred to the smaller catchpen area and formal observations ended, one animal from each group was removed within a week for trauma, and two more were removed from each group over the next two months. The proximity of breeding groups introduced uncontrolled variables in addition to the reduction in space, precluding inclusion of these data in the analysis.
Injury Pattern: Injuries to the animal removed for trauma from the cut-canines group were described as "necrotic wounding" and the animal was euthanized. Injuries to the two trauma animals from the intact-canines group were described as "multiple lacerations and scratches involving limbs and body." One of these was found dead; the other was euthanized.
+--------------------+----------+----------+
| | Cut | Intact |
+--------------------+----------+----------+
| Noncontact | | |
| Mild | 2.02 | 0.70 |
| More intense | 6.21 | 3.51 |
| | | |
| Total | 8.24 | 4.21 |
+--------------------+----------+----------+
| Contact | | |
| Mild | 0.31 | 0.14 |
| Hard | 0.21 | 0.05 |
| Prolonged | 0.10 | 0.02 |
| | | |
| Total | 0.62 | 0.21 |
+--------------------+----------+----------+
Table 1: Mean Rate of Aggressive Behaviors Per Session
Affiliation and Aggression: There was no significant difference between the two groups of monkeys in the number of affiliative groupings (sign test, p=.32, n.s.). Means for the different classifications of agonism are in Table 1. Immediately following introduction, contact aggression was higher in the cut-canines group, but only prolonged contact aggression approached significance (t�� =-1.37, p=.09). Total noncontact aggression was greater for the animals with cut canines (t-- =-2.77, p=.01), especially for open mouth threats (t-- =-2.28, p=.03), stare threats (t�� =-1.69, p=.05), and supplants (t�� =-2.44, p=.02). The combined categories (mild, more intense) also differed significantly for the two groups. The amount of total agonism did not differ over time (first 6 weeks vs. the rest of the study). See Table 2.
+--------------+-------------+-------------+
| |First 6 weeks|Rest of Study|
+--------------+-------------+-------------+
| Noncontact | | |
| Cut Canine | 9.63 (10.1) | 7.38 (5.4) |
| Intact | 8.56 (7.0) | 1.54 (1.6) |
| | | |
| Contact | | |
| Cut Canine | 1.50 (2.8) | 0.08 (0.3) |
| Intact | 0.44 (1.2) | 0.08 (0.3) |
+--------------+-------------+-------------+
Table 2: Mean Rates of Total Aggressive Behaviors/Session (standard deviations in parentheses).
Discussion
Cutting the canine teeth of rhesus monkey males housed in an all-male group was not associated with reduced trauma. It was associated with significantly increased noncontact aggression. Following introduction, contact aggression was actually higher in the group with cut canines. While noncontact aggression by the intact-canines animals decreased following initial introduction, it remained at higher levels for animals with cut canines. It may be that males evaluated the possible costs of aggression during the initial introduction (Popp & DeVore, 1979). While it cannot be stated whether males with canines avoided physical confrontations or males without canines provoked physical confrontations, it is clear that there was more aggression in the group with the potential to do the least damage.
From a husbandry viewpoint, the introduction procedure was successful since 21 of the 26 "social misfits" were still in social groups after 5 months. There were no differences in affiliative behavior between the two groups, so social success was likely due to the complexity of housing and social makeup of the groups rather than the condition of the teeth. More animals were removed from the two social groups after transferral to smaller catchpen areas, which supports the interpretation that housing style was more important in promoting social integration than either the presence or absence of canine teeth.
The advantages of cutting canine teeth should be questioned. From a clinical standpoint, the damage caused by animals with cut canines can still be significant, since crushing bites can result in necrosis and a poor prognosis. If males are to be returned to breeding groups, cut canines are a liability, as such males cannot threaten and control aggression effectively. While males on research projects housed with other males may not need teeth for defense, the increased noncontact aggression in the group without canines suggests that intact teeth send an important message about the risks of aggression, a message that apparently reduces overt aggression. In this study, cutting canine teeth did not facilitate social integration, and in fact was associated with increased aggression.
References
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227-267.
Bernstein, I. S. & Ehardt, C. L. (1985). Intragroup agonistic behavior in rhesus monkeys. International Journal of Primatology, 6, 209-226.
Bernstein, I. S., Gordon, T. P. & Rose, R. M. (1974a). Aggression and social controls in rhesus monkey ( Macaca mulatta ) groups revealed in group formation studies. Folia Primatologica, 21, 81-107.
Bernstein, I. S., Gordon, T. P. & Rose, R. M. (1974b). Factors affecting the expression of aggression during introductions to rhesus monkey groups. In R. F. Holloway (Ed.), Primate Aggression, Territoriality and Xenophobia (pp 211-240). New York: Academic Press.
McCully, C. L., Godwin, K. S., Brown, P., Mandrell, T., Bayne, K. A. L., Southers, J., & Poplack, D. G. (1992). A social housing strategy for rhesus monkeys used frequently in research. Contemporary Topics, 31, 33-34.
Popp, J. A. & DeVore, I. (1979). Aggressive competition and social dominance theory: Synopsis. In D. A. Hamburg & E. R. McCown (Eds.), The Great Apes (pp 317-338). Menlo Park, CA: Benjamin/Cummings.
-------------------------------------------------------------------
Authors' address: Tulane RPRC, 18703 Three Rivers Rd, Covington LA 70433.
This research was supported by NIH grant RR00164 to the Tulane RPRC and FDA breeding contract 223-91-1100 (JLB, principal investigator). Tulane Primate Center is fully accredited by AAALAC and adheres to the standards of the 1985 Animal Welfare Act. The authors would like to thank R. M. Daigle, K. Phillippi-Falkenstein, and M. McCullar for technical support, and Dr. E. L. Zucker, Ms. R. M. Daigle, and Ms. J. A. Snyder for comments on the manuscript.
* * *
Results of Providing Swings to Individually
Housed Rhesus Monkeys (Macaca mulatta)
Sandy Dexter and Kathryn Bayne
National Institutes of Health
Swings of a variety of materials and designs have been used for group-housed primates. Some examples are polyvinyl chloride (PVC) piping (Bayne et al., 1989), geometric shapes on chains (O'Neill et al., 1990), suspended milk crates, flexible hoses, and suspended platforms with chains (O'Neill, 1988). The use of swings for individually-housed primates has not, however, been explored systematically in terms of benefits, design, and construction. Several parameters were used here in evaluating swings including: 1) durability and sanitizability; 2) cost-effectiveness for possible wide-scale implementation; 3) simplicity of construction; and 4) potential for the swing design to interfere with ongoing research or husbandry routines.
Materials and Methods
Subjects: The subjects were nine adult rhesus monkeys (Macaca mulatta) of both genders, representing a random selection of the available primate population. They ranged in weight from 3.2 to 6.6 kg, and in age from 5 to 8 years. All had been individually housed for over 3 years in the same animal holding room in 4.3 ft� cages and in accordance with the guidelines established by the Guide for the Care and Use of Laboratory Animals (1985). The animals were on a 12L:12D cycle and were fed NIH Open Formula Monkey Diet at 0730 and 1430 daily. During the course of the study the animals did not receive any food treats, nor did they have any other enrichment devices in their cages.
Experimental Design: Four different swing types were examined during the experimental portion of the study: rope and Nylabone� (ROPE), vine and Nylabone� (VINE), flexible PVC, and Nylabone� (TUBE) (Photo 1), and flexible PVC with 2" diameter rigid PVC pipe (PIPE) (Photo 2). The ROPE, VINE, and TUBE types were wrapped around or inserted through a dog bone toy so the monkey could perch on it while swinging. In the case of the PIPE swing, the rigid PVC served the same purpose. All four types were constructed to hang a consistent 25" from the top of the cage, allowing space at the bottom of the cage for the animal to lie down.
Photo 1: (Left to right) rope and Nylabone", (ROPE) vine and Nylabone" (VINE), flexible PVC and Nylabone" (TUBE) swings.
The ROPE swing was composed of hemp rope measuring 42" in length wrapped around a Nylabone� Knot Bone (9" of rope was wrapped around the bone and secured with a hose clamp). The rope was attached to the top of the cage (8" of it was secured with a hose clamp). In addition to swinging, it was anticipated that the animals would manipulate and groom the rope portion of the swing. Because rope is not easily sanitized, it was replaced every two weeks in accordance with the cage sanitation schedule.
The VINE swing was composed of artificial vine made of a vinyl-coated steel rod manufactured to resemble natural vine (used widely in zoos with natural habitats), and a Nylabone� Knot Bone. A 3" snap hook was attached to a 2" loop of the core steel of the vine that suspended the swing from the cage ceiling. The body of the swing was 19" with a 1" piece of the core steel left uncovered to pass through a drilled Nylabone�, secured with a metal sleeve.
The TUBE swing was constructed using 37" of 3/4" diameter clear flexible PVC wrapped around a Nylabone� Soup Bone and attached to the top of the cage by weaving 6" of PVC through the top of the cage. An additional 6" of PVC were required to wrap around the bone and secure with a hose clamp.
Photo 2: Flexible PVC with rigid PVC pipe (PIPE) swing.
The PIPE swing was fabricated by threading a 40" length of flexible PVC through the rigid PVC pipe and weaving each end of the tubing through the top of the cage (this required 6" of flexible PVC to attach the swing at either side of the cage ceiling).
Data were collected utilizing a barcode recording system that combined a lap-top Tandy 102 computer and an IBM-PS2 to record the frequency and duration of selected behaviors during the observation sessions (Line, 1987). All subjects were habituated to the presence of an observer for one week prior to the start of the study. The duration and frequency of a variety of aberrant behaviors were recorded during both the baseline and experimental phases. During the baseline portion of the study, behavioral data were collected for each animal during nine 30-minute sessions (total 81 sessions for all subjects, or 40.5 hours). Experimental data were collected during three 30-minute sessions for each of the four swing types (totalling 12 sessions per animal for a total of 108 experimental sessions, or 54 hours). This information was then collapsed into four main categories of activity for purposes of statistical analysis: 1) locomotion; 2) stereotypic; 3) self-directed; and 4) cage-directed. During the experimental phase, the frequency and duration of swing-directed activities were also recorded. Swing activities included: 1) manipulating or biting the swing and 2) sitting or swinging on it. All subjects were exposed to each of the four swing types for 3 weeks during the experimental portion of this study.
Statistical Analysis: A Wilcoxon Sign Ranks Test was used to evaluate the subjects for significant differences in behavioral profiles as a result of the presence of the swings. Frequency and duration of baseline behaviors were first compared to behaviors exhibited when each swing type was present. Next, frequency and duration of baseline behaviors were compared to behaviors exhibited over all experimental sessions.
sec
400 Y
Y
Y Y
300 X Y Y
Y X Y X Y
X Y X Y X Y
200 X Y X Y X Y
X Y X Y X Y
X Y X Y X Y
100 X Y X Y X Y
X Y X Y X Y
X Y Y X X Y Y X Y
0 X Y X Y X Y X Y X Y X Y
Locomotion Stereotypic Self Total Cage Groom
Abnormal
Figure 1: Mean duration of recorded behaviors of sessions for all subjects by experimental condition.
Results
Results showed no significant reduction (p> .05) in abnormal behaviors during the experimental phase compared to the baseline con- dition (Figure 1). Based on these findings, no further analyses using multivariate analysis of variance were performed. All of the swing types were manipulated either orally or digitally for a mean of 4% of experimental sessions with the TUBE swing eliciting the most interaction. However, the PIPE was the only design actually used for swinging (0.4% of experimental sessions or 6 occasions; see Figure 2).
X: Manipulate Y: Swing
(sec)
80
X
X X X
60 X X X X
X X X X
X X X X
40 X X X X
X X X X
X X X X
20 X Y X X X
X Y X X X
X Y X X X
0 X Y X X X
PVC Pipe Vine Rope Flex. PVC
Figure 2: Mean duration of swing use per session for all subjects.
Discussion
In prior studies of enrichment techniques, an enrichment device or technique was deemed effective by our laboratory if the presence of that device resulted in a significant reduction in specific abnormal behaviors (e.g., Bayne et al., 1991). According to this criterion, swings are not effective enrichment devices. No significant reduction in atypical behaviors was observed following the introduction of the swing. Even though all the swing types could be removed, the presence of a swing in the cage presented practical and safety problems. The swing interfered with the squeeze apparatus during restraint of an animal. This interference was not improved by detaching the swing and letting it drop to the cage floor. Furthermore, the process of detachment could easily place a staff member at risk of being wounded by the monkey, since unlatching the swing required close proximity to the cage bars. The swing design could be improved by imitating designs for perches that collapse or move through cage bars (Reinhardt & Pape, 1991; Schmidt et al., 1989; Watson, 1991).
It is possible that individually housed younger animals might be more inclined to utilize a swing. For example, smaller animals would have more room in a cage for swinging and thus might be more inclined to use such a device. Certainly, O'Neill (1988) has documented a greater use of three-dimensional space by younger rhesus monkeys than by older ones. It has been noted in our facility that group-housed older animals tend to utilize the cage furniture that is secured (Bayne, personal observation). Previous work by Kopecky and Reinhardt (1991) showed that older rhesus had a clear preference for perches over swings when animals were exposed to both forms simultaneously; however, a clear avoidance of swings by individually-housed older animals could not be determined from their study.
As the search for new ways to enrich the lives of captive primates continues, it becomes increasingly clear that strategies must be investigated thoroughly before widescale implementation. Enrichment for nonhuman primates can be costly, and providing items that are not effective in reducing abnormal behaviors or that occupy an insignificant portion of the animals' daily activities should not be given priority in an enrichment program. Since research has shown varying degrees of success in reducing behavioral pathology using the same enrichment device at different facilities (Bayne et al., 1993), a sound strategy for evaluating an enrichment technique would entail providing the device to a small number of animals prior to full-scale implementation.
References
Bayne, K., Dexter, S., Hurst, J., Strange, G., & Hill, E. (1993). Kong� toys for laboratory primates: Are they really an enrichment or just fomites? Laboratory Animal Science, 43, 78-85.
Bayne, K., Mainzer, H., Dexter, S., Campbell, G., Yamada, F., & Suomi, S. (1991). The reduction of abnormal behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology, 23, 23-35.
Bayne, K., Suomi, S., & Brown, B. (1989). A new monkey swing. Labora- tory Primate Newsletter, 28[4], 16-17.
Guide for the Care and Use of Laboratory Animals (1985). NIH publication No. 85-23. Revised.
Kopecky, J. & Reinhardt, V. (1991). Comparing the effectiveness of PVC swings versus PVC perches as environmental enrichment objects for caged female rhesus macaques (Macaca mulatta). Laboratory Primate Newsletter, 30[2], 5-6.
Line, S. W. (1987). Portable computer/bar code system for recording behavioral observations. Animal Behavior Society Meeting, Williams, MA.
O'Neill, P. (1988). Developing effective social and environment enrichment strategies for macaques in captive groups. Lab Animal , 17[4], 23-36.
O'Neill, P. L., Novicky, P., & George, E. (1990). Preliminary evaluation of Prima-Hedron play structures for non-human primates. Lab Animal, 19[5], 40-41.
Reinhardt, V. & Pape, R. (1991). An alternative method for primate perch installation. Lab Animal, 20[8], 47-48.
Schmidt, E. M., Dold, G. M. & McIntosh, J. S. (1989). A perch for primate squeeze cages. Laboratory Animal Science, 39, 166-167.
USDA, APHIS (1991). Animal welfare, standards, final rule. Federal Register, 56, No. 32, 9CFR, Part 3.
Watson, D. S. B. (1991). A built-in perch for primate squeeze cages. Laboratory Animal Science, 41, 378-379.
-------------------------------------------------------------------
Authors' address: NIH, NCRR, Vet. Resources Program, Behavior and Nutrition Unit, 9000 Rockville Pike, Bldg 14D, Rm 313, Bethesda, MD 20892.
* * *
Captive Galago and Bushbaby Diet
Jeannette Ward
Memphis State University
The behavioral research laboratories at Memphis State University have housed colonies of galagos since 1970. For the first 15 years we had wild-born Galago senegalensis and G. moholi. During most of this period the galagos were fed Purina cat chow, canned applesauce and/or a variety of prepared baby foods including fruits, vegetables, cereals with and without fruit, and meats. This diet was supplemented with mealworms (Tenebrio larvae) for all animals and yogurt for pregnant and nursing mothers. They seemed to thrive on this diet with many animals surviving for 9-10 years.
At present, we have some captive-born G. moholi and a large breeding group of bushbabies (Otolemur garnettii), most of which are also captive-born. Since 1988 we have fed a diet, recommended by the Duke University Primate Center, consisting of high protein primate chow, a variety of fresh fruit and vegetables, daily yogurt for all G. moholi, and yogurt for pregnant and nursing O. garnettii. Both the bushbabies and galagos eagerly consume mealworms, crickets, and roaches, but we supplement only with mealworms because hard insect body parts have been known to puncture soft tissues and can be particularly dangerous for the galagos. Mealworms in several sizes may be purchased from Grubbco [1-800-222-3563] or a mealworm colony can be easily established on a substrate of bran meal and Irish potatoes.
Both bushbabies and galagos are quite catholic in their food acceptance. They have occasionally been offered a variety of human foods and have consumed all items readily. Anything sweet is strongly preferred, as are sources of protein. We were surprised to discover that every one of more than 70 bushbabies excitedly grabbed and immediately consumed fresh-killed "feeder goldfish" on first presentation. Only 8 of these bushbabies were wild-born; none of the others had ever seen a fish before. We subsequently found that 19 of 38 bushbabies would attempt to capture live fish in water (Watson, et al., in press). For captive bushbabies, fishing might be a useful dietary supplement as well as a behavioral enrichment. We have not offered fish to galagos.
Our galago diets are also supplemented with gum arabic (also known as gum acacia, used as an ice cream binder). It is relatively inexpensive when purchased in quantity as a powder. Mixed with water, it becomes a thick gummy mass, which we place on tongue depressors or ice cream sticks. The galagos spend much time licking it off the sticks. Other methods of presenting the gum, as well as information about the dietary value of gums, may be found in a report by Kelly (1993).
References
Kelly, K. (1993). Environmental enrichment for captive wildlife through the simulation of gum-feeding. Animal Welfare Information Center Newsletter, 4[3], 1-10.
Watson, S. L., Schiff, M., & Ward, J. P. (in press). Effects of Modeling and Lineage on Fishing Behavior in the Small-eared Bushbaby Otolemur garnettii. International Journal of Primatology.
-------------------------------------------------------------------
Author's address: Dept of Psychology, Memphis State University, Memphis, TN 38152.
* * *
Time to Ban Imports? Yet Another Response
Joe R. Held
It is always good to hear from and about old friends; thus I enjoyed reading the notes on the issue of primate imports by Hobbs, Kingston, Baulu, and Stanley in recent issues of this publication. They remind me of the old adage "The more things change, the more they remain the same." I sincerely hope that those of us who truly care about primates and their conservation are not getting into a situation in which we'll lose sight of the great strides made worldwide in improving human health and well-being, thanks to the judicious use of nonhuman primates in biomedical and behavioral research. Also I hope that we don't become so busy bickering among ourselves that we expend our energies dealing with questionable and divisive controversies and overlook the need for working together to find logical, clear-cut solutions to the many problems concerning the complex issues of primate supply and use. All four of these individuals are genuinely interested and active in primate conservation, and have made some very valid points. But they have in some ways neglected to deal with a basic issue now facing us in some countries: a coalition between antiscience and animal protection groups that is attempting, through any means possible, to stop the use of animals in biomedical and behavioral research.
Hobbs especially surprised me with his negative remarks regarding the World Health Organization. I was surprised because, as I recall, I first met Keith many years ago when we were both serving on a WHO committee set up to provide advice on better utilization and conservation of African primates. WHO, like any large bureaucracy, is imperfect, but it is the best the world has for its type of mission. Those of us concerned with primate availability and conservation need all of the help we can get and, contrary to the implications in Hobbs' letter, the PAHO Peruvian Primate Project has made many important contributions to learning more about New World primate populations and how to conserve them. He is right that it has not been a commercially viable operation, but it was not intended to be.
Those interested in this subject would be well advised to read an article by Bowden and Smith, "Conservationally sound assurance of primate supply and diversity" (ILAR News , 1992, 34[4], 53-56). This article gives some interesting background information on the history of primate utilization and supply, much of which is germane to the current debate.
It seems to me that what the world needs now is a consortium of governments, international organizations, academic institutions, commercial enterprises, and individuals who could and would contribute to a revolving fund for environmental protection and the worldwide conservation of natural resources, including primates and various plant species that have a potential for future medicinal value. The fund could be administered under the aegis of an existing international body such as WHO, with guidance from a "Board of Directors" made up of scientifically competent individuals from countries that had made appropriate donations to the fund, plus additional scientifically competent individuals from those countries having such resources, and which would be willing to allow their development. Hopefully various countries could also be encouraged to modify their tax laws so that donations to the fund could be tax-deductible to encourage greater monetary contributions, especially from the pharmaceutical industry and others likely to reap future benefits, as well as from interested individuals. Profits from the sale of primates or plants from the participating source countries would be added to the fund on a regular basis.
In this way there might be some hope of preserving these valuable resources for the future. In 1978 the U.S. Government Interagency Primate Steering Committee published a "National Primate Plan," which contained some arguments for supporting primate conservation activities in source countries. I submit that those same arguments are still valid today, and can be used to defend the position that imports should continue to be permitted. These arguments are:
It also needs to be recognized that currently existing positive activities in places such as Peru, Mauritius, Indonesia, the Philippines, Barbados, other nations in the Caribbean and others in various parts of the world will need to have a cash flow if they are to continue making their important contributions. Banning imports would be counterproductive in that it would stop that cash flow, and eventually sound the death knell for those programs.
-------------------------------------------------------------------
Author's address: 1300 Crystal Dr., Apt. 505, Arlington, VA 20222.
* * *
A reader asks: "Why is the head (chair) of the Canadian Church (and many others) called a 'Primate'? Was that a term created de novo for the modern Anglican communion, or does it have some history?"
Simon Kershaw answers: "It certainly wasn't created in modern times. The Archbishops of Canterbury and York 'agreed' on the titles 'Primate of All England' and 'Primate of England' (or rather their Latin equivalents, respectively) in the reign of William I (of 1066 and all that fame).
"This was the end of a long dispute between the two sees for primacy, and York was forced to concede. The document recording this still exists with the signatures of the two archbishops ('ego concedo,' writes York), several bishops as witnesses, and the marks of the illiterate William and his queen.
"The monkeys in the zoo would be impressed by the squabbling between York and Canterbury for primacy. On one occasion Canterbury, arriving first, sat in the position of honour. When York arrived he sat himself down on Canterbury's lap. There was comment at the time of York behaving like a baby sitting on another's lap."
-------------------------------------------------------------------
Mr. Kershaw's address: Smallworld Systems Ltd, Brunswick House,
61-69 Newmarket Rd, Cambridge, CB5 8EG, England.
This material appeared on the electronic mailing list Anglican,
and is reproduced with permission.
* * *
Research and Educational Opportunities
Pathology of Lab Animals Course
The Armed Forces Institute of Pathology and the American Registry of Pathology are co-sponsoring a continuing education course entitled, "Pathology of Laboratory Animals," May 31-June 3, 1994 at the Uniformed Services University of Health Sciences, Bethesda, MD. This course is designed to teach attendees how to describe both gross and microscopic lesions in a variety of major organs, with lectures on interpretation and description of electron micrographs as well.
For application forms and programs, contact AFIP, Washington, DC 20306-6000 [301-427-5231; FAX: 301-427-5001]. Tuition fee is $300 before May 2, $350 thereafter. Registration fee for active duty military, Dept of Defense civilians, fulltime permanent Dept of Veterans Affairs employees (not residents or fellows), and commissioned officers of the PHS with authorized approval is only $100.
Earthwatch
Earthwatch provides volunteers and funds to scholars doing fieldwork. Among the projects (and scholars) in their 1994 catalog are studies of the Northwestern Peru Biosphere Reserve (A. G. Cook, F. Encarnacion, & J. Flinders), the Brazilian rainforest (A. Keuroghlian), red howler monkeys in Venezuela (G. Agoramoorthy), toque macaques in Sri Lanka (W. Dittus), and Kenyan wildlife (D. Harper & P. Hickley). Volunteers, who pay their own transportation expenses plus a share of the costs of the expedition, learn to participate in fieldwork. Scientists may submit a two-page outline of their research subject and design, dates, budget, and plans for the use of volunteers to the Center for Field Research, 680 Mt Auburn St., Box 403, Watertown, MA 02272 [617-926-8200; FAX: 617-926-8532]. Volunteers apply to the same address.
Volunteer Field Workers, Venezuela
Volunteer field assistants are needed for an on-going field study of red howler monkey (Alouatta seniculus) demography and social behavior in Venezuela. Requirements: good physical condition, interest in watching monkeys, ability to work long hours in harsh field conditions. A working knowledge of Spanish, ability to drive a 4WD jeep, and previous field experience will be assets. Volunteers must pay for their own air travel, but living expenses may be provided depending on availability of funds. Send letter of interest, CV and names of three references to: Dr. G. Agoramoorthy, c/o Prof. Minna J. Hsu, Dept of Biology, National Sun Yat-Sen Univ., Kaohsiung 804, Taiwan [FAX: 886-7-5319333; e-mail: hsumin@cc. nsysu. edu. tw]. -- From primate-talk.
Volunteers, California
The International Center for Gibbon Studies (ICGS), a nonprofit organization located in Santa Clarita, CA, needs full-time volunteers during the non-summer months--September to May--to assist in behavioral observations of captive gibbons ("lesser apes"). Other duties include feeding, general maintenance, and data entry. Hard work, honesty, and a love for animals are required.
ICGS is the only facility in the world devoted exclusively to the preservation, study and propagation of gibbons. We currently house 27 gibbons, the second largest group of gibbons in North America. Personal rewards include the gratification of contributing to the well-being of the ICGS gibbons and learning more about these highly endangered apes. For further information, contact ICGS, PO Box 800249, Santa Clarita, CA 91380 [801-296-2737; FAX: 801-296-1237]. -- Posted to primate-talk by Alan Mootnick, Facility Director.
* * *
Correction to the Directory of Graduate Programs
in Primatology and Primate Research (1994)
GEORGIA
*Georgia State University, Language Research Center, Dept. of Psychology
PROGRAM DESCRIPTION: Biobehavioral and cognitive studies of nonhuman
primates.
FACULTY AND THEIR SPECIALTIES: Sue Savage-Rumbaugh (primate cognition, biopsychology, primatology); Duane M. Rumbaugh (project director); Rose Sevcik (developmental comparative psychology); Shelly
Williams (learning and communication); David Washburn (research psychology).
FOR FURTHER INFORMATION: Language Research Center, Georgia State University, University Plaza, Atlanta, GA 30303-3083.
* * *
Kathryn Bayne, Associate Director for Accreditation, AAALAC, 11300 Rockville Pike, Suite 1211, Rockville, MD 20852-3035 (as of May 2).
Rita U. Bellanca, 1549 NE 177th St. Apt 301, Seattle, WA 98155-5268.
William C. Cole, 40 Douglass Rd, Lansdale, PA 19446-1449.
Jess F. Deegan II, Cal. State Univ., Bakersfield, CA 93311-1099.
Jeff Edwards, Texas Veterinary Med. Diagnostic Lab., Drawer 3040, College Station, TX 77841.
Kenneth C. Gold, Lincoln Park Zoo Gorilla Project, 2200 N. Cannon Dr., Chicago, IL 60614.
Robert M. Letscher, 121 Pomona Rd, Pomona, NY 10970.
Preston A. Marx, AIDS Animal Model Lab., RR1 Longmeadow Rd, Tuxedo, NY 10987-9801.
Dennis R. Rasmussen, Florida State Univ., Panama Canal Branch, PSC #02, Box 2663, APO AA 34002.
Rosalind Ribnick, 1505 P St, Eureka, CA 95501-2545.
* * *
Comments on Guide Revisions
The committee appointed to revise the Guide for the Care and Use of Laboratory Animals by the National Research Council's Commission on Life Sciences Institute for Laboratory Animal Resources (ILAR) heard testimony at three meetings in Washington DC, San Francisco, and St. Louis this winter. Written comments may be sent to the committee, c/o Thomas Wolfle, ILAR, 2101 Constitution Ave NW, Washington, DC 20418, until May 30, but the committee would appreciate receiving comments as soon as possible.
SIV Isolated in Lab Worker
For the first time, medical researchers have isolated simian immunodeficiency virus (SIV) in the blood of a laboratory worker who was exposed to an infected animal. The findings, reported in the New England Journal of Medicine (1994, 330, 172-177) confirm that "the risk of human infection with SIV is no longer merely hypothetical," according to the research team led by Dr. Rima Khabbaz of the CDC. "The evidence of infection in our subject includes seroconversion, persistent seropositivity after the presumed exposure, increasing antibody titers with identification of seroreactivity to new viral gene products over time, isolation of SIV, and molecular evidence of an identical SIV sequence in DNA amplified at other times. Our molecular data show that the infecting virus is SIV and not HIV-2 and that this virus...is closely related to the SIV strain with which the researcher primarily worked.
"The SIV-infected researcher remains asymptomatic with no clinical or laboratory evidence of immunodeficiency 3.5 years after seroconversion in April, 1990...The clinical outcome of SIV infection in a human is unknown. It might be expected to be similar to that of its closest human counterpart, HIV-2, which is believed to be less pathogenic than HIV-1. The small viral burden and the inability to recover the virus consistently by culture or polymerase chain reaction may be indicative of a good prognosis...Preliminary data show a premature truncation in the nef gene of the SIV isolated from the researcher (but not in that of the SIV primarily worked with in his laboratory). This deletion might explain the small viral burden in our subject, which is consistent with observations in monkeys inoculated with SIV deleted in the nef gene.
In an accompanying editorial, "Simian immunodeficiency virus in people," (pp. 209-210), the editors discuss the similarities and differences in HIV-1, HIV-2, and SIVs and speculate that understanding these differences may lead to effective vaccines and drugs for AIDS.
Indonesia Bans Wild-caught Primate Exports
A Directive issued by Indonesia's Ministry of Forestry bans the export of wild-caught long-tailed and short-tailed macaques as of April 1, 1994. The Minister wrote, "the long-tailed macaque and the short-tailed macaque, wild animals which have been exploited by human beings for a long time without having been protected by law, are showing signs of decline in the wild." Indonesia exports about 10,000 pri- mates each year, primarily long-tailed macaques (Macaca fascicularis), but also pig-tailed macaques (M. nemestrina). Indonesia's exports make up about 30 percent of the worldwide primate trade. The U.S., the United Kingdom, and Japan are the leading importers of primates. The U.S. also provides imported animals for researchers in other countries following screening and testing in the U.S.
The primate trade community recognizes the problems associated with the trade in wild-caught primates and will be able to supply researchers' demand for primates with captive-bred animals. One primate breeder and exporter told the HSUS/HSI that there is currently a glut of captive-bred primates available for research. Until now the cost of purpose-bred primates has been higher than that of wild-caught animals. The captive, in-country breeding approach is being pursued for conservation of species, ensuring humane care and conditions for the animals, compliance with both international treaties governing exports and strict CDC import requirements regarding animal health, as well as for the economic interests of the supplying country. It is believed that these objectives, as well as the health and quality of purposebred animals, will justify any initial price differential. --- From a Humane Society of the United States news release and NABR Update, February 25, 1994.
* * *
ASP Conservation Committee
The Conservation Committee of the American Society of Primatologists is seeking nominations for ASP Conservation Awards and Grants. An award nomination is basically a letter of recommendation.
Subscription Award: provides the American Journal of Primatology to worthy individuals in source countries who otherwise would have little access to the scientific literature on nonhuman primates. Preference is given to individuals who will make the AJP available in a central place for colleagues to use. A nominating letter should describe the nominee, his or her primate-related activities, and why the subscription is needed.
Conservation Award ($500) : provides recognition and financial support for students and young investigators (not more than five years since terminal degree) from source countries. Nominators should provide the name, title, and full mailing address of a nominee, and a statement about the nominee's qualifications for the award and contribution to primate conservation.
Senior Biology and Conservation Award ($500 Honorarium): recognizes an individual without an advanced degree who has made a substantial contribution to conservation or related aspects of primatology a long period (five years or more). Nominees may be animal caretakers, research technicians, administrators, etc. Nominations should include a letter of support detailing the nominee's qualifications and contributions to primate biology and conservation.
Conservation Small Grant ($500): for conservation research or education, or other projects. Primatologists working in source countries are urged to apply or to help someone from a source country to do so. Grant proposals must be typed in English, should not exceed 2000 words, and should include a brief budget page. Recipients must agree that a brief progress report will be made, in a form suitable for publication in the ASP Bulletin , within 12 months of the award.
Arrangements are often made to have awards presented at public ceremonies by senior officials or U.S. Ambassadors. For an account of one such presentation, see the last issue of this Newsletter (33[1], 22). The deadline for submission of nominations and grant proposals is June 20, 1994. They should be sent to Ramon J. Rhine, Psychology Dept, Univ. of California, Riverside, CA 92521.
HSUS Russell and Burch Award
The Russell and Burch Award is given annually by the Humane Society of the United States to a scientist who has made an outstanding contribution toward the advancement of alternative methods in the areas of biomedical research, testing, or higher education. Alternative methods are those that can replace or reduce the use of animals in specific procedures, or refine procedures so that animals experience less pain or suffering. The award, which carries a $5,000 prize, is named in honor of British scientists who first articulated the "3Rs" approach of replacement, reduction, and refinement. The Award is a means of recognizing the important role that scientists themselves can play in advancing the welfare of animals in laboratories.
Send nominations by June 1, 1994, to Philip Mendoza, Laboratory Animal Programs, HSUS, 2100 L St, NW, Washington, DC 20037 [301-258-3042; FAX: 301-258-3082; e-mail: 76702,[email protected]]. Nominators should submit a letter explaining the nominee's suitability for the award and arrange to have supporting documents (e.g., relevant publications) forwarded.
SCAW Rowsell Award
The Scientists Center for Animal Welfare (SCAW) has established an award to honor Dr. Harry C. Rowsell, who has made major contributions in promoting animal welfare nationally and internationally. The first award was presented to Dr. Rowsell himself. Future awards will be given to persons who are known for commitment to fostering the dual goals of good science and the humane treatment of animals. To nominate an individual for the 1994 award, submit a description of the work you think meets these goals on one single-spaced page. Additional materials such as a curriculum vitae and project descriptions may be included. Send all materials to SCAW, Attn: Harry C. Rowsell Award, 4805 St. Elmo Ave, Bethesda, MD 20814, by July 15, 1994.
* * *
Information Requested and Available
Manuscripts Wanted
The Journal of Zoo and Wildlife Medicine has scheduled a special issue on nonhuman primate medicine for March 1995. Manuscripts focusing on health-related issues of nonhuman primates are being solicited. Deadlines are important! Submissions are due by June 1994. For more information, contact Joanne Paul-Murphy DVM, School of Veterinary Med., Univ. of Wisconsin, 2015 Linden Dr. West, Madison, WI 53706 [608-265-2608; e-mail: [email protected]].
STLV and Chimpanzee Collecting Practices
The STLV retrovirus has been isolated in both free-ranging and captive Old World monkeys from Africa and Asia. Nucleotide sequences of these isolates have shown a close relationship between STLV strains from a given region and human HTLV isolates taken from the same region. Multiple occurrences of interspecies transmission from monkey to man have been offered as an explanation. Transmission of primate T-cell lymphotropic viruses (PTLVs) from man to monkey must also be considered, however. It has been reported that it was once "common practice for animal catchers and dealers to inoculate newly-captured chimpanzees with pooled human blood for protection against human disease" (A. J. Zuckerman et al, Lancet, 1978, no. 8091, 652-654). If this practice was common in Africa and Asia, it might have occasioned the transmission of PTLVs from man to monkey on a significant scale. Although this would most directly affect captive primates and their progeny, it is also worth considering whether any of these inoculated animals could have been released or allowed to range freely, thereby infecting wild primates. This question is also relevant to current thinking on SIV and HIV. George Corey is searching for information on the practice of inoculating monkeys with human blood, as well as any other information on trapping and holding practices of the early 20th century that would be relevant to this question. Please contact Dr. Corey at The Duluth Clinic, 4886 Miller Trunk Hwy, Duluth, MN 55803 [218-727-8119; FAX: 218-722-8160].
New Genetic Database
Attention Geneticists, or anyone using genetic markers in primates: Phil Morin has started a new genetic database on the Gopher server system to facilitate communication among people using Simple Sequence Repeat (SSR) or Microsatellite loci for primate population genetic, paternity, or genome-mapping studies. This server will post information on primers that have been published or developed for human and non-human primate PCR amplification, and information on conditions for amplification on various primate species. He plans to update it on the 20th of each month.
If you have been working with SSR's on primates, you can get a copy of the form for submitting information and instructions for accessing the database by contacting Phillip A. Morin, Dept of Anthropology, Univ. of California, Davis, CA 95616-8522 [916-752-5598; FAX: 916-752-8885; email: [email protected]].
You can access the U.C. Davis Gopher directly through any Internet connection by getting into your account, then typing gopher gopher.ucdavis.edu Once in the UC Davis Gopher, go to the folder entitled research support. Inside of it will be the Primate Research Center folder, then the Primate Genetic Markers folder. Once you are connected, you can open and scan data files. If you find information you would like to copy to your own machine, you can either e-mail it to yourself or save it as a file in your account. If you use a Gopher access program, such as TurboGopher for the Macintosh, you have the option to save whatever document you are looking at as a text file on your terminal, which can subsequently be opened by whatever word processing program you use. As we go to press, there are three files in the PGM folder: Morin's data for chimpanzee SSR loci, his data for Rhesus SSR loci, and a copy of the information and data submission form.
* * *
Proyecto Peruano de Primatologia
The wild Peruvian nonhuman primate population is a valuable natural resource that, in recent years, has been threatened by expanding human populations destroying their habitats, uncontrolled trade, and lack of national and international supervision of animal capture and transport. The Peruvian Primatology Project was created by environmentally concerned organizations, including the Peruvian Ministries of Agriculture and Public Health, San Marcos University, and the Peruvian Amazonia Research Institute, in cooperation with PAHO/WHO, to halt the depletion of Peru's wild primate population.
Since its inception in 1972, the Project has * founded a wildlife census program for the 32 indigenous nonhuman primate species; * established The Center for Primate Reproduction and Conservation in Iquitos; * established three natural island habitats to serve as cen- ters for repopulation trials and areas for the study of natural breeding and behavior; * served as a source of community employment and support for schools, health care, and agricultural training programs; * encouraged awareness of the need for primate conservation and population management within the community.
Two distinct primate populations are maintained: * Free roaming natural populations in national parks and on islands, serving as subjects for ecological, reproductive, and ethological field studies; * Colony-reared populations raised under controlled conditions to ensure a continuous supply of well-socialized, high quality animals of known health status. Nonendangered and nonthreatened species only (e.g. Aotus nancymai, A. vociferans, Saimiri boliviensis peruviensis, S. sciureus macrodon, Cebus apella, Cebuella pygmae, and Saguinus spp.) may be transferred to other facilities. For more information, contact Interagency Research Animal Committee, Ms. Suzanne Moore, NIH, Bldg 14A, Rm 100, 9000 Rockville Pike, Bethesda, MA, 20892 [301-496-5424] or Proyecto Peruano de Primatologia, Apt. 621, Iquitos, Peru [FAX: 232-951] or PAHO, Coordinator, Vet. Public Health Program, 525 23rd St N.W., Washington, DC 20037 [202-861-3190].
Purpose-bred Monkeys
The Laboratory Animal Center of Kunming, Chinese Academy of Sciences, was established in 1958 and is breeding 14 species of monkeys for sale. The center has over 1000 animals, including Macaca mulatta , M. fascicularis, M. arctoides, M. nemestrina, M. assamensis, M. thibetana, Rhinopithecus bieti, Nycticebus pygmaeus, and N. coucang. They can supply from 60 to 100 animals, 18-36 months of age, per year. The center has clinical laboratories, nursery care, and computerized genealogic and clinical records. They can supply specific pathogen-free rhesus, antibody negative for Herpes simiae, SRV, SIV, STLV-1 and type D retrovirus. They are also seeking collaborative biomedical research using their animals. Contact Dr. Ye Yan, Director, Lab. Animal Center of Kunming, Chinese Academy of Science, Kunming, Yunnan, China (PRC) 650223 [5140390 or 8182475; FAX: 0871-5151823]. -- From the PSIC New Listings.
Time-Labeled Fetal Tissues
A series of nonhuman primate fetuses (longtailed macaque, Macaca fascicularis) will be made available in the coming year, allowing interested investigators to track cohorts of cells labeled on specific days of gestation through the full course of fetal development. Fetuses labeled with tritiated thymidine on days 42, 55, 70, and 90 of gestation will be taken at four or five intervals ranging from one week to six months after labeling. Other fetal age cohorts may be available in the future. This project is partially supported by NCRR/NIH through the Regional Primate Research Center at the University of Washington. For specimens or more information, contact Dr. Anita Hendrickson, Univ. of Washington SM-20, Seattle, WA 98195 [FAX: 206-543-1524; e-mail: [email protected]]. -- Posted on CompMed.
* * *
Training and Enrichment Workshop
The next Primate Training and Enrichment Workshop will be held May 1-4, 1994, at the M. D. Anderson Cancer Center, Dept of Veterinary Sciences, Bastrop, TX. Caregivers and supervisors working with primates in zoos and laboratories are especially encouraged to attend. This hands-on workshop, taught by professionals from M. D. Anderson and Active Environments, Inc., will focus on environmental enrichment and positive reinforcement training to improve primate well-being. The format will be a combination of discussion, demonstration, and lecture, emphasizing group participation. The registration fee of $225 includes all workshop materials, a t-shirt, and most meals during the four days. Attendance is limited to 30 participants.
For more information, or to register, contact Marilyn Hill, PTEW Coordinator, MDACC, Rt. 2, Box 151-B1, Bastrop, TX 78602 [512-321-3991; FAX: 512-332-5208]. -- primate-talk.
Continuing Education and Training
The NIH Office for Protection from Research Risks is cosponsoring an Animal Welfare Education Workshop with Purdue University, in West Lafayette, IN, May 5-6, 1994. The topic will be "Training and Education: Institutional Improvement -- Crisis Prevention." The general theme will center on continuing education and training (CET) as mandated by NIH and USDA. Two areas will be included: differential CET programs and approaches for different types of personnel, and special topics for CET programs such as anesthesia/pain/surgery, euthanasia, and occupational health.
The workshop is open to institutional administrators, members of IACUCs, lab animal veterinarians, investigators, technicians, as well as any person sharing responsibility for the management of an institutional animal care and use program. The regular registration fee for the day-and-a-half program is $150; $50 for grad students, post-docs, and technicians. The fee includes workshop materials, two continental breakfasts, coffee breaks, and two lunches. Continuing education credits will be offered through the Continuing Education Office of Purdue University. For information, contact Ms. Lisa D. Snider, Administrative Asst, Lab. Animal Program, Purdue University, 1071 South Campus Courts-D, West Lafayette, IN 47907-1071 [317-494-7206; FAX: 317-494-0793].
Animal Welfare Training
An Animal Welfare Training Exchange Workshop is planned for September 15-16, 1994, in Research Triangle Park, NC, for people who coordinate training programs for researchers who work with lab animals. The purpose is to exchange training ideas and successful program strategies among research institutions, and to improve individual programs without having to reinvent the wheel. For information, contact Sally Tyree, Lab Animal Resources, Burroughs Wellcome Co., 3030 Cornwallis Rd, Research Triangle Park, NC 27709 [919-315-3988, e-mail: [email protected]].
Animal Welfare Information Requirements
The Animal Welfare Information Center (AWIC) will hold a series of workshops, 10-11 March, 23-24 June, 22-23 September, and 8-9 December, 1994, at the National Agricultural Library, 10301 Baltimore Blvd, Beltsville, MD 20705-2351. The subject will be, Meeting the Informational Requirements of the Animal Welfare Act, focusing on documenting, for Institutional Animal Care and Use Committees, that a thorough literature search was conducted regarding alernatives. Each workshop will be limited to 12 persons. Contact AWIC at the above address or [301-504-6212; FAX: 301-504-5472; e-mail: [email protected]].
* * *
Fulbright Scholar Awards
Fulbright opportunities are available for university lecturing or advanced research in nearly 140 countries. Awards range from two months to a full academic year, and many assignments are flexible to the needs of the grantee. Openings exist in almost every area of the humanities, sciences, and arts. Applications are encouraged from professionals outside academe, as well as from faculty at all types of institutions.
The basic eligibility requirements for a Fulbright Scholar award are U.S. citizenship and the Ph.D. or comparable professional qualifications. For lecturing awards, university or college teaching experi- ence is expected. Language skills are needed for some countries, but most lecturing assignments are in English. The deadline to apply for research or lecturing in all world areas is August 1, 1994. There are other deadlines for special programs. For information and applications, contact the Council for International Exchange of Scholars, 3007 Tilden St, N.W., Suite 5M, Box GNEWS, Washington, DC 20008-3009 [202-686-7877; e-mail (for application requests only): [email protected]].
Postdoctoral and Associateship Programs
The National Research Council announces the 1994 Resident, Cooperative, and Postdoctoral Research Associateship Programs to be conducted on behalf of federal agencies or research institutions whose 140 participating research laboratories are located throughout the United States. The programs provide opportunities for Ph.D. scientists and engineers of unusual promise and ability to perform research on problems largely of their own choosing yet compatible with research interests of the sponsoring laboratory.
Associateships will be awarded for research in fields including biological, health, and behavioral sciences and biotechnology. Most of the programs are open to both U.S. and non-U.S. nationals, and to both recent Ph.D. degree recipients and senior investigators. Awards are made for one or two years, renewable to a maximum of three years. Financial support is provided for allowable relocation expenses and for limited professional travel for the duration of the award. The host laboratory provides the Associate with assistance including facilities, support services, equipment, and travel necessary for the conduct of the approved research program.
Information on specific research opportunities and participating federal laboratories, as well as application materials, may be obtained from the Associateship Programs (TJ 2094/D2), National Research Council, 2101 Constitution Ave, N.W., Washington, D.C. 20418 [FAX: 202-334-2759]. Deadlines for Applications are April 15 and August 15, 1994.
Neurologic Integrity in Fetus and Neonate
The National Institute of Child Health and Human Development (NICHHD) and the National Institute of Neurological Disorders and Stroke (NINDS) invite applications for studies in the development of effective technologies to assess the integrity and function of the developing brain in the human fetus and newborn. The long-term goal of this research is the identification of newborns with brain dysfunction due to early, repetitive or chronic intrauterine central nervous system (CNS) influences/insults which may result in Sudden Infant Death Syndrome and developmental disabilities including cerebral palsy.
The types of research approaches being sought include, but are not restricted to, studies in animals that model early, and/or recurrent, and/or chronic CNS influences/insults in utero and include assessments of central nervous system integrity in the fetus and neonate. The relevance to the human fetus and neonate of animal models must be demonstrated. The models should link the severity of outcome with the nature of the insult (e.g., type, timing, duration, location). Examples of insults of interest include but are not limited to: hypoxia, hypoglycemia, hypovolemia, infectious agents, and exposure to commonly abused substances.
Direct inquiries to Giovanna Spinella, M.D., Div. of Convulsive, Developmental and Neuromuscular Disorders NINDS, Federal Bldg, Rm 8C-10, Bethesda, MD 20892 [301-496-5821]. Application receipt date is April 29, 1994.
Academic Research Enhancement Award
The National Institutes of Health (NIH) is making a special effort to stimulate research in educational institutions that provide baccalaureate training for a significant number of the nation's research scientists but historically have not been major recipients of NIH support. Academic Research Enhancement Award (AREA) funds are intended to support new research projects or expand ongoing research activities proposed by faculty members of eligible institutions in areas related to the health sciences. NIH is inviting grant applications at this time for AREA grants to be awarded competitively in Fiscal Year (FY) 1995.
All domestic health professional schools and other academic institutions offering baccalaureate or advanced degrees in the sciences related to health are eligible, except those that have received research grants and/or cooperative agreements from the NIH totaling more than $2 million per year (direct and indirect costs) in each of four or more years during the period from FY 1987 through FY 1993. Several applications proposing different research projects may be submitted by an applicant institution. Proposed Principal Investigators * must not have active research grant support at the time of award of an AREA grant; * may not submit a regular NIH research grant application for essentially the same project as a pending AREA application; * are expected to conduct the majority of their research at their own institution, although limited access to special facilities or equipment at another institution is permitted; * may not be awarded more than one AREA grant at a time nor be awarded a second AREA grant to continue the research initiated under the first AREA grant.
Although this award is nonrenewable, it will enable qualified individual scientists within the eligible institutions to receive support for feasibility studies, pilot studies, and other small-scale research projects preparatory to seeking more substantial funding from the NIH research grant programs. The application receipt deadline is June 22, 1994. For program guidelines and applications, contact Academic Research Enhancement Award, Office of Grants Information, Div. of Research Grants, Westwood Bldg, Rm 449, Bethesda, MD 20892 [301-594-7248; FAX: 301-594-7045].
Whitehall Foundation Grants
The Whitehall Foundation is interested in funding scholarly research in the area of neurobiology defined as follows: Invertebrate and vertebrate (exclusive of human beings) neurobiology, specifically investigations of neural mechanisms involved in sensory, motor, and other complex functions of the whole organism as these relate to behavior. Types of awards offered are: (1) Research Grants of up to three years for established scientists of all ages working at accredited institutions. These grants will not be awarded to investigators who have already received, or expect to receive, substantial support from other sources, even if it is for an unrelated purpose. Research grants normally range from $10,000 to $40,000 annually. (2) Grants-in-Aid for researchers at the assistant professor level who experience difficulty in competing for research funds because they have not yet become firmly established. Grants-in-aid are awarded for a one-year period and do not exceed $15,000.
Funds may not be used for P.I. salary replacement or summer salary, construction projects or laboratory renovation, travel to conferences or for consultation, secretarial services, tuition, or fellowships. To obtain application forms, applicant should write a one-page letter briefly describing the project and send it to the Whitehall Foundation, Inc., 251 Royal Palm Way, Suite 211, Palm Beach, FL 33480 [407-655-4474; FAX 407-659-4978].
Oral Wound Healing, Tissue Regeneration
The National Institute of Dental Research (NIDR) invites grant applications to conduct multidisciplinary basic and clinical research on wound healing and tissue regeneration associated with the orofacial region. Applications that address the healing, regeneration and repair of oral tissues following periodontal diseases, trauma, and surgical treatment of birth defects such as cleft lip and palate and cancer are sought.
Research topics may include: * Molecular and cellular basis of healthy tissues * Cytokines, growth factors, and biological response modifiers * Biomaterials * Nutrition * Aging, systemic and behavioral conditions * Radiation and chemotherapy * Models of oral wounds * Technology transfer and clinical application of basic science.
Direct inquiries to: Mohandas Bhat, Director, Craniofacial Development and Disorders Program [301-594-7648], Matthew A. Kinnard, Director, Oral Soft Tissue Diseases and AIDS Program [301-594-7641], Dennis F. Mangan, Director, Periodontal Diseases Program [301-594-7641]. Address for all the above is: Extramural Program, NIDR, Westwood Bldg, Rm 509, Bethesda, MD 20892.
* * *
Malaria Info Phone Number
The Centers for Disease Control and Prevention telephone numbers from which health-care providers and the general public can obtain information about the treatment of malaria infections and to which they can report malaria cases acquired in the United States has changed: the new number is 404-488-7760. The malaria hotline number for recommendations for the prevention of malaria remains the same: 404-332-4555.
Dengue Fever
Dengue is a mosquito-transmitted acute disease characterized by the sudden onset of fever, headache, myalgia, rash, nausea, and vomiting. It is endemic in most tropical areas of the world and can occur in U.S. residents returning from international travel, with an incubation period of 7-10 days. Most infected persons experience mild illness, but infection may result in a severe form of the disease, dengue hemorrhagic fever (DHF), characterized by fever, low platelet count, hemorrhage, and a leaky capillary syndrome evidenced by hemoconcentration, hypoalbuminemia, or pleural or abdominal effusions. In 1992, suspected and confirmed cases of dengue were reported in U.S. travellers returning from Thailand, the Philippines, Puerto Rico, India, Nepal, St. Bartholomew, Honduras, Jamaica, and Colombia.
The prevention of dengue relies on avoidance of exposure to mosquitoes. The Aedes species that transmits dengue may bite at any time during the day, although the peak activity occurs during the early morning and late afternoon. Use of mosquito repellent and protective clothing at all times is recommended. Aedes aegypti usually is present in peridomestic settings and is found most often in dark areas such as closets, bathrooms, behind curtains, and under beds. The risk for exposure may be lower for tourists in some settings, including beaches, hotels with well-kept grounds, and heavily forested areas and jungles. Because of the anticoagulant properties of aspirin products, acetaminophen products are recommended for the management of fever. -- From Morbidity and Mortality Weekly Reports, 1994, 43, 97-99.
* * *
T O N K I N * * M * A * N
* B * N * A L O U A T T A
Y S T U M P * * E * L * S
* C * C * P A L L I A T A
T U R K E Y * * L * N * L
* R * L * * A P E * T * I
R A R E R * * * R E A P S
I * O * E L Y * * M * I *
O * A * D * * I M P A L A
N O R T H E R N * E * E *
A * I * E * * D E R M A *
P E N N A N T I * O * T *
O * G * D * * A D R I A N
* * *
Animal Behavior Society
The annual meeting of the Animal Behavior Society, hosted by the Regional P.R.C., University of Washington, will be held 23-28 July, 1994. 28 July will be a joint meeting with ASP. For more information, contact Jim Ha, Primate Center SJ-50, Univ. of Washington, Seattle, WA 98195 [206-543-2420; FAX: 206-685-0305; e-mail: [email protected]].
Behavioural Brain Research
A NATO Advanced Institute, Behavioural Brain Research in Naturalistic and Semi-naturalistic Settings: Possibilities and Perspectives, will be held 10-20 September 1994, at Acquafredda di Maratea, Italy. This interdisciplinary conference is limited to approximately 80 graduate students, postdoctoral scientists and experienced researchers in related fields and includes lectures, poster presentations and practical demonstrations in telemetry, postmortem tract tracing, stereology, and morphometry.
For further information and application forms, contact Dr. L. Ricceri, Lab. Fisiopatologia O.S., Ist. Superiore di Sanita, Via le Regina Elena 299, 00161, Rome, Italy [FAX: +39-6-4957821; e-mail: fos@irmiss].
Wildlife as Research Models
The Scientists Center for Animal Welfare will sponsor a half-day seminar entitled "Wildlife Mammals as Research Models: In the Laboratory and Field" at the annual AVMA conference in San Francisco on July 12, 1994. The co-chairs of the seminar are Kathryn A. L. Bayne (NIH) and Michael Kreger (AWIC). Subjects to be covered include Management in the laboratory; Positive reinforcement in training; Marking and tracking; and Non-capture methods. For more information, contact Conferences, SCAW, 4805 St. Elmo Ave, Bethesda, MD 20814 [301-654-6390; FAX: 301-907-3993].
Meeting Notes
Forest Canopies--Ecology, Biodiversity and Conservation: November 10-13, 1994, Marie Selby Botanical Gardens, Sarasota, Florida. Contact: Dr. Meg Lowman, Director of Research, Shelby Botanical Gardens, 811 South Palm Avenue, Sarasota, FL 34236 [813-366-5730; FAX: 813-366-9807].
Primate Society of Japan, 10th Annual Meeting, 18-19 June, 1994, at the Life Science Inst., Sophia Univ., Tokyo, Japan. Convener: Dr. Takashi Kitahara. Contact: Dr. Khoshi Norikoshi, Life Science Inst., 7-1 Kioichou, Chiyodaku, Tokyo, 102, Japan [03-3238-3488].
Lab. Animal Management Assn, June 16-18, 1994. Mid-Year Forum, "New Ever-Changing Research: Its Impact on Laboratory Animal Facility Design and Management," Houston, TX. Contact Valeri Lansford, Program for Animal Resources, Inst. of Biosciences and Technology, Texas A&M Univ., 2121 W. Holcombe Blvd., Houston, TX 77030-3303 [713-677-7452].
International Congress of Vertebrate Morphology, July 31-Aug 4, 1994, Univ. of Chicago. Contact: Sue Herring, Chair, ICVM Organizing Committee, Dept of Orthodontics, SM-46, Univ. of Washington, Seattle, WA 98195 [206-543-3203; FAX: 206-685-8163; e-mail: [email protected]].
Society for Conservation Biology, 7-11 June, 1995 at Fort Collins, CO. Contact: Richard L. Knight, Dept. of Fishery and Wildlife Biology, Colorado State Univ., Fort Collins, CO 80523 [303-491-6714].
* * *
Thanks to Our Proofreaders
We would like to pause and thank three friends and colleagues who have been proofreading the Newsletter without pay or recognition: Anne Richardson, Deborah van Dam, and Sue Boatwright. They have been contributing to the increasing quality of our "product."
George Ettlinger
We were very sorry to receive a note from Madeline Ettlinger, saying that her husband George died on December 15 following a heart attack. George was a physiological and behavioral psychologist. He retired to England in 1989 from the University of Bielefeld, in Germany. A colleague wrote to me: "If you have time, read some of his papers. You will find evidence throughout of a very keen intellect, a fine methodologist, and a good, clear writer. What more could one ask for?"
James Bowen
James Bowen, of the White Sands Research Center and the Coulston Foundation, died on March 4, 1994. He publications included eighteen papers on chimpanzees. He was coeditor, with C. E. Graham, of Clinical Management of Infant Great Apes (Alan R. Liss, 1985). Donations may be made to a scholarship fund to be established in his memory. Checks should be made out to the Coulston Foundation and sent to the Foundation, 1300 LaVelle Rd, Alamogordo, NM 88310.
* * *
Postdoc in Behavioral Medicine
Postdoctoral research training in behavioral medicine using nonhuman primate models is available in the Dept of Comparative Med., Bowman Gray School of Med. of Wake Forest Univ. Research opportunities include the effects of social factors on disease processes, and the role of individual differences in disease susceptibility. The successful applicant will have a Ph.D. or D.V.M., an interest in behavioral influences on disease processes, computer and data analysis skills. For more information, contact Carol Shively, Dept of Comparative Med., Bowman Gray School of Med. of Wake Forest Univ., Medical Center Blvd, Winston-Salem, NC 27157-1040 [910-716-7045; FAX: 910-764-5818; e-mail: cshively%[email protected]].
Primate Care Technicians
Yerkes Regional Primate Center currently has two openings for Primate Care Technicians at the Main Center in Atlanta. Duties are providing feed, water, and enrichment to a colony of 1500 primates of nine different species; sanitizing caging and equipment; preparing diets; observing animals and reporting abnormalities; assisting in restraint & transport of animals; receiving and storing supplies. Work is in indoor and outdoor areas, including exposure to inclement weather and possibly dangerous animals. Technicians may work with animals exposed experimentally to hazardous agents, such as radioisotopes, chemicals, and infectious agents. Hours are 8:30 AM to 5:00 PM, including alternate weekends and holidays.
Qualifications are * a high school diploma or GED and six months experience OR * an Associate's degree in related field and six months experience OR * AALAS Certification as an Assistant Laboratory Animal Technician OR * Bachelor's degree in related field. Contact: Human Resources Department, Emory University, 1762 Clifton Rd, Atlanta GA 30329.
* * *
Recent Books and Articles
(Addresses are those of first authors)
Books
*Laboratory Animal Welfare: A Guide to Reference Tools, Legal Materials, Organizations, Federal Agencies. C. R. Nyberg, M. A. Porta, & C. Boast. Twin Falls, ID: BN Books, 1994. 391 pp. [Price: $100, from BN Books, Box 1657, Twin Falls, ID 83303-1657]
* Juvenile Primates: Life History, Development, and Behavior. M. E. Pereira & L. A. Fairbanks (Eds). New York, NY: Oxford University Press, 1993. 400 pp. [Price: $52]
* The Use of Tools by Human and Nonhuman Primates. A. Berthelet & J. Chavaillon (Eds). New York: Oxford University Press, 1993. 448 pp. [Price: $66.40]
* Marmosets and Tamarins: Systematics, Behaviour, and Ecology. A. B. Rylands (Ed.). New York: Oxford University Press, 1993. 400 pp. [Price: $72]
* Comparative Primate Pathology. G. B. D. Scott. New York: Oxford University Press, 1993. 304 pp. [Price: $88]
Audiovisual Materials
* 1993 Pathology of Laboratory Animals. [Price: $10, on 3.5" or 5.25"
disks, in IBM or Mac format. Order from Maj. B. Williams, Training
Officer, Dept of Vet. Pathology, AFIP, Washington, DC 20306-6000]
. . Complete syllabus from the 1993 POLA course sponsored by the Armed
Forces Inst. of Pathology.
*Pathology Slide Sets. [Price: $15. Order from Reg. of Comp. Pathol-
ogy, AFIP, Washington, D.C. 20306-6000]
. . Slide sets illustrating 16 primate cases, complete with clinical
histories, results, and discussions, from the 1993 Primate Pathology
Workshop, held in conjunction with the U.S. and Canadian Academies of
Pathology meeting.
* Animal Care Matters. M.I.T. Committee on Animal Care. 24 minutes.
[Price: $20, payable to M.I.T. Order from Animal Care Matters, MIT, 37
Vassar St, 45-105, Cambridge, MA 02139]
. . A video program for education of animal research personnel. It
includes discussions of ethical and moral issues concerning animal
research, the responsibility of research personnel for ensuring humane
treatment of research animals, applicable legislative and regulatory
guidelines, the roles and functions of IACUCs, and questions about
alternatives to animals in research.
Bibliographies
* Simian and Human Retroviruses in Nonhuman Primates: Infection, Dis- ease and Animal Model Studies. A Bibliography, 1992-1993 Update. C. Johnson-Delaney. Seattle: Primate Information Center, 1994, 43 pp. (428 citations, primate & subject indexes, and a list of additional literature proceedings) [Price: $10.00. Order from PIC, RPRC, SJ-50, Univ. of Washington, Seattle, WA 98195]
* Focus On: Veterinary Science & Medicine. Monthly database on disk. [Price: $215 for 12 issues (1 year), from Inst. for Scientific Information, 3501 Market St, Philadelphia, PA 19104-9981]
Booklets
* Summary of New Animal Welfare Regulations. R. R. Smeby. Bethesda: SCAW, 1994. 15 pp. [Price: $4.50 (1 to 9 copies), $3 (10 or more), from SCAW, 4805 St. Elmo Ave, Bethesda, MD 20814]
* Research Helping Animals. Washington, DC: NABR, 1993. 20 pp. [Price:
$3, from Foundation for Biomed. Research, 818 Connecticut Ave, NW,
Suite 303, Washington, DC 20006]
. . This booklet, intended for junior high school children, was funded
by Ciba Pharmaceuticals.
Catalogs
* Primate Library Report: Audio-Visual Acquisitions. December, 1993.
No. 15. (WRPRC, 1220 Capitol Ct, Madison, WI 53715-1299)
. . New listings at the Wisconsin Center's circulating library.
* Federal Laboratory & Technology Resources: A Guide to Services,
Facilities, and Expertise. 5th Ed. Natl Technical Info. Service &
Natl Technology Transfer Ctr, 1993. 728 pp. [Price: $65 + $3 handling,
from NTIS, U.S. Dept of Commerce, 5285 Port Royal Rd, Springfield, VA
22161)
. . Over 1950 listings, indexed by subject, state, resource name, and
agency.
Handbooks
* IATA Live Animal Regulations (20th ed.). [Price: $58 to the Americas, Asia, Australia, and the Pacific; CHF 105 to Europe, Africa, and the Middle East. Available in English, French, or Spanish. Order from Publications Asst, IATA, 2000 Peel St, Montreal, Quebec H3A 2R4, Canada]
* Handbook: Animal Models of Human Disease (Nineteenth Fascicle). C.
C. Capen, T. C. Jones, L. K. Johnson, & G. Migaki (Eds). Washington,
DC: Registry of Comparative Pathology, 1993. [Price: $15. Send check
or money order payable to UAREP to Registry of Comparative Pathology,
AFIP, Washington, DC 20306-6000]
. . Twenty new studies and 10 supplemental updates from the American
Journal of Pathology and the Comparative Pathology Bulletin . One
(on megaloblastic anemia) involves a primate model (cebus monkey).
Magazines, Newsletters, and Reports
* Animal Welfare Information Center Newsletter, 1993, 4[4], (NAL,
AWIC, Beltsville, MD 20705)
. . Includes an article, "The use of behavioral management techniques
to reduce or eliminate abnormal behavior," by G. Laule.
* Community Conservation Consultants, Fall/Winter, 1993. (Howlers Forever, Inc., RD 1, Box 96, Gays Mills, WI 54631).
* The Newsletter, 1994, 5[3]. (Primate Fnd. of Arizona, P.O. Box 20027, Mesa, AZ 85277-0027)
* Neotropical Primates: A Newsletter of the Neotropical Section of the IUCN/SSC Primate Specialist Group, 1993, 1[4]. (Conservation Intl, Av. A. A. Caram 820/302, 31275-000, Belo Horizonte, Minas Gerais, Brazil)
* ILAR News, 1993, 35[2]. (ILAR, 2101 Constitution Ave, NW, Washington, DC 20418)
*SCAW Newsletter, 1993, 15[4]. (SCAW, 4805 St. Elmo Ave, Bethesda, MD 20814).
* Sulawesi Primate Newsletter, 1994, 1[3]. [Subscription: $5 (individuals), $10 (institutions), from N. Bynum, 1126 John Jones Rd, Bahama, NC 27503; also available by e-mail from [email protected]]
New Journals
* In Touch: For the Humane Use and Care of Laboratory Primates. 1994,
1[1]. (923 Laurel St, Junction City, OR 97448)
. . "...[A] forum for the sharing of ideas and solutions to improve the
lives of nonhuman primates throughout the research community." The
initial issue contains articles by R. S. Fouts, D. H. Fouts, M. L. A.
Jensvold, & M. D. Bodamer; D. R. Haskin; and E. O'Connor & V. Reinhardt.
Anatomy & Physiology
*Relationship between social factors and pituitary-adrenocortical
activity in female rhesus monkeys (Macaca mulatta). Gust, D. A.,
Gordon, T. P., Hambright, M. K., & Wilson, M. E. (Yerkes RPRC, Emory
Univ., Atlanta, GA 30322). Hormones and Behavior, 1993, 27, 318-331.
. . In two groups, one long-term and including kin, the other recently
formed of unrelated animals, neither group nor dominance rank was a
predictor of cortisol levels, based on regression analysis. However,
rank significantly correlated with cortisol levels in the established
but not in the recently formed group. The data suggest that cortisol
levels are influenced not only by negative interactions, such as
receiving bites, but also by positive interactions such as receiving
grooms. A higher rate of reconciliatory behaviors in the recently-formed group may account for the lack of a dominance rank/cortisol
relationship.
*The size of the neocortex in relation to ecology and social structure
in monkeys and apes. Sawa-guchi, T. (Dept of Neurophysiology, PRI,
Kyoto Univ., Inuyama, Aichi 484, Japan). Folia Primatologica, 1992,
58, 131-145.
. . Relative sizes of the neocortex (RSNs) are related to diet and
social structure: frugivorous anthropoids had higher values of RSNs
than folivorous ones, and polygynous anthropoids had significantly
higher values than monogynous ones. RSNs were also positively correlated with the size of the troop.
*Urinary enzyme concentrations in the owl monkey (Aotus nancymae).
Weller, R. E., Malaga, C. A., Buschbom, R. L., & Ragan, H. A. (Battelle Pacific NW Labs, P.O. Box 999, P7-52, Richland, WA 99352).
Journal of Medical Primatology, 1993, 22, 340-347.
. . The activity of 3 urinary enzymes was evaluated in 71 adult owl
monkeys, 15 of which showed evidence of renal dysfunction. Urinary
enzymes and creatinine ratios were significantly correlated with indices of renal damage and could identify active renal disease.
*Comparison of blood and urine analytes between two karyotypes of owl
monkey, Aotus nancymaeand Aotus vociferans. Weller, R. E., Buschbom, R. L., Malaga, C. A., & Ragan, H. A. (Address same as above).
Journal of Medical Primatology, 1993, 22, 368-373.
. . Statistically significant differences were noted in serum calcium,
serum phosphorus, serum sodium, serum potassium, serum urea nitrogen,
urine calcium, calcium clearance, and fractional clearance of calcium
between the two karyotypes, suggesting that A. vociferans regulate
calcium-phosphorus and electrolyte homeostasis in a manner different
from that of A. nancymae.
*Immunohistochemical distribution of leucocyte antigens in lymphoid
tissues of cynomolgus monkeys (Macaca fascicularis). Li, S.-L., Kaaya, E., Feichtinger, H., Biberfeld, G., & Biberfeld, P. (P. B., Immunopathology Lab., Karolinska Hospital, S-10401 Stockholm, Sweden).
Journal of Medical Primatology, 1993, 22, 285-293.
. . Of 54 clusters of differentiation (CD) antigens, 39 were expressed
essentially with the same immunostaining patterns in the monkey as in
human lymphoid tissues. By immunoblotting L26 (CD20) detected a 35 Kd
molecule in the monkey lymph node. Our observations indicated that
most of the CD antigens are expressed and can be studied in lymphoid
tissues of cynomolgus monkeys.
*Enzyme immunoassays for ovarian steroid metabolites in the urine of
Macaca fascicularis. Shideler, S. E., Shackleton, C. H. L., Moran,
F. M., Stauffer, P., Lohstroh, P. N., & Lasley, B. L. (ITEH, Univ. of
California, Davis, CA 95616). Journal of Medical Primatology, 1993,
22, 301-312.
. . In vivo studies using C�� labeled estradiol (E�) and progesterone
(Po) were performed to characterize the time course and metabolic fate
of circulating E� and Po. Co-chromatography of human, orangutan, and
macaque luteal phase urine samples demonstrated the presence of a
steroid conjugate peak in all 3 species that was identified as being
androsterone and etiocholanolone glucuronides. An enzyme immunoassay
for urinary metabolites of Po was developed subsequently for Macaca
spp. using a monoclonal antibody that cross-reacted with both C-19 and
C-21 metabolites.
*Visual optics and retinal cone topography in the common marmoset
(Callithrix jacchus). Troilo, D., Howland, H. C., & Judge, S. J.
(Dept of Psychology, Uris Hall, Cornell Univ., Ithaca, NY 14853).
Vision Research, 1993, 33, 1301-1310.
. . Similarities and differences between the marmoset eye and those of
humans and rhesus monkeys are described.
*Ocular development and visual deprivation myopia in the common marmoset (Callithrix jacchus). Troilo, D. & Judge, S. J. (Address same
as above). Vision Research, 1993, 33, 1311-1324.
. . The growth of the marmoset eye and the development of refraction
are described, in normal and visually deprived cases.
*Clinical implications of the fibre order in the optic pathway of primates. Reese, B. E. (Neuroscience Res. Inst., Univ. of California,
Santa Barbara, CA 93106). Neurological Research, 1993, 15, 83-86.
. . A review of recent studies of the optic pathways of primates and
other animals shows these studies contradict classical accounts, but
are consistent with incongruous field defects and functionally selective visual impairments documented in the clinical literature.
*The effects of diet, age, and sex on the mineral content of primate
bones. Grynpas, M. D., Hancock, R. G. V., Greenwood, C., Turnquist,
J., & Kessler, M. J. (Samuel Lunenfeld Res. Inst., 600 Univ. Ave,
Toronto, Ont M5G 1X5, Canada). Calcified Tissue International, 1993,
52, 399-405.
. . The Cayo Santiago rhesus colony was changed from a 15% protein diet
to one with 24-25% protein in 1969. Skeletons, of known age, sex, parity, and matrilineage, of animals fed each diets, were analyzed.
Results show that increasing protein content in the diet does not
change the bulk mineral content of primate bones. It was also found
that calcium content is lower during middle age than in youth and old age.
*Ambient light modifies gonadotropin-releasing hormone pulse generator
frequency in the rhesus monkey. O'Byrne, K. T., Thalabard, J.-C., Chiappini, S. E., Chen, M.-D., Hotchkiss, J., & Knobil, E. (E. K., Lab.
for Neuroendocrinology, Univ. of Texas Med. School, Rm 7128 MSMB,
Houston, TX 20708). Endocrinology, 1993, 133, 1520-1524.
. . In the rhesus monkey the activity of the GnRH pulse generators is
evidenced by sharp increases in hypothalmic multiunit electrical
activity that are coincident with LH pulses in the peripheral circulation. Changes in pulse generator frequency, observed morning and
night, also occurred when lights were turned off or on at times other
than the normal illumination period. These changes did not occur when
the animals were awakened with loud noises at such times. This direct
action of light, however, is superimposed upon a diurnal rhythm.
Animal Models
*Abstracts from the 11th Annual Symposium on Nonhuman Primate Models
for AIDS, held September 19-22, 1993. Journal of Medical Primatology, 1993, 22, 442-468.
*Immunopathology of experimental autoimmune uveoretinitis in primates.
Fujino, Y., Li, Q., Chung, H., Hikita, N., Nussenblatt, R. B., Gery,
I., & Chan, C.-C. (C.-C. C., Bldg 10, Rm 10N206, NIH/NEI, Bethesda, MD
20892). Autoimmunity, 1992, 13, 303-309.
. . The eyes and pineal glands from 10 rhesus monkeys immunized with
S-antigen were studied using routine histopathological and immunohistochemical techniques. Seven of 10 animals developed uveitis between
19 and 33 days after immunization. An influx of B lymphocytes and the
formation of subretinal fibrosis differentiate the disease in the monkey from that in the rat and mouse.
*Evidence that polyreactive antibodies are deposited in rejected discordant xenografts. Geller, R. L., Bach, F. H., Turman, M. A., Casali,
P., & Platt, J. L. (Baxter Healthcare Corp., Rte 120 and Wilson Rd,
Round Lake, IL 60073). Transplantation, 1993, 55, 168-172.
. . Findings are consistent with the hypothesis that idiotypes of polyreactive natural antibodies are broadly crossreactive. They also suggest that the polyreactive and xenoreactive IgM, which have been
detected based on in vitro assays, may be deposited in a xenogeneic
organ graft.
*Immunogenicity and safety of a recombinant vaccinia virus vaccine
expressing the carcinoembryonic antigen gene in a nonhuman primate.
Kantor, J., Irvine, K., Abrams, S., Snoy, P., Olsen, R., Greiner, J.,
Kaufman, H., Eggensperger, D., & Schlom, J. (J. S., Lab. of Tumor
Immunology & Biology, NCI, NIH, Bldg 10, Rm 8B07, 9000 Rockville Pike,
Bethesda, MD 20892). Cancer Research, 1992, 52, 6917-6925.
. . The administration of a human carcinoembryonic antigen (CEA) recombinant vaccine to rhesus monkeys induces both a humoral and a cell-mediated immune response directed against human CEA and appears to be
safe.
*Effects of excess selenomethionine on selenium status indicators in
pregnant long-tailed macaques (Macaca fascicularis). Hawkes, W. C.,
Willhite, C. C., Craig, K. A., Omaye, S. T., Cox, D. N., Choy, W. N.,
& Hendrickx, A. G. (USDA-ARS Western Human Nutrition Research Ctr,
P.O. Box 29997, Presidio of San Francisco, CA 94129). Biological
Trace Element Research, 1992, 35, 281-297.
. . Based on tests with 40 animals, plasma, erythrocyte, and hair selenium concentrations may be useful for monitoring and preventing the
toxicity of L-selenomethionine administered to humans in cancer chemoprevention trials.
*Pathology induced by interleukin-6. Ryffel, B., Mihatsch, M. J.,
Woerly, G. (Inst fur Toxikologie, Eidgenoss. Tech. Hochschule, Univ.
Zurich, CH8603 Schwerzenbach/Zurich, Switzerland). International
Review of Experimental Pathology, 1993, 34A, 79-89.
. . Recombinant human interleukin-6 (IL-6) was administered at high
doses to mice, rats, and rhesus monkeys. IL-6 had an immunostimulatory
and hemopoietic (especially on megakaryocytes) effect in all species.
An acute-phase response was most pronounced in the monkeys, which was,
however, not associated with any significant histopathological liver
change. No evidence of glomerular pathology was found.
*Experimental hepatitis E: Pathogenesis in cynomolgus macaques
(Macaca fascicularis). Longer, C. F., Denny, S. L., Caudill, J. D.,
Miele, T. A., Asher, L. V. S., Myint, K. S. A., Huang, C.-C., Engler,
W. F., LeDuc, J. W., Binn, L. N., & Ticehurst, J. R. (Dept of Virus
Diseases, WRAIR, Washington, DC 20307-5100). Journal of Infectious
Diseases, 1993, 168, 602-609.
. . Eight animals were inoculated with hepatitis E virus (HEV).
Results support the concept that experimental hepatitis E has an initial phase in which hepatic HEV replication is accompanied by the
onset of hepatitis and a later phase in which the appearance of anti-HEV is accompanied by progression of the hepatitis.
*Interruption of vascular thrombus formation and vascular lesion formation by dietary n-3 fatty acids in fish oil in nonhuman primates.
Harker, L. A., Kelly, A. B., Hanson, S. R., Krupski, W., Bass, A.,
Osterud, B., FitzGerald, G. A., Goodnight, S. H., & Connor, W. E.
(Div. of Hematology & Oncology, Emory Univ. School of Med., P.O.
Drawer AR, Atlanta, GA 30322). Circulation, 1993, 87, 1017-1029.
. . In baboons, dietary n3FAs in high doses eliminate both vascular
thrombus formation and vascular lesion formation after mechanical vascular injury, while largely sparing hemostatic function and modestly
reducing blood thrombotic responses.
*Childhood consumption of dietary polyunsaturated fat lowers risk for
coronary artery atherosclerosis in African green monkeys. Wolfe, M.
S., Parks, J. S., Morgan, T. M., & Rudel, L. L. (L. L. R., Dept of
Comp. Med., Bowman Gray School of Med., Medical Center Blvd, Winston-Salem, NC 27157-1040). Arteriosclerosis and Thrombosis, 1993, 13,
863-875.
. . Age, gender, and dietary fat type independently affected plasma
lipid and apolipoprotein concentrations. Total plasma cholesterol and
apoB concentrations were lower in animals consuming polyunsaturated
fat than in those getting saturated fats from birth into young adulthood.
*Comparative pathology of recombinant murine interferon-gamma in mice
and recombinant human interferon-gamma in cynomolgus monkeys. Terrell,
T. G. & Green, J. D. (Dept of Safety Evaluation, Genentech, Inc., South
San Francisco, CA 94080). International Review of Experimental
Pathology, 1993, 34B, 73-101.
. . Human IFN-gamma is active on nonhuman primate cells, though not at
the same level as on human cells. Multidose studies in cynomolgus
monkeys with rHulFN-gamma for 28 or 90 days were predictive of many of
the dose-limiting clinical toxicities. Qualitative similarity was
observed between toxicity studies employing rHulFN-gamma in the cynomolgus monkey and rMulFN-gamma in the mouse.
*Pharmacokinetic evaluation of anti-HIV drug interactions: Effect of
zidovudine on 2'-3'-dideoxyinosine kinetics in monkeys. Qian, M., Swagler, A. R., Mehta, M., Vishwanathan, C. T., & Gallo, J. M. (J. M.
G., Dept of Pharmaceutics, College of Pharmacy, Univ. of Georgia, Athens, GA 30602). Antiviral Chemistry & Chemotherapy, 1993, 4,
155-159.
. . AZT does not alter the pharmacokinetics of dideoxyinosine (ddI) in
long-tailed macaques at the dose studied, providing a basis for
rational dosage design for combined ddI and AZT treatments in HIV
infection.
*Constant low-dose oestradiol replacement accelerates skeletal maturation and growth in ovariectomized adolescent rhesus monkeys. Wilson,
M. E., Gordon, T. P., & Tanner, J. M. (Yerkes RPRC of Emory Univ.,
Field Stn, 2409 Taylor Ln., Lawrenceville, GA 30243). Journal of
Endocrinology, 1993, 137, 519-527.
. . Juvenile monkeys were ovariectomized and were given either an oestradiol (OE�)-implant or no treatment. Results indicate that, in the
absence of OE�, growth in female monkeys has periods of acceleration
and that OE� may enhance this preset pattern. The growth-promoting
effects of OE� are determined by the degree of skeletal maturity. Once
a certain degree of skeletal maturity is attained, bones may be primed
for maximal growth but, as skeletal maturity continues to advance, the
effect of OE� on growth diminishes.
*Surfactant replacement therapy in utero for prevention of hyaline
membrane disease in the preterm baboon. Galan, H. L., Cipriani, C.,
Coalson, J. J., Bean, J. D., Collier, G., & Kuehl, T. J. (T. J. K.,
Scott & White Clinic, 2401 S. 31st St, Temple, TX 76508). American
Journal of Obstetrics & Gynecology, 1993, 169, 817-824.
. . A single treatment with surfactant in utero significantly improved
the clinical course, but did not completely prevent, hyaline membrane
disease in an established animal model.
*Platelet-derived growth factor and dexamethasone combined with a collagen matrix induce regeneration of the periodontium in monkeys.
Rutherford, R. B., Ryan, M. E., Kennedy, J. E., Tucker, M. M., & Charette, M. F. (Univ. of Michigan School of Dentistry, Ann Arbor, MI
48109-1078). Journal of Clinical Periodontology, 1993, 20, 537-544.
. . The presence of substantial amounts of regenerated periodontium
including increased height of the alveolar bone; fill of vertically
resorbed interdental alveolar septa in PDGF/dexamethasone/CM-treated
lesions suggests that this combination may provide a new therapeutic
agent for the regeneration of lesions of periodontitis associated with
horizontal as well as angular bony defects.
*Primate models of age-related cognitive deficits. Cole, B. J. (Dept
of Neuropsychopharmacology, Schering AG, Mullerstr. 170-178, D-1000
Berlin 65, Germany). Neuroscience Research Communications, 1993,
13, S43-S46.
. . A review of studies indicating that humans and nonhuman primates
show similar profiles of age-related cognitive deficits and neuropathology.
*Lack of susceptibility of various primates and woodchucks to hepatitis C virus. Abe, K., Kurata, T., Teramoto, Y., Shiga, J., & Shikata,
T. (Dept of Pathology, NIH, Toyama 1-23-1, Shinjuku-ku, Tokyo 162,
Japan). Journal of Medical Primatology, 1993, 22, 433-434.
. . Cynomolgus, rhesus, green, and Japanese monkeys, Doguera baboons,
and woodchucks do not appear to be susceptible to hepatitis C virus.
Behavior
*Rate of takeovers in groups of Hanuman langurs (Presbytis entellus)
at Jaipur. Reena, M. & Ram, M. B. (Dept of Zoology, Univ. of
Rajasthan, Jaipur, India). Folia Primatologica, 1992, 58, 61-71.
. . Population density rather than habitat disturbances seems to be
correlated with takeovers. Aggressiveness, a marked transition period,
interim residencies, 'sneaky' copulations, and multimale phases were
some of the typical features accompanying takeovers.
*DNA fingerprinting reveals that infant care by male Barbary macaques
(Macaca sylvanus) is not paternal investment. Paul, A., Kuester, J.,
& Arnemann, J. (Inst fur Anthropologie, Burgerstrasse 50, D-W-3400
Gottingen, Germany). Folia Primatologica, 1992, 58, 93-98.
. . DNA data shows that kinship is neither a primary nor a necessary
factor in the 'male care' system of the Barbary macaque.
*Alloparental care in free-ranging Lemur catta at Berenty Reserve,
Madagascar. Gould, L. (Dept of Anthropology, Campus Box 1114, Washington Univ., St. Louis, MO 63130). Folia Primatologica, 1992, 58,
72-83.
. . Group members of all age/sex classes participate to some degree in
alloparental care of infants. Rank or sex of infant did not affect the
amount of alloparental care received. Infants and helpers all receive
benefits from participation in allocare.
*Social position and personality in Macaca fascicularis. Welker, C.,
Schafer-Witt, C., & Voigt, K. (Zoologie u. vergleich. Anatomie, Primatenethologie, Univ. Kassel, D-W-3500 Kassel, Germany). Folia Primatologica, 1992, 58, 112-117.
. . Individuals do not need the support of mothers and/or siblings to
attain a high or low position within the group hierarchy. Infants of
high-ranking mothers exhibited strategies different from those of low-ranking mothers. Depending on their behavior, they achieved a higher
or a lower position.
*Adoption of an additional infant by a Western lowland gorilla
(Gorilla gorilla gorilla). Blersch, B. H. & Schmidt, C. R.
(Nordstrasse 4, CH-8800 Thalwil, Switzerland). Folia Primatologica.
1992, 58, 190-196.
. . An account of how a 10-year-old gorilla at the Zurich Zoo, while
caring for her own 14-month-old son, adopted a female neonate.
*Costs of maternal care: Infant-carrying in baboons. Altmann, J. &
Samuels, A. (Dept of Ecology & Evolution, Univ. of Chicago, 940 E.
57th St, Chicago, IL 60637). Behavioral Ecology and Sociobiology,
1992, 29, 391-398.
. . Although carrying a growing infant is an increasingly costly behavior, during the period of nutritional dependence energetic costs to
the mother are greater if an infant travels independently instead of
being carried by its mother. Yet infants increased locomotor independence at a younger age than predicted by a simple model of maternal
energetic efficiency. Trade-offs in energetic economy may enhance a
mother's future reproduction at the expense of her present infant, may
enhance survival of the present infant by promoting early acquisition
of developmentally essential skills, or may suggest the importance of
additional factors that influence the mother's and infant's behavior.
*Maternal protectiveness and response to the unfamiliar in vervet monkeys. Fairbanks, L. A. & McGuire, M. T. (Dept of Psychiatry & Biobehav. Sci., Univ. of California, 760 Westwood Plaza, Los Angeles, CA
90024). American Journal of Primatology, 1993, 30, 119-129.
. . To test the hypothesis that more protective mothers produce more
cautious offspring, maternal protectiveness was varied by the introduction of new breeding males. Mothers responded to the presence of
new males by increased protectiveness toward infants born in the year
following the introductions. Immatures who had been born in New Male
years were more cautious and had significantly longer latencies to
approach a novel stimulus than those who had been born in Resident
Male years.
*Sons of low-ranking female rhesus macaques can attain high dominance
rank in their natal groups. Manson, J. H. (Dept of Anthropology, Univ.
of Michigan, 1054 LSA Bldg, Ann Arbor, MI 48109-1382). Primates,
1993, 34, 285-288.
. . Five adult and subadult sons of middle- and low-ranking female rhesus were observed to hold high dominance rank in their natal groups
during a 12-month study. Three of these males also experienced high
mating success during at least one mating season. These findings
present a challange to the hypothesis that natal transfer functions to
increase male access to fertile females.
Care
*Environmental enrichment for captive wildlife through the simulation
of gum-feeding. Kelly, K. (Dept of Pathology, Nat. Zool. Park, Wash-
ington, DC 20008). AWIC Newsletter, 1993, 4[3], 1-2, 5-10.
. . A complete discussion of the how and why of feeding gums to certain
animals in captivity.
*Regurgitation and reingestion in a captive chimpanzee (Pan troglodytes). Morgan, L., Howell, S. M., & Fritz, J. (J. F., Primate Fnd.
of Arizona, P.O. Box 20027, Mesa, AZ 85277-0027). Lab Animal, 1993,
22[8], 42-45.
. . Behavioral training coupled with a dietary change (e.g., replace
fruit with vegetables and browse) may reduce regurgitation temporarily, but constant attention to the individual, the diet, and intermittent training may all be necessary to control the frequency of the
behavior.
Conservation
*A reintroduction program for the conservation of the black howler
monkey in Belize. Horwich, R. H., Koontz, F., Saqui, E., Saqui, H., &
Glander, K. (RD 1, Box 96, Gays Mills, WI 54631). Endangered Species
UPDATE, 1993, 10[6], 1-6.
. . Details of the transfer of 14 animals, 12 of which survived after
10 months of observation.
Development
*Cross-sectional growth standards for captive baboons: I. Organ weight
by chronological age. Mahaney, M. C., Leland, M. M., Williams-Blangero, S., & Marinez, Y. N. (Dept of Genetics, Southwest Fnd. for
Biomed. Research, P.O. Box 28147, San Antonio, TX 78228-0147). Journal of Medical Primatology, 1993, 22, 400-414.
*Cross-sectional growth standards for captive baboons: II. Organ
weight by body weight. Mahaney, M. C., Leland, M. M., Williams-Blangero, S., & Marinez, Y. N. (Address same as above). Journal of
Medical Primatology, 1993, 22, 415-427.
. . Organ weight reference standards from necropsy reports for 634
healthy, pedigreed, captive female and male baboons.
Disease
*Analysis of envelope changes acquired by SIVmac239 during neuroadaption in rhesus macaques. Anderson, M. G., Hauer, D., Sharma, D. P.,
Joag, S. V., Narayan, O., Zink, M. C., & Clements, J. E. (J. E. C.,
Dept of Molecular Biology & Genetics, Johns Hopkins Univ. School of
Med., Baltimore, MD 21205). Virology, 1993, 195, 616-626.
. . Construction of an infectious recombinant virus containing the
tat, rev, and env genes from 17E and the remainder of the genome
from the parental virus SIVmac239 resulted in a virus that had the
macrophagetropism of 17E virus isolated from brain.
*Multiple viral determinants contribute to pathogenicity of the
acutely lethal simian immunodeficiency virus SIVsmmPBj variant.
Novembre, F. J., Johnson, P. R., Lewis, M. G., Anderson, D. C.,
Klumpp, S., McClure, H. M., & Hirsch, V. M. (V. M. H., Lab. of Infectious Diseases, NIAID, Rockville, MD 20852). Journal of Virology,
1993, 67, 2466-2474.
. . Although the disease course of SIV-induced immunodeficiency is generally measured in months to years, a disease syndrome that results in
death in 5 to 14 days has been described in pig-tailed macaques
infected with the SIVsmmPBj (PBj) strain. This study suggests that
multiple viral genes of PBj are critical for development of acute disease. The env gene but not the long terminal repeat PBj was required
for acute disease induction; however env must act in concert with
another gene(s) of the PBj genome.
*Spermatogenesis and hormone levels in rhesus macaques inoculated with
simian immunodeficiency virus. Nadler, R. D., Manocha, A. D., &
McClure, H. M. (Yerkes RPRC, Emory Univ., Atlanta, GA 30322). Journal
of Medical Primatology, 1993, 22, 325-329.
. . The presence of sperm in testicular tissue of rhesus macaques that
died as a result of infection with SIV was related to age and body
weight. Depressed testosterone levels were not associated with elevated LH levels. The data suggest that azoospermia in the SIV-infected
macaques was due to cachexia and not a direct effect of virus on the
testis, supporting a similar hypothesis regarding azoospermia in men
infected with HIV.
*Dubin-Johnson-like syndrome in golden lion tamarins (Leontopithecus
rosalia rosalia). Schulman, F. Y., Montali, R. J., Bush, M., Citino,
S. B., Tell, L. A., Ballou, J. D., Hutson, T. L., St. Pierre, M.,
Dufour, J. F., Gatmaitan, Z., Johnson, F. B., & Arias, I. M. (Dept of
Pathology, Nat. Zoo. Park, Smithsonian Inst., Front Royal, VA 22630).
Veterinary Pathology, 1993, 30, 491-498.
. . On routine blood screens, persistent conjugated hyperbilirubinemia
was discovered in two groups of closely related adult female golden
lion tamarins. The clinicopathologic presentation was similar to that
described for Dubin-Johnson syndrome in humans.
*Infrequent shedding and transmission of herpesvirus simiae from seropositive macaques. Weir, E. C., Bhatt, P. N., Jacoby, R. O., Hilliard,
J. K., & Morgenstern, S. (Sect. of Comp. Med., Yale Univ. School of
Med., P.O. Box 208016, New Haven, CT 06520-8016). Laboratory Animal
Science, 1993, 43, 541-544.
. . Surveys of the macaque colony at Yale over three years suggest that
shedding of B virus from seropositive macaques is uncommon, when the
animals are subjected to common laboratory procedures or environments,
and that transmission is rare in singly-housed animals.
*Herpesvirus encephalitis in a ruffed lemur (Varecia variegatus).
Kornegay, R. W., Baldwin, T. J., & Pirie, G. (Dept of Vet. Pathology,
School of Vet. Med., L.S.U., Baton Rouge, LA 70803). Journal of Zoo
and Wildlife Medicine, 1993, 24, 196-203.
. . A herpesvirus has not been previously reported as a cause of encephalitis in lemurs. Possible antemortem procedures for early diagnosis
of suspected cases could include viral culture and histopathologic,
immunofluorescent, or ultrastructural examination of brain biopsies,
or polymerase chain reaction of cerebral spinal fluid.
*Disseminated B virus infection in a cynomolgus monkey. Simon, M. A.,
Daniel, M. D., Lee-Parritz, D., King, N. W., & Ringler, D. J. (Div.
of Comp. Pathology, New England RPRC, Southborough, MA 01772-9102).
Laboratory Animal Science, 1993, 43, 545-550.
. . A cynomolgus monkey was euthanized one week following dystocia
because of severe peritonitis. B virus was isolated from serum. As
systemic B-virus infection was not diagnosed before death, these findings underscore the need for universal precautions when handling
blood, fluids, or tissues from macaques.
*Mesocestoides infection in captive olive baboons (Papio cynocephalus
anubis). Hubbard, G. B., Gardiner, C. H., Bellini, S., Ehler, W. J.,
Conn, D. B., & King, M. M. (Dept of Lab. Animal Med., Southwest Fnd.
for Biomed. Research, P.O. Box 28147, San Antonio, TX 78228-0147).
Laboratory Animal Science, 1993, 43, 625-627.
. . The baboon can and does serve as an intermediate host and reservoir
for Mesocestoides lineatus tetrathyridia, a tapeworm. Researchers,
pathologists, and those working with research animals must be aware of
the occurrence, appearance, and possible effects of this parasite.
*Asymptomatic polyarteritis in a cynomolgus monkey. Albassam, M. A.,
Lillie, L. E., & Smith, G. S. (Parke-Davis Research Inst., 2270 Speakman Dr., Mississauga, Ontario L5K 1B4, Canada). Laboratory Animal
Science, 1993, 43, 628-629.
. . First report of spontaneous polyarteritis in the cynomolgus monkey.
The pathogenesis is unknown.
*Simian foamy virus type 3 (SFV-3) in latently infected vero cells:
Reactivation by demethylation of proviral DNA. Schweizer, M., Fleps,
U., Jackle, A., Renne, R., Turek, R., & Neumann-Haefelin, D. (D.
N.-H., Abt. Virologie, Inst fur Med. Mikrobiologie u. Hygiene, Univ.
Freiburg, Hermann-Herder-Strasse 11, D-7800 Freiburg, Germany).
Virology, 1993, 192, 663-666.
. . Cell cultures latently infected with SFV-3 were established by suppressing lytic infection in Vero cells with 3'-azido-3'-deoxythymidine
(AZT) and homologous antibodies. Over two years of subcultivation, no
spontaneous expression of proviral genes could be detected. However,
the demethylating agent 5'-azacytidine reactivated lytic infection,
proving conservation of the complete viral genome. Comparison of proviral DNA from latently and lytically infected cells supports the
notion that methylation is instrumental in keeping SFV-3 infection in
latency.
*Alkaptonuria in a cynomolgus monkey (Macaca fascicularis). Johnson,
E. H. & Miller, R. L. (New Mexico RPR Lab., New Mexico State Univ.,
Holloman AFB, NM 88330). Journal of Medical Primatology, 1993, 22,
428-430.
. . An 8-year-old wild caught animal has alkaptonuria, a condition
characterized by the passage of normal-colored urine which darkens
upon standing. No other clinical manifestations, such as deposits of
pigment in the skin or mucous membranes or development of ochronitic
arthritis, were observed.
*Viral persistence of simian type D retrovirus (SRV-2/W) in naturally
infected pigtailed macaques (Macaca nemestrina). Moazed, T. C. &
Thouless, M. E. (Dept of Pathobiology, School of Public Health, Univ.
of Washington, Seattle, WA 98195). Journal of Medical Primatology,
1993, 22, 382-389.
. . In viremic animals infected pre-, peri-, or postnatally by the
mother, proviral DNA was limited, while in other viremic animals, it
was found in many tissues. In nonviremic, seropositive macaques, provirus was detected in bone marrow and/or ileocecal junction.
General
*Xenotransplantation: State of the art. Hammer, C. (Inst. of Surgical
Research, Clinic Grosshadern, Ludwig-Maximilians Univ., Marchioninistr. 15, 81366 Munchen 70, Germany). Transplantation Proceedings,
1993, 25 (Suppl 3), 35-37.
. . The author believes that xenografting may soon prove to be an
alternative to allografts, although to date neither the elimination of
single factors nor the mitigation of whole systems have led to clinically relevant survival times.
*Chimpanzees in AIDS research: A biomedical and bioethical perspective. van Akker, R., Balls, M., Eichberg, J. W., Goodall, J., Heeney,
J. L., Osterhaus, A. D. M. E., Prince, A. M., & Spruit, I. (J. W. E.,
3 Deggs Circle, Newtown Square, PA 19073). Journal of Medical Primatology, 1993, 22, 390-392.
. . A consensus view of the appropriate utilization of chimpanzees in
AIDS research developed at a meeting of a group of bioethicists and
scientists involved in AIDS research with chimpanzees. The paper considers which types of studies are scientifically justifiable in this
species, the conditions under which such studies should be carried
out, and the conditions which should be encouraged for postexperimental retirement of these animals.
Genetics
*Non-invasive sampling and DNA amplification for paternity exclusion,
community structure, and phylogeography in wild chimpanzees. Morin,
P. A., Wallis, J., Moore, J. J., Chakraborty, R., & Woodruff, D. S.
(Dept of Biology, Univ. of California, San Diego, La Jolla, CA 92093).
Primates, 1993, 34, 347-356.
. . Amplification of DNA from shed hair is an important new tool for
the acquisition of genetic information that will facilitate the study
and management of both captive and free-ranging chimpanzees. Hierarchical analysis of population genetics of chimpanzees has revealed
high historical levels of gene flow and large effective population
sizes, as well as substantial divergence between the West African
subspecies and those from Central and East Africa. Analyses have cast
light on divergence of subspecies, kin selection, and results of habitat fragmentation.
*Power and limits of DNA-profiling in primate populations: Paternity
assessment in rhesus macaques from Cayo Santiago. Krawczak, M., Berard, J. D., Bercovitch, F. B., Schmidtke, J., & Nurnberg, P. (Abt.
Humangenetik, Med. Hochschule, Hannover, Germany). Primates, 1993,
34, 395-402.
. . In 11/15 cases, sires could be identified by comparing the multilocus DNA profiles of 19 males in a social group to those of the corre-
sponding mother/child dyads. All 19 males could be excluded in the
remaining four cases. In a second social group paternity testing was
impeded by a deficit of maternal bands and by an increased extent of
band sharing of mothers and their infants. Possible explanations are
discussed.
*Research-oriented genetic management of nonhuman primate colonies.
Williams-Blangero, S. (Dept of Genetics, Southwest Fnd. for Biomed.
Research, P.O. Box 28147, San Antonio, TX 78228-0147). Laboratory
Animal Science, 1993, 43, 535-540.
. . The statistical power of experiments using nonhuman primates can be
improved when animals are selected on the basis of their genetic values or genotypes for experimentally relevant traits, because the quantified genetic variation among subjects can thus be minimized. Incorporating experimental needs into the overall genetic management plans
for captive breeding colonies helps ensure the long-term viability of
colonies for meeting the demands of both breeding and research.
Instruments & Techniques
*Fast scan magnetic resonance imaging and Doppler ultrasonography of
uteroplacental hemodynamics in the rhesus monkey (Macaca mulatta).
Panigel, M., Dixon, T., Constantinidis, I., Sheppard, S., Swenson, R.,
McLure, H., Campbell, W. E., Huddleston, J., Polliotti, B., & Nahmias,
A. (Emory Univ. School of Med., Pediatric Infectious Diseases, 69 Butler St, SE, Atlanta, GA 30303). Journal of Medical Primatology,
1993, 22, 393-399.
. . Fast scan magnetic resonance imaging, color Doppler ultrasonography, and magnetic resonance angiography appear suitable for noninvasive monitoring of the effects of drugs on placental function.
*Methods for timing of pregnancy and monitoring of fetal body and
brain growth in squirrel monkeys. Logdberg, B. (Inst. of Environmental
Med., Univ. of Lund, Solvegatan 21, S-223 62 Lund, Sweden). Journal
of Medical Primatology, 1993, 22, 374-379.
. . Tests were made of various combinations of vaginal examination,
bioassay of chorionic gonadotropin, measurement of maternal body
weight and uterine fundus height, and determination of fetal biparietal diameter by ultrasonography and magnetic resonance imaging. The
usefulness of these methods in developmental and teratological studies
is discussed.
*Cryopreservation of vervet monkey semen and recovery of progressively
motile spermatozoa. Seier, J. V., Conradie, E., Oettle, E. E., &
Fincham, J. E. (Experimental Biology Program: Primate Unit, MRC Tygerberg, P.O. Box 19070, Tygerberg 7505, South Africa). Journal of
Medical Primatology, 1993, 22, 355-359.
. . A method of cryopreserving semen from Cercopithecus aethiops has
been developed, yielding a mean post-thaw recovery of 63.6% of prefreeze progressive motility.
*New method for in vivo recording of myometrial activity in the squirrel monkey, Saimiri sciureus. Crane, L. H., Kuehl, T. J., & Dukelow,
W. R. (W. R. D., Endocrine Res. Center, Michigan State Univ., East
Lansing, MI 48824). American Journal of Primatology, 1992, 28, 73-78.
. . Videolaparoscopy was used to visualize reproductive tract contractility. Frequency, direction, and duration of contractions can be
directly observed with this method.
*[Annual sperm concentration variation in semen collected by electroejaculation in the cynomolgus monkey (Macaca fascicularis).] Okamoto,
M. (Nat. Inst. of Radiological Sci., 4-9-1 Anagawa, Inage-ku, Chiba-shi, Chiba 263, Japan). Experimental Animals, 1994, 43, 25-31.
(Japanese with English summary).
. . Results suggest that annual variation in sperm concentrations of
semen collected by electroejaculation is due to differences in the
contraction of muscles adjacent to the accessory genital gland and
ampulla elicited by electrical stimulation.
*Exteriorized chronic aorto-caval arteriovenous access shunts in the
baboon (Papio cynocephalus). Bass, A., Krupski, W. C., Hanson, S.
R., Dodson, T., Lumsden, A., White, D., Harker, L. A., & Kelly, A. B.
(A. B. K., Div. of Hematology & Oncology, Emory Univ. School of Med.,
P.O. Drawer AR, Atlanta, GA 30322). Journal of Medical Primatology,
1993, 22, 331-339.
. . A chronic arteriovenous blood access shunt composed of silicone
rubber tubing extensions bonded to vascular graft polytetrafluoroethylene cuffs, for creating anastomoses with the distal aorta and
inferior vena cava, remained patent for 9 (+/- 2.5) months.
Pharmacology & Anesthesia
*[Changes in cerebral metabolism and cerebral blood flow under Halothane and Isoflurane in increasing concentrations.] Brussel, T., Van
Aken, H., & Fitch, W. (Klinik u. Poliklinik fur Anesthes. u. operative
Intensivmed., Westfalische Wilhelms-Univ. Munster, Albert-Schweitzer-Str. 33, 4400 Munster, Germany). Anasthesiologie Intensivmedizin Notfallmedizin Schmerztherapie, 1992, 27, 414-420 (German, with English summary).
. . In two studies on baboons, low concentrations of both anesthetic
agents decreased the cerebral blood flow parallel to the reduction of
the cerebral metabolic rate for oxygen. Higher anesthetic
concentrations led to an increase in cerebral blood flow despite a
further decrease of the cerebral metabolism.
Reproduction
*[In vitro fertilization using Japanese monkey (Macaca fuscata) eggs
and cynomolgus monkey (Macaca fascicularis)] frozen/thawed spermatozoa. Sankai, T., Shimizu, K., Cho, F., & Yoshikawa, Y. (Tsukuba Primate Center for Med. Sci., NIH of Japan, 1 Hachimandai, Tsukuba-shi,
Ibaraki 305, Japan). Experimental Animals, 1994, 43, 45-50 (Jap-
anese with English summary).
. . Sixteen of 18 mature oocytes were judged to have been successfully
fertilized based on the presence of a male pronucleus and/or second
polar body; 12 oocytes then cleaved; and three developed into the
6-cell stage.
*Vulnerability of placental antibody transfer and fetal complement
synthesis to disturbance of the pregnant monkey. Coe, C. L., Kemnitz,
J. W., & Schneider, M. L. (Dept of Psychology, Univ. of Wisconsin,
1202 W. Johnson St, Madison, WI 53706). Journal of Medical Primatology, 1993, 22, 294-300.
. . Maternal and fetal/infant antibody levels were assessed throughout
pregnancy and at birth to evaluate the prenatal transmission of IgG in
the rhesus monkey. A marked increase in IgG occurred primarily during
the last two weeks of pregnancy, resulting in low antibody titers in
premature infants. When gestation length was normal, placental transfer of IgG was resistant to both dexamethasone treatment and a prolonged period of prenatal stress during pregnancy, although prenatal
stress affected other aspects of infant development.
*Baboons adjust secondary sex ratio in response to predictors of sex-specific offspring survival. Wasser, S. K. & Norton, G. (Dept of Animal Health & Conservation & Res. Center, Nat. Zoo. Park, Smithsonian
Inst., Front Royal, VA 22630). Behavioral Ecology and Sociobiology,
1993, 32, 273-281.
. . Social (e.g., birth order) and ecological (January rainfall) conditions that predicted improved survivorship of a given sex were positively correlated with more births of that sex than of the other.
*Serotonergic influences on male sexual behavior of rhesus monkeys:
Effects of serotonin agonists. Pomerantz, S. M., Hepner, B. C., &
Wertz, J. M. (Dept of Physiology, Univ. of Pittsburgh School of Med.,
Pittsburgh, PA 15261). Psychopharmacology, 1993, 111, 47-54.
. . Evidence shows that copulatory behavior of rhesus monkeys is influenced by 5-HT receptor stimulation, the direction of the effect
depending on the subtype of 5-HT receptor being stimulated.
*Effects of an oral contraceptive on sexual behavior of chimpanzees
(Pan troglodytes). Nadler, R. D., Dahl, J. F., Gould, K. G., & Collins, D. C. (Yerkes RPRC, Emory Univ., Atlanta, GA 30322). Archives
of Sexual Behavior, 1993, 22, 477-500.
. . Data indicate that despite generally adverse effects of an oral
contraceptive (OC) on certain physiological and behavioral dimensions
of sexuality, social factors ultimately determined the degree of
response to the OC. This research identifies three specific areas
affected adversely by OCs (female genital tissues, sexual proceptivity, and sexual attractivity) and suggests that the individuals who
are most vulnerable to such effects of OCs are those with unusually
high sensitivity to hormones and those with relatively low sexual
activity or preexisting problems in sexual adjustment.
*Serum and urinary concentrations of sex hormones and genital swelling
during the menstrual cycle of the gibbon. Nadler, R. D., Dahl, J. F.,
& Collins, D. C. (Address same as above). Journal of Endocrinology,
1993, 136, 447-455.
. . Genital swelling increased during the follicular phase with
increasing concentrations of estradiol, reached maximal swelling in
association with the mid-cycle peaks in the estrogens and LH and began
detumescence with the initial increases in progesterone during the
luteal phase, making it a useful marker for monitoring progress of the
menstrual cycle and the presumptive time of ovulation.
*Sexual behavior and serum levels of prolactin, testosterone, and
estradiol in young and old rhesus males. Chambers, K. C. & Phoenix, C.
H. (Univ. of Southern California, Dept of Psychology, SGM 501, Los
Angeles, CA 90089-1061). Physiology & Behavior, 1992, 52, 13-16.
. . The correlation between prolactin levels and different measures of
sexual behavior were not significant for either old or young male rhesus monkeys.
*Male rank, reproductive behavior, and reproductive success in free-ranging rhesus macaques. Berard, J. D., Nurnberg, P., Epplen, J. T.,
& Schmidtke, J. (Caribbean PRC, Univ. of Puerto Rico, Med. Sci. Campus, P.O. Box 906, Punta Santiago, PR 00741). Primates, 1993, 34, 481-489.
. . Paternity assessment through DNA fingerprinting of one birth cohort
in a social group suggests that the effect of rank on male reproductive success is not a predictable correlation, but a conditional probability.
*Male dominance rank and reproductive success in primate groups. de
Ruiter, J. R. & van Hooff, J. A. R. A. M. (Ethology & Socioecology
Group, Padualaan 14, P.O. Box 80.086, 3508 TB Utrecht, Netherlands).
Primates, 1993, 34, 513-523.
. . In a study of wild M. fascicularis, a relatively strong correla
tion was found between rank and reproductive success, which is attributable to selective mating by the alpha male during fertile periods of
the females. Such clear results have not usually been seen, and the
differences in outcome in relation to study conditions, group sizes,
and possible differences between species are discussed.
*Male dominance and genetically determined reproductive success in the
mandrill (Mandrillus sphinx). Dixson, A. F., Bossi, T., & Wickings,
E. J. (Sub-Dept of Animal Behaviour, Univ. of Cambridge, Madingley,
Cambridge CB3 8AA, England). Primates, 1993, 34, 525-532.
. . Behavioral and genetic studies of a large, semi-free ranging mandrill colony in Gabon have demonstrated that clear-cut relationships
exist between male secondary sexual development, social dominance,
copulatory behavior, and reproductive success in this species.
*Mating strategy and reproductive success of male patas monkeys
(Erythrocebus patas). Ohsawa, H., Inoue, M., & Takenaka, O. (Primate Res. Inst., Kyoto Univ., Inuyama, Aichi 484, Japan). Primates,
1993, 34, 533-544.
. . Observation of wild patas groups in Cameroon showed that groups are
multi-male during the year of a takeover of the resident male's position, but single-male thereafter, until another takeover. Females do,
however, mate with males outside of their group.
-------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
* * *
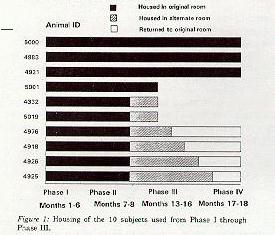{kind=link}
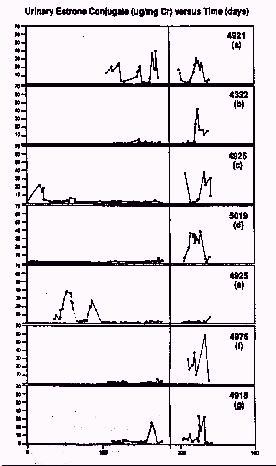{kind=link}
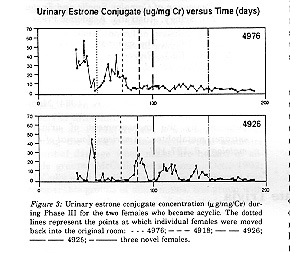{kind=link}
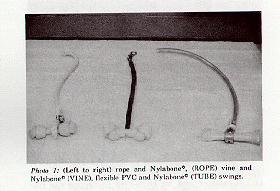{kind=link}
{kind=link}
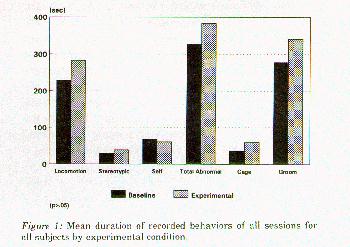{kind=link}
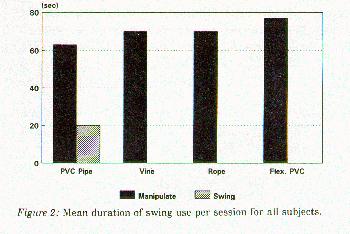{kind=link}
{kind=link}