 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
VOLUME 35 NUMBER 2 APRIL 1996
Articles and Notes
Successful Application of PCR-amplified DNA Markers for Paternity Determination in Rhesus Monkeys (Macaca mulatta) and Chimpanzees (Pan troglodytes), by J. J. Ely, M. L. Campbell, D. L Gonzalez, & W. H. Stone ...... 1
Toys for the Boys: Environmental Enrichment for Singly Housed Adult Male Macaques (Macaca fascicularis), by C. Brinkman ...... 4
Use of Two Food Foraging Devices by Singly Housed Baboons, by D. A. Pyle, A. L. Bennett, T. J.Zarcone, J. S. Turkkan, R. J. Adams, & R. D. Hienz ...... 10
News, Information, and Announcements
Travelers' Health Notes: Myiasis Warning Again ...... 4
Workshop Anouncements ...... 15
Primates in Art and Illustration ...... 16
Resources Wanted and Available ...... 16
Announcements From Publications ...... 17
. .
Journal of Medical Primatology; African Primates; ACLAD Newsletter;
JAAWS: New Animal Welfare Journal
Award Nominations: ASP Conservation Awards and Grants; W. W. Howells Prize ...... 18
Awards Granted: Whitehair Honored ...... 19
Addenda to the Directory of Graduate Programs in Primatology and Primate Research ...... 19
Grants Available ...... 20
. .
Conservation Biology Awards; Models for HIV and AIDS; Academic Research
Enhancement Award, Small Grants on Aging; Internet Connection; L. S. B.
Leakey Foundation Grants
Information Requested or Available ...... 22
. .
Hepatic Capillariasis; International Directory of Primatology --
1996; More WWW Addresses; Enrichment Device Ideas
Education and Volunteer Opportunities ...... 23
. .
Volunteer, Costa Rica; Wildlife Capture Course; Summer Intern -- Humane
Society; Wildlife Research Expedition, Peru; Program in Primate Behavior;
National Zoo Lecture Center; Primates of the World; University Research
Expeditions Program; Animal Technician, Texas
Meeting Anouncements ...... 25
. .
Animal Research in Industry; Ethical Issues of Animal Research;
International Society for Applied Ethology; New World Primate Taxon Advisory
Group; Primate Society of Japan; Canadian Association for Laboratory Animal
Science; National IACUC Affairs; Australian Primate Society; Harvard
Department of Anthropology; Primate-Talk Calendar
News Briefs ...... 28
. .
Univ. of Washington Press Release; Gorilla Birth at Los Angeles Zoo; Life
and Death at Woodland Park Zoo; Chimpanzee with AIDS Euthanized
Departments
Address Changes ...... 27
Positions Wanted or Available ...... 26
. .
Johns Hopkins; Animal Facility Supervisor, Athens, GA; Primate-Jobs;
Sabbatical Replacement, Kansas; Denver Zoo; Lefini Gorilla Manager
Recent Books and Articles ...... 30
* * *
John J. Ely, Misty L. Campbell, Daniel L Gonzalez, and William H.
Stone
Department of Biology, Trinity University
Introduction
Prior to the establishment of the NIH Regional Primate Research Centers in 1962, pedigreed nonhuman primates were not widely available, thus limiting the possibilities of genetic research. But now availability of approximately five rhesus generations and three chimpanzee generations affords many opportunities for biomedical research where genetically-defined animals are necessary, such as studies of transgenic animal models, models for human AIDS (Desrosiers, 1990), and complex (polygenic) human diseases (MacCluer, 1993; Stone, et al., 1987). The efficient use of animal models requires accurate pedigrees. This can best be accomplished by using genetic markers capable of corroborating and ascertaining parentage. In effect, genetic markers are indispensable tools for genetic management, allowing the formulation of rational breeding plans for maintaining genetic variation, minimizing inbreeding, and providing sound research strategies even when the focus is not genetics (Ely & Ferrell, 1992; Williams-Blangero, 1993). For example, genetic markers allow evaluation of the effects of common ancestry (i.e., shared genes) on experimental outcomes, since related animals may exhibit correlated responses for any trait under genetic control (Williams-Blangero, 1993). Thus, potentially misleading results are possible unless pedigrees have first been confirmed with genetic markers. Even with fastidious husbandry and record-keeping, up to 5.5% errors are common in primate pedigrees (VandeBerg, 1992).
Until recently, the genetic markers of choice for monitoring pedigrees included biochemical polymorphisms, blood types and lymphocyte antigens (Stone et al., 1993). Although still useful, these classical markers are neither as efficient nor as convenient as the newer DNA markers (Smith et al., 1992; Stone et al., 1993). Fortunately, there are nearly 4000 human-derived short tandem repeat (STR) DNA markers commercially available which can be screened to determine their utility for genetic typing of nonhuman primates. Here we report the use of human-derived STR markers amplified by polymerase chain reaction (PCR) for solving disputed parentage cases in rhesus monkeys and chimpanzees with very high efficiency.
Methods
Rhesus samples were obtained from animals housed at the University of Texas M. D. Anderson System Cancer Center, Bastrop, TX (Bastrop); the Wisconsin RPRC, Madison, WI; and the Laboratory of Comparative Ethology, NICHD, Poolesville, MD. Chimpanzee samples were obtained from Bastrop; the Primate Foundation of Arizona, Mesa, AZ; New Iberia Research Center, New Iberia, LA; and the Coulston Foundation, Holloman AFB, NM.
Five to 10 ml of whole blood were collected into vacuum tubes containing EDTA, an anti-coagulant, and shipped to our laboratory by overnight courier. High molecular weight DNA was extracted by a salting-out method (Miller et al., 1988) and resuspended in 500-1000 micro-l of full-strength buffering solution (10 mM Tris-Cl, 0.2 mM EDTA, pH 7.5) depending on yield. These samples were used as template DNA in PCR reactions using human-derived PCR primers obtained from Research Genetics (Huntsville, AL). Primers were randomly selected from all 22 human autosomes to determine which ones amplified rhesus or chimpanzee DNA. The goal was to identify primers which amplified homologous, polymorphic loci in these nonhuman primate species. Primers which amplified were optimized for MgCl2 concentration (1.5-3.5 mM) and buffer pH (8.5-10.0) using a commercially available PCR optimization kit (Invitrogen, PCR Optimizer). Otherwise, standard PCR cocktails consisted of 50 ng of each primer (forward and reverse), 200 micro-M of dATP, dGTP, dTTP, 2.4 micro-M of dCTP and 2.5 micro-M of 33P-dCTP, 0.5 U Taq polymerase (Promega), 50 mM KCl, 10 mM Tris-Cl, pH 8.3, and 15 ug template DNA in a reaction volume of 15 micro-l. PCR was performed with an initial 5 min denaturation at 94deg.C followed by 30 cycles of 1 min/94deg. denaturation, 45 sec/55deg. annealing, and 1.5 min/72deg. extension plus 2 sec autoextend per cycle, with a final extension of 10 min at 72deg.C. PCR reactions were performed on an Ericomp TwinBlock thermal cycler in 96-well microtiter trays with a mineral oil overlay. Four microliters of PCR product were diluted 1:2 with a denaturing loading dye (formamide/EDTA/xylene cyanol/bromophenol blue), loaded into 6% denaturing polyacrylamide gels and electrophoresed for 2.5 hr at 60W. Gels were dried for 30 min on a vacuum pump, wrapped in Saran wrap, then inserted in autoradiograph cassettes with Kodak XAR-5 X-ray film and allowed to expose at room temperature (22+/- 4deg.) for 3-4 days, before developing.
Since the alleles at STR loci are codominant, allele frequencies were determined directly by gene counting. Gene diversity (equivalent to expected heterozygosity) was estimated as one minus the sum of the squared allele frequencies (Nei, 1987). Paternity exclusion probabilities were caluculated using the generalized paternity exclusion index (Chakravarti and Li , 1983).
Results
In the process of screening 480 STR markers from human chromosomes I through X, we identified 381 primers which amplified rhesus DNA. Of these, 145 markers (38%) appeared to amplify homologous loci, as determined by similarity of the length of the amplified fragment to the known lengths of the human alleles. From these, we selected 14 STR markers for paternity testing which were polymorphic, as indicated by tests on a sample of five unrelated rhesus, and which exhibited Mendelian segregation in an 8-member, 3-generation rhesus pedigree. This battery of markers was then studied for its application to paternity testing. As shown in Table 1, each locus exhibited from 2 to 7 alleles (mean = 3.6), with gene diversity between about 20% to about 78% (mean = 58%). The estimated mean paternity exclusion probabilities for these 14 markers ranged from 0.089 to 0.737 (mean = 0.356), with a combined mean exclusion probability, PE, of 0.99999. In other words, the probability that a false sire could be correctly excluded by one or more of these 14 markers is nearly 100%.
We used these 14 STR markers in an attempt to resolve paternity cases. First we used our "progressive paternity exclusion" strategy to reduce the number of putative sires by testing several biochemical polymorphisms including the transferrins (Stone et al., 1993). After that, we ended up with 16 paternity cases involving 37 putative sires, with up to five sires per case. Thirteen of the 16 paternity cases (81%) were definitively resolved using the STR markers, by excluding all putative sires, except one, at two or more loci. We concluded that the the remaining putative sire, who was not excluded at any locus, was indeed the true sire.
Locus Number Gene P(e)1
(Marker) of Diversity
Alleles
D2S175 3 0.623 0.515
D7S559 7 0.648 0.487
D3S1271 3 0.494 0.290
GAAT1D9 3 0.426 0.216
GATA2A01 2 0.459 0.177
D8S271 4 0.691 0.720
D9S197 2 0.198 0.089
D1S194 3 0.560 0.301
D1S215 5 0.781 0.737
D1S306 5 0.580 0.508
D6S266 4 0.720 0.567
D2S102 4 0.660 0.698
D6S284 3 0.640 0.347
GATA11A07 3 0.540 0.273
Mean 3.6 0.580 0.356
Cumulative --- --- 0.99999
1. Mean exclusion probability
Table 1: STR loci (markers) used in rhesus monkey paternity determination.
Locus Number Gene P(e)1
(Marker) of Diversity
Alleles
D1S1679 5 0.722 0.861
D2S177 6 0.750 0.646
D8S1130 2 0.486 0.184
D8S1477 3 0.611 0.326
D9S921 4 0.514 0.318
D10S1435 4 0.653 0.820
D10S674 3 0.625 0.331
D11S1984 7 0.819 0.711
D11S1986 3 0.486 0.243
D13S788 4 0.625 0.784
D13S325 7 0.764 0.692
D15S569 3 0.569 0.305
Mean 4.3 0.635 0.518
Cumulative --- --- 0.99998
1. Mean exclusion probability
Table 2: STR loci (markers) used in chimpanzee paternity determination.
We studied another set of 35 human STR markers for use in chimpanzee paternity cases. Of these, eight (23%) did not amplify. However, 27 primers (77%) amplified a chimpanzee DNA fragment in the expected size range: 19 (70%) detected polymorphic loci, while eight of these (30%) detected monomorphic loci. As shown in Table 2, 12 of these 19 polymorphic markers exhibit between two and seven alleles each (mean = 4.3), with gene diversity between about 49% and about 76% (mean = 63.5%). Locus-specific mean exclusion probabilities range from 0.184 to 0.861 (mean = 0.518), with a cumulative mean exclusion probability of 0.99998. These 12 markers were used in eight paternity cases involving a total of 23 putative sires, with up to four putative sires per case. Six of the eight paternity cases (75%) were unequivocally resolved using this battery of STR markers.
Discussion
To our knowledge, this is the first report of the successful use of PCR-amplified human STR markers for routine paternity testing in nonhuman primates. Human-derived STR markers have been used in other studies involving nonhuman primates, including population history, individual identification, and phylogenetics (Bowcock et al., 1994; Deka et al., 1994; Garza et al., 1995; Morin et al., 1994; Pascall et al., 1994). The application of STRs to parentage determination in nonhuman primates has been suggested, but not previously reported, as part of routine colony maintenance and genetic management (Garza et al., 1995; Morin et al., 1993). Other researchers reported the cloning of species-specific STR markers (Takenaka et al., 1993), but this labor-intensive approach yielded a small number of markers which limited their use in other species and their application to genetic studies such as linkage.
We have demonstrated that human-derived STR markers can be applied to routine paternity testing in rhesus monkeys and chimpanzees with a high degree of success. These STR markers are much more efficient than the classical genetic markers. For example, the three most useful classical markers routinely used in our laboratory (transferrin, carbonic anhydrase II, diaphorase I) averaged 4.7 alleles per locus, but only 38% average heterozygosity. More importantly, the mean exclusion probability was 0.25559, with a cumulative exclusion probability of only 0.61030. In contrast, using our battery of 14 STR markers, we were able to attain a cumulative mean exclusion probability of 0.99999 in rhesus monkeys. Similarly, the 12 STR markers used in chimpanzees gave a cumulative mean exclusion probability of 0.99998. We anticipate that our ability to solve paternity cases will become even more powerful as we identify more polymorphic STR markers. We also expect to reduce the amount of time and cost involved in paternity testing by developing multiplexed PCR reactions by identifying three to four STR markers, easily distinguishable by their different allele sizes, to run in the same lane on a single gel.
Finally, it is important to understand why genetic tests sometimes fall short of the theoretical expectations. We typed 37 putative sires involved in 16 rhesus paternity cases with up to 14 STR markers each. Each marker defines a DNA type, or genotype, at a single locus. Among the 37 putative sires, we defined 147 genotypes which enabled us to exclude 27 putative sires and make 13 paternity determinations. On average, 5.4 (147/27) genotypes were needed per exclusion, while 11.3 (147/13) genotypes were required per case. However, the remaining three disputed parentage cases, which were refractory to resolution despite using many polymorphic markers, clearly involve animals who share more than the expected number of alleles. Many shared alleles can indicate common descent, that is, inbreeding. We are not the first to infer that a very high exclusion index sometimes translates into a smaller number of conclusive paternity determinations due to inbreeding (Smith et al., 1992). Even highly efficient DNA markers cannot compensate for lack of sound breeding practices as part of routine colony maintenance.
References
Bowcock, A. M., Ruiz-Linares, A., Tomfohrde, J., Minch, E., Kidd, J. R., & Cavalli-Sforza, L. L. (1994). High resolution of human evolutionary trees with polymorphic microsatellites. Nature 368, 455-457.
Chakravarti, A. & Li, C. C. (1983). The effect of linkage on paternity calculations. In R. H. Walker (Ed.), Inclusion Probabilities in Parentage Testing (pp. 411-422). Arlington, VA: American Association of Blood Banks.
Deka, R., Shriver, M. D., Yu, L. M., Jin, L., Aston, C. E., Chakraborty, R., & Ferrell, R. E. (1994). Conservation of human chromosome 13 polymorphic microsatellite (CA)n repeats in chimpanzees. Genomics 22, 226-230.
Desrosiers, R. C. (1990). Simian immunodeficiency viruses. Annual Review of Immunology 8, 557-578.
Ely, J. & Ferrell, R. E. (1992). Use of genetic markers in the genetic management of chimpanzees. In J. Erwin & J. L. Landon (Eds.), Chimpanzee Conservation and Public Health (pp. 119-126). Rockville, MD: Diagnon Corp.
Garza, J. C., Slatkin, M., & Friemer, N. B. (1995). Microsatellite allele frequencies in humans and chimpanzees, with implications for constraints on allele size. Molecular Biology and Evolution 12, 594-603.
MacCluer, J. W. (1993). Applications of pedigree analysis to animal models for complex diseases. In C. F. Sing & C. L. Hanis (Eds.), Genetics of Cellular, Individual, Family and Population Variability (pp. 122-139). New York: Oxford University Press.
Miller, S. A., Dykes, D. D., & Polesky, H. F. (1988). A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Research 6, 1215.
Morin, P. A., Wallis, J., Moore, J. J., Chakraborty, R., & Woodruff, D. S. (1993). Non-invasive sampling and DNA amplification for paternity exclusion, community structure, and phylogeography in wild chimpanzees. Primates 34, 347-356.
Morin, P. A., Moore, J. J., Chakraborty, R., Jin, L., Goodall, J., & Woodruff, D. S. (1994). Kin selection, social structure, gene flow and the evolution of chimpanzees. Science 265, 1193-1201.
Nei, M. (1987). Molecular Evolutionary Genetics. NY: Columbia University Press.
Pascall, V. L, Bisol, G. D., Dobosz, M., d'Aloja, E., Paonessa, G., & Mereu, U. (1994). Chimpanzee DNA profiles on trial. Nature 367, 692-693.
Smith, D. G., Rolfs, B., & Lorenz, J. (1992). A comparison of the success of electrophoretic methods and DNA fingerprinting for paternity testing in captive groups of rhesus macaques. In R. D. Martin, A. F. Dixson, & E. J. Wickings (Eds.), Paternity in Primates: Genetic Theories and Tests (pp. 32-52). Basel: Karger.
Stone, W. H., Treichel, R. C .S. & VandeBerg, J. L. (1987). Genetic significance of some common primate models in biomedical research. In Animal models: Assessing the Scope of Their Use in Biomedical Research (pp. 73-93). New York: A. R. Liss, Inc.
Stone, W. H., Ely, J. J., Manis, G. S., & VandeBerg, J. L. (1993). Classical genetic markers & DNA markers: A commensal marriage. Primates 34, 365-376.
Takenaka, O., Takasaki, H., Kawamoto, S., Arakawa, M., & Takenaka, A. (1993). Polymorphic microsatellite DNA amplification customized for chimpanzee paternity testing. Primates 34, 27-35.
VandeBerg, J. L. (1992). Biochemical markers and restriction fragment length polymorphisms in baboons: Their power for paternity exclusion. In R. D. Martin, A. F. Dixson, & E. J. Wickings (Eds.), Paternity in Primates: Genetic Theories and Tests (pp. 18-31). Basel: Karger.
Williams-Blangero, S. (1993). Research-oriented genetic management of nonhuman primate colonies. Laboratory Animal Science 43, 535-540.
* * *
In the July, 1993 issue of the LPN, we printed a report of myiasis (the invasion of human or animal tissue by the larvae of flies) in a scientist who had been conducting research in the rain forest of Costa Rica. In the December, 1995 issue of Missouri Medicine (92[12], 764-765), K. J. Mott & P. C. Anderson report on a zoologist who returned from a field trip in a Peruvian rainforest with two Dermatobia hominis in the skin of his hand. Physicians must be aware of their patients' travel histories!
"In Central and South America, D. hominis is the most common intruder into the skin of explorers. The infestation begins insidiously after the eggs are deposited on the skin of the unaware host. The comforting warmth of the skin stimulates the eggs to hatch. The larvae then burrow into the skin to complete development before reemerging approximately 50 days later.
"Dermatologically, soon after penetration of the skin by the larva, a small 2-3 mm papule develops that is not likely to be noticed in a bug-ridden jungle. The tiny papule soon enlarges into a dome-shaped nodule, at which point the clinical appearance may resemble a furuncle, particularly when the lesion becomes pointed as the posterior end of the larva protrudes from the wound. An intermittent foul-smelling discharge, commonly mistaken for `pus', is actually sloughed larval remnants. Severe piercing pain, a reliably classic symptom of myiasis, is attributed to the tearing of the host's tissues by the oral hooks of the larva during feeding. Removal of the larva is in itself curative."
* * *
Cobie Brinkman
Division of Psychology and Centre for Visual Science, Australian
National University
Introduction
In the last ten years, ever since providing for the "psychological well-being" (PWB) of captive nonhuman primates (Animal Welfare Act, 1985) was first demanded, "environmental enrichment" has been considered an essential part of such provision. No agreed-upon definition of PWB exists because it is likely to be specific to both the species (Clarke & Mason, 1988; Thomas & Lordon, 1989) and the individual (Boccia et al., 1995); but a reasonable degree of PWB is assumed when an animal is in good health, its behavioral repertoire approaches that seen in wild populations, and abnormal behaviors are reduced or absent (Novak & Suomi, 1989). Environmental enrichment (EE) may thus be defined as any measure which promotes expression of natural, species-specific behaviors and a decrease in, if not disappearance of, abnormal behaviors.
An impressive list of EE devices, toys, and husbandry procedures can be collated from the literature (e.g., Reinhardt & Reinhardt, 1992), and several books have appeared (Novak & Petto, 1991; Segal, 1989). But there are problems. Many toys have no attraction beyond their novelty value (Fazji et al., 1989; Reinhardt & Reinhardt, 1992), and some devices such as food puzzles require a considerable investment of human labor and thus, money (Moazed & Wolff, 1988). Regarding husbandry procedures, because macaques are social animals, housing of previously singly caged macaques in pairs or in groups appears to conform to the definition of PWB, and can produce long-lasting results, especially for females or for adults paired with an infant (Reinhardt, et al., 1995). However, pairing or grouping of adult males has often proved to be difficult because of aggression (Crockett, et al., 1994), and such pairings may be easy only when animals are related, have grown up together, or when there is a considerable age difference, as in adult males and infants (Reinhardt et al., 1995). For males in particular, it may be unreasonable to assume that for every monkey in captivity, a compatible partner exists and that this partner can be found, as pointed out by Crockett (1993).
Because of increased captive breeding, more males will be available for research. Because of experimental demands, such animals may have to be housed singly but may be involved in experiments for many months. To provide for PWB for such single male macaques is a formidable challenge to animal house staff and researchers. Few papers deal with the specific problems of a small number of animals actively involved in research. This paper describes some toys, devices, and procedures which have been found to be useful (or not!) for adult male macaques housed singly for a number of years while involved in behavioral research into control of movement.
The Boys
Five male longtailed macaques (Macaca fascicularis) provided data for this paper. All were captive-born, weaned between 6 and 12 months of age, then reared in a mixed-sex peer group with visual and auditory contact with the natal group and, when 2 to 3 years old, transferred to an all-male group of juveniles. Two monkeys were half-brothers from the same natal group, the others were unrelated. All had been housed singly for short periods (2 to 3 weeks), and all but one spent up to three years as breeder males before the start of this study, at which time they were aged between 6 and 13 years and weighed 6 to 10 kg. Several times/week, they took part, for 1 to 2 hours, in different tests of motor behavior for food rewards. These experiments lasted for 2 to 3 years. Housing, cage furniture, toys and devices, and foods, the most common sources of EE (Bayne, 1991), will be presented below. All were used for periods of 1 to 3 years, and some for 4 to 5 years, until the present day. All aim at extending the "psychological space" (Chamove, 1989) of an animal with restricted physical space.
The Toys
In Australia, few laboratories use nonhuman primates. Therefore, the National Health and Medical Research Council's Policy on the Use of Non-human Primates in Medical Research (1995) is not as prescriptive as the U.S. equivalent. Expected is an environment that "meets the behavioural demands of the species", allows the animal to "display species-specific behaviours" and, preferably, "appropriate social groupings". On the other hand, contact with conspecifics must be provided when adult males require single housing because of aggression.
Housing: There is no prescribed cage size or shape, and cages are usually built locally. The males of this study were housed in cages with a floor area of 95 x 100 cm (appr. 3'1" x 3'3"), and a height of 110 cm (3'8", inside dimensions). Walls and roof were made of 1" x 2" galvanized steel mesh while vertical steel bars, 5 cms (2") apart, were used for the front, which included a 40 x 40 cm (1'4") sliding door. The floor was made of stainless steel 5 x 5 cm mesh over a removable, stainless steel sawdust tray (Figure 1). The cage was mounted on a 50 cm (1'8")-high trolley. Cages were kept in sets of 5, 4, 3, or 2, so animals could see and hear each other, in conventional animal holding rooms. Compatible animals, such as the two half-brothers, were allowed tactile contact as well, by placing the cages with the fronts facing each other. However, very little contact, such as allogrooming, was ever seen, and clear dominance rankings existed for all animals despite single housing.
Figure 1: Monkey in cage with toys.
Cage furniture: Each cage was fitted with a perch 40 cm (1'4") below the roof, made of two hardwood slats pushed through the mesh and held in place by large drop-in bolts (Figure 1). Perches were preferred for eating, resting, and sleeping. Perches that were easier to clean were made of 5 cm diameter PVC piping (orange conduit or bright yellow gas pipe), secured in the same way. Animals spent more time on perches made of two pipes, 5 cm apart, than on a single-pipe perch. Each cage also had one or two sun-dried hardwood branches (Figure 1). These were not secured. The animals would sit on them, and three of the animals liked "rearranging the furniture", handling and lifting the branches.
Toy or device Manipu- Use by Perceived
lation animals Benefit
------------------------------------------------
Radio No Not -
applicable
Balls Yes ST -
Bones (rawhide) Yes ST -
Mirror, metal, Yes ST +
suspended
Mirror, silvered No LT, I +
fixed on wall
Plastic rings Yes LT, I +
Phonebooks Yes LT, I +
Cans Yes LT, I ++
Lids of cans Yes LT, I ++
Rope Yes LT, P ++
Fishing line Yes LT, I +++
reels
Coconuts Yes LT, I +++
Cardboard (boxes Yes LT, I +++
and tubes)
Food tubes Yes LT, I +++
Wood `chewing' Yes LT, P +++
barbell
Garden hose Yes LT, P +++
Plastic bottle Yes LT, P +++
(suspended)
Plastic bottle Yes LT, P +++
(in cage)
Explanation of abbreviations and symbols: ST: short term, novelty value only;
LT, I: Long term (years) if given intermittently; LT, P: can be used
continuously, for years; -: no real benefit; +: used by minority, 1/5 or 2/5;
++: used by majority, 3/5 or 4/5; +++: used by 5/5 monkeys.
Table I: Toys and devices assessed for longterm use and benefit.
Toys and devices: A number of toys and devices have been tried. If possible, they had to be cheap, cleanable (cleaned toys were given back to the original owner to prevent them from acting as fomites, cf. Bayne et al., 1993), and require little extra human labor. They are listed in Table I. It can be seen that some, such as balls, mirrors and rawhide bones (Eaton et al., 1993), were of little interest to the animals; others were used by at least some monkeys for long periods. The latter include 3 cm
(1.25") plastic rings cut from 40 cm (1'4") diameter PVC pipe; large infant formula or instant coffee cans and their metal lids, which animals would use as hand-held mirrors; thick cotton horse-lead ropes clipped to the cage, which were chewed on, and which at least one animal would thread across and around perch and branches; and coconuts (Taff & Dolhinow, 1989), which were groomed bare if not broken first. Very successful toys, used extensively by all animals, were cardboard boxes or tubes and cedar wood in the shape of barbells. Pine was found to be too soft and hardwoods too hard, but cedar was slowly chewed into wood chips by the animals (which did not seem to ingest any wood, or suffer upsets if they did). Lengths of garden hose, threaded through one or more cages, were used for manipulation, climbing, and chewing (again without apparent ingestion or adverse effects). Brightly colored plastic fishing line reels were hung on hoses and ropes for more manipulation and chewing. By far the most successful enrichment was plastic two-liter milk bottles, or soft drink, juice, or syrup bottles; not rinsing the latter before giving them to the monkeys added extra flavor to the animals' experience. Bottles were either suspended from the cage front with a clip and short chain, or put inside the cage. Animals would manipulate them, throw them, chew them, and tear them into pieces. Clear (PET) bottles were preferred (apparently because they make more noise when handled) over softer, opaque bottles, and monkeys would play for days even with PET pieces. Labels were systematically "groomed" off. Sometimes monkeys were given empty shampoo bottles with caps removed but only lightly rinsed. The olfactory stimulus appeared to be of great interest because the animals carried the bottles around, sometimes for hours, before destroying them. Plastic bottles have retained their appeal for over four years now.
Food: Until recently, no primate chow was produced in Australia. Instead, animals were fed 4 to 8 ounces per day of a mix made up of commercial rat, rabbit, goat, and sheep pellets, corn kernels, and black, unshelled sunflower seeds. Because the species is "crab-eating" in the wild, cat biscuits were added. The mix was supplemented daily by a medium apple, half a banana, and some cabbage. The mix could be put into the plastic bottles, with a cut-off large funnel, or mixed into (sterilized) sawdust in the tray or a large horse feed bin hung at the front of the cage. Animals would forage for long periods in the tray, but not in the bin. Breakfast cereals or molassed horse grain mix were sometimes substituted. Other fruits and vegetables were often used. The animals have sampled just about anything grown in this country and apparently enjoyed it, with the exception of some pungent (onions, leeks) or bland (lettuce, commercial tomato and strawberries) foods. Fruits and vegetables were cut up and placed on top of the cage. The animals could retrieve them by pushing them to one of a few 5 x 5 cm holes cut into the ceiling to make it into a large "puzzle-box" (cf Reinhardt, 1993). To prevent food falling onto the floor, a 5 cm-high barrier was erected around the cage top (Figure 1). Corn kernels or sunflower seeds were sometimes given in a PVC pipe with a screw top at one end, easily made from plumbing supplies. Small holes drilled in the pipe allowed food to fall out when the tube was rolled or handled. Browse was also given: poplar or willow branches in summer, lucerne hay in winter.
Discussion
Because "psychological well-being" has no clear definition, its assessment is problematical (Novak et al., 1995; Novak & Suomi, 1989; Thomas & Lordon, 1989) and, therefore, so are the effects of enrichment. The latter usually are measured as decreases in nonfunctional, abnormal behaviors, such as stereotypies, and increases in species-specific behaviors, or in time spent with environmental enrichment devices (e.g., Bryant, et al., 1988); but not all studies present quantitative data (see reviews by Fajzi et al., 1989; Reinhardt & Reinhardt, 1992). In the present study, animals were free from abnormal behaviors as they changed from social to single housing, unlike the rhesus studied by Goosen (1989), possibly because they had been in single cages several times before and, therefore, did not experience the change as stressful. Avoidance of stress, should single housing be necessary, either for experimental or medical reasons, was, in fact, the aim of having periods of single housing as part of a captive-born monkey's standard "education". Also, from the start, animals were provided with EE and involved in the experimental training program, which in itself can be enriching (Laule, 1993). After two to three years, no abnormal behaviors had developed (Chamove, 1993, James, 1993) and the animals were judged to behave in a manner appropriate to their species, age, and sex (James, 1993). No formal quantitative data are available, but because of the long periods of time during which the same animals have been kept in single housing, and during which these cheap toys, devices, and procedures have been tried and retried, others faced with the problems of singly housed social animals may find this qualitative evaluation of use. No other studies have reported on EE exposure for such long periods (Reinhardt & Reinhardt, 1992). It should also be noted that the animals used here are longtailed macaques. This species appears to be more susceptible to stress than the rhesus monkey (Clarke & Mason, 1988). The use of this species is increasing and its EE needs, as compared to those of rhesus monkeys, should be further explored.
Although cage size is not strongly correlated with well-being when assessed over short periods (Crockett et al., 1993; Crockett and Bowden, 1994; Line et al., 1991), when animals are kept in them for a number of years it may become important. The size of the cages used here allowed the animals both walking and climbing space, and they could sit on the perch without their tails touching the ground (Figure 1). The cage has almost twice the floor area and is 50% higher than the U.S. equivalent for animals this size (USDA, 1991). It is larger than the European standard range, and close to that of the UK (Poole & Hubrecht, 1994). While this size may still not be adequate, it has kept the animals in good physical condition. When transferred to their present quarters, where space is increased four to eight times, the motor weakness and incoordination seen by others (O'Neill, 1989), when conventionally singly caged animals were given access to a large exercise area, were absent.
An inspection of Table I reveals that successful toys are ones that can be manipulated and carried around. Shape also matters (Mench, 1994): monkeys all but ignored plastic balls (cf Line and Morgan, 1991) but did not tire of a variety of plastic bottles. All used wooden barbells intensively. This contrasts with the variable use by rhesus monkeys of wooden sticks or branches (Champoux et al., 1987; Line & Morgan, 1991), and virtual ignoring of wooden blocks by longtailed macaques (Brinkman, unpublished observations). The importance of texture is reflected in a preference for cardboard boxes or tubes over phone books, and the interest shown in coconuts (cf. Taff & Dolhinow, 1989) compared to simple balls. Use of colored piping, hose, rope, and reels may provide a more pleasing surround for trichromatic nonhuman and human primates. The interest shown in fragrant bottles indicates that olfaction, hitherto largely neglected as enrichment, warrants further attention. Auditory stimulation is almost equally neglected. The monkeys appeared to ignore a radio (cf. O'Neill, 1989), except when attendants switched stations; but when a radio can be controlled by the animals, it is more effective (Line et al., 1991). In the present study, monkeys seemed to use cans and PVC food tubes to create noises by rolling them against the mesh or dropping them repeatedly.
Common to all toys which never lost appeal (hoses, cardboard tubes, bottles, wooden barbells) was that animals could chew and tear them. Such chewing could be a form of stereotypy which cannot be indulged in conventional single caging (except for bar-biting, a behavioral abnormality seen in some domestic animals; see Broom & Johnson, 1993, p.77). However, such behavior was never seen in these monkeys during previous periods of single-caging, and animals did not resort to it in association with stressful events, e.g., when changing cages, when visitors were present, or during other procedures known to be stressful (Crockett et al., 1993; Line et al.,1991). Rather, the oral exploring, chewing, and tearing may represent behaviors normally associated with foraging and food collection in the wild: free-ranging macaque populations are notoriously destructive. In zoos, their environment soon becomes bare unless specially protected (Coe, 1989). The opportunity to chew and tear has also benefitted the animals' dental health: no animal has required any cleaning of its teeth in the past few years.
Offering different kinds of food provides motor, sensory, olfactory and gustatory variety. Often, foods and fruits from gardens (and bottles of all kinds) have been brought in by attendants and other researchers. Their increased awareness and appreciation of the monkeys' behavioral needs, generated by the enrichment program, has been one of the most valuable outcomes, since the animals continue to benefit (Petto, et al., 1992).
References
Bayne, K. (1991). Alternatives to continuous social housing. Laboratory Animal Science, 41, 355-359.
Bayne, K. A. L., Dexter, S. L., Hurst, J. K., Strange, G. M., & Hill, E. E. (1993). Kong toys for laboratory primates: Are they really an enrichment or just fomites? Laboratory Animal Science, 43, 78-84.
Boccia, M. L., Laudenslager, M. L., & Reite, M. L. (1995). Individual differences in macaques' responses to stressors based on social and physiological factors: Implications for welfare and research outcomes. Laboratory Animals, 29, 250-257.
Broom, D. M. & Johnson, K. G. (1993). Stress and Animal Welfare. London: Chapman and Hall.
Bryant, C. E., Rupniak, N. M. J., & Iverson, S. D. (1988). Effects of different environmental enrichment devices on cage stereotypies and auto-aggression in captive cynomolgus monkeys. Journal of Medical Primatology, 17, 257-269.
Chamove, A. S. (1989). Assessing the welfare of captive primates -- a critique. In Laboratory Animal Welfare Research -- Primates (pp. 39-49). Potters Bar, UK: Universities Federation for Animal Welfare.
Chamove, A. S. (1993). Non-Human Primates at the Australian National University. Report to the Deputy Vice Chancellor, ANU.
Champoux, M., Hempel, M., & Reinhardt, V. (1987). Environmental enrichment with sticks for singly-housed adult rhesus monkeys. Laboratory Primate Newsletter, 26[4], 5-7.
Clarke, A. S. & Mason, W. A. (1988). Differences in three macaque species in responsiveness to an observer. International Journal of Primatology, 9, 347-364.
Coe, J. C. (1989). Naturalizing habitats for captive primates. Zoo Biology, Supplement 1, 89-100.
Crockett, C. (1993). Primate well-being is not promoted by suit. Laboratory Primate Newsletter, 32[2], 1-2.
Crockett, C. M. & Bowden, D. M. (1994). Challenging conventional wisdom for housing monkeys. Laboratory Animal, 24[2], 29-33.
Crockett, C. M., Bowers, C. L., Bowden, D. M., & Sackett, G. P. (1994). Sex differences in compatibility of pair-housed longtailed macaques. American Journal of Primatology, 32, 73-94.
Crockett, C. M., Bowers, C. L., Sackett, G. P., & Bowden, D. M. (1993). Urinary cortisol responses of longtailed macaques to five cage sizes, tethering, sedation, and room change. American Journal of Primatology, 30, 55-74.
Eaton, G. G., Kelley, S. T., & Iliff-Sizemore, S. A. (1993). Rawhide "chew-bones" reduce abnormal behavior in individually-housed adult rhesus macaques. American Journal of Primatology, 30, 308.
Fajzi, K., Reinhardt, V., & Smith, M. D. (1989). A review of environmental enrichment strategies for singly caged nonhuman primates. Laboratory Animal, 18[2], 23-35.
Goosen, C. (1989). Influence of age of weaning on the behaviour and well-being of rhesus monkeys. In Laboratory Animal Welfare Research -- Primates (pp. 17-22). Potters Bar, UK: Universities Federation for Animal Welfare.
James, A. E. (1993). A Report on the Housing Conditions of the Non-Human Primate Colony at the Australian National University. Report to the National Health and Medical Research Council.
Laule, G. (1993). Addressing psychological well-being: Training as enrichment. Shape of Enrichment, 1, 12-13.
Line, S. W., Markowitz, H., Morgan, K. N., & Strong, S. (1991). Effects of cage size and environmental enrichment on behavioral and psychological responses of rhesus macaques to the stress of daily events. In M. A. Novak & A. J. Petto (Eds.), Through the Looking Glass: Issues of Psychological Well-being in Captive Nonhuman Primates (pp. 160-179). Washington DC: American Psychological Association.
Line, S. W. & Morgan, K. N. (1991). The effects of two novel objects on the behavior of singly caged adult rhesus monkeys. Laboratory Animal Science, 41, 365-399.
Mench, J. A. (1994). Environmental enrichment and exploration. Laboratory Animal, 23[2], 38-41.
Moazed, T. C. & Wolff, A. V. (1988). The raisin board as an environmental enrichment tool for laboratory primates. Laboratory Primate Newsletter, 27[1], 16.
National Health and Medical Research Council (1995). Policy on the Use of Non-human Primates in Medical Research. Unpublished draft.
Novak, M. A. & Petto, A. J. (Eds.). (1991). Though the Looking Glass: Issues of Psychological Well-being in Captive Nonhuman Primates. Washington DC: American Psychological Association.
Novak, M. A., Rulf, A., Munroe, H., Parks, K., Price, C., O'Neill, P. & Suomi, S. J. (1995). Using a standard to evaluate the effects of environmental enrichment. Laboratory Animal, 24[6], 37-42.
Novak, M. A. & Suomi, S. J. (1989). Psychological well-being of primates in captivity. ILAR News, 31[1], 5-15.
O'Neill, P. (1989). A room with a view for captive primates: Issues, goals, related research and strategies. In E. F. Segal (Ed.), Housing, Care and Psychological Well-being of Captive Laboratory Primates (pp. 135-160). Park Ridge: Noyes Publications.
Petto, A. J., Russell, K., Watson, L., & Lareau-Alves, M. (1992). Promoting psychological well-being in a biomedical research facility: Sheep in wolves' clothing. Humane Innovations and Alternatives, 6, 366-373.
Poole, T. & Hubrecht, R. (1994). Letter to the Editor. Laboratory Animal, 23[7], 51.
Reinhardt, V. (1993). Using the mesh ceiling as a food puzzle to encourage foraging behavior in caged rhesus macaques (Macaca mulatta). Animal Welfare, 22, 165-172.
Reinhardt, V., Liss, C., & Stevens, C. (1995). Social housing of previously single-caged macaques: What are the options and the risks? Animal Welfare, 4, 307-328.
Reinhardt, V. & Reinhardt, A. (1992). Quantitatively tested environmental enrichment options for singly-caged nonhuman primates: A review. Humane Innovations and Alternatives, 6, 374-383.
Segal, E. F. (Ed.). (1989). Housing, Care and Psychological Well-being of Captive Laboratory Primates. Park Ridge: Noyes Publications.
Taff, M. A. & Dolhinow, P. (1989). Langur monkeys (Presbytis entellus) in captivity. In E. F. Segal (Ed.), Housing, Care and Psychological Well-being of Captive Laboratory Primates (pp. 291-304). Park Ridge: Noyes Publications.
Thomas, R. K. & Lordon, R. B. (1989). What is psychological well-being? Can we know if primates have it? In E. F. Segal (Ed.), Housing, Care and Psychological Well-being of Captive Laboratory Primates (pp. 13-26). Park Ridge: Noyes Publications.
* * *
Danielle A. Pyle, Amy L. Bennett, Troy J. Zarcone, Jaylan S. Turkkan, Robert
J. Adams, and Robert D. Hienz
The Johns Hopkins University School of Medicine
Introduction
Since the advent of USDA regulations requiring research facilities to provide environmental enrichment as a means for promoting psychological well-being in nonhuman primates, numerous strategies for providing for psychological well-being have been proposed by the primate community (Bayne, 1991). In laboratory settings where experimental protocols require individual housing, one prevalent enrichment strategy has been to provide "enrichment devices" to promote psychological well-being (e.g., Fajzi et al., 1989). Little scientific data exists, however, to support the usefulness of these devices in increasing well-being in nonhuman primates, particularly for those housed singly (Bloomsmith et al., 1991).
A large variety of enrichment devices are available for use with nonhuman primates, including feeding devices to promote foraging-type behaviors (e.g., Bloom & Cook, 1989). Engaging in feeding behaviors accounts for most of a nonhuman primate's daily activity in the wild (O'Neill et al., 1989). In captivity, feeding devices appear to engage non-human primates for longer periods of time than non-food devices. Further, studies have shown a reduction in aberrant behaviors in rhesus monkeys (Bayne et al., 1991) and chimpanzees (Maki et al., 1989) when food devices are present. One could conclude that such feeding devices do indeed promote well-being, since engaging in species-typical behaviors such as grooming and foraging are thought to indicate an increase in well-being (Novak and Suomi, 1991). The purpose of the present study was to determine the effects of two foraging devices, P-nut Butter Rolls® and Prima-Foraging Grooming Boards®, on the behavior of singly caged baboons in an experimental laboratory setting.
Methods
Subjects: Six adult male baboons (Papio anubis and P. cynocephalus), weighing 21 to 32 kg, were individually housed in the same colony room in regulation Group 4 primate cages equipped with a seating bench and free access to water. All baboons were maintained on a 12-hr light/dark cycle (lights on/off at 6 a.m./ 6 p.m.). All baboons could see and hear each other at all times. During the course of this study, all baboons participated in daily behavioral signal detection experiments. Food intake was controlled at 200 g/day. About 90% of this amount was obtained during daily 100-min experimental sessions. Baboons performed a signal detection task in which each correct detection of an acoustic signal was rewarded with a 500-mg Bio-Servreg. pellet. The remaining food was given at the end of day in the form of Purina monkey chow, along with two pieces of fresh fruit. A physical health examination was conducted once a month. All animals received a tuberculin test by the veterinary staff every three months. Animal care was in accordance with current NIH guidelines concerning the humane treatment of nonhuman primates.
Apparatus: A P-Nut Butter Rollreg. (Bio-Environmental Modifiers, Frankfort, Kansas) and a Prima-Foraging Grooming Boardreg. with Fleece Padsreg. (Bio-Serv, Frenchtown, New Jersey) served as the enrichment devices. The P-Nut Butter Roll was a device with two concentric tubes. The innermost tube was free-moving about its axis and was typically coated with creamy peanut butter (approximately 1/4 cup); the outer tube revolved around the inner tube, and had six 2 in. long x 3/4 in. wide slots for accessing the peanut butter. The Prima-Foraging Grooming Board was a dual-plated autoclavable stainless steel device. A fleece pad (90% polyester, 10% acrylic) covered the board and the board was then attached to a bracing frame. The fleece pad was coated with a small drizzling of dark corn syrup and 1 cup of mixed flavors of Prima-Foraging Crumbles (Bio-Serv, Frenchtown, New Jersey) were pressed onto the pad.
To monitor the movements of these foraging devices, wireless activity monitors were attached to each device. Each activity monitor was a 5.5 x 3.3 x 1.5 cm (LWH) unit enclosed in a plastic case (Personal Activity Monitor, manufactured by Individual Monitoring Systems, Baltimore MD). Details of the monitor's use with nonhuman primates have been described previously (Hienz et al., 1992). The monitor's internal microprocessor records "activity counts" whenever the device undergoes any accelerative movement. A monitor is sensitive to accelerations exceeding 1 m/s/s. Each activity count indicates an above-threshold acceleration within a .22-sec interval. In the present study, these activity counts were summed over 15-min intervals (i.e., summed over 4096 consecutive .22-sec bins). An activity monitor was attached to the inside of the internal tube of each P-Nut Butter Roll, or was bolted to the back of each grooming board. Each foraging device was suspended on the outside of the cage by a chain so that touching the device caused the entire device to move. Both devices were accessible through the front of a cage, but could not be pulled into a cage. Activity monitors were removed every two weeks to retrieve the data.
To record the general movements of baboons, activity monitors were placed inside protective metal cases and the cases then mounted on soft leather neck collars (3/4-in wide) that were custom-fitted to each baboon's neck. Each animal could move the case freely along the collar. The collars are well-tolerated by baboons, and do not interfere with eating or restrict body movements (Hienz et al., 1992). Each animal wore a monitor for a minimum 2-week baseline period prior to introduction of the foraging devices.
General Procedures: Three weeks prior to the start of the experiment, all enrichment devices were removed from the cages, and human interactions were recorded. Technicians entered the room under the following conditions: 1) to perform daily maintenance (e.g., to clean pans, etc.); 2) to transfer an animal from its home cage to an experimental chamber for daily behavioral sessions; 3) to mount enrichment devices on cages; and 4) to deliver afternoon supplemental feedings (4:30 pm). During testing, all food devices were removed from the baboons' cages prior to daily behavioral sessions, and were subsequently loaded with food and placed back on all animals' cages every weekday at 3:15 pm, five minutes prior to the onset of video taping sessions. Because baboons did not perform in behavioral sessions on weekends, the devices were loaded and attached at varied times between 6:00 am and 1:00 pm on weekend days. All animals were exposed to the enrichment devices over a six-week period. For the first two weeks of testing, one of the two enrichment devices was placed in a baboon's cage; the device was then removed for two weeks, followed by presentation of the other device for the next two weeks. The order of presentation of the two foraging devices is shown in Table 1. Because feeding schedules differed on weekends, only weekday data were employed for the present analyses.
Data Collection/Analysis: Videotapes were collected via three closed-circuit video cameras (Panasonic, WV-1410 CCTV cameras) mounted to the ceiling and connected to a videotape recorder programmed for automatic taping. Each camera was oriented towards a different pair of baboons, and an automated switch box randomly selected which pair was videotaped at the following times: 3:20-3:40 pm, 3:45-4:05 pm, and 4:10-4:30 pm, Monday through Friday. Each pair of baboons was videotaped during one of these times each day. Samples of each baboon's behaviors thus were obtained soon after receiving a device. Behavioral scoring of the videotapes was performed with an Event Scoring System (Princeton Economics, Inc., Princeton, MA) using the behavioral categories presented in Table 2.
Animal Baseline Weeks Weeks Weeks Weeks
1-2 3-4 5-6 7-8
-----------------------------------------
AC - - PR - GB
BE - PR - GB -
CA - - GB - PR
DR - PR - GB -
FR - - PR - GB
WE - GB - PR -
Table 1: Weekly enrichment device schedule for all baboons. PR = P-Nut Butter Roll, GB = grooming board.
Behavior Definition
------------------------------------------------
Stereotypy Any pattern of movement repeated more
than two consecutive times.
Self-directed Any behavior directed towards an
behavior animal's own body (e.g., grooming).
Cage-directed Any behavior in which the animal
manipulated part of the cage (e.g.,
manipulating a door lock).
Bracing An animal supporting himself above
the floor of the cage and the bench
by holding the top or sides of the cage.
Toy play Any contact between an animal and the
enrichment device.
Locomotion Any movement not included in the above
categories.
Inactive No movement observed.
Table 2: Behavioral categories used for scoring video tapes.
Videotapes of all baboons were scored during both device-absent and device-present testing phases. Samples of the use of each foraging device were taken from a minimum of three video segments representing intervals A) immediately after device presentation during the first week, B) one hour after device presentation during the first week, and C) immediately after device presentation during the second week. Comparing data from segments A and B allowed an assessment of any changes in device use over the initial exposure period, while comparison of segments A and C allowed assessment of changes over the 2-week period. One observer scored all video segments. Reliability was assessed by having a second observer score 20% of the segments; reliability was greater than 80% for all segments scored by both observers.
Results
The recorded movements of the foraging devices are displayed in Figure 1, which shows 24-hr movement patterns for grooming boards (left column) and P-Nut Butter Rolls (right column) for each baboon. A peak in device activity occurred immediately after each device was presented (3 pm), with this peak decreasing within the next two hours. Occasional device movements were recorded overnight. Movements of the foraging devices resumed in the morning after the lights came on. Frequently, animals continued to interact with the devices on the following morning even though no food was observed on the devices at this time. In general, the same patterns of movement occurred for both devices. Paired t-tests of total daily activity across the two-week period showed that five of six baboons moved the P-Nut Butter Rolls more than the grooming boards (p < .01). This finding may be due to increased difficulty in retrieving food from the P-Nut Butter Roll, which would create more movement, or it may simply demonstrate a food preference. Analyses of the videotapes revealed that the most common ways in which the baboons interacted with a grooming board were for them to either brace the board with one hand and groom the fleece with the other, or press the board to the cage with both hands and lick the fleece. For the P-Nut Butter Roll, baboons typically braced the cylinder to the cage in some manner (e.g., pressing the cylinder to the cage), and then extracted peanut butter by either digging at the slot with one finger, or shoving all the fingers of one hand into the slot and then licking the fingers.
Figure 1: Average 24-hour patterns of movement for grooming boards (left column) and P-Nut Butter Rolls (right column) recorded from each baboon. Plotted are the number of minutes of movement of each device per 15-min interval over a 24-hr period, averaged across the two-week device exposure period (excluding weekend data). Lights-off times are indicated by the shaded areas.
Figure 2 shows the total minutes of device movements per day recorded from the grooming board (left) and the P-Nut Butter Roll (right) for each animal, plotted across consecutive weekdays. Variations occurred in how much the devices were moved across the two-week exposure periods. In some instances, the devices were moved most frequently during the first 1-3 days of exposure to the devices (i.e., grooming boards for baboons AC, CA, and WE; P-Nut Butter Rolls for baboons AC, BE, and DR), suggesting a "novelty" effect following initial exposure to the devices. Some baboons showed a gradual decrease, or habituation, in their interactions with the devices, as indicated by decreasing device movements over days (i.e., grooming boards for baboons AC, CA, and WE; P-Nut Butter Rolls for baboons AC, DR, and FR). In still other instances, some baboons moved the devices at about the same rate over the two-week period (grooming boards for baboons BE, DR, and FR; P-Nut Butter Rolls for baboons BE, CA, and WE). Finally, in two instances baboons interacted very little with the grooming board (baboons DR and FR). These differing movement patterns over time indicate that a range of individual differences exists among nonhuman primates in their interaction with foraging devices.
Figure 2: Total minutes of device movements per day for grooming boards (left) and P-Nut Butter Rolls (right) for each baboon, plotted across consecutive weekdays.
Table 3 shows the percentage of time that each baboon engaged in the six behavioral categories of Table 2 prior to being exposed to either of the two foraging devices. In general, most baboons engaged in general movements and/or rested. Baboons AC and WE, however, showed a relatively high percentage of stereotypic behaviors during this baseline period. Baboon AC repeatedly circled, braced, and sat on his bench, while baboon WE circled his cage repeatedly.
Stereo- Self- Cage- Bracing General Resting
Animal typic Directed Directed Movement
DR 0.0 19.0 5.0 8.0 51.0 17.0
AC 25.0 0.0 0.0 10.0 39.0 26.0
WE 44.0 1.0 0.0 0.0 36.0 19.0
FR 8.0 4.0 0.0 21.0 27.0 40.0
BE 3.0 18.0 5.0 4.0 57.0 13.0
CA 0.0 10.0 3.0 0.0 38.0 49.0
Table 3: Percentage of time occupied by behaviors during the baseline
video segment.
To determine whether or not foraging device use was correlated with these six behavioral categories, the percentage of time that baboons engaged in device use was plotted as a function of the percentage of time they engaged in each of these six behavioral categories prior to exposure to either foraging device. Of the six categories, stereotypy and self-directed behaviors were highly correlated with subsequent foraging device use. Figure 3 shows plots for P-Nut Butter Roll use (left) and grooming board use (right) as a function of the percentage of time that baboons had previously engaged in stereotypical behaviors. Baboons with greater amounts of prior stereotypical behavior also interacted with the foraging devices more. The correlation between stereotypic behavior and P-Nut Butter Roll use was extremely high (r = .951, p <. 005), whereas the correlation between stereotypic behavior and grooming board use was lower (r = .626, 0.2 > p > 0.1). Further, no animal exhibited stereotypic behaviors during the videotaping times when foraging devices were attached to the cages. Figure 4 shows P-Nut Butter Roll use (left) and grooming board use (right) as a function of the percentage of time that baboons engaged in self-directed behaviors. Baboons displaying lower amounts of self-directed behaviors interacted with the foraging devices less frequently. The correlation between self-directed behaviors and P-Nut Butter Roll use approached significance (r = .793, 0.1 > p > 0.05); the correlation between self-directed behaviors and grooming board use was lower (r = .616, 0.2 > p > 0.1). The remaining behavioral categories showed no significant correlations with foraging device use.
Figure 3: The percentage of time baboons engaged in using the P-Nut Butter Roll (left) and grooming board (right), plotted as a function of the percentage of time baboons engaged in stereotypical behaviors prior to introduction of the foraging devices. A linear correlation function and the corresponding Pearson's r value are shown.
Figure 4: The percentage of time that baboons were engaged in using the P-Nut Butter Roll (left) and grooming board (right), plotted as a function of the percentage of time baboons engaged in self-directed behaviors prior to introduction of the foraging devices. A linear correlation function and the corresponding Pearson's r value are shown.
Average food pellet intakes were also examined during baseline and testing periods to determine whether exposure to the foraging devices influenced the animals' performances in the signal detection task. No differences in overall pellet intake were found between the foraging device and non-device periods (p = .64), and daily plots of each animal's signal detection thresholds showed no systematic deviations from baseline performance levels.
Discussion
Our results show that singly caged adult male baboons readily interact with food-based foraging devices, and that stereotypical behaviors are reduced when the devices are present. In addition, the foraging devices elicited species-typical behaviors such as foraging and grooming, even after food appeared to be absent from the devices. These results indicate that food-based enrichment devices can have substantial benefits for singly caged baboons (e.g., reducing stereotypic behaviors). Similar results of reductions in abnormal behaviors following the introduction of feeding devices have been reported for rhesus monkeys (Bayne et al., 1991), chimpanzees (Maki et al., 1989), and cynomologus monkeys (Lam et al., 1991).
A finding of great potential significance is the correlation between foraging device use and the frequency of prior stereotypic and self-directed behaviors. In the present study, foraging devices were used more frequently by those baboons exhibiting relatively higher frequencies of stereotypic behaviors; conversely, the devices were used less frequently by baboons exhibiting relatively higher frequencies of self-directed behaviors (e.g., self-groom-ing). These correlations suggest the importance of noting the nature of abnormal behaviors in nonhuman primates before prescribing enrichment strategies for treating such problems. For example, feeding devices might be particularly helpful for animals exhibiting stereotypic "pacing" behaviors, but less so for animals exhibiting self-directed behaviors such as hair-pulling. Clearly, additional research using other nonhuman primate species is needed to determine the generality of this finding.
Previous investigators have reported that nonhuman primates show no habituation to food-related devices (e.g., Boccia, 1989). In the present study, however, the use of activity monitors allowed for a more accurate assessment of the pattern of device use over time, with the resulting data showing that some habituation occurred in two baboons (see Figure 2). Such decreases in device use could be due to animals developing more efficient methods in removing food from the devices. On the other hand, baboons also interacted with the devices in the morning after the food had been removed, suggesting that the devices may have remained beneficial even when food was not available. This "residual" interaction with an empty foraging device would contribute additional movements to the total daily device movement times of Figure 2, which may be why more habituation to the foraging devices was not observed when total movement over 24-hrs was measured.
Both the P-Nut Butter Roll and grooming board proved compatible with ongoing research protocols in that the devices were easily incorporated into technicians' daily routines. Further, although the food retrieved from the devices slightly decreased daily pellet intakes, it did not appear to adversely affect the accuracy of the baboons' performances in their behavioral tasks. Evaluating other devices in this way will assist research facilities not only in providing environmental enrichment to meet USDA regulations, but also in providing enrichment that will be most beneficial to their nonhuman primates. Although an individualized approach to enrichment may not be convenient in some research settings, this approach may alleviate some aberrant behaviors and thus increase psychological well-being in singly caged nonhuman primates.
References
Bayne, K., Mainzer, H., Dexter, S., Campbell, G., Yamada, F., & Suomi, S. (1991). The reduction of abnormal behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology, 23, 23-35.
Bloom, K. R., & Cook, M. (1989). Environmental enrichment: Behavioral responses of rhesus to puzzle feeders. Lab Animal, 18, 25-31.
Bloomsmith, M. A., Brent, L. Y., & Schapiro, S. J. (1991). Guidelines for developing and managing an environmental enrichment program for nonhuman primates. Laboratory Animal Science, 41, 372-377.
Boccia, M. L. (1989). Long-term effects of a natural foraging task on aggression and stereotypies in socially housed pigtail macaques. Laboratory Primate Newsletter, 28[4], 18-19.
Fajzi, K., Reinhart, V., & Smith, M. D. (1989). A review of environmental enrichment strategies for singly caged nonhuman primates. Lab Animal 18[2], 23-35.
Hienz, R. D., Turkkan, J. S., Spear, D. J., Sannerud, C. A., Kaminsky, B. J., & Allen, R. P. (1992). General activity in baboons measured with a computerized, lightweight piezoelectric motion sensor: Effects of drugs. Pharmacology, Biochemistry, and Behavior, 42, 497-507.
Lam, K., Rupniak, N. M. J., & Iversen, S. D. (1991). Use of a grooming and foraging substrate to reduce cage stereotypies in macaques. Journal of Medical Primatology, 20, 104-109.
Maki, S., Alford, P. L., Bloomsmith, M. A., & Franklin, J. (1989). Food puzzle device simulating termite fishing for captive chimpanzees (Pan troglodytes). American Journal of Primatology, 1, 71-78.
Novak, M. & Suomi, S. J. (1991). Social interaction in non-human primates An underlying theme for primate research. Laboratory Animal Science 41, 308-314.
O'Neill, P., Price, C., & Suomi, S. (1989). Daily patterns in activity levels relative to age and sex in a free-ranging group of rhesus monkeys. American Association of Laboratory Animal Scientists Meeting, Little Rock, AR.
* * *
Welfare Information Requirements
The Animal Welfare Information Center (AWIC) will hold a series of 2-day workshops, April 4-5, August 1-2, and November 14-15, 1996 at the National Agricultural Library, 10301 Baltimore Blvd, Beltsville, MD 20705-2351. Each workshop will be limited to 20 persons. The subject will be meeting the informational requirements of the Animal Welfare Act, focusing on documenting, for IACUCs, that a thorough literature search was conducted regarding alternatives. These workshops are targeted for principal investigators, members of IACUCs, information providers, administrators of animal use programs, and veterinarians. There is no charge for these workshops. Contact AWIC at the above address or call 301-504-6212 [FAX: 301-504-7125; e-mail: [email protected]].
Captive Chimpanzees
The 1996 workshop on the Care and Management of Captive Chimpanzees will be held during the joint meeting of IPS and ASP in Madison, WI this August 11-16. T. Matsuzawa (PRI, Kyoto University) and J. A. R. A. M. vanHoof (Universiteit Utrecht) will speak on the application of information from the field to the captive environment. For information on the workshop, contact Linda Brent, Southwest Foundation for Biomedical Research, P.O. Box 28147, San Antonio, TX 78228 [210-675-1410; e-mail: [email protected]].
Animal Care and Use Programs
The NIH Office of Extramural Research (OER), Office for Protection from Research Risks (OPRR) is continuing to sponsor workshops on implementing the Public Health Service Policy on Humane Care and Use of Laboratory Animals. The workshops are open to institutional administrators, members of Institutional Animal Care and Use Committees, laboratory animal veterinarians, investigators, and other institutional staff who have responsibility for high-quality management of sound institutional animal care and use programs. Ample opportunities will be provided to exchange ideas and interests through question-and-answer sessions and information discussions.
A workshop titled "The Role of the Institutional Official in Compliance with the Animal Welfare Act and the PHS Policy on Humane Care and Use of Laboratory Animals" will be held May 16-17, 1996 at Wright State University, Dayton, OH, cosponsored by Central State University, Ohio University, and Wright State. There is a $150 registration fee.
For registration, contact Ms. Theresa Atwood, Office of Conferences and Events, Wright State Univ., 3640 Colonel Glenn Hwy, Dayton, OH 45435 [513-873-5512; fax: 513-873-5517].
Topics of particular importance to Institutional Officials to be addressed in this workshop include: the role of members of the IACUC the role of the Institutional Official in non-compliance methods for evaluating the effectiveness of an institution's program for animal care and use determining how effectively the program meets its mandates.
For information concerning future NIH/OER/OPRR Animal Welfare Education Workshops, contact Ms Darlene M. Ross, OPRR, NIH, 6100 Executive Blvd, Suite 3B01, MSC 7507, Rockville, MD 20892-7507 [301-496-8101, Ext. 233; fax: 301-402-0527].
Third Gorilla Workshop
The Third Gorilla Workshop, sponsored by the Pittsburgh Zoo, will be held 2-6 April, 1997 at Sheraton Station Square, Pittsburgh, PA. The focus will be on conservation, behavior, husbandry, reproduction, and field work. Contact Debbie McGuire or Roseann Gianbro, Pittsburgh Zoo, One Hill Rd, Pittsburgh, PA 15206-1178 [1-412-665-3794; Fax: 1-412-665-3661].
* * *
An exhibit titled "Primates in Art and Illustration" will be held in conjunction with the IPS/ASP Joint Meeting in Madison, WI, August 11-16, hosted by the Wisconsin RPRC. The committee for the exhibit welcomes suggestions for artists to contact as well as submissions from primatologists.
Exhibit dates will be August 9-30. In addition to primatologists attending the meeting, we anticipate considerable interest in this exhibit from people in Madison and the surrounding area.
Painters, illustrators, sculptors and photographers are invited to submit samples of their work related to nonhuman primates. Works which are anthropomorphic in nature are discouraged. We anticipate being able to mount approximately 50-55 works in two galleries, with each artist being represented by not more than 3-5 items. Works will be juried by a panel of primatologists and local area artists. Unless specially arranged, all works will be for sale with the gallery taking a 20% commission, plus an additional 10% designated for the IPS/ASP Conservation Fund. Not only is the Conservation Fund important, but people will be more likely to purchase items if they know that part of the purchase goes toward preservation of endangered primates.
Only works related to nonhuman primates will be considered. Please send 3-5 slides or photographs of your work to: IPS/ASP Art Exhibit, c/o Larry Jacobsen, Wisconsin RPRC, Univ. of Wisconsin, 1220 Capitol Ct, Madison, WI 53715-1299 [for further information, phone 1-608-263-3512; fax 1-608-263-4031; or e-mail: [email protected]].
An exhibit (no sales) of children's art focused on primates will also be mounted in the Theater Galleries. The IPS/ASP Education Committees are putting this together. Contact Sean Evans (14805 S.W. 216 St, Miami, FL 33170) or Gabriele Lubach (Harlow Primate Lab., 22 N. Charter St, Univ. of Wisconsin, Madison, WI 53715).
* * *
Telemetry for Multiple Subjects
Prof. Dr. H. Rothe reports that a data-collecting system for simultaneous collecting of heart rates and behavioral data of up to eight small laboratory mammals has been developed by Dipl. Biol. Jens Kerl of the University of Göttingen. The data-logging program stores heart rate data together with the corresponding behavioral data for each animal. During a preliminary study on a pair of common marmosets (Callithrix jacchus), the system proved to be a powerful tool.
Further information is available from Jens Kerl, Inst. of Anthropology, Univ. of Göttingen, Ethologische Station Sennickerode, 37130 Gleichen, Germany [49-5592-1513; fax: 49-5592-1524; e-mail: [email protected]].
Gonadal and Semen Samples
Susan Bufton is undertaking a comparative study of gross morphological differences in spermatozoa, using scanning and transmission electron microscopy as a qualitative method. She would like to obtain samples from various species. Donated tissues can be in the form of fresh, frozen or preserved gonadal specimens or fresh, frozen or preserved semen samples. Full acknowledgement will be given. Contact Susan M. Bufton, c/o Tidewater Technical Laboratories, Ltd., Kempsville Office Park, 408 Oakmears Crescent, Suite 203, Virginia Beach, VA 23462 [804-422-0096; e-mail: [email protected]].
Primate Vocalizations
The Wisconsin RPRC Library makes a concerted effort to collect, catalog, and make accessible a wide range of recorded primate vocalizations. Without an effort to conserve this material, which may ultimately be the only audiovisual documentation for some endangered primates, these important vocalizations may be lost. "Whether you have one call or a full repertoire available, we would like to hear about your work. If you have audiotaped vocalizations you could contribute, we would like the best quality copies available. If you cannot part with the original recording, we accept copies which we may retain. We also have the facility to make copies. This process usually takes 2 weeks, after which the original is returned to the contributor. If you are planning any future research which may involve taping vocalizations, please keep us in mind. Any help in pointing us toward potential sources of material would also be appreciated." For further information, contact Ray Hamel, Special Collections Librarian, Wisconsin RPRC, 1223 Capitol Ct, Madison, WI 53715.
* * *
Journal of Medical Primatology
Munksgard Press has announced that "Due to problems between the New York University Medical Center and LEMSIP (Laboratory for Experimental Medicine and Surgery in Primates) the publication of Journal of Medical Primatology has been delayed and, subsequently, Dr. Jan Moor-Jankowski has asked us as publishers to be released as Editor-in-Chief."
Professor Jorg W. Eichberg of the Biomedical Primate Research Centre in Rijswijk, the Netherlands, will be the new Editor-in-Chief. The journal will be published bimonthly, i.e. a total of six issues including the special issue from the Annual Primate Centre Meeting. "Professor Eichberg is currently assembling a new Editorial Board and will announce the Board, including a new aim and scope, in the first 1996 issue of Journal of Medical Primatology." As a consequence of the delay, only four of the eight 1995 issues will be published.
African Primates
African Primates is in need of articles for issue no. 3 which is due to come out in mid-1996. Articles in French are particularly desired. African Primates publishes information relevant to the conservation of nonhuman primates and their ecosystems in Africa. Please refer to issue no. 1 for answers to your questions or contact Tom Butynski, Zoo Atlanta, P.O. Box 24434, Nairobi, Kenya [fax: 254-2-890615; e-mail: [email protected]]. It is hoped that this newsletter will enhance the conservation of African primates: by increasing interest in their survival, by alerting people to situations where primate species and populations are under threat, and by providing a forum for useful debate on some of the more pressing, controversial, and sensitive issues that have an impact on the conservation of these primates.
The success of this newsletter depends largely upon the willingness of those people involved with primate conservation in Africa to provide relevant information on research findings, field survey results, advances in field and laboratory techniques, field action alerts, book reviews, events, job announcements, funding possibilities and recent publications. African Primates also announces letter-writing campaigns and other activities which might benefit from the support of its readership.
African Primates is published bi-annually and distributed free of charge to all interested persons. About 3,000 copies are produced of each issue. The mailing list now stands at about 1,200 addresses. Please send (or e-mail) contributions to: Tom Butynski, Editor, African Primates, Zoo Atlanta, Africa Biodiversity Conservation Program, P.O. Box 24434, Nairobi, Kenya [254-2-745374; fax: 254-2-890615; e-mail: [email protected] or [email protected]].
ACLAD Newsletter
Stephen S. Morse, Secretary of the American Committee on Laboratory Animal Diseases (ACLAD) and Editor of the ACLAD Newsletter, invites the submission of 1- to 2-page summaries of research on lab animal diseases, diagnostic methods, or animal models, or relevant comments on any of these subjects. His address is Rockefeller University, 1230 York Ave, P.O. Box 120, New York, NY 10021-6399 [212-327-7722; fax: (212) 327-7172; e-mail: [email protected]]. The Newsletter is made available on the CompMed e-mail list, and by paper mail to ACLAD members.
JAAWS: New Animal Welfare Journal
The Journal of Applied Animal Welfare Science (JAAWS) is issuing a call for papers. Conceived to promote the emerging field of animal welfare science, the goal of the journal is to publish articles and reports on methods of experimentation, husbandry, and care that demonstrably enhance the welfare of animals. The first volume of JAAWS will be published in January, 1997 and thereafter on a quarterly basis. A coproject of the American Society for the Prevention of Cruelty to Animals and Psychologists for the Ethical Treatment of Animals, JAAWS will be published by Lawrence E. Erlbaum. Several types of manuscripts are accepted: "Articles" present new empirical data or a reevaluation of available data, or conceptual or theoretical analysis, and are up to 8000 words. These feature articles are accompanied by several invited critical commentaries on them, of up to 2500 words each. "Reports" present scientific experiments or demonstrations relating to some issue of animal welfare science and are about 5000 words.
The deadline for manuscripts for the inaugural issue is May 1, 1996. Send manuscripts and requests for subscription information to either: Kenneth J. Shapiro, PSYeta, P.O. Box 1297, Washington Grove, MD 20880 [phone and fax: 301-963-4751; e-mail: [email protected]] or Stephen Zawistowski, ASPCA, 424 East 92nd St., New York, NY 10128 (send books for review and other correspondence to K. J. S).
* * *
ASP Conservation Awards and Grants
Nominations for Conservation Awards and Grants are now being sought by the American Society of Primatologists (ASP). These awards and grants, funded from the ASP Conservation Fund, are a contribution to the larger conservation picture and a mechanism to recognize deserving colleagues and students, including those from primate habitat countries -- countries with native primate fauna -- for whom the prestige of an ASP award or grant can be a valuable aid to the recipient's conservation efforts. An award nomination is basically a letter of recommendation. A grant proposal should consist of a few narrative pages plus a budget page.
The Subscription Award provides the American Journal of Primatology to worthy individuals in habitat countries who otherwise would have little access to the scientific literature on nonhuman primates. The Society expects to support several such continuing awards. Preference is given to those who will make the journal available in a central place for colleagues to use, and to those who can justify brief reapplication every two years to build up their library of the journal. A nominating letter should describe the nominee's credentials and his/her primate-related activities, and explain why the nominee deserves to receive high priority.
The Conservation Award ($500) provides recognition and financial support for students and young investigators from habitat countries who demonstrate potential for making significant and continuing contributions to primate conservation. Those eligible include students, young investigators, and educators in countries with native nonhuman primates and for whom no more than five years have elapsed since receipt of the terminal degree. Nominators should provide the nominee's name, title and full mailing address and a statement about the nominee's qualifications for the award, focusing on past and potential contributions to primate conservation. Supporting letters from other individuals acquainted with the work of the nominee may be submitted. Past awards have been presented by U.S. Ambassadors or other senior officials, thereby obtaining favorable publicity for the award, its recipient, and primate conservation in the recipient's country.
Senior Biology and Conservation Award ($500 Honorarium) is one of ASP's highest honors. It is given to recognize an individual without an advanced degree who has made substantial contributions over many years to promote primate conservation either through direct action or via enhancement of biological knowledge or well-being of primates. Such contributions could arise from work done in field, laboratory or zoo settings. Nominees might work directly with primates or be engaged in activities supporting those who work with primates. Examples include park rangers, census takers, animal caretakers, research technicians, assistants or facilitators, and individuals involved in private enterprise benefitting primate conservation. Nominating letters should detail the nominee's qualifications, contributions to primate conservation, period of service, and full mailing address. Supporting letters from other individuals acquainted with the work of the nominee may be submitted. This award typically is presented at public ceremonies by senior officials.
Conservation Small Grants (up to $1,500 but usually $500) are given for conservation research or other projects, including conservation education. ASP and IPS members working in habitat countries are especially urged to apply or to help someone from a habitat country submit a meaningful project, which can be a portion of a larger effort. Grant proposals must be typed in English, should not exceed 2000 words, and should include a brief budget page. Recipients of grants must agree that a brief progress report, in a form suitable for publication in the ASP Bulletin, will be made within 12 months of the award.
Evaluation and Application Procedure: With the exception of requests for emergency support, which can be considered at any time for immediate action, the Conservation Committee will make its recommendations for awards and grants to the ASP Executive Committee at its annual meeting. Successful nominees and applicants will be informed following the meeting and their names published in the ASP Bulletin. The 1996 deadline for submission of nominations and grant proposals is June 30. They should be sent to Ramon J. Rhine, Psychology Dept, University of California, Riverside, CA 92521.
W. W. Howells Prize
The W. W. Howells Prize is awarded by the Biological Anthropology Section of the American Anthropological Association to honor a book in the area of biological anthropology. The book should inform a wider audience of the significance of physical/biological anthropology in the social and biological sciences and demonstrate a biocultural perspective. Nominated works should represent the highest standards of scholarship and readability. The book may have multiple authors, but may not be an edited volume. Books must be published within the last three to four years and once nominated will remain on the list for three to five years, depending on their date of publication.
Previous winners of the Prize are: 1993: Dorothy Cheney and Robert Seyfarth (1990) How Monkeys See the World: Inside the Mind of Another Species. University of Chicago Press. 1994: Matt Cartmill (1993) A View to a Death in the Morning: Hunting and Nature through History. Harvard University Press. 1996 (awarded in 1995): William C. McGrew (1992) Chimpanzee Material Culture. Cambridge University Press.
Nominations, in the form of a letter and accompanying material, such as book reviews, should be sent to: Karen Rosenberg, Department of Anthropology, University of Delaware, Newark, DE 19716 [302-831-1855; fax: 302-831-4002; e-mail: krr@udel.edu].
* * *
Leo A. Whitehair, who has directed the National Center for Research Resources Comparative Medicine Program for about seven years, received the 1995 Griffin Award, given by the American Association of Laboratory Animal Science, in recognition of his ongoing efforts to improve the care and quality of animals used in biological and medical research. Dr. Whitehair oversees NCRR's Regional Primate Research Centers Program, the Laboratory Animal Sciences Program, the AIDS Animal Models Program, and training programs in animal medicine. The award was given at AALAS's national meeting in October.
* * *
* University of California, Davis, Psychology Department
PROGRAM NAME: Comparative Psychology and Physiological Psychology are
specializations within the Psychobiology program.
FACULTY & THEIR SPECIALTIES: Leo M. Chalupa (central mechanisms of vision,
prenatal development of sensory systems in the mammalian brain); Richard G.
Coss (developmental psychobiology, evolution, experimental aesthetics,
antipredator behavior); Michael S. Gazzaniga (cognitive neuroscience); Kenneth
R. Henry (audition, physiological psychology, behavioral genetics,
developmental psychobiology, aging); Leah A. Drubitzer (evolutionary
neurobiology); George R. Mangun (human cognitive neurophysiology); Peter R.
Marler (animal communciation, neuroethology); William A. Mason (primate
social behavior); Sally P. Mendoza (behavioral endocrinology, physiological
basis of primate social relationships, stress and reproduction); Robert M.
Murphey (behavior of domesticated ungulates, genetic correlates of behavior,
psychopathology); Donald H. Owings (communication and antipredator behavior,
ground squirrel behavior); Robert Sommer (environmental psychology, abnormal
psychology, action research); Niels G. Waller (behavior genetics,
psychometrics, and personality).
FOR FURTHER INFORMATION: Graduate Admissions, Department of Psychology,
University of California, Davis, CA 95616.
* Emory University, Yerkes Regional Primate Research Center
PROGRAM DESCRIPTION: Behavior and Biology of Primates Training Program:
Postdoctoral training is available in several sciences that contribute to our
understanding of the behavior and biology of primates. These include: primate
behavior, including learning, memory, cognition, communication, social behavior
and psychopharmacology; molecular medicine; neurosciences; vision; and primate
models of human diseases. Training facilities: Training facilities of the
Yerkes Center including its Field Station, as well as a wide variety of other
laboratories at the Main Station, are available. Funding for Research
Associates and Research Fellows generally is derived from individual research
grants at the center or fellowships awarded by public and private agencies.
FOR FURTHER INFORMATION: Tom Gordon, Associate Director for Scientific
Programs, Yerkes Regional Primate Research Center, Emory University, Atlanta,
GA 30322.
* * *
Conservation Biology Awards
The Sophie Danforth Conservation Biology Fund was established by the Roger Williams Park Zoo and the Rhode Island Zoological Society to help protect the world's threatened wildlife. Each year we award grants of up to $1,000 to individuals or institutions working in conservation biology. Projects and programs that enhance biodiversity and maintain ecosystems receive the highest funding priority. Field studies, environmental education programs, development of techniques that can be used in a natural environment, and captive propagation programs that stress an integrative and/or multi-disciplinary approach to conservation are also appropriate. Proposals for single species preservation, initial surveys, or seed money for technique development are not appropriate.
Recipients are required to acknowledge the Roger Williams Park Zoo and the Rhode Island Zoological Society in any publications that result from the project. Recipients must also submit a progress report that includes an update on the status of the project. This report is due one year after funding.
All proposals must be submitted by May 1, 1996. Applications will be reviewed by a committee of Zoo, RI Zoological Society, and outside advisors. Grants will be awarded in July, 1996. For applications and further information contact Dr. Anne Savage, Director of Research, Roger Williams Park Zoo, Elmwood Ave, Providence, RI 02905 [401-785-3510; FAX: 401-941-3988; e-mail: [email protected]].
Models for HIV and AIDS
The National Cancer Institute and the National Institute of Allergy and Infectious Diseases are seeking to encourage investigator-initiated grant applications for the development of useful and predictive biochemical, cellular, in vivo and mathematical models for the preclinical evaluation of new therapies against HIV disease and AIDS-related malignancies. The availability of well-characterized in vitro and in vivo models would accelerate the pace of evaluation of different paradigms of disease progression and would facilitate the discovery of successful treatments, including drugs, vaccines, gene therapy, and immune modulators. Collaborative arrangements involving more than one institution are encouraged, including participation of the pharmaceutical industry where appropriate. Although the goal of this announcement is to stimulate the development of diverse types of efficient and predictive biochemical, cellular, in vivo, and mathematical models for the evaluation of new agents for the treatment of HIV disease and AIDS-related malignancies, priority will be given to in vivo models if the number of meritorious applications exceeds funds available.
For information, contact Dr. Nava Sarver, Div. of AIDS, NIAID, Solar Bldg, Rm 2C01, 6003 Executive Blvd, MSC 7620, Bethesda, MD 20892-7620 [301-496-8197; fax: 301-402-3211; e-mail: [email protected]] or Dr. Mary K. Wolpert, DCTDC, NCI, Executive Plaza North, Rm 832, 6130 Executive Blvd, MSC 7450, Bethesda, MD 20892-7450 [301-496-8783; fax: 301-496-8333; e-mail: [email protected]].
Academic Research Enhancement Award
The National Institutes of Health (NIH) is continuing to make a special effort to stimulate research in educational institutions that provide baccalaureate training for a significant number of the nation's research scientists, but historically have not been major recipients of NIH support. NIH is inviting grant applications at this time for Academic Research Enhancement Award (AREA) grants to be awarded competitively in FY 1997. The AREA funds are intended to support new research projects or expand ongoing research activities proposed by faculty members of eligible institutions in areas related to the health sciences.
The application receipt date is June 26, 1996. Questions regarding eligibility, policies, procedures, and other administrative aspects of the NIH AREA program should be referred FIRST to the Office of Sponsored Programs at the educational institution. Issues that remain AFTER consultation with the institutional Office of Sponsored Programs may be directed to: Special Programs, Office of Extramural Research, NIH, 6701 Rockledge Dr., Rm 6186 - MSC 7910, Bethesda, MD 20892-7910 [301-435-2770; fax: 301-480-0146; e-mail: [email protected]].
Small Grants on Aging
The National Institute on Aging (NIA) is seeking small grant (R03) applications to stimulate and facilitate the entry of promising new investigators into targeted, high priority areas of aging research. This Small Grant Program provides support for pilot research that is likely to lead to a subsequent individual research project grant (R01) or a First Independent Research Support and Transition (FIRST) (R29) award application and/or a significant advancement of aging research.
To be eligible for this award, the proposed Principal Investigator must, at a minimum, be an independent investigator at the beginning of her/his research career as defined by the eligibility requirements for a FIRST (R29) award. Established investigators proposing research unrelated to a currently funded research program are also eligible to apply for these grants.
The Small Grant program is designed to support independent basic and clinical scientists who are interested in entering the area of aging research. Targeted topics include the following: Aging muscle: basic underlying mechanisms; Molecular basis of cardiovascular aging; Aging immunobiology: basic underlying mechanisms; Animal models of aging Neural mechanisms of age-related changes in attention; Aging protein: Protein structure changes with aging with special emphasis on the nervous system; Mechanisms underlying changes in spatial and temporal processing in sensory systems of the aging brain; Mechanisms underlying changes in sleep processes in older organisms.
Application receipt dates are July 12 and November 12, 1996. Only one Small Grant application may be submitted by a principal investigator per receipt date. Direct inquiries to: Biology of Aging Program, David B. Finkelstein [301-496-6402; fax: 301-402-0010; e-mail: [email protected]]; Behavioral & Social Research, Jared Jobe [301-496-3137; fax: 301-402-0051; e-mail: [email protected]]; Neuroscience & Neuropsychology of Aging Program, Judy Finkelstein [301-496-9350; fax: 301-496-1494; e-mail: [email protected]]. The address for all is: NIA, Gateway Bldg, Suite 2C212, 7201 Wisconsin Ave, MSC 9205, Bethesda, MD 20892.
Internet Connection for Medical Institutions
The National Library of Medicine (NLM) is encouraging the development of a communications infrastructure to promote the rapid interchange of medical information nationally and throughout the world. This infrastructure is based upon the Internet, a network of networks that was developed with the support of the National Science Foundation. Internet access provides health professionals engaged in education, research, clinical care, and administration with a means of accessing remote databases, libraries, NLM's Grateful Med, DOCLINE, and Loansome Doc; of transferring files and images; and of interacting with colleagues throughout the world. To accelerate the pace with which health-related institutions become part of the electronic information web, NLM is offering grants to support Internet connections.
Support is available for development and enhancement of multi-institution networks including extending connectivity to outlying sites, or otherwise furthering NLM's goal of expanding information outreach. Institutions will usually be expected to fund the local area network and support other costs of the gateway system. Grant funds may be used to expand access to the Internet in an institution with an existing connection.
Prospective applicants are asked to submit a letter of intent by April 17, 1996. The application receipt date is May 16, 1996. Direct inquiries to Ms. Frances E. Johnson, Div. of Extramural Programs, NLM, Bldg 38A, Rm 5S-520, Bethesda, MD 20894 [301-496-4221; fax: 301-402-0421; e-mail: [email protected]].
L. S. B. Leakey Foundation Grants
The L. S. B. Leakey Foundation was formed to further research into human origins, behavior, and survival. Recent priorities have included research into the environments, archaeology, and human paleontology of the Miocene, Pliocene, and Pleistocene; into the behavior, morphology, and ecology of the great apes and other primate species; and into the behavioral ecology of contemporary hunter-gatherers. Other areas of study related to human evolution have been funded occasionally.
Advanced pre-doctoral students, as well as established scientists, are eligible for General Research Grants, normally given to the exploratory phases of promising new projects that most closely meet the stated purpose of the Foundation. A special Fellowship for Great Ape Research promotes long-term research on the behavior and ecology of wild populations of great apes, especially if, in addition to the basic scientific goals of the project, the work contributes to the development or testing of models of human evolution. Both continuing and new projects wlll be considered. Strong preference is given to post-doctoral applicants prepared to make a long-term commitment to the study site. The Paleoanthropology Award is intended for potentially long-term, multidisciplinary research programs which seek to recover the physical and/or cultural remains of early humans and their hominid ancestors. Franklin Mosher Baldwin Memorial Fellowships are awarded to Africans who seek to complete an advanced degree in anthropology at a major institution and is limited to a two-year program of advanced training towards an M.A., Ph.D., or equivalent. Priority is given to students of disciplines related to human evolution.
For information on application procedures, deadlines, etc., contact the L. S. B. Leakey Foundation, 77 Jack London Square, Suite M, Oakland, CA 94607-3750 [510-834 -3636; fax: 510-834-3640].
* * *
Hepatic Capillariasis
Carles Juan Salles and his co-workers have recently diagnosed acute toxoplasmosis and chronic hepatic capillariasis in a white-faced saki (Pithecia pithecia). Since hepatic capillariasis seems to be a disease rarely reported in primates, they are making an international survey to determine the incidence and course of this disease among captive primates. If you have any data, please contact: Carles Juan Salles, Dept. de Patología y Producciones Animales, Facultat de Veterinaria, Univ. Autónoma de Barcelona, 08193 Bellaterra, Barcelona, Spain [Fax: 3-581-3142; e-mail: [email protected]].
International Directory of Primatology -- 1996
The third edition of the International Directory of Primatology is currently in preparation. This is a major directory to the field, including more than 300 organizations active in primate research, conservation, and education. Information can be found about species held, research programs, training and educational opportunities, primates in zoological gardens, primate societies, and informational programs. Contact information for 2200 people, including the current membership of the International Primatological Society, is provided. Prepared by staff and affiliates of the Wisconsin RPRC, the third edition will also include a listing of field studies and a roster of primate-related World Wide Web sites. Over 1,000 copies of the second edition were distributed to individuals, primate organizations and libraries worldwide.
Questionnaires for the third edition have been sent out to over 400 primate programs worldwide. The deadline for return of questionnaires is April 30, although newly identified programs will receive extended time. Organizations or programs with a focus on primatology are welcome to contact the coordinator about being added to the directory. Commercial organizations willing to help support production and distribution costs will be given detailed listings in the Contributor Section.
Completion of the directory is scheduled for July/August, 1996 and copies will be available at the IPS/ASP Meetings in Madison, WI August 11-16. The price is not yet set. For further information contact Larry Jacobsen, IDP Coordinator, Wisconsin RPRC, 1220 Capitol Court, Madison, WI 53715-1299 [608-263-3512; e-mail: [email protected]].
More WWW Addresses
All of the following URLs (WWW addresses) begin with "http://" which has been omitted to save space.
* U.S. Dept Health & Human Services: www.os.dhhs.gov
* FDA: www.fda.gov/cdrh/cdrhhome.html
* Center for Veterinary Medicine: www.cvm.fda.gov
* National Institutes of Health: www.nih.gov
* Program for Monitoring Emerging Diseases:
www.healthnet.org/promed.html
* World Health Organization: www.who.ch
* ACLAM:
chopin.osp.uh.edu/~rocky/aclam/hdg1055.htm
* The Scientist -- on the Penn Library Web
www.the-scientist.library.upenn.edu/
* South Texas Primate Observatory:
www.nodehaven.com/stpo
* Southwest Foundation for Biomedical Research: www.sfbr.org
* National Association for Biomedical Research:
www.fiesta.com.nabr
* Foundation for Biomedical Research: www.fiesta.com.fbr
* Lincoln Park Zoo: www.lpzoo.com
* Primate Handedness and Brain Lateralization Research:
www.indiana.edu/~primate/index.html
* Orangutan Foundation International:
www.ns.net/orangutan
* African Primates at Home:
www.indiana.edu/~primate/primates.html
* East African Research Sites:
www.indiana.edu/~primate/sites.html
* Primate Society of Great Britain (PSGB):
www.ana.ed.ac.uk/PSGB/
* Malaysia Dept of Wildlife and National Parks:
www.jaring.my/hilitan/
* National University of Hanoi Zoological Museum:
coombs.anu.edu.au/~vern/hnu-zoo-mus.html
* Vietnam Inst. Ecology & Biological Resources:
coombs.anu.edu.au/~vern/iebr.html
* Parks and reserves in Vietnam:
coombs.anu.edu.au/~vern/parks.html
* University of Barcelona Ethology Lab:
www.ub.es/SERP/EtoHom/index.html
* Psychologists for the Ethical Treatment of Animals:
www.envirolink.org/arrs/psyeta/index.html
* Psychology Preprint Bulletin Board:
www.ccsnet.com/prep
Enrichment Device Ideas
Anne Small, an intern at UTMD Anderson Science Park, is compiling data on enrichment devices currently in use around the world. She hopes to update or offer something similar to Chimpanzoo's Enrichment Manual and Sally Boysen's work. Please send contributions to: Anne Small, 2605 Whitis Ave, #215, Austin, TX 78705 [e-mail: [email protected]]. Please try to include the following: materials and cost, assembly instructions or diagram, study data on use (if applicable), and any other relevant information. Please indicate if you wish the submitted material to be cited in any way other than bibliographical form.
* * *
Volunteer, Costa Rica
Jardín Gaia, a wildlife Rescue Center in Quepos, Costa Rica, is seeking an unskilled volunteer to take care of animals, including cleaning cages and distributing food. This person must be able to volunteer for at least 2-3 weeks. Ability to speak some Spanish is helpful. A recent tetanus shot and T. B. certification is required.
All volunteers are charged $10 per day to cover cook's salary and food for 3 meals. Lodging and transportation within the country are provided. Contact Jardín Gaia Official Wildlife Rescue Center, P.O. Box 182, Quepos, Costa Rica, 6350 [506-777-0535; fax: 506-777-1004; e-mail: [email protected]].
Wildlife Capture Course for Veterinarians
A wildlife capture course for veterinary surgeons will be presented in the Kruger National Park, South Africa by the Price Forbes Chair in Wildlife Diseases (University of Pretoria) and the National Parks Board, 2-8 June 1996 (repeated 25-31 August). The course will include training in capture of a variety of species, including elephants, antelopes and lions, and darting techniques from vehicles and helicopter It will cost US$1100. For more information, contact Prof. D. G. A. Meltzer, Dept. of Veterinary Tropical Diseases, Fac. of Vet. Sci., Onderstepoort, 0110 South Africa [27 12 529 8077; fax: 27 12 529 8312; e-mail: [email protected]]; or Dr E. Stan Miller (Stan) [27 12 529 8229 or 27 12 573841; e-mail: [email protected]].
Humane Society Seeks Summer Intern
The Animal Research Issues Section of The Humane Society of the United States (HSUS) is seeking a summer intern to help with various projects and gain valuable professional experience with animal protection. This paid position lasts three months, beginning sometime between May 1 and June 1. The successful applicant will be a college student (high-school graduates are eligible), preferably pursuing studies in the life sciences, and will have a demonstrable commitment to animal advocacy. The work location will be the HSUS's operations center located in Gaithersburg, MD (a suburb of Washington, DC).
Please send a cover letter and resume to: Jonathan Balcombe (Associate Director for Education, Animal Research Issues), HSUS, 2100 L Street, NW, Washington, DC 20037 [301-258-3046; fax: 301-258-3082; e-mail: [email protected]].
Wildlife Research Expedition, Peru
The Dallas Zoological Society and the Detroit Zoological Institute announce their Third Wildlife Research Expedition to Peru to study and census rare primates of the Peruvian Amazon, June 5-19, 1996. A group of volunteers will work with experienced zoo personnel, local assistants, and Peruvian students to map the travels of seven species of primates (squirrel, woolly, titi, and two species of capuchin monkeys, sakis, and red uakaris), noting population densities and spacing.
This will not be the usual travel experience. In fact, it is an adventure only for the physically hardy. The primate expedition entails regular eight-hour treks through the forest, swamps, and hills of the rain forest. Occasionally, balancing acts over fallen timber that span streams or gullies may be necessary, as well as clambering over fallen logs, under branches, and around thorny bushes. Always a wary eye is kept for the local herpetofauna. Days on the primate study begin around 6 am and conclude sometime after 8 pm with dinner, discussions, and clean-up.
Living conditions are comfortable but primitive. Participants will sleep in large 3-person tents (or you may bring your own single-person tent), bathe in the stream, and utilize pit toilets. So, pack your sense of humor first and prepare to be amazed by the power of nature in a wonderland where the birds bark like dogs and the primates (and dogs) sing like birds.
Cost of the trip is $2,100, excluding airfare. Some of the trip expenses are tax deductible. For more information please contact: Cynthia Bennett, Research Curator, or Wanda Weaver, Dallas Zoo, 621 East Clarendon Dr.,
Dallas, TX 75203 [214-670-6833; e-mail: [email protected] or [email protected]]; or Scott Carter, Mammal Curator, Detroit Zool. Inst., P.O. Box 39, Royal Oak, MI 48068-0039 [810-398-0903].
Program In Primate Behavior
This summer Florida State University, Panama Canal Branch, will offer an intensive training program in the scientific study of primate behavior. The program, June 16 to July 26, 1996, will be held on the 33-hectare island of Isla Tigre in Gatun Lake. Eight tamarins (Saguinus geoffroyi) have recently been released on the island. These tamarins allow the close approach of observers and are individually identified by bead collars. These will be the primary focus of behavioral and ecological research conducted during the class.
Students taking the program will take concurrent courses in Primate Behavior, Ecology, and Research Topics in Psychology and will therefore earn 9 semester hours of credit for the successful completion of the program. This is an intensive program designed for students who have a serious interest in the study of primate behavior and ecology. The cost with round trip air fare from Florida will be $2,708, while the cost for students from Panama will be $2,237. For more information, contact Ms. Rita Wright, College Programs, 210 A Williams Bldg, Florida State University, Tallahassee, FL 32306-4044 [e-mail: [email protected]].
National Zoo Lecture Center
The National Zoological Park's Lecture Center is recording lectures and placing them on-line in Real Audio format. The URL is http://www.si.edu/organiza/museums/zoo/homepage/hilights/lectures.html.
Primates of the World
The Anthropology Department at California State College, Fullerton, with the help of generous donors, is sponsoring an exhibit and lecture series on "Primates of the World", through October, 1996. The focus will be on conservation and education. The exhibit will be on the third floor of the Humanities Building, on the campus in Fullerton, CA. For more information about the exhibit, contact Lori Sheeran [714-773-2765; e-mail: [email protected]], Joseph Nevadomsky [714-449-5335], or the Anthropology Office [714-773-3626].
Lectures will be at 2:00 Sunday afternoons, in the Gabrieleno Room of the University Center. There will be refreshments. Lot B facing State College Blvd. is the closest free parking. Lectures in the series will include the following April 14, 1996: Wendy Hoole on "The Orangutan" May 12, 1996: Ardith Eudey on "Primate Diversity and Conservation in Thailand and Vietnam: A Study in Contrasts and Similarities" June 16, 1996: Lori Sheeran on "The Ecology and Conservation of Gibbons". Additional lectures will be announced. For information about the talks, contact Anne Handberry, President of the Association of Friends and Docents of the Anthropology Museum at [714-773-3626].
University Research Expeditions Program
The University Research Expeditions Program this year includes expeditions to Costa Rica and Belize. Students of all ages and experience are invited to participate, as well as make a tax-deductible contribution. For information about taking, or leading, expeditions, contact UREP at Desk D04, University of California, Berkeley, CA 94720-7050 [510-642-6586; fax: 510-642-6791; e-mail: [email protected]].
Animal Technician -- Texas Snow Monkey Sanctuary
The Texas Snow Monkey Sanctuary/Arashiyama West is looking for an intern or volunteer for a 6-month position with the possibility of extension. Duties will include, but not be limited to, care and feeding of caged sanctuary animals and assistance in capture and relocation of a large troop of monkeys.
Qualifications include any experience working with wild or captive primates, valid driver's license, and the ability to work unsupervised. A small stipend is offered, and living accomodations are available. Travel, meals, and lodging are provided to interns and volunteers.
Applicants should be prepared to work in extreme climatic conditions (temperatures in south Texas reach above 100deg. in the summer months). There will be a probationary period of one month. All applicants should include resume and references. The applicant may have time to conduct a behavioral project of his/her own.
Contact Tracy Wyman, Asst Director, Texas Snow Monkey Sanctuary, P.O. Box 702, Dilley, TX 78017 [210-378-5775; fax: 1-210-378-5881; e-mail [email protected]].
* * *
A conference entitled "Animal Research in Industry: Current Issues Now and in the Twenty-first Century" will be held by the Scientists Center for Animal Welfare in Baltimore, MD, May 9-10, 1996. For more information, contact Conferences, SCAW, Golden Triangle Bldg One, 7833 Walker Dr., Suite 340, Greenbelt, MD 20770 [301-345-3500; fax: 301-345-3503].
A conference entitled "Ethical Issues of Animal Research" will be held at the Poynter Center for the Study of Ethics, Indiana Univ., Bloomington, May 30-June 1, 1996. It is jointly sponsored with the Kennedy Institute of Ethics, Georgetown University, and is part of an annual series of meetings on animal ethics that the Kennedy Institute has sponsored. This is a multi-disciplinary conference for those interested in the profound questions of ethics and animal use. Well-balanced, contrasting viewpoints concerning the use of animals in biomedical and agricultural research will be presented. Leading experts in the field will address the following topics: The moral standing of animals, including religious perspectives Major philosophical arguments for and against animal research Ethical decision-making in IACUCs Personal ethical conflicts Genetic manipulation of animals. Contact Prof. Kenneth Pimple, Poynter Ctr, Indiana Univ., 410 N. Park Ave, Bloomington, IN 47405 [812-855-0261; Fax: 812-855-3315; e-mail: [email protected]].
The 30th International Congress of the International Society for Applied Ethology will be held in Guelph, Ontario, Canada Wednesday 14th to Saturday 17th August 1996. The congress will include a plenary session on "The Behaviour and Welfare of Laboratory Animals". Further information may be obtained from Dr. Ian Duncan [e-mail: [email protected]].
New World Primate Taxon Advisory Group meeting, 19 May, 1996, follows the Western Regional American Zoological Association meetings in Denver, CO. It is intended to bring together managers and scientists who are investigating some aspect of New World Primate genetics. Contact: Jean Dubach, Brookfield Zoo, Dept of Conservation Biology, Genetics Lab, 3300 Golf Rd, Brookfield, IL 60513 [708-485-0263 Ext. 502; Fax: 708-485-3532; e-mail: [email protected]].
The Twelfth Annual Meeting of the Primate Society of Japan will be held 27-29 June, 1996 at the Osaka University Convention Center, Suita City, Osaka. Contact: PSJ Annual Meeting Organizing Committee [81-6-879-8045; Fax: 81-6-879-8010; e-mail: [email protected] or [email protected]].
The 35th Annual Conference of the Canadian Association for Laboratory Animal Science / L'association canadienne pour la technologie des animaux de laboratoire (CALAS/ACTAL) will be held July 8-10, 1996, in Charlottetown, Prince Edward Island, Canada. The theme of the Conference is "Prawns to Primates - Back to the Future". The program includes workshops, scientific sessions in laboratory animal science, a poster session, and an autotutorial section. Scheduled workshops will include: Post-operative Analgesia in Laboratory Animals; Facility Design and Operational Considerations in the Use of Cage Isolation Equipment; and Documentation and Reports for an Effective Animal Care and Use Program (CCAC sponsored). For more information please contact: Dr. Don McKay, CALAS/ACTAL National Office, Biosciences Animal Service, CW 401 Biological Sciences Building, Edmonton, Alberta, Canada T6G 2E9 [403-492-5193; Fax: 403-492-7257; e-mail: dmckay@ gpu.srv.ualberta.ca].
The Scientists Center for Animal Welfare (SCAW) and the Univ. of Texas Health Science Center will co-sponsor a Conference on National IACUC Affairs on December 5-6, 1996 in San Antonio, TX. The conference will be based on the results of SCAW's national survey of IACUCs in academic and industrial institutions. Members of IACUCs, researchers, regulatory personnel, veterinarians, and administrators are among those invited to attend. For more information, contact SCAW (address above).
The Australian Primate Society's Annual Meeting will be held 6-8 December, 1996, at the Wellington Zoo, Wellington, New Zealand. For more information, contact Graeme Crook, CSIRO Div. of Human Nutrition, Animal Services, Majors Rd, O'Halloran Hill, South Australia 5158. [6182980336; Fax: 6183770004; e-mail: [email protected]].
The BioAnthro wing of the Harvard Dept. of Anthropology is sponsoring a series of talks at the Peabody Museum's Putnam Lab, all at 3:30 pm on Fridays and open to the public. 26 April, Susan Lipson will speak on "The impact of variation in ovarian hormone levels on the probability of successful conception;" 10 May, Alan Mootnick will speak on "Gibbon systematics and conservation."
The Primate-Talk Calendar is available on the PRIMATE INFO NET via the WRPRC Web Page (http://www.primate.wisc.edu/pin) or their gopher site (gopher.primate.wisc.edu).
* * *
Animal Services Operations Manager, Johns Hopkins
The Division of Comparative Medicine of the Johns Hopkins University School of Medicine is seeking an Operations Manager for its section of Animal Services. This individual will supervise a staff of 42 animal care technicians in the daily care of over 20,000 animals ranging from rodents to domestic livestock and nonhuman primates. The AAALAC-accredited institutional animal holding facilities comprise approximately 95,000 square feet on three geographically separated campuses. The Operations Manager is responsible for overseeing all aspects of day-to-day animal husbandry, purchasing and maintenance of supplies and equipment, and hiring, training, and direction of a large technical staff. He or she must be able to interact with research investigators and staff on questions related to animal care and housing.
The requirements for this position include (a) a Bachelor's degree in animal husbandry or biological science and five years of relevant experience or (b) ten years of such experience in laboratory animal facility management; good oral and written communicative skills; and an in-depth knowledge of legal requirements and guidelines. This position offers excellent benefits.
Send resume with salary expectations referring to position 96M2132 to Don Horan, Sr. Employment Specialist, School of Medicine, The Johns Hopkins University, P.O. Box 2454, Baltimore, MD 21203-2454.
Animal Facility Supervisor, Athens, GA
There is an opening for a Animal Facility Supervisor to join the team of dedicated animal care technicians in the University of Georgia's College of Arts & Science's Animal Resources program. The position entails supervising one or more of the facilities for small animals, including nonhuman primates. There is a staff of two full-time care technicians and 6-10 part-time students. This particular position is directly linked to the polyclonal antibody production service. The successful candidate will have experience in supervising student technicians, and should have a BS degree in Animal Science. AALAS certification is prefered, but can be obtained once in the position. Interested persons can contact Robin M. Kavanaugh, Animal Resources Manager, Franklin College of Arts & Sciences, B021 Life Science Building, University of Georgia, Athens, GA 30602 [706-542-6083; fax: 706-542-0149; e-mail: [email protected]].
Primate-Jobs
The Wisconsin Regional Primate Research Center has announced the availability of an internet-based jobs listing service for the field of primatology. Primate-Jobs can be accessed at http://www.primate.wisc.edu/pin/jobs. Currently this service is located in the Primate Info Net menu, "WRPRC Library Outreach Services."
Organizations with positions to fill and individuals looking for jobs should post this information on the Primate-Jobs board. In addition to descriptive information about the position, you will be asked to select the type of position available or wanted -- professional, post-doctoral, field, internship or volunteer -- and also to set a date when the posting will expire. All interactions with the job service are electronic and can be managed from your computer.
Those looking for placement opportunities in primatology can consult the "Positions Available" listings. Organizations seeking potential candidates can review the "Positions Wanted" listings. Note that since this service is just beginning, it will be a while before a significant number of listings is available.
Any job -- research, teaching, curatorship, TAG member, field worker, sanctuary volunteer, foundation president, zoological garden docent -- which relates to nonhuman primates can be listed. The position must have some involvement with primate research, conservation, or education, and may be in disciplines such as biomedicine, anthropology, or zoology. Positions in conservation agencies, primate centers or laboratories, government organizations, educational programs, sanctuaries, zoological gardens, museums or other primate-related programs are welcome.
Sabbatical Replacement, Kansas
Tabor College, a small church-related undergraduate institution located in Hillsboro, Kansas, announces a one-year sabbatical leave replacement position for 1996-97 for a person who can teach courses in general zoology, ecology, environmental science, and general botany. If you would enjoy living in a small town and teaching in an atmosphere that emphasizes small classes and maximum student interaction, please contact Dr. Lon Fendall, Academic Dean, Tabor College, Hillsboro, KS 67063 or Max Terman at [email protected] for more information.
Director of Research and Conservation, Denver Zoo
The Denver Zoological Gardens is seeking to fill a newly established position, with responsibility for the development and implementation of research and conservation programs. This position requires an earned doctoral degree in a scientific field related to the conservation of animal populations; several years of postdoctoral zoological research; strong managerial and communications experience; and a proven record of grant development and scientific publication. It will require the application of a broad range of administrative, financial, and communictions skills; the ability to work effectively with employees, volunteers, board members, and the general public; and ongoing leadership and oversight of scientific research projects.
Please do not contact or submit materials to the Denver Zoological Gardens. Send letter, including salary history and current resume to: Executive Search, Management Advisors, Inc., 5675 DTC Blvd., Suite 230, Englewood, Colorado 80111-3216.
Lefini Gorilla Manager
Howletts Wild Animal Park, home of the world's largest captive gorilla colony, manages a rescue and rehabilitation project for lowland gorillas in the Republic of the Congo, Central Africa. We are seeking someone of senior keeper/assistant curatorial status, with several years of proven hands-on experience in managing great apes, some experience of the tropics (preferably Africa), and an ability to work under harsh and difficult circumstances. The successful applicant will be in good health and physcially robust, and will be able to work as part of a team. The language of the country is French and an ability to speak French is essential. We can provide travel, meals, and lodging for interns and volunteers. The manager may be single or married, but the local circumstances do not allow for children. The Congo has two rainy seasons per annum, and tropical diseases are common. We are offering a 2- or 3-year contract which includes negotiable salary, private health insurance, accommodation, transport, weekly living allowance and annual return airfares to the manager's home country. Contact Mr. Mike Lockyer or Mr. Chris Furley, Port Lympne Wild Animal Park, Lympne, Hythe, Kent, U.K. CT21 4PD [fax: 44-1303-264944].
* * *
Marc Bekoff, 777 Juniper Ave, Boulder, CO 80304-1763.
Steven G. Gilbert, Biosupport, Inc., 14716 NE 87th St., Redmond, WA 98052-3400.
Stan Dannemiller, Baxter Healthcare Corp., Route 120 & Wilson Rd., WG2-1S, Round Lake, IL 60073-0490.
Stephen P. Easley, Easley and Associates, Professional Consultants, 1403 Granada, Alamogordo, NM 88310.
Dr. A. E. James, Chinese University, Laboratory Animal House, Shatin, NT., Hong Kong.
Journal of Medical Primatology, Prof. Jorg Eichberg, Editor, Biomedical Primate Research Centre, P.O Box 3306, 2280 GH Rijswijk, Netherlands.
Stephanie Torlone, 13601 E. Palomino Dr., Fort Lauderdale, FL 33330-2232.
* * *
Advantages:
1. No parasites.
2. No snakes roaming around.
3. You don't have to look for the animals.
4. It's cheap.
5. You know what the monkeys are eating and, in the case of multiple species housing, you know who displaces whom on feeding competition.
Disadvantages:
1. Children.
2. Your ears might no longer be functional after a week from the screaming. Buy earplugs.
Conclusions:
1. The zoo is a wonderful place.
2. Just don't let the people in.
Michael Passman, New Mexico State University -- as posted on primate-talk
* * *
Univ. of Washington Press Release
Officials at the University of Washington's Primate Field Station at Medical Lake, WA, responded to the death of a 5-month-old macaque monkey last November by notifying the U.S. Department of Agriculture's Animal and Plant Health Inspection Service, dismissing the employee responsible for the incident, and taking steps to prevent a recurrence.
As part of a regular site inspection, USDA inspectors on Dec. 7 confirmed that the macaque had died from apparent water deprivation. The water supply to the room where the animal was housed had been turned off over a weekend.
A post-mortem examination by field station personnel showed that the animal had been ill and possibly dehydrated before being deprived of water. The USDA report stated that the death would likely not have occurred if the animal had not had a prior medical problem, since no other animal in the room died.
The employee responsible for the room was dismissed from employment for failing to check the automatic watering device and for entering a false report that indicated the device had been checked.
The facility has taken appropriate steps to prevent a recurrence of this type of incident. -- University of Washington Press Release, January 5, 1996
Gorilla Birth at Los Angeles Zoo
A gorilla was born at the Los Angeles Zoo on 17 January, 1996. The infant, a female, was born to Angel, an 8-year-old female, and Tzambo, a 23-year-old male. This was Angel's first pregnancy; she was born at the zoo and reared in her natal group. Tzambo is wild-caught, and has sired only one other offspring. The infant was full term; weight at birth was 5 lbs. 10 1/2 oz., gestation was 257 days.
Unfortunately, the infant was found abandoned on the floor, unmoving, and her body temperature was below 74 degrees. After the infant was warmed and stabilized, we attempted to re-introduce her, but the mother showed no interest in her. She is being hand-reared in the zoo's nursery. She is strong and appears to be thriving at this writing, and we hope to attempt an early introduction to her natal group.
In L.A. we have made every attempt to allow females to rear their infants since we stopped routinely pulling all gorillas at birth in 1987. That year, we had three infants born, all reared by their mothers. Of these three mothers, two were hand-reared with no prior offspring and no experience with infants, and one was a mother who had had several infants pulled. Our philosophy is to leave infants with the mother unless leaving it obviously risks the infant's life, as was the case here. Interestingly, this mother was mother-reared in a stable group with another infant 4 months older than she was. In other words, she had everything going for her. -- From reports on Primate-Talk by Jennifer J. Chatfield, Gorilla Keeper, Los Angeles Zoo
Life and Death at Woodland Park Zoo
Woodland Park Zoo announced the birth of Jumoke and Congo's first western lowland gorilla baby. The five-and-a-half-pound female was born Monday, February 12, at 11:23 pm PST. Jumoke and Congo's infant marked the seventh successful gorilla birth at Woodland Park Zoo.
Congo was born in Cameroon around 1959. On breeding loan from the Honolulu Zoo, Congo came to Woodland Park from Phoenix Zoo in 1992. His transfer to Woodland Park provided his first-ever experience with a grassy, naturalistic habitat. Congo had never successfully mated with his two previous mates, Fifi and Hazel, at Honolulu and Phoenix, respectively.
Just two weeks after his daughter's birth, on February 27, Congo suffered a seizure and died suddenly while being examined by the animal health staff. Gorilla keepers noticed on the 25th that Congo was lethargic. He showed very little interest in eating or drinking and behaved as if he was suffering from some non-specific pain. Zoo animal health staff initially diagnosed Congo to be afflicted with Balantidium, causing diarrhea and excessive gas. He was treated with an antibiotic and ibuprofen for pain. The keepers were offering fluids and treats to encourage him to eat at 10:00 am on the 27th, when he suffered a seizure. The animal health staff immobilized Congo to further examine him and assess his condition but, while they were examining him, he stopped breathing and died in spite of resuscitation attempts.
The preliminary postmortem indicated no obvious cause of death. Complications related to age (average life expectancy for a captive gorilla is 40 years) and a very severe systemic infection, possibly associated with abscessed teeth, were apparent. A fractured upper left canine and an upper right second molar were found to be abscessed (he had had prior dental problems resulting in root canals and fillings). Also found was a systemic infection causing infarcts, including 50% loss of left kidney and small portions of the liver. There was also hemorrhage in the lungs, possibly as a result of resuscitation attempts, and some lesions in the heart. The actual cause of death will not be known until a histology review.
Congo epitomized the true "gentle giant" spirit of his species and was a wonderful animal to work with and to be around. The zoo's staff is devastated by this loss. We hope to to attempt an early introduction to her natal group. -- Laurence Gledhill, Woodland Park Zoo, on Primate-Talk
Chimpanzee with AIDS Euthanized
The first animal to develop the clinical disease of Acquired Immune Deficiency Syndrome (AIDS) as a result of infection with the human immunodeficiency virus (HIV-1) was euthanized February 13. The animal, a 15-year-old Yerkes Primate Research Center chimpanzee named Jerom, was humanely euthanized because medical tests, including one conducted that morning, indicated that his anemia was becoming more severe and untreatable. Anemia, which also occurs in people with AIDS, results from the failure of the bone marrow to produce adequate numbers of oxygen-carrying red blood cells.
"There was no evidence that Jerom was in pain before his death," said Thomas R. Insel, M.D., director of the Yerkes Center, which is part of Emory University. "But, it was clear to the veterinarians, the researchers and myself that Jerom had reached terminal stages of the disease and was beyond the point of recovery. We wanted to prevent further deterioration of his clinical condition. There was little likelihood that additional information would be gained by prolonging his life."
With Jerom's death, the number of Yerkes chimpanzees involved in AIDS research totals 12. Thus far, none of these animals, all of which live in pairs for social companionship, shows signs of AIDS. They will remain the core of the AIDS research efforts with chimpanzees, since there are no immediate plans to infect additional chimpanzees.
"Whether you're working with human patients or with nonhuman animals, AIDS research continues to raise ethical issues within the scientific and medical communities and within the public at large," Dr. Insel pointed out. "Studying and caring for the first animal with AIDS has been challenging. We have treated opportunistic infections aggressively but ultimately we were left fighting a relentless disease for which there still is no cure."
Jerom was one of 13 chimpanzees at Yerkes who were inoculated with HIV-1 as part of a research study conducted with the U.S. Centers for Disease Control and Prevention in Atlanta, to develop a primate model for research, including vaccine studies. The project was initiated over 10 years ago. Like most of the animals at Yerkes, the 13 chimpanzees were captive-born for research purposes.
Although Jerom became persistently infected with HIV -- his immune system produced antibodies against the virus, and virus was isolated repeatedly from samples of his blood -- from the time he was infected over a decade ago, clinical signs of AIDS did not occur until late summer of 1995.
Although he did not receive anti-viral drugs, Yerkes veterinarians treated Jerom for clinical disease that occurred secondarily to the HIV infection. In addition to receiving extensive care from Yerkes veterinarians, Jerom also was the focus of a great deal of personal attention from the center's primate caretakers and the center's behavioral scientist who is in charge of enrichment of the laboratory animals' environment.
Yerkes researchers and animal care experts also have been paying close attention to Nathan, one of the 12 remaining chimpanzees. He received an experimental blood transfusion from Jerom in September 1995. Nathan's CD4+ cells have dramatically declined but he has not yet developed clinical AIDS. That the virus affected Nathan's immune system more quickly than Jerom's was not a surprise, explained Dr. Harold McClure, who heads the Yerkes AIDS research studies, since scientists know that "passage" of an infectious agent from one animal to another can alter the virus and increase its pathogenicity.
The development of clinical AIDS in Jerom was reported by Yerkes scientist Dr. Frank Novembre at a scientific conference last fall, and on Jan. 30 at the Conference on Retroviruses and Opportunistic Infections in Washington, D.C.
Yerkes scientists have initiated studies to determine whether the HIV isolate from the chimpanzee with AIDS can infect and induce disease in pigtailed macaque monkeys. "The availability of a more readily available, less expensive monkey model for HIV-1 infection would be an extremely important animal model system for studies of the pathogenesis of HIV-1-induced disease as well as for the evaluation of AIDS drugs and vaccines," said Dr. McClure, Associate Director for Research Resources at Yerkes. -- From a February 13 press release by the Yerkes Primate Research Center, Emory University
* * *
(Addresses are those of first authors)
Books
Environmental Enrichment in Captive Primates: A Survey and Review. L. A. Dickie. M.Phil. Dissertation, 1994. 88 pp. [Price: [[sterling]]10.00/US$25.00, from UFAW, 8 Hamilton Close, South Mimms, Potters Bar, Hertfordshire EN6 3QD, England]
Formulary for Laboratory Animals. C. T. Hawk & S. L. Leary (Compilers). Ames, IA: Iowa State Univ. Press, 1995. [Price: $17.95. Order from ISU Press, 2121 S. State Ave, Ames, IA 50014-8300]
Audiovisual Material
Science in Action. Foundation for Biomedical Research. Video for grades K-4. Eight minutes. [Price: $20, including activity booklet and sheets, from FBR, 818 Connecticut Ave, NW, Suite 303, Washington, DC 20006]
Primates of the Tropical Forest (1996 Calendar). [Price: USA $11.95; Canada: $16.95, from Duke University Primate Center, 3705 Erwin Rd, Durham, NC 27705]
Bibliographies
Hepatitis Research in Nonhuman Primates. A Selective Bibliography, 1992-1994 Update. D. Paros. Seattle: Primate Information Center, 1995, 19 pp. (245 citations, primate & subject indexes) [Price: $6.50. Stock #95-003. Order from PIC, RPRC, Univ. of Washington, SJ-50, Seattle, WA 98195]
Cardiovascular Pharmacology in Primates (Exclusive of Atherosclerosis). A Selective Bibliography, 1985-January 1995. M. McLean. Seattle: Primate Information Center, 1995, 22 pp. (249 citations, primate & subject indexes) [Price: $6.50. Stock #95-004. Ordering information same as above]
Booklets
Annual Resource Guide 1995: Special Edition of Continuing Listings. Seattle: Primate Information Center, 1995. [PSIC, PIC, RPRC, Box 357330, Univ. of Washington, Seattle, WA 98195-7330]
Careers with Animals (revised edition). 12 pp. UFAW, 1995. Price: [[sterling]]0.50/US$1.50, from UFAW, 8 Hamilton Close, South Mimms, Potters Bar, Hertfordshire EN6 3QD, England]
Magazines and Newsletters
AAALAC Communiqué, Fall, 1995 [11300 Rockville Pike, Suite 1211, Rockville, MD 20852-3035]
Animal Welfare, February, 1996, 5[1].
. .
Contents include "On comparing the behaviour of zoo- housed animals
with wild conspecifics as a welfare indicator," by J. S. Veasey, N. K. Waran
& R. J. Young; and "Space utilization by captive-born baboons (Papio
sp.) before and after provision of structural enrichment," by A. L. Kessel
& L. Brent.
Animal Welfare Information Center Newsletter, Winter 1995/1996, 6[2-4].
Annales de la Fondation Fyssen, 4th trimester, 1995, No. 10. [194, rue
de Rivoli, Paris 1e, France]
. .
Reports in French, including "Le développement des bases neurales de la
reconnaissance visuelle chez le primate," by O. Pascalis.
Boletín de la Asociación Primatológica Española, Enero 1996, 3[1]. [Área de Psicobiol., Fac. de Psicol. (Dpcho. 36), Univ. Autónoma de Madrid, 28049-Madrid, Spain]
Chinese Primate Research and Conservation News, 1995, 4[1]. [Primate
Specialist Group, Mammalogical Soc. of China, 19 Zhongguancun Lu, Inst. of
Zool., Acad. Sinica, Beijing 100080, China]
. .
Abstracts and announcements, in English and Chinese.
Gorilla Gazette, December 1995, 9[1]. [Columbus Zool. Park
Assoc., 9990 Riverside Dr., Box 400, Powell, OH 43065-0400]
. .
Reports on gorillas (and other apes) in various zoos and other locations.
Gorilla Gazette, January 1996, 10[1].
. .
Includes a personal report on the tragic recent fire at the Philadelphia Zoo,
by J. Unger, the Head Keeper at World of Primates.
IPPL News, November 1995, 22[3]. [IPPL, P.O. Box 766, Summerville, SC 29484]
Noldus News, 1995, 2[2]. [Costerweg 5, P.O. Box 268, 6700 AG
Wageningen, Netherlands]
. .
Noldus Observer is available for Windows.
Our Animal WARDS, Winter, 1996. [WARDS, Inc., 1660 L St N.W., Suite 612, Washington, DC 20036-5603].
Pongo Quest: Newsletter of the Orangutan Foundation International,
Fall/Winter 1995, 7[1]. (O.F.I., 822 S. Wellesley Ave, Los Angeles, CA
90049)
. .
News, announcements, and an article on captive orangutan management by L.
Perkins.
Positively Primates, 1995, 1[3-4]. [DuMond Conservancy, 14805 S.W. 216 St, Miami, FL 33170]
The Newsletter, 1996, 7[3]. [Primate Fnd. of Arizona, P.O. Box
20027, Mesa, AZ 85277-0027]
. .
Includes chimpanzee diets from the Dallas and Auckland Zoos.
Proceedings
Human/Research Animal Relationship. L. Krulisch, S. J. Mayer, & R. C. Simmonds (Eds.). Greenbelt, MD: SCAW, 1996. [Price: $35. SCAW, Golden Triangle Bldg One, 7833 Walker Dr., Suite 340, Greenbelt, MD 20770]
Reports
Primate Report, 1995, 43. [German Primate Center (DPZ),
Kellnerweg 4, 37077 Göttingen, Germany]
. .
The Annual Scientific Report 1994 of the German Primate Center.
Primate Report, 1996, 44. [Address same as above]
. .
Abstracts of the 4th Congress of the Gesellschaft für Primatologie. Also
"Some hematological and biochemical values of normal blood in captive
moustached tamarins (Saguinus mystax)," by V. F. Poleschuk et
al.; "The present status of living primates and experimental primate research
in China," by B. P. Tian, W. Z. Ji, & Y. Z. Peng; "A survey of Hoolock
gibbon (Hylobates hoolock) in Southern Assam, India," by A. Choudhury;
and abstracts from a workshop on fruit selection and seed dispersal by
primates.
Primate Report, 1996, 45. [Address same as above]
. .
Papers from the first IPS Symposium on the History of Primatology, held during
the XVth Congress of the IPS.
Special Journal Issues
Callitrichid social structure and mating system: Evidence from field
studies. American Journal of Primatology, 1996, 38[1].
. .
Contents: Callitrichid social structure and mating system: Evidence from
field studies, by A. Savage & A. J. Baker. Habitat and the evolution of
social and reproductive behavior in Callitrichidae, by A. B. Rylands. Wild
Callithrix groups: Stable extended families? by S. F. Ferrari & L.
J. Digby. Social and seasonal influences on reproductive biology in male
moustached tamarins (Saguinus mystax), by P. A. Garber, L. Moya, J. D.
Pruetz, & C. Ique. Immigration in wild groups of golden lion tamarins
(Leontopithecus rosalia), by A. J. Baker & J. M. Dietz. Saddle-back
tamarin (Saguinus fuscicollis) reproductive strategies: Evidence from a
thirteen-year study of a marked population, by A. W. Goldizen, J. Mendelson, M.
van Vlaardingen, & J. Terborgh. Demography, group composition, and
dispersal in wild cotton-top tamarin (Saguinus oedipus) groups, by A.
Savage, L. H. Giraldo, L. H. Soto, & C. T. Snowdon. Social behavior of
wild moustached tamarins, Saguinus mystax, at the Estación
Biológica Quebrada Blanco, Peruvian Amazonia, by E. W. Heymann.
Nonhuman primate models for AIDS VI. Journal of Medical Primatology,
1995, 24[3]. Guest Editor: J. W. Eichberg.
. .
Contents: Introduction, by J. Moor-Jankowski. AIDS-like disease following
mucosal infection of pig-tailed macaques with SIVsmmPBj14, by P. N. Fultz, R.
Schwiebert, & J. Stallworth. Isolation and characterization of the first
simian immunodeficiency virus from a feral sooty mangabey (Cercocebus atys)
in West Africa, by Z. Chen, P. Telfer, P. Reed, L. Zhang, A. Gettie, D. D.
Ho, & P. Marx. Origins of simian immunodeficiency virus infection in
macaques at the New England Regional Primate Research Center, by K. G.
Mansfield, N. W. Lerche, M. B. Gardner, & A. A. Lackner. VCAM-1 expression
and leukocyte trafficking to the CNS occur early in infection with pathogenic
isolates of SIV, by V. G. Sasseville, J. H. Lane, D. Walsh, D. J. Ringler,
& A. A. Lackner. Differences in viral distribution and cell adhesion
molecule expression in the intestinal tract of rhesus macaques infected with
pathogenic and nonpathogenic SIV, by J. D. Stone, C. C. Heise, D. R. Canfield,
M. J. Elices, & S. Dandekar. Assessing genetic-based therapies for AIDS
using the simian immunodeficiency virus, by C. L. Endres, E. Berg-quam, M. K.
Axthelm, & S. W. Wong. Neutralising epitopes of simian immunodeficiency
virus envelope glycoprotein, by K. A. Kent. Fourth annual survey of worldwide
HIV, SIV, and SHIV challenge studies in vaccinated nonhuman primates, by J. T.
Warren & M. A. Levin.
1996 Buyers Guide. Lab Animal, 1995, 24[11]
Human health significance of animal models. Lab Animal, 1996,
25[2].
. .
Includes "The rhesus monkey: Providing insight into obesity and diabetes," by
N. Bodkin.
Systematic Review of Southeast Asian Longtail Macaques: Macaca fascicularis (Raffles, 118211). J. Fooden. Fieldiana, Zoology New Series, No. 81. [Price: $35.00, Field Museum of Natural History, Library--Publ. Div., Roosevelt Rd at Lake Shore Dr., Chicago, IL 60605-2498]
Supplements
Neotropical Primates, 1995, 3, Suppl. [Conservation
International, Minas Gerais, Brazil]
. .
"Species and subspecies of neotropical primates: Conservation status
categories adopted by the IUCN in 1994."
Anatomy & Physiology
Correlation of CSF 5-HIAA concentration with sociality and the timing
of emigration in free-ranging primates. Mehlman, P. T., Higley, J. D.,
Faucher, I., Lilly, A. A., Taub, D. M., Vickers, J., Suomi, S. J., &
Linnoila, M. (LABS, P.O. Box 557, Yemassee, SC 29945). American Journal of
Psychiatry, 1995, 152, 907-913.
. .
In adolescent male rhesus macaques living in naturalistic settings, CSF 5-HIAA
concentration is positively correlated with affiliative sociality. Animals
with low CSF 5-HIAA concentrations exhibit less social competence and emigrate
from their social groups at a younger age than those with higher
concentrations.
Social effects and circadian rhythms in squirrel monkey pituitary-adrenal
activity. Lyons, D. M., Ha, C. M. G., & Levine, S. (Dept of Psychiatry
& Behav. Sci., Stanford Univ. School of Med., Stanford, CA 94305).
Hormones and Behavior, 1995, 29, 177-190.
. .
Observations and measurements suggest that squirrel monkeys housed without
companions hypersecrete cortisol at a morning peak because adrenal
responsiveness to ACTH is enhanced. Low circulating ACTH levels in turn are
maintained by robust glucocorticoid feedback mechanisms that inhibit the
synthesis or release of pituitary ACTH.
Primate rod and cone photoreceptors may differ in glucose accessibility.
Nihira, M., Anderson, K., Gorin, F. A., & Burns, M. S. (M. S. B., Dept of
Ophthalmology, UC Davis School of Med., 1603 Alhambra Blvd, Sacramento, CA
95816). Investigative Ophthalmology & Visual Science, 1995, 36,
1259-1270.
. .
Brain glycogen phosphorylase immunoreactivity was found in cone, but not rod,
photoreceptors in the eyes of adult humans and monkeys. Rod and cone
photoreceptors have the biochemical capability to transport exogenous glucose
from the circulation. Cones could be more resistant to acute reductions in
circulating glucose during hypoglycemia. However, during hypoxic insult,
glycogen-olysis and anaerobic glycolysis could result in increased production
of intracellular lactic acid, potentially predisposing the cone to acidotic
damage.
Recombinant human megakaryocyte growth and development factor stimulates
thrombocytopoiesis in normal nonhuman primates. Farese, A. M., Hunt, P.,
Boone, T., & MacVittie, T. J. (Exp. Hematology, Armed Forces Radiology
Research Inst., 8901 Wisconsin Ave, Bethesda, MD 20889-5603). Blood,
1995, 86, 54-59.
. .
Recombinant human megakaryocyte growth and development factor (r-HuMGDF) was
administered subcutaneously to normal, male rhesus monkeys once a day for 10
consecutive days at 2.5, 25, or 250 micro-g/kg of body weight. Results suggest that
the single protein MGDF is a major physiological regulator of
thrombocytopoiesis.
Hematological and serum biochemical values in pregnant and postpartum females
of the squirrel monkey (Saimiri sciureus). Suzuki, T., Suzuki, N.,
Shimoda, K., & Nagasawa, H. (Inst. of Ozaki Lab., Hamri Co, Ltd., 2638-2
Ozaki, Sanwamachi, Sashima-gun, Ibaraki 306-01, Japan). Experimental
Animals, 1996, 45, 39-43.
. .
Values from 18 animals, from 12 weeks before to 12 weeks after normal births,
plus control values from nonpregnant females.
Regional distribution and characterization of nitric oxide synthase activity
in the brain of the common marmoset. Gerlach, M., Oehler, D., Blum-Degen, D.,
Lange, K. W., Mayer, B., Reichmann, H., & Riederer, P.
(Univ.-Nervenklinik, Abt. Klin. Neurochem., Julius-Maximilians-Univ.,
Füchsleinstr. 15, D-97080 Würzburg, Germany). NeuroReport,
1995, 6, 1141-1145.
. .
The brain of the common marmoset shows no hemispheric asymmetry of specific
NOS activity.
Animal Models
The pathogenicity of behavior and its neuroendocrine mediation: An
example from coronary artery disease. Manuck, S. B., Marsland, A. L., Kaplan,
J. R., & Williams, J. K. (Behavioral Physiology Lab., 506 Old Engineering
Hall, 4015 O'Hara St, Univ. of Pittsburgh, Pittsburgh, PA 15260).
Psychosomatic Medicine, 1995, 57, 275-283.
. .
In cynomolgus monkeys several psychosocial variables, e.g., social instability
and behavioral dominance and subordination, promote coronary atherogenesis,
either independently or in interaction. Animals exhibiting a heightened
cardiac responsivity to stress also develop the most extensive coronary
lesions. Social stress causes injury to arterial endothelium and, among
chronically stressed animals, impairs endothelium-dependent vasomotor responses
of the coronary arteries.
Chemotherapy-induced hemolytic uremic syndrome: Description of a
potential animal model. Hillyer, C. D., Duncan, A., Ledford, M., Barrett, T.
J., Klumpp, S. A., Anderson, D. C., McClure, H. M., & Winton, E. F. (Emory
Univ. Hospital Blood Bank, 1364 Clifton Rd, NE, Atlanta, GA 30322). Journal
of Medical Primatology, 1995, 24, 68-73.
. .
Data from four rhesus monkeys that developed fatal hemolytic uremic syndrome
after high-dose chemotherapy indicate that these animals may model this
syndrome in oncology and bone marrow transplant patients.
Tumor necrosis factor-alpha in serum of macaques during SIVmac251 acute
infection. Clayette, P., Le Grand, R., Noack, O., Vaslin, B., Le Naour, R.,
Benveniste, O., Théodoro, F., Frétier, P., & Dormont, D.
(Lab. Neuropath. exp. de Neurovirologie, CEA/DSV/DPTE/SSA, 60-68 ave. Division
Leclerc, B.P.6, 92265 Fontenay aux Roses, Cédex, France). Journal of
Medical Primatology, 1995, 24, 94-100.
. .
SIVmac251-infected cynomolgus macaques could be a model to explore the
interdigitation existing between HIV and tumor necrosis factor in acute and
chronic infection, and to develop new therapeutic strategies.
Zidovudine treatment prolongs survival and decreases virus load in the central
nervous system of rhesus macaques infected perinatally with simian
immunodeficiency virus. Rausch, D. M., Heyes, M. P., Murray, E. A., &
Eiden, L. E. (Sect. on Molecular Neurosci., Lab. of Cell Biology, NIMH, Bldg
36, Rm 3A17, Bethesda, MD 20892). Journal of Infectious Diseases, 1995,
172, 59-69.
. .
Two of 5 infected, untreated monkeys developed motor impairment within 6
months following infection; motor impairments did not occur in infected,
zidovudine-treated monkeys until after the drug was discontinued.
Reactivation of human immunodeficiency virus type 2 in macaques after simian
immunodeficiency virus SIVmac superinfection. Petry, H., Dittmer, U.,
Stahl-Hennig, C., Coulibaly, C., Makoschey, B., Fuchs, D., Wachter, H., Tolle,
T., Morys-Wortmann, C., Kaup, F.-J., Jurkiewicz, E., Lüke, W., &
Hunsmann, G. (Dept of Virology & Immunology, DPZ, Kellnerweg 4, D-37077
Göttingen, Germany). Journal of Virology, 1995, 69,
1564-1574.
. .
The presence of an apathogenic viral variant seems to retard the disease
occurring after infection with a pathogenic virus, rather than to confirm total
protection. This partial protection appears to depend on a specific
proliferative T-cell response early after infection.
Early intrathecal events in rhesus macaques (Macaca mulatta) infected
with pathogenic or nonpathogenic molecular clones of simian immunodeficiency
virus. Smith, M. O., Heyes, M. P., & Lackner, A. A. (Dept of Clinical
Sci., College of Vet. Med. & Biomed. Sci., Colorado State Univ., Fort
Collins, CO 80523). Laboratory Investigation, 1995, 72,
547-558.
. .
Findings support the following hypotheses of early events in SIV
neuropathogenesis: (a) Pathogenic virus invades the CNS within days of
i.v. inoculation and elicits an intrathecal immune response, including
intrathecal synthesis of IgG and macrophage activation; (b) the immune response
initially is associated with a decreased virus load in the CNS; (c) as
immunodeficiency develops, virus load in the CNS increases once again; and (d)
both virus and host factors are important in determining the course of
events.
Distribution of SIV in lymph nodes of serially sacrificed rhesus monkeys.
Baskin, G. B., Martin, L. N., Murphey-Corb, M., Hu, F.-S., Kuebler, D., &
Davison, B. (Dept of Pathology, Tulane RPRC, 18703 Three Rivers Rd, Covington,
LA 70433). AIDS Research and Human Retroviruses, 1995, 11,
273-285.
. .
One male and one female with high levels of antigenemia and one each with a
lower level were sacrificed at 2, 4, 8, and 24 weeks after inoculation with
SIVDeltaB670. Results showed that disease course is determined within 2 weeks
after experimental inoculation with uncloned pathogenic SIV, and that viral RNA
is widespread in the lymph nodes of animals with rapidly or slowly progressing
disease.
Progression to AIDS in macaques is associated with changes in the replication,
tropism, and cytopathic properties of the simian immunodeficiency virus variant
population. Rudensey, L. M., Kimata, J. T., Benveniste, R. E., &
Overbaugh, J. (J. O., Dept of Microbiology, SC-42, Univ. of Washington,
Seattle, WA 98195). Virology, 1995, 207, 528-542.
. .
This study demonstrates important similarities in the phenotypic viral changes
that accompany development of AIDS in SIV and HIV-1 infections.
Seroepizootiology of Helicobacter pylori gastric infection in nonhuman
primates housed in social environments. DuBois, A., Fiala, N., Weichbrod, R.
H., Ward, G. S., Nix, M., Mehlman, P. T., Taub, D. M., Perez-Perez, G. I.,
& Blaser, M. J. (Dept of Med., Uniformed Services Univ., 4301 Jones Bridge
Rd, Bethesda, MD 20814-4799). Journal of Clinical Microbiology, 1995,
33, 1492-1495.
. .
Plasma collected over a year from 196 rhesus monkeys housed in indoor gang
cages, outdoor corrals, or free-ranging forest demonstrate that these animals
may serve as a model for human infection.
Effects of Helicobacter pylori infection on gastric mucosal
defense factors in Japanese monkeys. Fujiyama, K., Fujioka, T., Murakami, K.,
& Nasu, M. (Second Dept of Internal Med., Oita Med. Univ., Hasama-machi,
Oita, 879-55 Japan). Journal of Gastroenterology, 1995, 30,
441-446.
. .
Colonization by H. pylori injures the gastric mucosa by depressing the
gastric mucosal defense factors and, consequently, the cell kinetics are
accelerated.
Nitroxidergic innervation in dog and monkey renal arteries. Okamura, T.,
Yoshida, K., & Toda, N. (N. T., Dept of Pharmacology, Shiga Univ. of Med.
Sciences, Seta, Ohtsu 520-21, Japan). Hypertension, 1995, 25,
1090-1095.
. .
Renal arteries are innervated by nitric oxide-mediated vasodilator and
adrenergic vasoconstrictor nerves, and depression of the vasodilator nerve
function potentiates the contraction caused by adrenergic nerve excitation.
Rhesus macaque with cecal fistula: A model for studying gut mucosal immunity.
Schmucker, D. L., Kato, T., Owen, R. L., Anderson, J., Miller, C., &
Hayworth, M. F. (Cell Biol. & Aging [151E], Dept Vet. Affiars Med Ctr, 4150
Clement St, San Francisco, CA 94121). Regional Immunology, 1995,
6, 238-242.
. .
Cecal fistulae were well tolerated by two adult female rhesus in studies of
secretory antibody response to direct immunization of the gut mucosa. This
model may facilitate in vivo studies of factors that modulate antigen
uptake and regulate intestinal immune responses.
Opioid and nonopioid analgesic drug effects on colon contractions in monkeys.
Ferraz, A. A. B., Cowles, V. E., Condon, R. E., & Schulte, W. J. (R. E. C.,
Dept of Surgery (Rm 3510), Med. Coll. of Wisconsin, 9200 W. Wisconsin Ave,
Milwaukee, WI 53226). Digestive Diseases and Sciences, 1995, 40,
1417-1419.
. .
Recordings of colon contractions of four cynomolgus macaques after colon
surgery showed that morphine increased the frequency of phasic and decreased
the frequency of migrating contractions in the colon. Ketorolac does not
affect the frequency of colon contractions.
Effects of morphine and ketorolac on thermal allodynia induced by
prostaglandin E2 and bradykinin in rhesus monkeys. Negus, S. S., Butelman, E.
R., Gatch, M. B., & Woods, J. H. (ADA Research Center, McLean Hospital, 115
Mill St, Belmont, MA 02178-9106). Journal of Pharmacology and Experimental
Therapeutics, 1995, 274, 805-814.
. .
Relative to morphine, ketorolac had a similar efficacy and slightly lower
potency in blocking BK-induced allodynia. Ketorolac also displayed a slower
onset but a longer duration of action than morphine.
Osteoarthritis in cynomolgus macaques II. Detection of modulated proteoglycan
epitopes in cartilage and synovial fluid. Carlson, C. S., Loeser, R. F.,
Johnstone, B., Tulli, H. M., Dobson, D. B., & Caterson, B. (Dept of Comp.
Med., Bowman Gray Sch. of Med. at Wake Forest Univ., Med. Center Blvd,
Winston-Salem, NC 27157-1040). Journal of Orthopaedic Research, 1995,
13, 399-409.
. .
Measurement of the epitope recognized by 7-D-4 in synovian fluid or, by
immunohistochemical or Western blot methods, in articular cartilage has
potential use as a marker of severity of naturally occurring osteoarthritis.
Animal models for Lyme disease. Barthold, S. W. (Sect. of Comp. Med., Yale
Univ. Sch. of Med., P.O. Box 208016, 333 Cedar St, New Haven, CT 06520-8016).
Laboratory Investigation, 1995, 72, 127-129.
. .
The rhesus model offers some clear potential for investigating mechanisms of
neuroborreliosis, which cannot be obtained with other models or human clinical
material.
Primate models of cholinergic dysfunction. Liberini, P. & Cuello, A. C.
(Dept of Biomed. Sci. & Biotech., Div. of Pharmacol. & Exp. Therapeut.,
Brescia Univ. School of Med., Via Valsabbina 19, 25124 Brescia, Italy).
Functional Neurology, 1995, 10, 45-54.
. .
Mature basal forebrain cholinergic neurons degenerate as a consequence of the
removal of their target. Both fimbria fornix transection and cortical
devascularization seem to be useful tools in the assessment of potential
neurotrophic and neuroprotective properties of pharmacological agents.
A macaque model for hantavirus infection. Groen, J., Gerding, M., Koeman, J.
P., Roholl, P. J. M., van Amerongen, G., Jordans, H. G. M., Niesters, H. G. M.,
& Osterhaus, A. D. M. E. (A. D. M. E. O., Inst of Virology, Erasmus Univ.,
P.O. Box 1738, 3000 DR Rotterdam, Netherlands). Journal of Infectious
Diseases, 1995, 172, 38-44.
. .
The experimental infection of cynomolgus macaques with Puumala virus, which
causes nephropathia epidemica (NE) in humans, results in a mild form of acute
renal infection similar to mild forms of NE in humans.
Simian varicella virus antibody response in experimental infection of African
green monkeys. Gray, W. L., Gusick, N. J., Fletcher, T. M., & Soike, K. F.
(Dept of Microbiol. & Immunol., Univ. of Arkansas for Med. Sci., Little
Rock, AR 72205). Journal of Medical Primatology, 1995, 24,
246-251.
. .
Results demonstrate that the simian varicella model offers a useful approach
for investigating immune mechanisms in human varicella zoster virus
infections.
Ischemic neuronal damage specific to monkey hippocampus: Histological
investigation. Tabuchi, E., Ono, T., Nishijo, H., Endo, S., & Kuze, S. (T.
O., Dept of Physiology, Fac. of Med., Toyama Med. & Pharm. Univ., Sugitani,
Toyama 930-01, Japan). Brain Research Bulletin, 1995, 37,
73-87.
. .
The CA1 subfield of the monkey (M. fuscata) hippocampus is the precise
region of the brain most susceptible to ischemic insult in the primate
forebrain, and after a critical time (15-min ischemia in the procedure used)
ischemic cell changes occur suddenly and extensively. Ischemia due to
occlusion of 8 arteries for 10-15 min could produce a model of human amnesia
caused by transient ischemic insult.
Physiological results of monkey brain ischemia, and protection by a calcium
blocker. Nishijo, H., Tabuchi, E., Ono, T., Fukuda, M., Kuze, S., & Endo,
S. (T. O., Address same as above). Brain Research Bulletin, 1995,
37, 89-101.
. .
A calcium entry blocker can protect neurons from mild ischemia-induced injury
and might ameliorate morphological damage and functional impairment of the
brain due to ischemia in patients who suffer transient anoxic or hypoxic
injury.
Effect of transcranial magnetic stimulation on cerebral function in a monkey
model. Yamada, H., Tamaki, T., Wakano, K., Mikami, A., & Transfeldt, E. E.
(Dept of Orthopedic Surgery, Wakayama Med. College, Wakayama National College
of Technology, Wakayama 640, Japan). Electroencephalography and Clinical
Neurophysiology, 1995, 97, 140-144.
. .
Transcranial magnetic stimulation had no effect on monkeys performing a task
requiring spatial short-term memory.
Does experimentally-induced amblyopia cause hyperopia in monkeys? Kiorpes, L.
& Wallman, J. (Center for Neural Sci., 4 Washington Pl., Rm 809, NYU, New
York, NY 10003). Vision Research, 1995, 35, 1289-1297.
. .
Refractive errors were assessed in 19 M. nemestrina raised with
experimentally produced strabismus or unilateral defocus. Refractive changes
were correlated with changes in axial length and vitreous chamber depth,
suggesting that amblyopia may cause alterations in eye growth and late-onset
hyperopia.
Long-term effects of taurine deprivation on retinal morphology in rhesus
monkeys. Imaki, H., Neuringer, M., & Sturman, J. A. (Dept of
Developmental Biochem., NY State OMRDD, Inst. for Basic Research in
Developmental Disabilities, Staten Island, NY 10314). Biomedical Letters,
1994, 50, 211-226.
. .
Taurine-deprived monkeys showed retinal changes, persisting long after
differences in taurine concentrations had disappeared.
Cold stress-induced recurrences of herpetic keratitis in the squirrel monkey.
Varnell, E. D., Kaufman, H. E., Hill, J. M., & Thompson, H. W. (LSU Eye
Center, 2020 Gravier St, Suite B, New Orleans, LA 70112). Investigative
Ophthalmology & Visual Science, 1995, 36, 1181-1183.
. .
Squirrel monkeys infected by the ocular route with herpesvirus type 1 were
exposed to temperatures about 5deg.C lower than normal for periods as short as
12 hours, producing significant recurrences of keratitis.
Pharmacokinetics of antisense oligonucleotides. Agrawal, S., Temsamani, J.,
Galbraith, W., & Tang, J. (Hybridon Inc., 1 Innovation Dr., Worcester, MA
01605). Clinical Pharmacokinetics, 1995, 28, 7-16.
. .
A positive review of the pharmacokinetics and in vivo stability of
S-oligonucleotide in mice, rats, and monkeys.
Neurotoxicity of glucocorticoids in the primate brain. Uno, H., Eisele, S.,
Sakai, A., Shelton, S., Baker, E., DeJesus, O., & Holden, J. (Wisconsin
RPRC, Univ. of Wisconsin, Madison, WI 53706). Hormones and Behavior,
1994, 28, 336-348.
. .
Prenatal administration of a potent glucocorticoid induced an irreversible
deficiency of the hippocampal neurons, high plasma cortisol at the circadian
baseline, and post-stress levels in juvenile rhesus monkeys, suggesting that
the hippocampus mediates negative feedback of cortisol release.
Comparison of skeletal muscle laser Doppler flowmetry to changes in central
hemodynamics in detecting the physiological response to moderate hemorrhage.
Rodriguez, A. A., Gardner, G. P., LaMorte, W. W., Obi-Tabot, E. T., Valeri, C.
R., & Hirsch, E. F. (Div. of Surgery, Boston Univ. Sch. of Med., Boston,
MA 02118). Journal of Surgical Research, 1995, 58, 189-192.
. .
Laser Doppler (LD) flow and hemodynamic parameters were measured following two
episodes of bleeding and reinfusing 250 cc of blood. Results suggest that LD
flowmetry is more sensitive to tissue hypoperfusion than commonly measured
central parameters, and may be useful in aiding the resuscitation of critically
ill patients.
Effect of recombinant human osteogenic protein-1 on healing of segmental
defects in non-human primates. Cook, S. D., Wolfe, M. W., Salkeld, S. L., &
Rueger, D. C. (Dept of Orthop. Surg., Tulane Univ. Sch. of Med., 1430 Tulane
Ave, New Orleans, LA 70112-2699). Journal of Bone and Joint Surgery,
1995, 77-A, 734-750.
. .
Recombinant human osteogenic protein-1 implants elicited healing in large
segmental bone defects that was as good as or better than that achieved with
autogenous bone grafts.
Effect of ductal patency on organ blood flow and pulmonary function in the
preterm baboon with hyaline membrane disease. Morrow, W. R., Taylor, A. F.,
Kinsella, J. P., Lally, K. P., Gerstmann, D. R., & deLemos, R. A.
(Pediatric Cardiology, Children's Hosp. of Michigan, 3901 Beaubien Blvd,
Detroit, MI 48201). Critical Care Medicine, 1995, 23, 179-186.
. .
Early ductal ligation did not result in improved cardiac output, increased
organ blood flow, or improved pulmonary function. Gradual constriction of the
ductus arteriosus may play an important role in succsesful cardiovascular
adaptation in the premature infant.
Intrastriatal cografts of autologous adrenal medulla and sural nerve in
MPTP-induced parkinsonian macaques: Behavioral and anatomical assessment.
Watts, R. L., Mandir, A. S., & Bakay, R. A. E. (Dept of Neurol., Emory
Univ. Sch. of Med., 6000 WMB, P.O. Drawer V, Atlanta, GA 30322). Cell
Transplantation, 1995, 4, 27-38.
. .
Animals undergoing autologous cografts demonstrated improved motor performance
while a control animal continued in a chronic, stable parkinsonian state.
Behavior
Long-tailed macaques avoid conflicts during short-term crowding.
Aureli, F., van Panthaleon van Eck, C. J., & Veenema, H. C. (Yerkes RPRC,
Emory Univ., Atlanta, GA 30322). Aggressive Behavior, 1995, 21,
113-122.
. .
Data from 42 individual M. fascicularis in different age-sex classes
showed that only mild forms of aggression increased under short-term crowded
conditions. Crowding resulted in a decrease in counteraggression,
allogrooming, playing, and exploring, along with an increase in huddling with
companions. The animals reduce the risk of severe aggression when
interindividual distances are small by simply decreasing the level of activity.
When aggression does occur during crowding, tension reduction mechanisms such
as reconciliation and redirection are used as often as in control conditions.
Temperament in nonhuman primates. Clarke, A. S. & Boinski, S. (Harlow
Primate Lab., Univ. of Wisconsin, 22 N. Charter St, Madison, WI 53711).
American Journal of Primatology, 1995, 37, 103-125.
. .
Studies of temperament in nonhuman primates have provided a useful framework
for characterizing consistent variations among primates in behavioral and
physiological responsivity at both the individual and species levels. The
temperamental dimension that has been most useful is that of bold/shy or
fearful/unfearful in response to novelty. Characteristics desplayed by
individuals appear to have a constitutional basis, yet are to some extent
modifiable by experience. Tentative evidence suggests that bold primate
species may utilize more active and instrumental foraging strategies than less
bold species.
Exploratory behavior and learning in laboratory marmosets (Callithrix
jacchus jacchus): Comparisons between experimental-cage and home-cage
activity. Forster, F. C. (Dept of Psychology, Lancaster Univ., Lancaster LA1
4YF, England). Primates, 1995, 36, 501-514.
. .
In an isolated experimental cage, marmosets which learned a lever-pressing
task showed higher general activity levels than controls; in the group home
cages, the mere addition of novel objects resulted in enhancement of
exploratory behavior. Although the activities associated with learning tended
to decline over time, the novel objects remained interesting throughout the
experiment.
Care
Normal and abnormal behaviors of laboratory animals: What do they mean?
Bayne, K. (11300 Rockville Pike, Suite 1211, Rockville, MD 20852-3035). Lab
Animal, 1996, 25[1], 21-23.
. .
"Knowledge of an animal's behavior will help care staff judge when an animal's
actions are appropriate or inappropriate. By furthering our understanding of
normal behavior, we can provide better husbandry to our research animals, and
make a more refined animal model available to the investigator."
Pigtail macaque performance on a challenging joystick task has important
implications for enrichment and anxiety within a captive environment. Lincoln,
H., III, Andrews, M. W., & Rosenblum, L. A. (SUNY Health Sci. Center, Dept
of Psychiatry, Box 120, Brooklyn, NY 11203). Laboratory Animal Science,
1995, 45, 264-268.
. .
The number of daily trials on a joystick task decreased when the task was made
more difficult, with more time required to complete each trial with the
difficult task. Preferences among locations became more pronounced with the
more difficult task. Preferences apparently derived, at least in part, from
levels of induced anxiety.
Preferential and motivational testing to improve animal well-being. Fraser, D.
(Centre for Food & Animal Res., Bldg 94, Agriculture & Agri-Food
Canada, Ottawa K1A 0C6, Canada). Lab Animal, 1996, 25[1],
27-31.
. .
"Can we, in fact, assume that what an animal chooses is really the option that
best promotes its welfare?"
Refining the blood collection procedure for macaques. Reinhardt, V. (4605
Crescent Rd, Madison, WI 53711). Lab Animal, 1996, 25[1],
32-35.
. .
Training nonhuman primates to cooperate during venipuncture in their home
environment can eliminate significant cortisol responses.
Simian bites and bacterial infection. Goldstein, E. J. C., Pryor, E. P., III,
& Citron, D. M. (2021 Santa Monica Blvd, Suite 640E, Santa Monica, CA
90404). Clinical Infectious Diseases, 1995, 20, 1551-1552.
. .
Three patients with simian bites and resultant infection are described. A
literature review revealed brief mentions of 132 cases of simian bites.
Infection, despite antimicrobial therapy, and complications, such as
osteomyelitis and flexion contractures, occurred frequently.
Dermatitis in a rhesus macaque after autologous bone marrow transplantation.
What's your diagnosis? Feldman, S. H., Metzger, M., & Hoyt, R. F., Jr.
(Facility Vet., Research Court Fac., Lab. of Animal Med. & Surg., NHLBI,
NIH, 5 Research Ct, Rm 1B18, Rockville, MD 20850). Lab Animal,1995,
24[3], 17-19.
. .
Staphlococcus aureus, present in 5% of some healthy populations, in
this case caused skin lesions in a rhesus three months post-transplantation.
Development
Comparative analysis of weight gain, hand/wrist maturation, and dental
emergence rates in chimpanzees aged 0-24 months from varying captive
environments. Marzke, M. W., Young, D. L., Hawkey, D. E., Su, S. M., Fritz,
J., & Alford, P. L. (Dept of Anthropology, Arizona State Univ., Box 872402,
Tempe, AZ 85287-2402). American Journal of Physical Anthropology, 1996,
99, 175-190.
. .
Results of a mixed longitudinal study of 175 animals are suggestive of
significant sex and environmental effects on the variables monitored,
justifying further analysis and continuation of the study. The study provides
norms specific to sex, rearing, and colony environments with which individual
colony animals may be compared, and also provides a standard with which the
dental emergence status and hand/wrist maturation of fossil apes and hominids
may be compared.
Effects of age, sex, and heredity on measures of bone mass in baboons
(Papio hamadryas). Kammerer, C. M., Sparks, M. L., & Rogers, J.
(Southwest Fnd. for Biomed. Research, P.O. Box 28147, San Antonio, TX
78228-0147). Journal of Medical Primatology, 1995, 24,
236-242.
. .
Bone mass and bone density were estimated in 219 pedigreed baboons by
radiographic morphometry.
Neuronal number and size are preserved in the nucleus basalis of aged rhesus
monkeys. Voytko, M. L., Sukhov, R. R., Walker, L. C., Breckler, S. J., Price,
D. L., & Koliatsos, V. E. (Dept of Comp. Med., Bowman Gray Sch. of Med.,
Wake Forest Univ., Med. Center Blvd, Winston-Salem, NC 27157-1040).
Dementia, 1995, 6, 131-141.
. .
Neurons in the nucleus basalis of Meynert (NBM) were analyzed morphometrically
in 21 rhesus monkeys ranging in age from 9 to 33 years. Numbers of NBM
cholinergic neurons are stable with age; NBM neurons become hypertrophic in
older animals; morphometric indices of cholinergic neurons are associated with
cognitive function.
Effects of early rearing environment on immune responses of infant
rhesus monkeys. Lubach, G. R., Coe, C. L., & Ershler, W. B. (Wisconsin
RPRC, Univ. of Wisconsin, Madison, WI 53706). Brain, Behavior, and
Immunity, 1995, 9, 31-46.
. .
Nursery-reared monkeys had significantly lower proportions of CD8 cells and
lower natural killer cell activity than did mother-raised monkeys. Their
lymphocyte proliferation responses to mitogen stimulation were higher than
those of mother-raised monkeys. Abnormal early rearing may have long-lasting
effects on the immune system, which could have health consequences later in
life.
Developmental changes in brain metabolism in sedated rhesus macaques and
vervet monkeys revealed by positron emission tomography. Jacobs, B., Chugani,
H. T., Allada, V., Chen, S., Phelps, M. E., Pollack, D. B., & Raleigh, M.
J. (Lab. of Quant. Neuromorphology, Dept of Psychology, Colorado College, 14 E.
Cache La Poudre, Colorado Springs, CO 80903-3298). Cerebral Cortex,
1995, 3, 222-233.
. .
The period between 2 and 6 months of age is a time of peak metabolic activity,
which coincides with increased histologic and behavioral activity in the
developing monkey.
Disease
Emerging viruses. Le Guenno, B. (Inst. Pasteur, 25 et 28, rue du
Docteur-Roux, 75724 Paris Cédex 15, France). Scientific American,
October, 1995, 44, 30-37.
. .
An overview of the recent epidemics of hemorrhagic fevers from newly
discovered viruses.
Transmission of Ebola virus (Zaire strain) to uninfected control monkeys in a
biocontainment laboratory. Jaax, N., Jahrling, P., Geisbert, T., Geisbert, J.,
Steele, K., McKee, K., Nagley, D., Johnson, E., Jaax, G., & Peters, C.
(USAMRIID, Frederick, MD 21702-5011). Lancet, 1995, 346,
1669-1671.
. .
Transmission of Ebola virus to two of three control rhesus monkeys that did
not have direct contact with experimentally inoculated monkeys held in the same
room is reported. The most likely route of infection was aerosol, oral, or
conjunctival exposure to virus-laden droplets secreted or excreted from the
inoculated monkeys.
Protozoal diseases of nonhuman primates. Hubbard, G. B. (Southwest Fnd. for
Biomed. Research, P.O. Box 28147, San Antonio, TX 78228-0147). Seminars in
Avian and Exotic Pet Medicine, 1995, 4, 145-149.
. .
Generally, protozoan infections are a minor problem in nonhuman primates
unless the animals are stressed. Most protozoa are opportunistic and can be
found as natural nonpathogenic inhabitants of their nonhuman primate hosts.
The simian herpesviruses. Eberle, R. & Hilliard, J. (J. H., address same
as above). Infectious Agents and Disease, 1995, 4, 55-70.
. .
This review "summarizes what is known about the natural history and pathogenic
potential of simian alphaherpesviruses. Recent research into the molecular
biology of this group of viruses is also reviewed, and recent advances toward
development of diagnostic tests based on these data are discussed."
Mycobacterial infections of animals: Pathology and pathogenesis. Hines, M.
E., II, Kreeger, J. M., & Herron, A. J. (Veterinary Diagnostic &
Investig. Lab., Univ. of Georgia, Coll. of Vet. Med., Tifton, GA 31793).
Laboratory Animal Science, 1995, 45, 334-351.
. .
An overview of mycobacterial disease in various animal species, with special
emphasis on the mycobacteria that cause disease, the lesions they produce, and
the pathogenesis of resulting disease.
Mycobacterium kansasii infection in squirrel monkeys (Saimiri
sciureus sciureus). Brammer, D. W., O'Rourke,
C. M., Heath, L. A., Chrisp, C. E., Peter, G. K., & Hofing, G. L.
(Parke-Davis Pharm. Res., 2800 Plymouth Rd, Ann Arbor, MI 48105). Journal
of Medical Primatology, 1995, 24, 231-235.
. .
Documentation of asymptomatic infections of M. kansasii in four of five
tuberculin-positive squirrel monkeys.
The new GB hepatitis viruses. Zuckerman, A. J. (WHO Collaborating
Centre for Reference & Research on Viral Diseases, Royal Free Hospital
School of Med., London, UK). The Lancet, 1995, 345, 1453-1454.
. .
Preliminary results on the molecular cloning and genomic characterization of
the GB agent are sketched.
Identification of two flavivirus-like genomes in the GB hepatitis agent.
Simons, J. N., Pilot-Matias, T. J., Leary, T. P., Dawson, G. J., Desai, S. M.,
Schlauder, G. G., Muerhoff, A. S., Erker, J. C., Buijk, S. L., Chalmers, M. L.,
Van Sant, C. L., & Mushahwar, I. K. (Virus Discovery Group, Exp. Biology
Research, Abbott Labs, North Chicago, IL 60064). Proceedings of the
National Academy of Sciences of the U.S.A., 1995, 92,
3401-3405.
. .
The known viral agents of hepatitis do not account for all of the cases of
hepatitis of purported viral etiology. Diagnostic reagents developed from the
genomic sequences of GBV-A and GBV-B may determine whether either of these
viruses are the causative agents in hepatitis cases of unknown etiology.
Ultrastructural and histochemical mesangial alterations in callitrichid IgM
nephropathy (Primates: Platyrrhina). Brack, M. & Weber, M. (DPZ,
Kellnerweg 4, D-37077 Göttingen, Germany). Nephron, 1995,
69, 286-292.
. .
Alterations during different stages of spontaneously occurring IgM nephropathy
in Callitrichidae were determined.
Evolution, Genetics, and Taxonomy
Owl monkeys (Aotus) are highly divergent in mitochondrial
cytochrome c oxidase (COII) sequences. Ashley, M. V. & Vaughn, J.
L. (Dept of Biol. Sciences, Univ. of Illinois, 3262 SES, 845 W. Taylor St,
Chicago, IL 60607-7060). International Journal of Primatology, 1995,
16, 793-806.
. .
Aotus COII data support the existence of multiple species of
Aotus, with origins predating late Pleistocene climatic events.
Although A. nancymae and A. azarae both live south of the Amazon
River and have been considered members of the same species group, phylogenetic
analysis of the COII sequences does not support a close relationship between
them.
General
A Whistle Blower's Bitter Battle. P. V. Walker. Chronicle of Higher
Education, March 15, 1996, A34.
. .
A discussion of the legal battle between J. Moor-Jankowski and New York
University administration over the disposition of LEMSIP.
Instruments & Techniques
Comparison of fluorescein angiography with microvascular anatomy of
macaque retinas. Weinhaus, R. S., Burke, J. M., Delori, F. C., &
Snodderly, D. M. (Schepens Eye Research Inst., 20 Staniford St, Boston, MA
02114). Experimental Eye Research, 1995, 61, 1-16.
. .
A comparison of high-quality angiograms with complete maps of capillary
networks determined histologically.
Salivary cortisol in nursery-reared rhesus monkeys: Reactivity to peer
interactions and altered circadian activity. Boyce, W. T., Champoux, M.,
Suomi, S. J., & Gunnar, M. R. (M. R. G., Inst. of Child Development, 51 E.
River Rd, Univ. of Minnesota, Minneapolis, MN 55455). Developmental
Psychobiology, 1995, 28, 257-267.
. .
Saliva cortisol samples were obtained by giving each animal, while in its
individual home cage, a 6-in. cotton dental roll pretreated with sugared
fruit-drink crystals and dried. Subjects were allowed to mouth the cotton roll
for 10 min. during each collection period. Saliva was then expressed from the
cotton and analyzed by radioimmune assay. Subjects had higher average cortisol
concentrations when the sampling followed peer-socialization sessions, compared
to no-socialization days.
Fecal steroid analysis of ovarian cycles in free-ranging baboons. Stavisky,
R., Russell, E., Stallings, J., Smith, E. O., Worthman, C., & Whitten, P.
L. (Lab. of Reproductive Biology, Yerkes RPRC, Emory Univ., Atlanta, GA 30329).
American Journal of Primatology, 1995, 36, 285-297.
. .
Field and laboratory tests of serial sampling, solid phase extraction, and
microradioimmunoassay methods for the collection, preservation, and analysis of
fecal steroids. Estradiol and progesterone concentrations in the
field-extracted samples exhibited high recovery and statistically significant
correlations (P < 0.05) with concentrations in the lab-extracted
samples, suggesting that solid phase extraction could provide a useful
alternative to freezing.
Polymerase chain reaction and a liquid-phase, nonisotopic hybridization for
species-specific and sensitive detection of malaria infection. Oliveira, D.
A., Holloway, B. P., Durigon, E. L., Collins, W. E., & Lal, A. A.
(Immunology Br., Div. of Parasitic Diseases, NCID, CDC, 4770 Buford Hwy,
Mailstop F12, Chamblee, GA 30341). American Journal of Tropical Medicine and
Hygiene, 1995, 52, 139-144.
. .
This assay is sensitive, specific, simple, and reliable, detecting mixed
infections and low-level parasitemia.
Nutrition
Differences in cholesterol metabolism in juvenile baboons are
programmed by breast- versus formula-feeding. Mott, G. E., Jackson, E. M.,
DeLallo, L., Lewis, D. S., & McMahan, C. A. (Dept of Pathology, Univ. of
Texas Health Sci. Ctr, San Antonio, TX 78284-7750). Journal of Lipid
Research, 1995, 36, 299-307.
. .
Results suggest that breast- vs formula-feeding in baboons imprints
differences in bile acid synthesis, regulation of LDL receptor expression, and
HDL-C subfraction concentrations.
Pharmacology & Anesthesia
The effects of multiple administrations of sevoflurane to cynomolgus monkeys:
Clinical pathologic, hematologic, and pathologic study. Soma, L. R., Tierney,
W. J., Hogan, G. K., & Satoh, N. (Univ. of Penn. School of Vet. Med., NBC
Campus, Kennett Square, PA 19348-1692). Anesthesia and Analgesia, 1995,
81, 347-352.
. .
Sevoflurane adminstered at 1.0 and 1.6 times the minimum alveolar anesthetic
concentration for 72 hours over an 8-week period was well tolerated, except for
transient early increases in serum enzymes.
Reproduction
1994 International Studbook: Golden Lion Tamarin: (Leontopithecus
rosalia rosalia). J. D. Ballou & A. Sherr, Studbook Keepers. 259 pp. (Dept
of Zool. Research, NZP, Smithsonian Inst., Washington, DC 20008).
. .
Includes special management notes, a section on diseases, and a reprint
of "Diseases and their control for reintroduction of the endangered golden lion
tamarin (Leontopithecus spp.)," by R. J. Montali, M. Bush, J. Hess, J.
D. Ballou, D. G. Kleiman, & B. B. Beck.
The primate endometrium: Morphological and secretory changes during early
pregnancy -- implications of the insulin-like growth factor axis. Fazleabas,
A. T., Hild-Petito, S., & Verhage, H. G. (Univ. of Illinois, Dept of
OB-GYN, 820 S. Wood St (M/C 808), Chicago, IL 60612-7313). Seminars in
Reproductive Endocrinology, 1995, 13, 120-132.
. .
This review compares and contrasts the morphological and biochemical changes
associated with implantation and early pregnancy in the human, baboon, and
rhesus macaque.
Are daughters more costly to produce for Japanese macaque mothers? Sex of the
offspring and subsequent interbirth intervals. Takahata, Y., Koyama, N.,
Huffman, M. A., Norikoshi, K., & Suzuki, H. (Naruto Univ. of Education,
Naruto, Tokushima 772, Japan). Primates, 1995, 36, 571-574.
. .
A tendency for longer interbirth intervals following the birth of daughters
than of sons was seen in a sample of 237 live births recorded over 30 years.
Behavioral changes across the menstrual cycle in isosexual groups of bonnet
macaques (Macaca radiata). Bassoff, T. B. (20 Plaza St [F1], Brooklyn,
NY 11238-4929). Primates, 1995, 36, 549-562.
. .
Behavioral changes were recorded over a six-month period for two lab-housed
triads of bonnet macaques. An increase in contact aggressive hehaviors during
the early-mid follicular phase and the mid-late luteal phase does not suggest a
simple hormonal correlate, as these two phases are characterized by high levels
of estrogen and progesterone, respectively. However, the distribution of
nonaggressive hierarchical behaviors suggest that this category of agonistic
interaction may be related to mating competition.
Effects of chronic melengestrol acetate contraceptive treatment on perineal
tumescence, body weight, and sociosexual behavior of hamadryas baboons
(Papio hamadryas). Portugal, M. M. & Asa, C. S. (C. S. A., St.
Louis Zoo, Forest Park, St. Louis, MO 63110). Zoo Biology, 1995, 14,
251-259.
. .
Treatment with melengestrol acetate can be recommended based on its ability to
suppress ovarian cyclicity without causing social disruption. However, because
this and other progestins stimulate weight gain, their use is contraindicated
in overweight or obese animals.
Xenotransplantation
Successful survival of primates receiving transplantation with "dead,"
nonbeating donor hearts. Gundry, S. R., Fukushima, N., Eke, C. C., Hill, A.
C., Zuppan, C., & Bailey, L. L. (Div. of Cardiothoracic Surgery, Dept of
Surgery, Loma Linda Univ. Med. Center, 11234 Anderson St, Loma Linda, CA
92354). Journal of Thoracic and Cardiovascular Surgery, 1995, 109,
1097-1102.
. .
Baboon hearts recovered 15 to 31 minutes after cardiac standstill caused by
anoxia can be transplanted, reperfused, and reanimated without difficulty with
the use of clinically applicable methods. The use of donors with nonbeating
hearts could be extended to human cardiac transplantation, greatly expanding
the donor pool.
Secondary organ allografting after a primary "bridging" xenotransplant. Ye,
Y., Luo, Y., Kobayashi, T., Tani-guchi, S., Li, S., Niekrasz, M., Kosanke, S.,
Baker, J., Mieles, L., Smith, D., & Cooper, D. K. C. (Oklahoma
Transplantation Inst., Baptist Med. Center, 3300 NW Expressway, Oklahoma City,
OK 73112). Transplantation, 1995, 60, 19-22.
. .
In four baboons that had undergone previous xenotransplants, subsequent organ
allografting was not followed by hyperacute, antibody-mediated, or accelerated
cellular rejection. Organ xenotransplantation using discordant or concordant
donor species does not prohibit subsequent allotransplantation.
Red cell compatibility testing in baboon xenotransplantation. Triulzi, D. J.
& Jochum, E. E. (Dept of Pathology, Univ. of Pittsburgh Med. Center, 3636
Blvd Allies, Pittsburgh, PA 15213). Transfusion, 1995, 35,
756-759.
. .
Description of a simple technique to determine the ABO group in baboons, to
aid in the selection of baboon donors for hepatic xenotransplantation.
Heteroagglutinins and their significance in baboon hepatic
xenotransplantation. Triulzi, D. J., Jochum, E. A., Marino, I. R., &
Starzl, T. E. (Address same as above). Transplantation, 1995, 60,
127-131.
. .
Based on data from two human patients who received baboon livers, human
antibaboon heteroagglutinins do not appear to be associated with hyperacute or
cellular rejection, but their role in the complement-mediated injury, suspected
in both cases, cannot definitively be excluded.
Virus infection of baboons. Plagemann, P. G. W. (Dept of Microbiology, Univ.
of Minnesota, Minneapolis, MN 55455). Nature, 1995, 377, 98.
. .
Simian hemmorrhagic fever, which is endemic in some populations of Papio
anubis and P. cyanocephalus, could be transferred to humans by
xenotransplantation.
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; FAX: 401-863-1300]
Judith_Schrier@brown.edu
Current and back issues of the Newsletter are available on the
World Wide Web at
http://www.brown.edu/Research/Primate
The Newsletter is supported by U. S. Public Health Service Grant RR-00419 from the Comparative Medicine Program, National Center for Research Resources, N.I.H.
Cover illustration of a squirrel monkey (Saimiri sciureus sciureus) by Susan D. Meier (see Natural and Artificial Minds, SUNY Press, 1993, with permission)
Copyright (c) 1996 by Brown University
Copy Editor: Elva Mathiesen
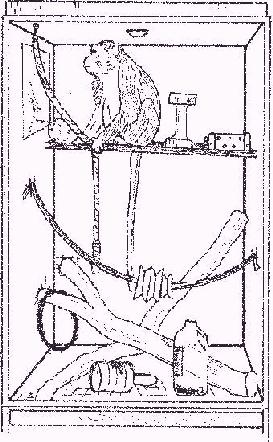{kind=link}
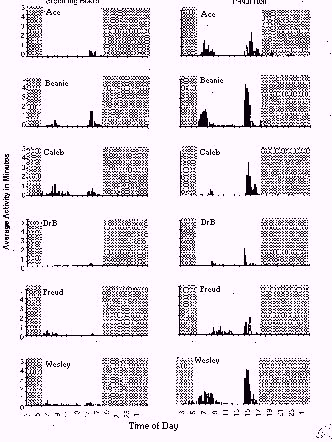{kind=link}
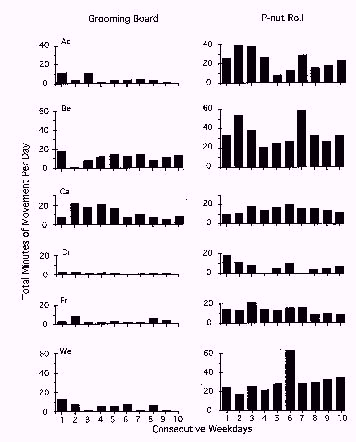{kind=link}
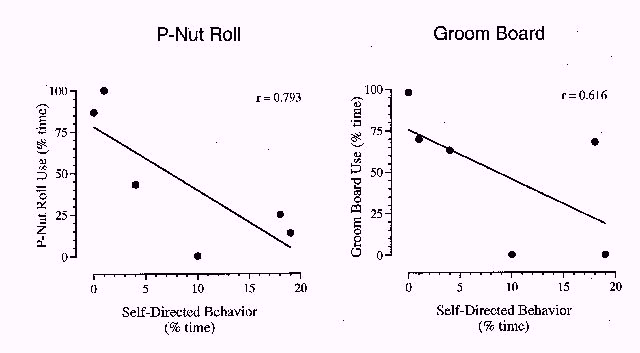{kind=link}
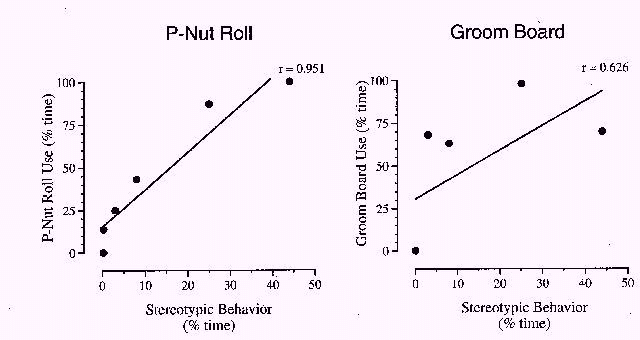{kind=link}