 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 40 NUMBER 1 JANUARY 2001
Behavioral Enrichment for Marmosets by a Novel Food Dispenser, by B. Voelkl, E. Huber, & E. Dungl......1
Puzzle Ball Foraging Device for Laboratory Monkeys, by C. M. Crockett, R. U. Bellanca, K. S. Heffernan, D. A. Ronan, & W. F. Bonn......4
Who’s Enriching Whom? The Mutual Benefits of Involving Community Seniors in a Research Facility’s Enrichment Program, by N. Megna & J. Ganas......8
Early vs. Natural Weaning in Captive Baboons: The Effect on Timing of Postpartum Estrus and Next Conception, by J. Wallis & B. Valentine......10
Discussion: Enrichment for Lemurs......14
News, Information, and Announcements
Meeting Announcements......3
Workshop Announcements......7
...Teaching Research Ethics; Orangutan Reintroduction and Protection
Call for Papers: AAZV......13
News Briefs......15
...Ape Caper Foiled; Death of a Chimpanzee at Coulston; Anna Mae Noell, Owned “Chimp Farm”; LEMSIP Chimps Retire to Texas; Three New Lemur Species; Clinton Signs Great Ape Conservation Bill Into Law
Letters: Howling Howlers......16
Awards Granted......17
...PCWS Conservation Grant; NCRR Primate Grants
Award Nominations: Fyssen Foundation......17
Research and Educational Opportunities......20
...Continuing Education for Lab Animal Technicians; Primate Behavior and Ecology Program - Panama; Field Course in Animal Behavior - Georgia and Africa; Research in the Biology of Aging; Summer Apprenticeship: Chimpanzees and ASL
Information Requested or Available......22
...International Veterinary Information Service; New Video Available; Introducing PIN WEB LINKS; E-mail News Groups: ZooNews Digest and Zoo Biology; More Interesting Web Sites
Grants Available......23
...ACLAM Foundation Request For Proposals; National Research Service Awards for Senior Fellows; Behavioral Science Award For Rapid Transition
Resources Wanted and Available......24
...Species Information Service Launched; Material Exchanges; Primate-Enrichment.Net; Free Software for Studying Behavior
Announcements from Publications......36
...International Journal of Comparative Psychology; Institute for Laboratory Animal Research; APE Boletín on the Web
Departments
Address Change......7
Primates de las Américas...La Página......18
Positions Available......19
...Clinical Lab Animal Veterinarian/Assistant Director; Primatology/Psychology - Bucknell University; Animal Behavior/Biology - Bucknell University; Gorilla Keeper - Georgia; Biological Anthropologist - Florida; Life Sciences Job Resource
Recent Books and Articles......25
* * *
Behavioral Enrichment for Marmosets by a Novel Food Dispenser
Bernhard Voelkl, Edith Huber, and Eveline Dungl
Konrad Lorenz Institute for Evolution and Cognition Research
Introduction
Environmental enrichment for captive primates is unquestionably an important part of managing research facilities and zoos. This is recognized by the U.S. government, which emphasized that "...in addition to providing the required standards of veterinary care and husbandry, regulated research facilities must...promote the psychological well-being of primates used in laboratories" (APHIS, 1995). To meet these demands, captive environments should encourage the expression of social and ecological skills. These should include foraging - searching for food - and food processing, both manual and oral.
In the wild, marmosets spend 50 to 60% of the day foraging (Rylands & deFaria, 1993), while in captivity feeding takes just a few minutes if the food is offered on an open plate. Animals in the wild have to spend more time feeding than in captivity, partly because some food sources have to be processed first, but mainly because they have to search for food, while captives often get all their food offered at the same time and place every day. To increase the animals' foraging behavior, it is advisable to distribute food over the whole living area and/or over the day (Novak & Drewson, 1989; Snowdon & Savage, 1989; Buchanan-Smith, 1997). One way of distributing food over the day is to use the food dispenser presented here. This dispenser has been constructed for feeding marmosets live mealworms. Besides exudates and fruit, insects are an important constituent of the diet of marmosets (Rylands & deFaria, 1993) and tamarins (Garber, 1993). By providing insects, we not only enrich the animals' lives but also supply them with important proteins which are not available from plants.
The Food Dispenser
The dispenser consists of a cylindrical storage tank (14 cm in diameter) with a funnel-shaped bottom (inclination about 10°) and a tube (10 mm in diameter) connecting the funnel with the interior of the cage. The tank can be detached from the dispenser (Figure 1) for cleaning. The tank is filled with about 30 g of mealworms, which crawl over the opening of the funnel, lose their grip, and slide through the tube into the cage, becoming prey for the monkeys. This dispenser provides the monkeys with a relatively small number of mealworms (about 300), more or less evenly distributed over a few hours, through a simple mechanism (Figure 2).
Figure 1: The food dispenser consists of a cylindrical storage tank (A) set in a tray (B), and a tube (C) connecting the funnel of the tank with the interior of the cage (E). The inclination of the tube (a) is about 60°. The dispenser is fixed at the cage with two hooks (D).
Figure 2: Mean number of mealworms eaten per minute using the dispenser (dotted line, flattened, 10 trials), or offering the mealworms in a bowl (solid line, flattened, 3 trials). The graphs are flattened by xi'= 1/3xi+2/9xi-1+1/1xi-2+2/9xi+1+1/9xi+2.
Subjects
The subjects were a group of 11 common marmosets (Callithrix jacchus) kept at the Konrad Lorenz Institute for Evolution and Cognition Research, Altenberg, Austria. All animals were born in captivity. The group lived in an indoor cage (2 x 3.5 x 3 m) equipped with branches, ropes, and living plants. The animals were fed fruits, vegetables, monkey pellets, insects, and protein supplements. They were kept at a daytime temperature of 26-30°C, and a night temperature of 21-23°C. Humidity was about 70-80%. In summer daylight was the main source of lighting, while in winter daylight fluorescent tubes were used to maintain a 12:12 hour light:dark cycle.
Method
Some enrichment devices encourage subjects to modify their behavior or to learn new skills (e.g. Marsy & Boussekey, 1992; Reinhardt, 1994; Citrynell, 1998). In the case of the mealworm dispenser we did not expect qualitatively new behavior: as soon as the mealworm falls to the floor of the cage it just has to be picked up - not a very sophisticated task at all. Rather we expected a quantitative shift in locomotor activity: we hypothesized that the subjects would inspect the dispenser frequently, to see if another mealworm were available. Therefore we decided to use a relative measurement for locomotor activity to assess whether the food dispenser has an effect on the monkeys' behavior.
An area of 40 x 40 cm was marked out around the mealworm dispenser, which was attached to the front of the cage. Over 10 days, we measured how often the monkeys entered that area during one hour when no mealworms were available (Control Phase) and when they were available (Test Phase). An increase in the monkeys' activity in front of the food dispenser during the Test Phase would not necessarily mean that their locomotor activity had increased. Such an increase could be caused purely by a local activity shift. It could even be that the animals were moving less, because they were waiting for the mealworms near the dispenser. To check for such an effect, another area of the same size was marked out at the rear of the cage. Only when the activity in the area around the dispenser rose while the activity in the rear area stayed constant, would we suggest that the locomotor activity of the subjects rose.
Results
Activity increased significantly in the area of the dispenser when mealworms were available (Wilcoxon Signed Rank Test, z = -2.803, p = 0.005). However, in the rear area of the cage, activity did not decrease (Wilcoxon Signed Rank Test, z = -1.376, p = 0.169). That means that this feeding device not only leads to a shift of the activity to the area of the feeding device but also increases the monkeys' total activity (Figure 3).
Figure 3: Number of times subjects entered the area around the dispenser (Dispenser) or the control area at the rear part of the cage (Rear), during 30 minutes when dispenser was filled with mealworms (Test) and during 30 minutes when no mealworms were available (Control). The bold line shows the median; the box the second and third interquartil; and the lines the range.
Due to the erratic movements of the mealworms, it is unpredictable when the next one will be provided - from just a few seconds up to several minutes. As it can take quite long until the next worm is delivered, and as the total period of time during which mealworms are dispensed amounts to several hours, the monkeys did not sit in front of the dispenser throughout the entire feeding period. At the beginning of the Test Phase the dominant female kept the other group members away from the dispenser, but she left after 10 to 20 minutes, leaving the other animals access to the dispenser. Thus the dispenser was not monopolized by a dominant individual for more than 10% of the total time.
Figure 4: A marmoset (Callithrix jacchus) in front of the dispenser, waiting for a mealworm.
Conclusion and Animal Welfare Implications
The dispenser offers several advantages over conventional feeding methods: (1) The food is distributed over a longer period of time. A food dispenser filled with 30 g of mealworms lasts about three hours, whereas the same amount of mealworms is consumed within four to six minutes when they are freely accessible. That means that the time spent foraging (= looking for food) increases. (2) The food is not permanently available but instead is available randomly. This unpredictability may raise the vigilance of the animals. (3) Maintenance is easy and does not require much additional time. The dispenser itself is attached to the outside mesh of the cage and can be refilled without opening the cage. (4) Even small amounts of food delivered by the dispenser have a strong effect on the behavior of the animals. This is important, because all enrichment activities related to food have to be incorporated into the feeding schedule; this is much easier when the amount of food needed for enrichment is low. (5) It is inexpensive. This is also important, because high additional costs are often used as an argument against behavioral enrichment. This rather simple apparatus can help to enrich the monkeys' foraging experience and thus reduce boredom and monotony. It can be used not only for marmosets and tamarins, but also for other insect-eating primates.
References
APHIS 1995 Factsheet: Regulatory enforcement and animal care. United States Department of Agriculture, Animal and Plant Health Inspection Service.
Buchanan-Smith, H. M. (1997). Environmental control: An important feature of good captive callitrichid environments. In C. Pryce, L. Scott, & C. Schnell (Eds.), Marmosets and tamarins in biological and biomedical research (pp. 47-53). Salisbury, UK: DSSD Imagery.
Citrynell, P. (1998). Cognitive enrichment: Problem solving abilities of captive white-bellied spider monkeys. Primate Eye, 66, 16-17.
Garber, P. A. (1993). Feeding ecology in the genus Saguinus. In A. B. Rylands (Ed.), Marmosets and tamarins: Systematics, behaviour, and ecology (pp. 273-295). Oxford: Oxford University Press.
Maier, W., Alonso, C., & Langguth, A. (1982). Field observations on Callithrix jacchus jacchus L. Zeitschrift für Säugetierkunde, 47, 334-346.
Marsy, P. & Boussekey, M. (1992). Environmental enrichment of the golden-bellied mangabey (Cercopithecus galeritus chrysogaster) groups in a zoological garden: Effects of puzzle feeder vs. food dispersion in litter. Proceedings of the XIVth Congress of the International Primatological Society, 307.
Novak, M. A. & Drewson, K. H. (1989). Enriching the lives of captive primates: Issues and problems. In E. F. Segal (Ed.), Housing, care, and psychological well-being of captive and laboratory primates (pp. 161-182). New Jersey: Noyes Publications.
Reinhardt, V. (1994). Caged rhesus macaques voluntarily work for ordinary food. Primates, 35, 95-98
Rylands, A. B. & de Faria, D. S. (1993). Habitats, feeding ecology, and home range size in the genus Callithrix. In A. B. Rylands (Ed.), Marmosets and tamarins: Systematics, behaviour, and ecology (pp. 262-271). Oxford: Oxford University Press.
Snowdon, C. T. & Savage, A. (1989). Psychological well-being of captive primates. In E. F. Segal (Ed.), Housing, care, and psychological well-being of captive and laboratory primates (pp. 161-182). New Jersey: Noyes Publications.
----------
First author's address: Institute of Zoology, University of Vienna, Althanstrasse 14, A-1090 Vienna, Austria [e-mail: [email protected]].
----------
The conference will focus on the management and technology needs of the laboratory animal science community, emphasizing information pertinent to the day-to-day management of laboratory animal facilities and career development for managers, directors, and administrators. The meeting is open to anyone involved in the field of laboratory animal science, including institutional administrators, members of Institutional Animal Care and Use Committees, laboratory animal veterinarians, investigators, researchers, regulatory personnel, managers, supervisors, and other staff who are responsible for laboratory animal care and use programs.
For additional information contact Carol Wigglesworth, Senior Policy Analyst, OLAW, Office of Extramural Research, NIH, RKL1, Suite 1050, MSC 7982, 6705 Rockledge Dr. [301-402-5913; e-mail: [email protected]]; or see <www.aalas.org/education/ meetings/MT_Conference/MT_Conf-index.htm>.
The Primate Society of Great Britain is having a one-day open meeting April 10, 2001, at the Bolton Institute, Bolton, Lancashire, England. This meeting is intended to reflect the diversity of current primatological research. Postgraduate research students particularly are encouraged to attend and give papers. For information or to offer a paper, contact Geoff Hosey, Biology and Environmental Studies, Deane Road, Bolton, BL3 5AB, UK [44 (0) 1204 903647; fax +44 (0) 1204 399074; e-mail [email protected]].
The American Society of Primatologists will hold the 2001 meeting in Savannah, Georgia. Armstrong Atlantic State University is sponsoring the event; our meeting will be the first to be held in the new science building that is currently under construction at Armstrong. The dates are Wednesday, August 8, 2001 (business meetings and the icebreaker), through Saturday, August 11, 2001. The banquet will be held Saturday evening at Historic Savannah Station. Information regarding the Savannah area can be located at <www.savannah-online.com/>.
Puzzle Ball Foraging Device for Laboratory Monkeys
Carolyn M. Crockett, Rita U. Bellanca, Kelly S. Heffernan*, Dennis A. Ronan, and Wayne F. Bonn
Regional Primate Research Center, University of Washington
Foraging is considered to be a crucial element in promoting psychological well-being in nonhuman primates. The USDA Draft Policy on Environmental Enhancement for Nonhuman Primates (U.S. Department of Agriculture, 1999) specifies that environmental enhancement plans should provide for daily foraging opportunities. At the Washington Regional Primate Research Center (WaRPRC), we define a foraging experience as "any food or drink enrichment that requires extra manipulation and prolongs consumption time, thus providing mental stimulation." Simple food treats, such as small pieces of fresh produce, do not meet our definition of foraging. Food treats are given about four days per week (Bellanca et al., 1998). In addition, animals currently have periodic foraging experiences (usually at least once every week or two), primarily frozen treats and browse. Some animals are also provided with foraging devices. Two years ago, we recognized the need to enlarge our inventory of such devices to increase the laboratory primates' foraging experiences. If every cage were equipped with a foraging device, each could, in principle, be provisioned daily to meet the anticipated USDA Policy.
We wanted a foraging device that was inexpensive, durable, and effective, and that could be sanitized during routine cage washing. Psychological Well-being (PWB) Program staff worked with Colony Facilities Maintenance staff to design, fabricate, and test a device which we call the "Puzzle Ball". An earlier design of this foraging device was developed at WaRPRC (Murchison, 1992) and later modified at the Tulane RPRC to include three access holes (M. Murchison and S. Falkenstein, pers. comm.). The design described here includes modified access holes and a redesigned attachment. The final product has been in use at the WaRPRC since autumn of 1998.
The Puzzle Ball Foraging Device
The Puzzle Ball foraging device is constructed of stainless steel hardware and a commercially available plastic Boomer Ball(r) (Grayslake, IL), 4" to 4.5" in diameter (Figure 1). The original device had a single 0.75" diameter access hole (Murchison, 1992). We conducted observations to determine the appropriate number and size of holes (see next section). The final design included one 1" and two 0.75" diameter access holes, located above the midline and approximately equidistant from each other. The 1" hole provides easier access for larger fingers, including those of adult baboons, and provides a larger target for personnel provisioning the puzzle. We also found that Puzzle Balls sanitized while affixed to cages in the cage washer got cleaner with three access holes rather than two. Three holes above the midline appeared to allow more water to flush through the ball and out of the 3/8" drain hole at the bottom. The balls received from the supplier vary from 5/32" to nearly 3/8" in thickness and from 4" to 4.5" in diameter. This variation has not notably affected durability or use of the foraging device. We request that shipments include as many colors as are available, and our inventory of puzzle balls includes blue, red, yellow, orange, green, purple and turquoise. Although we have not tested whether color variation makes a difference to the monkeys, visitors and staff remark that it makes the housing rooms much more colorful and appealing.
Figure 1: : Puzzle Ball foraging device fabricated from a Boomer Ball(r) and stainless steel hardware. The standard nut is secured with a drop of Locktight(r).
Because non-stainless hardware used in some previous enrichment devices had rusted, we opted for stainless steel. The chain (six 1.25" links, 1/8" trade size, Type 316L; including bent and split links) was selected for strength and to minimize finger entrapment. The length, about 5.25", is short to avoid entanglement (Bielitzki, 1992). After considerable discussion and some experimentation, we decided that the Puzzle Balls would be attached permanently on the cages with a split link of the type of chain used to suspend the device. Sharp edges of the split links are deburred with a torch before use. Maintenance shop personnel invented a simple device to easily open and close the split link (Figure 2). Using brass locks (our standard cage locks) to attach the Puzzle Balls is too expensive, and snap hooks are too often removed by the monkeys. Several other attachments we tried were either flimsy or potentially hazardous. For example, the "Du Clip" (resembles a large paper clip) was removed by some animals, and caused at least one pinching injury, according to veterinary staff.
Another feature of the Puzzle Ball is its attachment to the chain with a self-locking nut. When worn out, the Puzzle Ball can be removed from the chain and easily replaced. The nuts and bolt can be recovered and used again. So far, only a few of the several hundred Puzzle Balls in use have had to be thrown away because of excessive damage from chewing. We cannot distinguish when individual balls have been placed in service, but most of those that are two years old are still in use.
Figure 2: : Stainless steel device for attaching Puzzle Balls: A. Open split link hooked on end of chain. B. Split link hooked over cage wire; slot of device is slipped over half of link to be closed and is turned (three slots allow various angles of leverage). C. Closed split link. Opposite action opens link.
The cost of the device is less than $8 in materials ($5 per ball; $2.20 for stainless steel hardware) and about 10 minutes of fabrication time. A recent batch of 85 Puzzle Balls was fabricated in 10 hours. Our goal is for all individual cages to have foraging devices, so permanent attachment is the least expensive option. Although we will eventually need one Puzzle Ball per individual cage, including empty ones in the cage wash cycle, the expense will be easily compensated for by the elimination of personnel time to move the devices from cage to cage.
Puzzle Balls are attached outside of the cage. Location of attachment varies from cage to cage. Those with food boxes located centrally below the cage door usually have the Puzzle Ball attached toward one side, above where the perch is installed. Those with food boxes to one side of the door often have the Puzzle Ball attached above the food box so that most spilled items fall in the hopper. However, husbandry staff request that the ball not be so close to the hopper opening that biscuit feeding is obstructed.
Testing the Device
Methods: Behavioral observations on four pigtailed macaques (Macaca nemestrina: two adult males, one adult female, one juvenile female) and three longtailed macaques (M. fascicularis: one adult male, two adult females) helped determine the final design of the Puzzle Ball. Each animal was observed for 10 minutes after six pieces of cereal were placed in the ball attached to that subject's cage. Three types of ball were presented to the subjects, a different one on each of three different days. Type A most resembled the "Tulane" model with three 0.75" holes. Type B had two 0.75" holes. Type C had two 1" holes. Behavior was recorded every 30 seconds using the instantaneous scan sampling method (Crockett, 1996).
Results: Overall, the subjects manipulated the Puzzle Ball during 69.5% of the scan samples. These observations also verified that the animals rarely manipulated the self-locking nut and were not able to remove it. Four of the seven subjects were able to successfully empty (eat plus spill) at least one type of Puzzle Ball in less than 10 minutes. (Most spilled about as much as they ate.) For the successful animals, it took an average of five minutes to empty the puzzle. The record was achieved by an adult female longtailed macaque who removed and ate all six pieces of cereal from a Type C puzzle in less than one minute. The other successful subjects were the second female longtailed macaque, and one adult male and the adult female pigtailed macaques. Across all seven subjects, the Puzzle Ball was emptied in less than 10 minutes in 35% of the trials; 57% of these cases were with the Type C ball. About 50% of the food was spilled from Type A ball, about 30% from Type B, and only 20% from Type C. In the end, we decided that one 1" hole and two 0.75" holes would be a good compromise, to facilitate cleaning, to provide holes for various finger sizes, and to add an element of choice. We have not conducted any follow-up observations to see if animals preferentially take food items out of the larger hole. We have verified that juvenile and adult baboons (Papio cynocephalus) are also adept at using the Puzzle Ball.
Effect of Puzzle Ball on Abnormal Behavior
Methods: Because the Puzzle Balls are being fabricated and placed on cages over a period of many months, we were able to conduct an informal study on some animals with and without the devices. Seventeen individually housed animals 3-11 years of age (12 male and one female M. nemestrina, two male and two female M. fascicularis) were observed for at least four 10-min observations with and four 10-min observations without the Puzzle Ball. These animals had been referred to the PWB Program for behavioral assessment; their behavior was scored on our standardized data sheet. We have found that four 10-min observations (30-sec scan samples) are sufficient to characterize behavior profiles of most animals referred for assessment (Bellanca et al., 1999). All observations were made when the ball was empty. Our intention was to see if the presence of the ball had any effect on abnormal behavior during periods when it was empty. In other words, we wanted to know if any beneficial effects of having a regularly provisioned device extended beyond the time animals were actually foraging. During the time these observations were made, less than half of the cages had Puzzle Balls. Because the Puzzle Balls are permanently attached, animals were rotated into cages with them and without them on an unpredictable basis. Because the number of devices increased over time, the majority of the "without Puzzle Ball" observations occurred before the "with Puzzle Ball" observations. Results: The presence of a Puzzle Ball was associated with a significant reduction in abnormal behavior (locomotor stereotypies, self-stimulation, and potential self-injurious behavior combined) in animals referred to the PWB Program for behavioral assessment. The median proportion of scan samples when the subjects engaged in abnormal behavior was .188 without the Puzzle Ball and .075 with the puzzle ball (Wilcoxon Signed Rank Test, z-statistic = 2.5, p= 0.012; Velleman, 1997). Even though the Puzzle Balls were empty during observations, the subjects did manipulate them, although an average of only 1.6% of the time. During the same observations (Puzzle Ball present), the subjects manipulated their portable cage toys (at least one per cage) an average of only 0.9% of the time. (Eight of 17 manipulated neither Puzzle nor toy.)
Discussion
An underlying policy of the WaRPRC's Psychological Well-being Program is to test enrichment methods before adopting them for general use. We incorporate research into most aspects of our environmental enhancement and behavioral assessment activities. The development of the Puzzle Ball foraging device also illustrates the essential collaboration between staff responsible for environmental enrichment and those responsible for maintaining caging and other colony facilities.
The Puzzle Ball has proven to be a durable and effective foraging device. It is inexpensive, easy to make, and easy to clean. The device is simple and requires more manual dexterity than mental acuity. However, it increases consumption time for all but the most dexterous animals. There is probably more spillage because the device has three holes rather than two, but we compromised on the side of easier sanitizing. Although we continue to use existing PVC tube puzzles (as described in Heath et al., 1992, and Murchison, 1991, as well as larger models), we are unlikely to replace them as they wear out. Such puzzles are more expensive, are more labor intensive to clean and move from cage to cage, and also obstruct more of the cage front. Even when empty, the Puzzle Ball is used as an anchored object of manipulation. The balls are handled, chewed, and batted, sometimes by animals in neighboring cages, occasionally inspiring minor aggressive interactions or affiliative touching. The most frequent signs of wear are tooth marks around the access holes. We have found that Boomer Balls last much longer when made into Puzzle Balls than when put inside cages.
Because animals are usually successful at removing food items from the Puzzle Balls, and because the three access holes and the drain hole permit water to flush through them during cage washing, residue rarely accumulates. However, we recommend avoiding sticky items (raisins, mini-marshmallows) or items likely to become gummy when wet (pasta, sugary cereals). Such items also stick to the floor when spilled. Peanuts and dry cereal are our usual provisioning items. Filberts are a good alternative for animals not allowed peanuts. Although some animals helpfully hold the Puzzle Ball to facilitate loading, most are not so cooperative. We have experimented with various "loading" devices, such as modified funnels, but have not yet found a successful one. It is definitely easier to provision the 1" hole than the 3/4" holes. Some animals are grabby, so personnel must be careful, as always when working with nonhuman primates. Double gloves are recommended, and we wear Tyvex sleeves when provisioning puzzles of animals with infectious diseases.
We were pleased that the empty Puzzle Balls were associated with a reduction in abnormal behavior. This is a somewhat surprising result because effects of foraging devices on abnormal behavior rarely extend beyond the time spent emptying them (Novak et al., 1998). However, because the "without Puzzle Ball" observations tended to occur earlier than those "with Puzzle Ball", we suspect that the present result reflects a general increase in psychological well-being due to increased enrichment over time, including regular provisioning of the Puzzle Ball. Although the Puzzle Ball is relatively simple, more difficult puzzles are not necessarily more beneficial. Instead of being mentally stimulated, animals may give up trying to solve them (Heath et al., 1992; Novak et al., 1998).
Meeting the proposed goal of daily foraging (U.S. Department of Agriculture, 1999) can be facilitated by the use of foraging devices like the Puzzle Ball. Although our facility does not yet have them, we also support the idea of providing biscuits in forage feeders (Reinhardt & Garza-Schmidt, 2000). However, we do not believe that forage feeders can take the place of puzzle feeders, since the latter are provisioned with treat foods rather than regular biscuits. An ideal enrichment program would include both.
References
Bellanca, R. U., Crockett, C. M., Johnson-Delaney, C., DeMers, S. M., & Eiffert, K. (1998). Catering to catarrhines: Food enrichment at the University of Washington's Regional Primate Research Center. American Journal of Primatology, 45, 167-168.
Bellanca, R. U., Heffernan, K. S., Grabber, J. E., & Crockett, C. M. (1999). Behavior profiles of laboratory monkeys referred to a Regional Primate Research Center's Psychological Well-being Program. American Journal of Primatology, 49, 33.
Bielitzki, J. T. (1992). Letter to the editor: Enrichment hazards. Laboratory Primate Newsletter, 31, 36.
Crockett, C. M. (1996). Data collection in the zoo setting, emphasizing behavior. In D. G. Kleiman, M. E. Allen, K. V. Thompson, S. Lumpkin, & H. Harris (Eds.), Wild mammals in captivity: Principles and techniques (pp. 545-565). Chicago: University of Chicago Press.
Heath, S., Shimoji, M., Tumanguil, J. & Crockett, C. (1992). Peanut puzzle solvers quickly demonstrate aptitude. Laboratory Primate Newsletter, 31, 12-13. Murchison, M. A. (1991). PVC-pipe food puzzle for singly caged primates. Laboratory Primate Newsletter, 30, 12-14.
Murchison, M. A. (1992). Task-oriented feeding device for singly caged primates. Laboratory Primate Newsletter, 31, 9-11.
Novak, M. A., Kinsey, J. H., Jorgensen, M. J., & Hazen, T. J. (1998). Effects of puzzle feeders on pathological behavior in individually housed rhesus monkeys. American Journal of Primatology, 46, 213-227.
Reinhardt, V. & Garza-Schmidt, M. (2000). Daily feeding enrichment for laboratory macaques: Inexpensive options. Laboratory Primate Newsletter, 39, 8-10.
U.S. Department of Agriculture. (1999). Animal welfare: Draft policy on environment enhancement for nonhuman primates. Federal Register, 64, 38145-38150.
Velleman, P. F. (1997). Data desk: The new power of statistical vision. Ithaca, NY: Data Description Inc.
----------
Authors' address: Regional Primate Research Center, Box 357330, University of Washington, Seattle, WA 98195-7330 [e-mail: [email protected]]; *K. S. Heffernan is now at SNBL USA, Ltd. (Shinn Nippon Biomedical Laboratories, Ltd., Everett, WA 98203).
This research was supported by NIH grant RR00166. We thank N. Dickson and K. Hagerman for observation assistance; D. Shaw for inventing the attachment device; R. Shain for fabrication assistance; J. Gray for design input; K. Elias for editorial comments; and M. Domenowske for the illustrations.
----------
* * *
Workshop Announcements
Teaching Research Ethics
Indiana University's eighth annual Teaching Research Ethics Workshop will convene on the campus at Bloomington, Indiana, May 9-12, 2001. Session topics will include an overview of ethical theory; using animal subjects in research; using human subjects in clinical and non-clinical research; and responsible data management. Many sessions will feature techniques for teaching and assessing the responsible conduct of research.
For more information, contact Kenneth D. Pimple, Teaching Research Ethics Project Director, Poynter Center, Indiana Univ., 618 East Third St, Bloomington, IN 47405-3602 [812-855-0261; fax: 812-855-3315; e-mail: [email protected]]; or see [www.indiana.edu/~poynter/].
Orangutan Reintroduction and Protection
A workshop on Orangutan Reintroduction and Protection will be held June 15-18, 2001, at Wanariset-Samboja and Balikpapan, E. Kalimantan, Indonesia. It will mark the 10th anniversary of the Wanariset Orangutan Reintroduction Project (ORP). ORP is sponsoring this international workshop with the aims of presenting and evaluating its own operations and research over the last ten years, discussing future directions, and considering broader issues of orangutan conservation. For further information, abstract submission forms, or to discuss possible contributions, please contact Jeane Mandala, P.O. Box 500, Balikpapan 76103, Indonesia [+62 (0)542 413 069; fax: +62 (0)542 410 365; e-mail: [email protected]]; or Dr. A. Russon, Secretary, Scientific Advisory Board, Dept. of Psychology, Glendon College, 2275 Bayview Ave., Toronto, Ontario M4N 3M6, Canada Tel: [+1 416 736 2100 ext. 88363; fax: 1 416 487 6851; e-mail: [email protected]]. The deadline for abstracts is March 15, 2001.
* * *
Who's Enriching Whom? The Mutual Benefits of Involving Community Seniors in a Research Facility's Enrichment Program
Nancy Megna and Jessica Ganas
Yerkes Regional Primate Research Center Field Station and Antioch New England Graduate School
In hundreds of research facilities across the world live thousands of nonhuman primates that get lonely, bored, and frustrated. They often live in conditions which deprive them of the opportunity to express species-typical behaviors such as social interactions, foraging, and travel. These conditions can lead to abnormal behaviors such as self-mutilation, stereotypies, and depression. It is the duty of animal caretakers and veterinary and research staff to ensure that the psychological well-being of these animals is addressed. According to the Animal Welfare Act, Section 3.81, "Dealers, exhibitors and research facilities must develop, document, and follow an appropriate plan for environmental enhancement adequate to promote the psychological well-being of nonhuman primates."
In order to meet the needs of the 1700 nonhuman primates living in cages, runs, and compounds at the Yerkes Primate Research Center Field Station, located in Lawrenceville, Georgia, the authors, behavioral researchers Nancy Megna and Jessica Ganas, began APES, or "Alliance for Primate Enrichment by Seniors." Even though enrichment is required by law, time is often a major constraint for staff trying to create challenging and varied enrichment items for the animals. On the other end of the spectrum, time is something that nursing home residents have a lot of. We felt that perhaps seniors would be interested in creating enrichment treats for the Field Station's monkeys and apes. We contacted a local nursing home, the Medical Arts Health Facility in Lawrenceville, and spoke with the Director of Activities, Merri Kaye Robinson. After hearing what the program would entail, she enthusiastically agreed to help start APES.
The program began in January, 2000, with about ten residents who gathered for the first session. Since then, attendance has increased at each meeting. Often the seniors line up outside of their activities room ready to go before the session begins! The supplies used to make the enrichment treats are empty paper towel and toilet paper rolls collected by Yerkes employees, Dixie cups, and lunch bags. Food items are taken from the regular enrichment supplies of the Field Station or are donated by Yerkes employees or the nursing home. The foods include popcorn, nuts, pretzels, seeds, cereals, raisins, banana chips, currants, and uncooked noodles and rice. A mixture of items is placed in the cups, rolls or bags. Once the containers are half full, they are closed so that the food inside cannot be accessed without some effort by the animals. On their own, the seniors have divided duties based on manual dexterity. Some of the seniors crush items, some fill the cups, and some fold them closed. In one hour the seniors can make about 500 Dixie cups and 40 rolls, as well as many lunch bags.
Figure 1 Immediately, we began to notice how APES affected both seniors and primates. It has become the most highly attended activity at the Medical Arts Nursing Home; the level of enthusiasm is overwhelming. The seniors involved in the program take it very seriously. To them, this is their job - and they work very hard at it. They have gained a sense of self-worth and purpose because they are contributing to animal welfare. They take much enjoyment in repaying the animals for their contributions to medical research.
Figure 2: The apes and monkeys are benefiting tremendously from the program as well. The treats made for them are challenging - they have to work to get their food. There may be new treats inside each cup, bag, or roll, providing variety - and the food and paper both serve as enrichment. The care staff benefit from the time saved.
The residents at the Medical Arts Health Facility have contributed greatly to the well-being of Yerkes Field Station primates. The seniors deserved to be rewarded with more than a simple "Thank you," so we invited them to tour our facility. The mutual benefits of the program were apparent when seniors and primates were brought together, as shown by the residents' excitement and delight as they watched the primates enjoying the items they had prepared. Because some of the seniors are not ambulatory, we show them pictures and videotapes of the primates with the enrichment; some of the pictures are on a bulletin board at the nursing home. We will be scheduling more tours and thanking each senior with a certificate of appreciation in May during National Nursing Home Week.
The APES program has future plans. We will be expanding the repertoire of items that the seniors can prepare for the animals. Second, we plan to approach corporations and organizations for donations of food and/or materials. Third, we would like the Yerkes Main Center to participate in the program. We would also like to include other nursing homes in the area, since the program has been successful. Finally, by sharing our story of the program's success and demonstrating how easily it can be started and maintained, we hope to inspire other animal research facilities to start such programs.
* * *
----------
First author's address: 133 Sir Gregory Manor, Lawrenceville, GA 30044 [e-mail: [email protected]].
----------
* * *
Early vs. Natural Weaning in Captive Baboons: The Effect on
Timing of Postpartum Estrus and Next Conception
Janette Wallis and Buddy Valentine
University of Oklahoma Health Sciences Center
Introduction
The period following birth in baboons is characterized by cessation of the mother's reproductive cycles during the early phase of lactation. As in humans and anthropoid apes, nursing on demand tends to delay the resumption of reproductive cycles (Heinig et al., 1994; Vitzthum, 1994). As infants begin to suckle on a less consistent schedule, the mother's menstrual cycles resume.
When a wild infant baboon dies, regardless at what age, the mother typically resumes cycling within one month and becomes pregnant again within three months (Altmann et al., 1978). If the infant survives, however, lactation suppresses cycle resumption for about 12 months, although the infant may continue to suckle for up to 17 months (Altmann et al., 1977). In captivity, where nutrition is generally better than in the wild, cycle resumption has been reported to occur earlier - as early as six months postpartum (Rowell, 1966). At that age, the infant is still psychologically dependent on the mother, but eats enough solid foods to allow survival away from the mother. Thus, many laboratory facilities use six months (approximately 180 days) as an arbitrary age at which to separate infants from their mothers in the effort to stimulate the mother's reproductive cycles and maximize reproduction. At the University of Oklahoma Health Sciences Center, this policy was in practice for a number of years.
In both human and nonhuman primates, however, infants can suffer serious and lasting effects when separated - for even a brief period - from their mothers (Kaufman & Rosenblum, 1967; Palloni & Millman, 1986). For baboons, weaning too early can have negative effects on a youngster's behavioral and physical maturation (Glassman & Coelho, 1988; Rhine et al., 1980). The optimal environment for any young primate is with its mother (Pazol & Bloomsmith, 1993).
In 1998, we began a program to enhance and expand our baboon resource program. In addition to establishing an environmental enrichment plan, we are assessing the efficacy of early infant weaning on both the reproductive performance of the overall colony as well as the behavioral well-being of individual baboons. An ongoing study compares the progress of social development in early (forcibly) weaned vs. naturally weaned infants (to be published elsewhere). The present study investigated the timing of infant removal and its influence on postpartum estrus and subsequent conception in captive baboons (Papio spp.).
Subjects and Methods
The subjects were baboons (Papio sp.) living at the Baboon Resource Center at the University of Oklahoma Health Sciences Center. Although detailed histories are lacking for many of these subjects, the majority of the baboons appear to be of the olive subspecies (Papio hamadryas anubis), with a possibility that some have mixed ancestry with yellow baboons (Papio h. cynocephalus).
From the colony records dating from March 1998 through August, 1999, we examined the details of 73 recorded pregnancies of 45 adult females. Of the 73 pregnancies under review, six were deemed apparent miscarriages, four were stillbirths, and five were neonatal deaths (by age 9 days; none occurred after that). In addition, four of the infants were removed for human rearing before five days of age due to incompetent maternal care. Only the time delay until onset of postpartum cycles was assessed for these 19 cases; none were used in the analysis of gestation length. The remaining 54 cases (live births) were assessed for the colony's gestation length. We used an analysis of variance (ANOVA) to test for possible gender difference.
The subjects lived in five large social groups, in indoor, double-sided cages measuring 182 ft.2 each. The groups consisted of 10-12 adult females and various offspring. Three of the groups contained one adult male as a permanent resident, while the other two groups shared one male. The presence of a male influences the timing of reproductive parameters investigated in this analysis. In addition to the obvious parameters, such as time of conception and, consequently, parturition, a male's presence has even been reported to influence the timing of cycle resumption (Colmenares & Gomendio, 1988). Thus, for each analysis of these parameters, we omitted cases when a male was not present.
The past policy at this facility had been to forcibly wean infant baboons at approximately 180 days of age, and pair-house them with a like-aged peer. From March, 1998, to September, 1998, 21 infants were weaned at ages ranging from 167 days to 305 days. For the 18 cases involving an adult male living in the social group, we determined the following dates: conception, parturition, infant removal, onset of postpartum estrus, and next conception. Date of conception was arbitrarily estimated as the last day the mother exhibited full genital swelling, based on a number of studies which found ovulation to occur one or two days prior to detumescence (Gillman & Gilbert, 1946; MacLennan & Wynn, 1971; Hagino, 1974). Using these dates, we calculated the infants' ages at removal and several reproductive parameters. A Pearson's Correlation was used to determine the interrelationship between reproductive parameters. An ANOVA was used to assess the effect of forced weaning on these parameters.
Beginning in October, 1998, we ceased the routine practice of forcibly weaning infant baboons in our colony. Thus, we added to the analysis a comparison of reproductive parameters resulting when infants were forcibly weaned (regardless of age) vs. when infants remained with their mothers to be naturally weaned. There were 33 cases in the latter category. Only 21 of these included an adult male in the group and had subsequent reproductive events occurring during the period under review.
Results
The gestation length for 54 live baboon births was 181.6 ± 6.9 days. There were 32 female and 22 male infants. No twin births occurred and there was no gender effect on gestation length (female infants averaged 181.3 ± 7.2 days' gestation and males averaged 182.2 ± 6.6 days.
Predictably, in the 19 cases of miscarriage, stillbirth or early neonatal death (within 9 days), the mothers resumed having menstrual cycles quite rapidly. For these females, the first sign of postpartum estrous genital swelling appeared at an average of 28.5 days (range = 17-39 days). Effect of Early Weaning on Mothers' Cycles: For the 54 live births, there was a significant correlation between age at removal and duration from that time until postpartum estrus (r2 = -.72; p = .0007). This was a negative relationship, however; the earlier an infant was weaned, the longer it took for the mother to resume reproductive cycles. There was no correlation between infant removal age and the length of time to the next conception or birth.
Comparison of Age at Early Weaning: For cases in which infants were forcibly weaned, the data were divided into two groups: 8 infants removed at < 180 days of age (range 167-180 days), and 10 infants removed at > 180 days (range 181-305 days). The two groups differed somewhat in the time from infant removal to the onset of postpartum estrus, with the difference approaching significance (f = 4.16; p = .058). This finding resulted from the fact that postpartum cycles typically resumed before infant removal. Consequently, from the time of birth to the onset of postpartum estrus, the two groups showed no significant difference (mean of 173.4 and 167.7 days, respectively), nor were subsequent conception and parturition significantly influenced by time of infant removal. Comparison of Early Weaning vs. Natural Weaning: The 18 forced weaning cases (above) were compared to 21 cases in which infants were allowed to remain with their mothers to be naturally weaned. There was no significant difference between these two groups for the time between birth and the onset of postpartum estrus, the next conception, or the next parturition (Table 1). There was also no difference in the time between the onset of postpartum estrus until conception: 62.5 days vs. 59.0 days. Although none of these comparisons showed a statistically significant difference, the mean values were smaller for the naturally weaned cases than the forcibly-weaned cases (see Table 1).
| Weaned Early | Naturally Weaned | ANOVA | |
| Birth to Next Postpartum Estrus | 170.22 days (N=18) | 165.2 days (N=21) | F = .10 n.s. |
| Birth to Next Conception | 233.6 days (N=14) | 210.3 days (N=13) | F = 1.14 n.s. |
| Birth to Next Birth | 413.1 days (N=14) | 377 days (N=2) | F = 1.15 n.s. |
| Postpartum Estrus to Next Conception | 62.5 days (N=14) | 59.00 days (N=13) | F = .03 n.s. |
Table 1: Comparison of cases when infants were weaned early vs. when infants stayed with the mother for natural weaning
Only two births occurred during the study period to a mother who had naturally weaned her previous infant. The interbirth interval for those two cases averaged 377 days, compared to the average of 413.1 days for the other mothers. (For the live births that occurred after the study period, the mean for seven interbirth intervals was 384.9, still well below the figure when infants were removed.)
Discussion
The results of this study indicate that forced infant weaning did not improve reproductive productivity in our colony of baboons. In fact, the data indicate that most females resumed their reproductive cycles well before infant removal and, when given the opportunity for natural weaning, the females conceived while their previous infants were still dependent upon them (Table I). Based on these findings, we have encouraged the managers of our colony to allow natural weaning whenever possible. Unfortunately, some research protocols call for young subjects to live in small peer groups. Therefore, the colony managers' policy has not changed appreciably, although they have reduced pair-housing infants in favor of forming small peer groups.
The potential benefits of allowing infants to remain with their mothers throughout childhood are obvious. The first year of life for a baboon is a period of helplessness and dependency, during which the mother provides important companionship and care (Altmann, 1980). Rhine et al. (1984) suggested that the period centering upon the sixth month is a time when maturing motor, sensory, emotional, and exploratory systems are undergoing rapid integration with social learning. Cheney (1978) suggests that interactions with individuals of various ages other than the mother will facilitate the development of independence in a more relaxed and natural manner. Siblings and peers can provide a major source of social contact in early baboon life (Ransom, 1981). Allowing infants to be naturally weaned by their mothers provides crucial interaction that is required for normal behavioral and social development and, thus, may reduce the development of abnormal behavior. This is a key point for laboratory managers to consider. Because most captive research often focuses on the use of primates as "animal models" for humans, it is imperative that colony managers seek to produce the healthiest subjects possible. This must include attention to behavioral as well as physical health.
In addition to the benefits to psychological development, living in a complex social environment can facilitate learning skills for reproduction and parenting in baboons (Rhine et al. 1980; Hamilton et al., 1982). For example, our ongoing behavioral study indicates that infant males living with their mothers, in a mixed age social group, show playful attempts at sexual behavior (e.g., attempted mounting with erection) as young as one year of age. In contrast, we have a four-year-old forcibly weaned, human-reared male baboon who has yet to show any indication of sexual interest despite his living with many adolescent females (J. Wallis, unpublished data).
The results of this study may be a consequence of several changes in the subjects' environment. In addition to allowing natural weaning when possible, we have implemented a new enrichment program. Among other activities, our enrichment plan includes provision of novelty food items and the addition of climbing structures in the cages. The occasional food treats help to supplement the infants' normal diet. For baboons in the wild, a major determinant of weaning age is the availability of easily eaten "weaning foods" to supplement milk (Altmann 1980). Thus, through the novel treats, we may be providing enough supplemental foods to slightly alter suckling patterns.
The added climbing structures, inexpensively constructed with PVC pipe, metal conduit, and chain, enhance the use of vertical space, thus reducing crowding. Wilson (1972) noted that the number and complexity of structures present in captive primate environments are more important than the actual size of the enclosures. Adding moveable and nonmoveable objects to captive baboon cages results in higher levels of activities such as play and locomotion (Kessel & Brent, 1996). The benefits of such social play include the development of motor skills and facilitation of communication leading to social bonding (Fagen, 1993). Together, the reduced stress, increased motor activity, and presence of many infants of various ages approximates a more species-normal social group and may facilitate reproductive success and psychological well-being (Novak & Suomi, 1991).
The findings of this study suggest that the best strategy for baboon breeding programs is to allow natural weaning whenever possible. The benefits include:
Reproduction: Reproductive output for the naturally weaned group is comparable to that of the forcibly weaned group.
Management: Group housing reduces the use of individual or pair housing, which in turn reduces the animal care workload: cleaning and feeding of gang cages tends to be less time-consuming than care of individually caged subjects.
Compliance: Maintaining larger and more complex social groups allows for better compliance with the USDA guidelines (USDA, 1999).
Psychological well-being: Allowing infant baboons to stay with their mothers and grow up in a socially complex setting provides the opportunity for the development of more natural and normal social behavior.
References
Altmann, J. (1980). Baboon mothers and infants. Cambridge: Harvard University Press.
Altmann, J., Altmann, S. A., & Hausfater, G. (1978). Primate infant's effects on mother's future reproduction. Science, 201, 1028-1030.
Altmann, J., Altmann, S. A., Hausfater, G., & McCuskey, S. A. (1977). Life history of yellow baboons: Physical development, reproductive parameters, and infant mortality. Primates, 18, 315-330.
Cheney, D. L. (1978). Interactions of immature male and female baboons with adult females. Animal Behaviour, 26, 389-408.
Colmenares, F. & Gomendio, M. (1988). Changes in female reproductive condition following male take-overs in a colony of hamadryas and hybrid baboons. Folia Primatolgica, 50, 157-174.
Fagen, R. (1993). Primate juveniles and primate play. In M. E. Periera & L. A. Fairbanks (Eds.), Juvenile primates: Life history, development, and behavior (pp. 182-196). New York: Oxford University Press.
Gillman, J. & Gilbert, C. (1946). The reproductive cycle of the chacma baboon (Papio ursinus) with special reference to the problems of menstrual irregularity as assessed by the behaviour of the sex skin. South African Journal of Medical Sciences, 11, 1-54.
Glassman, D. M. & Coelho, A. M., Jr. (1988). Formula-fed and breast-fed baboons: Weight growth from birth to adulthood. American Journal of Primatology, 16, 131-142.
Hagino, N. (1974). Follicular maturation, ovulation, luteinization and menstruation in the baboon. In E. M. Coutinho & F. Fuchs (Eds.), Physiology and genetics of reproduction, Part A (pp. 323-342). New York, Plenum. Hamilton. W. J., Busse, C., & Smith, K. S. (1982). Adoption of infant orphan chacma baboons. Animal Behaviour, 30, 29-34.
Heinig, M. J., Nommsen-Rivers, L. A., Peerson, J. M., & Dewey, K. G. (1994). Factors related to duration of postpartum amenorrhea among USA women with prolonged lactation. Journal of Biosocial Science, 26, 401-409.
Kaufman, C. I. & Rosenblum, L. A. (1967). Depression in infant monkeys separated from their mothers. Science, 155, 1030-1031.
Kessel, A. L. & Brent, L. (1996). Space utilization by captive-born baboons (Papio sp.) before and after provision of structural enrichment. Animal Welfare, 5, 37-44.
MacLennan, A. H. & Wynn, R. M. (1971). Menstrual cycle of the baboon: I. Clinical features, vaginal cytology and endometrial histology. Obstetrics and Gynecology, 38, 350-358.
Novak, M. A. & Suomi, S. J. (1991). Social interaction in nonhuman primates: An underlying theme for primate research. Laboratory Animal Science, 41, 308-314.
Palloni, A., & Millman, S. (1986). Effects of interbirth intervals and breastfeeding on infant and early childhood mortality. Population Studies, 40, 215-236.
Pazol, K. A. & Bloomsmith, M. A. (1993). The development of stereotyped body rocking in chimpanzees reared in a variety of nursery settings. Animal Welfare, 2, 113-129.
Ransom, T. W. (1981). Beach troop of the Gombe. Lewisburg, PA: Bucknell University Press.
Rhine, R. J., Norton, G. W., Roertgen, W. J., & Klein, H. D. (1980). The brief survival of free-ranging baboon infants (Papio cynocephalus) after separation from their mothers. International Journal of Primatology, 1, 401-409.
Rhine, R. J., Norton, G. W., & Westlund. B. J. (1984). The waning of dependence in infant free-ranging yellow baboons (Papio cynocephalus) of Mikumi National Park. American Journal of Primatology, 7, 213-228.
Rowell, T. E. (1966). Forest living baboons in Uganda. Journal of Zoology, 149, 344-364.
United States Department of Agriculture, Animal and Plant Health Inspection Service. (1999). Final report on environment enhancement to promote the psychological well-being of nonhuman primates.
Vitzthum, M. J. (1994). Comparative study of breastfeeding structure and its relation to human reproductive ecology. Yearbook of Physical Anthropology, 37, 307-349.
Wilson, C. C. (1972). Spatial factors and the behavior of nonhuman primates. Folia Primatologica, 18, 256-256.
----------
Corresponding author: Janette Wallis, Department of Psychiatry and Behavioral Sciences, University of Oklahoma Health Sciences Center, P.O. Box 26901, Oklahoma City, OK 73026 [e-mail: [email protected]].
We acknowledge the support of the OUHSC Baboon Resource Center animal care staff, W. Hill, G. Spiegel, P. Griffith, and J. Weigant, as well as the veterinary staff, G. White, R. Wolf, and M. Cary. We wish to dedicate this paper to the memory of Ray Rhine, who taught the world so much about young baboons and what makes them tick. This work was supported by NIH/NCRR grant RR12317-01A1.
----------
The following responses were received:
From Jo-Ann C. Jennier, Jackson Zoo [[email protected]]: Our red-ruffed lemurs love to hang from the top of their cage by their hind feet. Perhaps you could try hanging treats (e.g., sweet potato) from a low tree limb to elicit this fascinating behavior. This would be enriching for both the lemurs and the observers.
From: JoAnne Kowalski, Philadelphia Zoo [[email protected]]: We've used coated mesh to make feeders. Basically just make a box out of it and cut holes in it large enough for the lemurs to get the food out in their fists. You can also drill holes in the side of PVC piping and put screw-on caps on each end. Hang them up as feeders. Depending on the species, some lemurs will like to have browse to mess with. You can also freeze fruit, make ice cubes with pieces of fruit, or bigger blocks of ice with food in it. You can also check on different foods to use - blueberries, peas, Cheerios, raisins, corn, green beans, etc. Hope this helps.
From: Jo Fritz, Primate Foundation of Arizona [[email protected]]: Try the National Academy of Sciences' Guide for the Care and Use of Laboratory Animals, as a beginning point. You can access it at <www.nap.edu/readingroom/books/labrats/>. For environmental enrichment, there are several places, but start with: the Animal Welfare Information Center: <www/nal.usda.gov.awic>; Dr. Viktor Reinhardt: <www.animalwelfare.com/Lab-animals.biblio/enrich.htm>; and the Primate Enrichment Web Site: <www.brown.edu/Research/Primate/enrich.html>. For general primatology questions try AskPrimate: <primate.wisc.edu/pin/askprim.html>; the American Society of Primatology: <asp.org>; Living Links Web Site: <www.emory.edu/LIVING-LINKS/>; Sci-Web, the Life Science Home Page: <www.sciweb.com>; The Electronic Zoo: <netvet/wustl.edu/e-zoo.html>; or the World Wide Web Virtual Library's Biotechnology Information: <www.cata.com/biotech>.
From Bob Lewis, former caretaker, [[email protected]]: Puzzle feeders can be found at the Primate Products Website: <www.primateproducts.com/products/prod03.html>. These expensive doodads are hung on the outside of the cage. The animal routes food and treats through a maze. Do NOT do like a former supervisor of mine and place a puzzle feeder IN the cage with a primate - in this case, a female rhesus macaque. Your US$200 investment will end up in the rubbish as scraps of polycarbonate plastic.
From: Ian Colquhoun, University of Western Ontario, Scientific Advisor to the Black Lemur Species Survival Plan [[email protected]]: It should be remembered that "environmental enrichment" can be achieved in some respects through straightforward, low-tech (i.e., "inexpensive") measures. With lemurs, we are dealing with arboreal species. A basic bit of environmental enrichment would be to ensure that they have as complex a system of raised substrata as possible, to the extent that the size of the enclosure(s) and the management protocols of the animal care staff allow. That is, give them lots of branches, of various diameters and set at various angles - ideally, you'd be trying to reconstruct the three-dimensional complexity of a tree canopy to the best extent possible. As a graduate student, I spent several summers working on the animal care staff of a small, seasonal municipal zoo. I earned the title "branch manager" from colleagues thanks to my drive to provide as many raised supports as possible for the zoo's primates. But, the proof was in the pudding, as they say - the branches were used and definitely gave the animals options on their use of space and time outside of the periods that food was being consumed.
News Briefs
Ape Caper Foiled
Washington, D.C.: A male orangutan named Junior made a brief escape from the National Zoo's Great Ape House and zoo visitors were warned away from the area until he was recaptured. Junior is about 30 years old and had never before left the Ape House, zoo officials said, even though there are a series of towers orangutans can use to go back and forth between that enclosure and the zoo's animal Think Tank.
Keepers were "astonished" when Junior climbed out on one of the 40-foot towers and managed to escape, said zoo spokesman Robert Hoage. The animal got out about 11:45 a.m. and headed in the direction of the zoo police station, Hoage said. His absence was almost immediately detected. About 22 minutes later, the orangutan was "darted" with a tranquilizer gun fired by a veterinarian. The tranquilized animal was to be examined and officials said he might be back on exhibit as early as that afternoon.
The towers the animals climb are surrounded by electric "hot" wires to keep them from straying into the park. If the wires fail, lights are supposed to come on to alert keepers. "We have to learn if the power went off or if he defeated the system," Hoage said.
During Junior's escape, zoo visitors were cleared away from the area and confined to a section of the zoo near the Elephant House. - From the Washington Post, August 29
Death of a Chimpanzee at Coulston
Dr. M. K. Izard, Director of Behavioral Enrichment at the Coulston Foundation, posted an announcement to Alloprimate on September 13 about the death of Ray, a chimpanzee owned by NIH and under the care of the Coulston Foundation. "Ray was an HIV-exposed animal. On Wednesday, he was noted to be hypoactive with pale lips and gums. He had a normal appetite. On Thursday, he had a good appetite and normal stool. He was scheduled for a physical exam on Friday, yet was found dead that morning. Pathology reports show that Ray died of a systemic fungal (Coccidiodes immitis) infection that is fatal to an immunosuppressed host. Treatment would have been unsuccessful due to the progressive nature of this disease."
Anna Mae Noell, Owned "Chimp Farm"
Anna Mae Noell, who with her late husband owned and operated Noell's Ark Chimp Farm in Palm Harbor, Florida, died October 15. She was 86. The couple ran the popular and sometimes controversial roadside zoo since 1971.
Billing their farm as a retirement home for many cast-off primates, the Noells developed a reputation for taking in abandoned, old, and sick animals - mostly apes and monkeys - from small zoos and private owners. Mr. Noell died in 1991.
In recent years, state and federal officials, as well as several animal rights activists, criticized the facility for keeping animals in poor conditions. Last year, the U.S. Department of Agriculture permanently revoked the sanctuary's permit to show the animals to the public.
Now called the Suncoast Primate Sanctuary, the facility is run by a private, non-profit organization. The organization recently broke ground for a new primate facility, and volunteers there hope that it will lead to a reinstatement of the license to exhibit the animals. The sanctuary is home to 27 chimpanzees, three orangutans, two gorillas, 21 monkeys, a bear, two turtles, two alligators and two goats. - from an article by Robert Farley in the October 17 St. Petersburg Times, posted to Alloprimate by Hope Walker
LEMSIP Chimps Retire to Texas
Carol Asvestas announced October 25 on Alloprimate that the National Sanctuary for Retired Research Primates has received eight of 16 chimpanzees due to be retired. They arrived on October 19. The remaining eight will arrive December 10th. These chimps were not a social group, so great lengths were taken to make sure socialization was successful. Because of the help given by Dr. Thomas Rowell, Dr. James Mahoney (who stayed a week to be certain all was well), Dr. Dana Hasselschwert, and Chief Technician Danny Bouttay, the transfer and socialization were successful and without incident. The chimps, originally at LEMSIP, were being temporarily housed at New Iberia Research Institute. The animals were socialized within three and a half hours and are doing beautifully. "I can't tell you how much it meant to us and the chimps to have the support from these wonderful caring people. I thank them all for their amazing efforts and this incredible achievement."
Three New Lemur Species
Scientists from the United States, Germany, and Madagascar have announced that they have discovered three previously unknown species of mouse lemurs (see abstract on page 35). Lemurs are among the world's most endangered species. The newly discovered lemurs represent a small but encouraging sign to conservationists. - From an Associated Press announcement, posted to Alloprimate, November 14
Clinton Signs Great Ape Conservation Bill Into Law
On November 1, President Clinton signed into law H.R. 4320, the Great Ape Conservation Act of 2000, that will provide funding for conservation programs designed to protect chimpanzees, bonobos, gorillas and orangutans. The bill authorizes $5 million in annual U.S. aid to support conservation and protection of the great apes by allocating grants to local wildlife management authorities and other organizations in Africa and Asia dedicated to protecting the apes and their habitat. Although habitat destruction is the major threat to great ape populations, unregulated hunting and the international bushmeat trade pose an even greater threat in many areas. While national laws prohibit the hunting of great apes in most cases, forestry and wildlife officials often lack the basic resources required for enforcement. Funding through this Act will bolster projects aimed at strengthening law enforcement and by providing necessary resources and training for park officials in Africa and Asia. - From a Humane Society press release, posted to Alloprimate, November 5
Only the Latin binomial has any "right" or "wrong". Even here there will be disagreements and proposed names. The International Commission on Zoological Nomenclature will make decisions from time to time based on various principles. Their position, however, is that there is no right or wrong for common or "trivial" names. Howler, howling, and "black monkey with five fingers" are all equally good. So are the names in non-English languages. Brachyteles is the muriqui is the wooly spider. Macaca fascicularis is the long tail macaque, longtailed macaque, cyno, irus, kra, kera, Java, Phillipine, etc. I always say the grammatically incorrect thing when I talk of pigtail monkeys. Pigtailed, berok, or such have no greater validity. We all have our favorite names. As long as we mention the Latin binomial on first usage, we can call it whatever we wish. I note that in Belize they call howler monkeys "baboons". Certainly not desirable from my perspective, but not "wrong". You will notice a certain perverse consistency in common names so that if a scientific name becomes invalid some people will use it as the common name. This is the origin for rhesus, cynomologus, irus, etc. Some people now even refer to Macaca arctoides as speciosa, instead of the bear macaque, stumptail, stumptailed, or miniature chimp (a reference in an old Montgomery Ward's catalog offering them for sale as pets).
Best regards, Irwin Bernstein, Dept of Psychology, University of Georgia, Athens, GA [e-mail: [email protected]].
* *
Howler - Howler Monkey - Howling Monkey: Is there a legitimate reason for preferring one over the other? What rules apply to determining common names? Several years ago I was confronted with this controversy. I wrote an article for an International Journal of Primatology issue on howlers (1998, 19[3]). Throughout, I used the common name "howler monkey" as had been my practice in previous papers. The guest editor of the collection, Margaret Clarke, and the editor of the journal, Russ Tuttle, insisted on "howling monkey." Katharine Milton, another contributor to the issue, and I lobbied hard for "howler monkey," but to no avail. In the end, Russ Tuttle decided that we could use "howling monkey" or "howler," but not "howler monkey." This was his editorial decision.
"Howling monkey" is not based on any preponderance of use or compelling grammatical argument that I can detect. Clarence Ray Carpenter initially used "howling monkey" but switched to "howler monkey" in his later works. To my mind, a "howling monkey" is a monkey that is howling; a "howler monkey" is a monkey capable of howling (and especially notable for this behavior). Common names of other fauna and flora do not reveal a consistent pattern. There are "digger wasps," "song sparrows," and "sting rays." But there are also "chipping sparrows" and "stinging nettles." In fact, relatively few living things are named after their behavior; most are named for appearance (woolly monkey) or after a person (Goeldi's monkey). So the issue comes down to personal preference.
We could solve the problem by using "howler" exclusively. At least one other creature I can think of has a name equivalent to "howler": the "screamer" of South America. It's a bird, but it is not called the "screamer bird." There is no other creature called "howler," so the additional descriptor "monkey" is not really necessary. As for "howling monkey," it sounds grating and strange to me, much as "howler monkey" must sound to the "howling" proponents. - Carolyn M. Crockett, Regional Primate Research Center, University of Washington, Seattle, WA 98195-7330 [e-mail: [email protected]].
* *
Given this assortment of advice, We will exercise our editorial prerogative: Authors may call monkeys by whatever common name makes them happy (except "pigtail" macaques, which has personal connotations for Us...make that "pigtailed", please), so long as they start with the Latin binomial. - The Editors
* * *
Awards Granted
PCWS Conservation Grant
The Primate Conservation and Welfare Society has announced the award of their first annual Primate Conservation Grant. The grantee, Mr. Nicholas Malone, is a graduate student at Central Washington University. His project is "Population Assessment of Displaced Hylobatids and Monitoring the Trade of Primates in Indonesia". Mr. Malone's application was reviewed by the PCWS Board of Directors as well as three outside reviewers. His project is an effort to monitor the sale of primates at various "bird markets" in Java and Bali and to conduct genetic analysis to develop a comparative database for future research. Investigations such as Mr. Malone's, working in collaboration with local people and NGO's, is a vital link in the long-term survival of highly endangered species.
Several other very worthy projects did not receive funding due to PCWS's funding limitations. PCWS plans to continue the small grant project in the years to come and would like to thank all of the PCWS Conservation Auction bidders who made this grant possible, as well as those who applied for funding.
Details of Mr. Malone's project, as well as some amazing photos from Indonesia, will be available in the next issue of Faces From the Forest - the PCWS newsletter. For your copy contact: [[email protected]].
NCRR Primate Grants
Among the more than 1000 new grants awarded in Fiscal Year 2000 by the National Center for Research Resources (NCRR), NIH, were the following:
* To help meet the increased demand for specific-pathogen-free (SPF) macaques for AIDS research, NCRR is funding five SPF rhesus macaque colonies at NCRR-supported primate centers in Massachusetts, Puerto Rico, Texas, Oregon, and California.
* A $2.5 million, five-year grant to the Primate Center Library at the Wisconsin RPRC will fund Coordinated Information Services for Primate Research. Headed by WRPRC Librarian Larry Jacobsen, the services will promote rapid sharing of information among primate researchers.
Applications should be sent on a form to be obtained from the Foundation, and will include: a C.V.; a list of publications of the applicant; the names of two senior scientists whom the applicant has asked to send testimonials to the Secretariat of the Foundation by the date indicted below; and a letter of acceptance from the inviting laboratory.
Fifteen copies of the completed files should be sent to the Secretariat of the Foundation, 195, rue de Rivoli, 7500l Paris, France. Deadline for receipt of applications by the Foundation: March 30, 2001.
2001 International Prize
An International Prize of 300.000 FF is awarded annually to a scientist who has conducted distinguished research in the areas supported by the Foundation. This prize was awarded in 1980 to Professor Andre Leroi-Gourhan; in 1981 to Professor William H. Thorpe; in 1982 to Professor Vernon B. Mountcastle; in 1983 to Professor Harold C. Conklin; in 1984 to R. W. Brown; in 1985 to P. Buser; in 1986 to D. Pilbeam; in 1987 to D. Premack; in 1988 to J. C. Gardin; in 1989 to P. S. Goldman-Rakic; in 1990 to J. Goody; in 1991 to G. A. Miller, in 1992 to P. Rakic; in 1993 to L. L. Cavalli-Sforza; in 1994 to L. R. Gleitman; in 1995 to W. D. Hamilton; in 1996 to C. Renfrew; in 1997 to M. Jouvet; in 1998 to A. Walker; and in 1999 to B. Berlin. Discipline considered for the 2001 prize: Comparative Ethology.
Nominations must be proposed by recognized scientists, and should include: a C.V. of the nominee; a list of his publications; and a summary (four pages maximum) of the research work upon which the nomination is based. Nominations should be sent in 15 copies to the Secretariat of the Foundation, 194, rue de Rivoli, 7500l Paris, France. Deadline for receipt of nominations: October 31, 2001.
La Universidad de la Habana, Sociedad Cubana de Antropología Biológica, Museo Antropológico Montané, Cátedra de Antropología Luis Montané y la Asociación Antropológica de Estudios Primatológicos: Eopithecus de México, les hacen una cordial invitación, al VII Simposio de Antropología Biológica "Luis Montané" y al III Congreso Los Primates como Patrimonio Nacional, durante el 26 de Febrero al 2 de Marzo del 2001,en la Universidad de la Habana, Cuba.
Esperamos contar con gran asistencia en el evento que incluirá las siguientes formas de participación: conferen-cias magistrales por invitación, mesas redondas, comunicaciones orales, y carteles científicos.
Inscripción: El pago de la inscripción al congreso se hará al registrarse en la Universidad de La Habana. Las tarifas son: Titular: US$100, Estudiante: US$50, Acompańante: US$70.
Correspondencia en México para el Congreso Los Primates Como Patrimonio Nacional III, con Braulio A. Hernández Godínez, [e-mail: [email protected]] y en La Habana, Cuba, para el VII Simposio de Antropología Física "Luis Montané", con el Lic. Armando Rangel Rivero, Secretario Ejecutivo Museo Antropológico Montané, Facultad de Biología, Universidad de La Habana, Calle25#455, entre J e I. El Vedado, Ciudad de La Habana 10400, Cuba [e-mail: montaneqcomuh.uh.cu].
Reflexiones sobre la relación hombre-mono en las diferentes culturas. Arturo González-Zamora. Maestría en Manejo de Fauna Silvestre, Instituto de Ecología A.C., Xalapa, Veracruz, México [e-mail: [email protected]].
Desde tiempos ancestrales, la relación que ha existido entre el hombre y la naturaleza ha quedado plasmada en grabados, motivos cerámicos, trabajos en metal, petroglifos, y otras manifestaciones culturales; indicando que para el hombre, la naturaleza no sólo representaba la base material que cubría sus necesidades básicas, sino que constituía un estimulo para sus emociones.
Para el caso de la relación hombre-mono, hubo manifestaciones muy claras de lo anterior. En Europa se pensó por mucho tiempo que entre el hombre y el mono solo existía un paso. Inclusive, muchas veces se le vió al mono como un hombre degenerado, víctima del pecado y sobre todo con una connotación de lujuria, condición negativa, que provocaba su degeneración. Sin embargo, esta impresión cambiaba según el proyecto cultural de la época.
Las relaciones entre el hombre y el mono han sido mediadas por la cultura. Con el paso del tiempo, el hombre ha ido construyendo diversos conceptos sobre este animal, especialmente tomándolo como el referente más cercano a él. Por ejemplo, en el aspecto mitológico, donde en el "origen" del mundo, los monos pertenecen al período de la humanidad intermedio entre los dioses, semidioses y hombres que narran estos mitos.
Para el resto de Mesoamérica la situación fue parecida y la presencia de monos en el contexto cultural de estos pueblos se llenó de fantasías y leyendas. Como ejemplos tenemos los relatos del Popol Vuh y Chilam Balam entre otros. En cuanto al Popol Vuh, es muy conocido el episodio en que Hunbatz y Hunchuen, excelentes músicos y artesanos, se convirtieron en monos, el primero en un "saraguato" o mono aullador (Gen. Alouatta) y el segundo en mono arańa (Gen. Ateles), debido a los celos que estos tenían por sus hermanos Hunahpú e Ixbalanqué. Como se mencionó, en muchos de estos lugares, el mono es visto como un animal muy parecido al hombre, el cual es muchas veces objeto de burla pues asemejan al propio hombre como un animal travieso y flojo.
Es claro que la concepción del mono, más allá de ser un animal cualquiera del medio natural, se ha conservado debido a una base de simbolismos tanto occidentales como mesoamericanos. Muchos de estos simbolismos son coincidentes en su carácter dinámico, (propio de cualquier símbolo), y continuamente se transforman para seguir reconstruyendo los conceptos humanos frente a los oríge-nes de su naturaleza. En síntesis, para la mentalidad europea, el mono era visto como un elemento que representaba la parte animal del hombre, lo que en sus orígenes, provocaba problemas bajo las ideas de la iglesia. En cambio, para la mentalidad mesoamericana el mono era alguien cercano y cotidiano, el cual les traía recuerdos sobre su pasado o rasgos muy relacionados con ellos. A pesar de lo anterior, en la actualidad las cosas han cambiado para los primates silvestres. Los pobladores que habitan cerca de fragmentos de selva en donde aún existen grupos de primates saben que ante la fragmentación del hábitat, "ahora ya no hay tantos como antes", y que quizás pronto pudieran dejar de verlos y oírlos. Así, muchas de estas comunidades se han organizado a fin de retomar ese pensamiento positivo sobre la fauna silvestre que les rodea, especialmente sobre los monos silvestres.
* * *
Positions Available
Clinical Lab Animal Veterinarian/Assistant Director
BIOQUAL, Inc. seeks a clinical veterinarian for the Department of Laboratory Animal Medicine (DLAM). BIOQUAL is an established contract research laboratory with a national reputation in infectious disease and related research, using a variety of laboratory animal species with an emphasis on nonhuman primates. Our AAALAC International-accredited facilities are located in Rockville, Maryland, conveniently situated between the National Institutes of Health and the Frederick Cancer Research and Development Center.
This position reports to the Vice President of the Division of Primate Biology and Medicine/Director, DLAM, and is responsible for enhancements to, and expansion of, the animal care and use program. Responsibilities include provision of clinical care, surgical support, and preventive medicine programs for approximately 600 nonhuman primates in addition to rabbits and rodents; consultation and collaboration with NIH scientists for animal model and protocol development in support of vaccine and discovery research; membership on the BIOQUAL ACUC; and development and training of technical staff.
Minimal qualifications for this position are a DVM/VMD degree and license to practice veterinary medicine in at least one state; completion of an ACLAM-recognized residency/postdoctoral training program in laboratory animal medicine; and either board certification by ACLAM or eligibility to take the ACLAM certifying examination in 2001. Strong interpersonal and communi-cation skills are essential, and experience with nonhuman primates is preferred.
BIOQUAL is located in the Maryland suburbs of the Washington, D.C., Metropolitan area. Opportunities exist for collaborative research, and a local ACLAM board-preparation study group is available for interested candidates. BIOQUAL offers a competitive compensation and benefits package, and is an Equal Opportunity Employer. Persons interested in this position should submit a letter of application, a CV, and the names, telephone numbers and addresses of three professional references to: Dr. Marisa St. Claire, Vice President, Division of Primate Biology and Medicine, BIOQUAL, Inc., 2501 Research Blvd, Rockville, MD 20850 [301-948-9565].
Primatology/Psychology - Bucknell University
Bucknell University's Department of Psychology and Program of Animal Behavior seek a primatologist for a tenure-track appointment as either assistant or associate professor (with a maximum of three years credit toward tenure). Preference will be given to candidates who conduct noninvasive or minimally invasive research with long-term socially-maintained groups. Indoor/outdoor facilities exist to house several species of primates (but not great apes. Research must have potential to attract and involve undergraduate and master's students. Teaching assignment includes core courses in both psychology and animal behavior at introductory, intermediate, and advanced levels of the curriculum, as well as courses in the general education curriculum. The successful candidate will share responsibility for three core courses in animal behavior. Other teaching assignments depend on the candidate's area of expertise, but may include courses in cognition, perception, learning, and physiological, developmental, social, personality, or abnormal psychology.
The Department is also seeking a psychologist in the area of animal learning for an entry-level tenure-track appointment at the rank of assistant professor. Required teaching includes courses in basic learning processes at the intermediate and advanced levels of the curriculum, participation in a team-taught introductory course, and an occasional general education course. Ability to teach statistics is desirable. Opportunity exists to create new courses according to interest and expertise. Research must address basic learning processes primarily or exclusively in non-primate animals.
Applicants for either position must have the PhD or be able to complete it by September, 2001. Send CV, materials demonstrating evidence of success or promise in teaching and research, and three letters of reference to: Primatology Search Committee (or Animal Learning Search Committee), Department of Psychology, Bucknell University, Lewisburg, PA 17837. Bucknell encourages applications from women and members of minority groups (EEO/AA). Contact K. A. Daubman, Chair [570-577-1200; e-mail: [email protected]], for more information about either position.
Animal Behavior/Biology - Bucknell University
Applications are invited for a tenure-track position at the assistant professor level (a maximum of three years may count toward tenure) beginning in August, 2001. A PhD is required. Applicants should be working in the area of vertebrate behavior and have expertise in population, quantitative, and evolutionary genetics. Teaching responsibilities will include introductory and advanced courses in animal behavior/biology, an upper-division course in one's area of expertise, and supervising undergraduate research. A cover letter addressing teaching philosophy and research goals, a CV, and three letters of recommendation should be submitted to: Dr. Wayne McDiffett, Chair, Biology Department, Bucknell University, Lewisburg, PA 17837 [570-577-1124; fax: 570-577-3537; e-mail: [email protected]]. For more information, see <www.bucknell.edu/department/biology>.
Gorilla Keeper - Georgia
Gorilla Haven is seeking an experienced ape keeper for the position of Assistant to the Director of Animal Care. The successful applicant, preferably having worked with gorillas, will be familiar with accepted husbandry practices and standards and have the maturity to supervise volunteers and make decisions responsibly. Duties will be varied and include animal care and exhibit cleaning plus overseeing the daily management of the facility in the absence of the Director of Animal Care. Gorilla Haven is a new facility and offers the opportunity to meet the challenges of growing with the planned expansion to four habitats (totaling 25 acres) over 20 years. The position will be available in April, 2001. Salary will be commensurate with experience. Please send resume and references to: Pete Halliday, Project Manager/Director of Animal Care, Gorilla Haven, P.O. Box 210, Morganton, GA 30560 [e-mail: [email protected]].
Biological Anthropologist - Florida
The University of West Florida, Department of Sociology and Anthropology, is seeking a Biological Anthropologist for a tenure-track Assistant Professor position beginning in August, 2001, to provide the Physical Anthropology component of a four-field undergraduate program and to participate in the development of a Master's program in Anthropology. The successful applicant must have a PhD in physical/biological anthropology at the time of the appointment. We are seeking a colleague who is broadly trained and interested in one or more of the following areas: paleodemography, human ecology, bioarcheology, skeletal biology, health and disease, and human variation. Research area is open, but must link to existing archeology and applied anthropology programs. Responsibilities include teaching general anthropology, basic courses in physical/biological anthropology, human evolution, and elective courses in area of specialization. The University of West Florida is a regional state university with almost 9000 students. The department has a very active historical archeology program, with a master's program in the Department of History. Interested candidates should send a letter of application detailing teaching and research interests, CV, and names of three references to Judith A. Bense, Chair, Dept of Sociology and Anthropology, University of West Florida, 11000 University Parkway, Pensacola, FL 32514. Applications must be received by February 16, 2001. Salary is negotiable. UWF is an affirmative action/ACCESS/equal opportunity employer. For more information, contact Dr. Bense [[email protected]]; or visit
Life Sciences Job Resource
BiologyJobs, at <www.BiologyJobs.com> , is a resource for job seekers and employers who are specifically interested in the life sciences. Biology job categories in the database include Agriculture, Anatomy/Physiology, Bioinformatics/Biotechnology, Botany, Cell Biology, Conservation, Ecology, Environmental Science, Genetics, Marine Science, Medicine, Natural Resources, Pharmacology, Toxicology, Veterinary Medicine, and Zoology.
Biology job seekers may search their database for jobs, register, and use their on-line tools to create a resume, post an existing resume, set up a "job agent", and receive e-mails with matching job opportunities in the Biological Sciences. Membership for registered job seekers is free.
Employers who register can post vacancies, set up "resume matching agents", and view resumes on-line. For a limited time, this service to employers is free.
* * *
Continuing education for laboratory animal technicians will be offered at the SUNY Delhi, College of Technology's Veterinary Science Program for 2001. The following is a list of courses offered to date; more may be added later. For further information contact Jackie Howard, 136 Farnsworth Hall, SUNY, Delhi, NY 13753 [607-746-4305; e-mail: [email protected]]. For registration, contact Amy Reynolds, Center for Continuing Education and Corporate Services, 315 Bush Hall, SUNY, Delhi, NY 13753 [607-746-4545].
Sunday, May 20 to Friday, May 25, 2001: Laboratory Animal Technologist Review, designed to aid the qualified candidate in preparing for the national AALAS Certification Examination for Laboratory Animal Technologists. The candidate must submit an application to AALAS according to protocol. Study material and accommodation materials will be mailed to you when your registration is received at SUNY Delhi. Instructor: Kenneth Pyle, LATG, and Staff; Continuing Education Units (CEU): 6.0; Fee: $450 (materials included); Registration deadline: May 4, 2001.
Wednesday, May 30 or Thursday, May 31, 2001: Primatology, an introduction to the biology and husbandry of nonhuman primates used in biomedical research, including hands-on experience in catching and restraining primates, administering drugs and compounds, and collecting samples; plus a description of different housing regimes, anesthesia, TB testing, and physiological data collection (i.e., ECG, blood pressure, and pulse oximetry). Instructor: Kenneth Pyle, LATG; CEU: 0.7; Fee: $200; Registration deadline: May 16, 2001. Registrants must have evidence of a negative TB test (P.P.D.) within the preceding 12 months.
Friday, June 1, 2001: Veterinary Dentistry: "Brush up" your skills or learn the basics of dental prophylaxis and periodontal therapy. Ultrasonic and Rotosonic techniques for felines, canines, and primates will be used. Instructor: Cheryl Peletz, LVT; CEU: 0.4; Fee: $200; Registration deadline: May 18, 2001.
Friday, June 8, 2001: Basic Lab Animal Biomethodology, including a review of basic biomethodology involving laboratory rodents. Procedures will include restraint, oral and injectable drug administration including anesthetics, implantation of osmotic pumps, blood collection procedures, surgical preparation techniques, and methods of animal identification including microchip implantation. Instructor: Alan Franks, LATG; CEU: 0.6; Fee: $200; Registration deadline: May 25, 2001.
Primate Behavior and Ecology Program - Panama
Florida State University (FSU)-Panama is offering a 4-week, 7-semester-hour Primate Behavior and Ecology Program this summer from June 22nd to July 21st. As a part of the training, students will conduct directed research projects on the endangered Panamanian tamarin (Saguinus geoffroyi) and live at the Primate Refuge and Sanctuary of Panama (PRSP). The PRSP is located on the Atlantic side of Panama in a cluster of beautiful islands in Gatun Lake. D. R. Rasmussen, Ph.D., Director of the Primate Refuge and Sanctuary of Panama, and Associate Professor at FSU-Panama branch, will be leading the program and teaching the courses.
The following courses will be taught as a part of the Primate and Behavior Ecology Program:
The cost of the program is $2795, which includes tuition, books, food and lodging, and travel within (but not to) Panama. The deadline for the receipt of a completed application form is January 19, 2001. For information, call FSU at 850-644-3272 or 800-374-8581, or see <www.fsu.edu/~cppanama/ipsp/Program.htm> or <www.international.fsu.edu> .
Field Course in Animal Behavior - Georgia and Africa
Georgia Tech and Zoo Atlanta present a four-week field course in animal behavior from June 18 - July 15, 2001, for 6 semester credits in psychology. The course takes place in Atlanta (4 days at the zoo), South Africa (3 days), and Kenya (20 days).
The priority of the course is to provide intensive instruction in animal behavior and observational methodology. In Atlanta, students are taught data collection methods and the behavior of East African mammals from a variety of zoo, academic, and field scientists. Students then use this information in Africa to conduct daily observations on a variety of species. Observations are conducted in both national parks and private reserves, which permits students to compare the behavior of a single species across settings. Students also use the comparative psychology approach to examine behavior across closely related taxa. Daily observations are supplemented by readings, discussions, and lectures by field scientists. The course also places a strong emphasis on conservation, and students read and discuss many of the issues related to conservation in Africa.
Spaces are limited to 12 students. The approximate cost of the course is $6100, and applications must be mailed by February 15, 2001 (early applications are encouraged). For additional information and an application, contact: Dr. Tara Stoinski, Coordinator of Primate Research, Zoo Atlanta, 800 Cherokee Ave, Atlanta, GA 30315 [404-624-5826; e-mail: [email protected]].
Research in the Biology of Aging
The American Federation for Aging Research (AFAR) is accepting applications for scholarships for research in the biology of aging. This program provides PhD and MD students the opportunity to conduct a 3-month research project under the auspices of a mentor. Each scholarship carries an award of $5000. Deadline for receipt of application is February 26, 2001. Contact AFAR, 1414 Ave. of the Americas, 18th Floor, New York, NY 10019 [212-752-2327; fax: 212;832-2298; e-mail: [email protected]]; or see <www.afar.org>.
Summer Apprenticeship: Chimpanzees and ASL
The Chimpanzee and Human Communication Institute (CHCI) is taking applications for their 10-week Summer Apprentice Program, to be held June 17 to August 24, 2001. Graduates, undergraduates, and postgraduates from various academic backgrounds are encouraged to apply. The research at CHCI involves a group of five chimpanzees who use the signs of American Sign Language (ASL). The chimpanzees reside at Central Washington University in Ellensburg, Washington, in a large, state-of-the-art facility.
Apprentices clean enclosures, prepare meals and enrichment, make observations of the chimpanzees, and participate in a research project. The first week is intensive training in laboratory jobs and chimpanzee behaviors. Apprentices are trained in humane care and research techniques. After several weeks each apprentice becomes more autonomous and has responsibilities in the research project.
The program fee is $1700 and there is a nonrefundable $25 application processing fee. This does not include housing and transportation. Inexpensive housing is available on campus. A course in ASL is highly recommended. For more information see <www.cwu.edu/~cwuchci/apprentice.html> ; or contact Mary Lee Jensvold, CHCI, CWU, Ellensburg, WA 98926 [[email protected]]. Application deadline is March 26, 2001.
* * *
International Veterinary Information Service (IVIS) is a not-for-profit organization dedicated to providing clinical information to veterinarians, veterinary students and animal health professionals worldwide, using Internet technologies. The IVIS Web site, <www.ivis.org/> , provides free access to original, up-to-date publications organized in edited electronic books, proceedings of veterinary meetings, short courses, continuing education (lecture notes, manuals, autotutorials and interactive Web sites), an international calendar of veterinary events, image collections, and more, with the help of private and corporate sponsors. Contact IVIS at P.O. Box 4371, Ithaca NY 14852 [e-mail: [email protected]].
New Video Available
A new video, Working Safely with Nonhuman Primates, is available through NIH's Office of Animal Care and Use, Intramural Research Program. The video covers nonhuman primate behavior and the proper use of personal protective equipment. It is currently online at: <grants.nih.gov/grants/olaw/PrimateVideo.cfm> .
Introducing PIN WEB LINKS
PIN WEB LINKS is a new feature on Primate-Science. This series will feature selected Web sites which relate to primate research, conservation, education, and animal care, with synopses of their contents. PIN <www.primate.wisc.edu/pin> currently links to 1,500 pages of information about primates, with an additional 30-40 new links being reviewed for inclusion each month. All sites presented in this series can be bookmarked or found through a PIN search. Depending on your mailer, you should be able to link directly from PIN WEB LINKS to the site(s) being reviewed. If you would like to suggest a site to be featured in this series, contact Matt Hoffman [e-mail: [email protected]], Acting Internet Services Librarian at the WRPRC.
E-mail News Groups
ZooNews Digest is a weekly compilation of links to Zoo News from all over the world. The Digest includes short articles, vacancies, and related information. Anyone can subscribe to this e-zine; send an e-mail to: [[email protected]]. Leave the subject and message area blank.
Zoo Biology is a discussion, question, and answer group dealing with the diverse range of Zoo Sciences. The Zoo Biology Group is concerned with all disciplines involved in the running of a zoological garden: captive breeding, husbandry, cage design and construction, diets, enrichment, management, record keeping, etc., It is specifically a forum for professional zoo staff. Exceptions may be considered in the case of students or allied professions. Do be prepared to receive up to 30 messages per day. To subscribe, send an e-mail to: [[email protected]]. Leave the subject and message area blank. At the same time please send a separate e-mail to [[email protected]], telling your work area and interests. Failure to do this will mean that your subscription will not be processed. All details will be kept strictly confidential.
To learn more about these zoo news groups, please visit <www.egroups.com/group/zoonewsdigest> .
More Interesting Web Sites
* 2000 IUCN Red List of Threatened Species: <www.redlist.org/>
* Animal Protection Institute exotic pet campaign: <www.exotic-pets.org>
* Australian primate welfare conference (Proceedings): <www.lisp.com.au/~primate/index.htm>
* AWIC animal behavior and welfare links : <www.nal.usda.gov/awic/links/links.htm>
* Conservation International: <www.conservation.org/>
* Database of postgraduate training programs in lab animal medicine: <www.aslap.org/career.htm>
* Duke University Primate Center: <www.duke.edu/web/primate/>
* Netherlands Centre Alternatives to Animal Use: <prex.las.vet.uu.nl/nca/>
* The Norwegian Reference Centre for Laboratory Animal Science & Alternatives: <oslovet.veths.no/NORINA/>
* Parasitology Online: <www.parasitology-online.com>
* Tropical Disease Research publications: <www.who.int/tdr/publications/publications/>
* * *
The American College of Laboratory Animal Medicine Foundation Committee members are pleased to announce our solicitation of research proposals in Laboratory Animal Science and Medicine. The deadline for pre-proposals is February 6, 2001. The 2001 Foundation awards will be distributed early in July. (See Grant Application Instructions and Tips on the ACLAM web site: <www.aclam.org>).
The mission of the ACLAM Foundation is to expand the body of knowledge in the fields of laboratory animal science and medicine through funding research grants. The ACLAM Foundation has developed a two-tiered system for competitive review of grant proposals. Submission of a pre-proposal is followed by a request for a detailed proposal to selected applicants. The Foundation will focus its funding on research related to laboratory animal science and medicine in the following five broad areas: * analgesia/anesthesia * animal behavior/well-being * diagnostics/diseases of laboratory animals * laboratory animal husbandry * toxicology (refining models or techniques in toxicology).
Successful grantees are encouraged to publish their results in peer-reviewed journals, and must agree to provide summary research reports in lay language suitable for inclusion in ACLAM Foundation communications, fund -raising solicitations, and the ACLAM Newsletter.
For more information, see the Web site above, or contact Dr. Martin Morin, Chairman, ACLAM Foundation, 208 Byford Dr., Chestertown, MD 21620 [410-810-1870; fax: 410-810-1869; e-mail: [email protected]].
National Research Service Awards for Senior Fellows
The Congress of the United States enacted the National Research Service Act (NRSA) Program in 1974 to help ensure that highly trained scientists will be available in adequate numbers and in appropriate research areas to carry out the nation's biomedical and behavioral research agenda. Under this congressional authority, the National Institutes of Health (NIH) awards NRSA senior fellowships to experienced scientists who wish to make major changes in the direction of their research careers or who wish to broaden their scientific background by acquiring new research capabilities. These awards will enable individuals with at least seven years of research experience beyond the doctorate, and who have progressed to the stage of independent investigator, to take time from regular professional responsibilities for the purpose of receiving training to increase their scientific capabilities. In most cases, this award is used to support sabbatical experiences for established independent scientists. This program is not designed for postdoctoral level investigators seeking to prove their research potential prior to independence. Potential applicants are encouraged to discuss their plans with a contact from one of the Institutes or Centers.
By the time of award, individuals must be citizens or noncitizen nationals of the United States or have been lawfully admitted to the United States for permanent residence. Individuals on temporary or student visas are not eligible. Before submitting an application, an individual must arrange for acceptance by an institution and sponsor. The applicant's sponsor should be an active investigator in the area of the proposed research who will directly supervise the candidate's research during the period of the fellowship. Individuals requesting foreign-site training rather than a domestic site must show in the application that the foreign institution and sponsor offer unique opportunities that are not currently available in the United States. Only if there is clear scientific advantage will foreign training be supported.
Application receipt dates are April, August, and December 5. Written and telephone inquiries concerning this program announcement are encouraged, especially during the planning phase of the application. For information on whom to contact, call NIH Office of Extramural Research: Walter Schaffer [301-496-9743; e-mail: [email protected]].
Behavioral Science Award For Rapid Transition
The National Institute on Drug Abuse (NIDA) seeks to facilitate the entry of beginning investigators into the field of behavioral science research. NIDA invites newly independent investigators to submit applications for small-scale, exploratory, or pilot research projects related to NIDA's behavioral sciences mission.
To be eligible for a Behavioral Science Award For Rapid Transition (B/START) award, the proposed principal investigator must be independent of a mentor at the time of award, but be at the beginning stages of her/his research career. No B/START award will be made to individuals in training status. The proposed principal investigator may not have been designated previously as principal investigator on a National Institutes of Health (NIH) or federally-supported research project or component (except a dissertation research grant) or have received similar support from another federal agency. Because this program is targeted at newly independent investigators, individuals who are more than five years beyond training status should consider other mechanisms of support.
To be appropriate for a B/START award, research must be primarily focused on behavioral processes and research questions. Thus, research on genetic, neural, or other biological processes would be appropriate for a B/START award only if its overall emphasis were on understanding behavioral processes or outcomes. All areas of behavioral research related to drug abuse are sought under this announcement including animal and human basic behavioral and cognitive research which is aimed at elucidating underlying behavioral and cognitive mechanisms, determinants, and correlates of drug abuse, and characterizing the consequences of drug abuse and addiction.
Direct inquiries regarding programmatic issues to: Cora Lee Wetherington, Div. of Neuroscience and Behavioral Research, NIDA, 6001 Executive Blvd, Rm 4282, MSC 9555, Bethesda, MD 20892-9555 [301-435-1319; fax: 301-594-6043; e-mail: [email protected]].
* * *
A Species Information Service (SIS) has been launched at the World Conservation Union (IUCN) Congress in Amman. It will be the world's most complete source of scientific and management information on species with data from 7,000 volunteers and 120 specialists representing almost every country in the world.
This IUCN database will consist of geographical and conservation information and population data that will be regularly updated. It will be available to conservationists, scientists, people who manage species, and members of IUCN, an umbrella organization of environmental institutions and agencies around the world.
According to the Red List, released every four years by the IUCN, as many as a quarter of mammals, reptiles, and freshwater fish, 20 percent of amphibians, and 12 percent of birds are threatened. The wandering albatross, sea otters, Chinese alligators, Iberian lynx and Philippine eagle are among the species which may face the same fate as the dinosaurs. Since the last Red List was published in 1996, 136 new species have been added. - From an October 6 Reuters report by Patricia Reaney, posted to Alloprimate
Material Exchanges
Hope Walker, of the Primate Conservation and Welfare Society, notes that many museums and other institutions exchange materials in order to mount temporary educational exhibits. She asks whether any institutions are interested in this sort of exchange in the fields of primatology. "If not, do any of you have excess primate/jungle/forest-related materials that you would be willing to donate to our Primate Education Center?"
She suggests a program that would allow exhibits to be changed at least four times per year. This would allow the public to come and see new things, and the institutions to promote different concepts. "We could loan each other materials, which would save all of us money, and allow the public to learn more." If anyone is interested in this, please contact Hope at P.O. Box 2101, Port Townsend, WA 98368 [e-mail: [email protected]]. "I'd be very keen to begin discussions. We have enough materials to put together a bushmeat-related exhibit (small but effective), which would include photos, posters, wire snares from Uganda, etc."
Primate-Enrichment.Net
Janette Wallis and Deborah Hartley, of the University of Oklahoma Health Sciences Center, announce a new Web site devoted to sharing ideas for nonhuman primate enrichment. The site is meant to be a "how-to" site only (i.e., it does not replace or compete with discussion lists or the scientific literature). Think of it as what Martha Stewart would create if she were devoted to the care and well-being of nonhuman primates.
Take a look at the site and consider offering your ideas for primate enrichment. CAUTION: The site is still in developmental stages, so don't be worried that not much is there yet. We want this to be a shared site (i.e., not just our own ideas!). We anticipate several ideas rolling in soon from individuals we've already contacted. So, check it out: <primate-enrichment.net>.
Free Software for Studying Behavior
Dan Blumstein, Chris Evans, and Janice Daniel have made available Jwatcher, which they call "a powerful and free tool for the quantitative analysis of behavior. It can be used to address any theoretical problem that requires a complex sequence of actions to be scored by a human observer." JWatcher can be used simply as an event recorder which logs the time at which keys are pressed. It was designed primarily for focal sampling, in which a single subject is observed continuously. The real power of JWatcher is, however, in its analysis routines. These calculate time budgets and provide statistics about the duration of behavioral states and the intervals between them.
JWatcher is currently free, available for downloading at <galliform.psy.mq.edu.au/jwatcher/>. "We suggest reading the 33-page user manual before downloading the program. On-line help will guide you through the early stages of using JWatcher. Feel free to use it, share it with others, and teach with it. We're seeking funding for future program development and would be very interested in your suggestions for additional features."
Books
* Primate Conservation Biology. G. Cowlishaw & R. Dunbar. Chicago: University of Chicago Press, 2000. [Price: $75 (cloth); $27(paper)]
* Natural Conflict Resolution. F. Aureli & F. B. M. De Waal (Eds.). Berkeley: University of California Press, 2000. [Price: $65 (cloth) $24.95 (paper)]
* Mahale: A Photographic Encounter with Chimpanzees. A. Hofer, M. A. Huffman, & G. Ziesler. Forward by J. Goodall. New York: Sterling Publishing, 2000. [Price: $24.95]
* Animal Biology and Care. S. E. Dallas. Ames, Iowa: Iowa State University Press, 2000. [Price: $26.95]
* Primate Males: Causes and Consequences of Variation in Group Composition. P. M. Kappeler (Ed.). Cambridge: Cambridge University Press, 2000 [Price: $85.00 (cloth); $37.95 (paper)]
* Biology, Brains, and Behavior: The Evolution of Human Development. S. T. Parker, J. Langer, & M. L. McKinney (Eds.). Santa Fe, NM: School of American Research Press, 2000. [Price: Cloth $60; Paper $24.95]
* Evolution and Human Behavior: Darwinian Perspectives on Human Nature. J. Cartwright. Cambridge, MA: MIT Press, 2000. [Price: $24.95]
* Encyclopedia of Human Evolution and Prehistory (2nd ed.). E. Delson, I. Tattersall, J. A. Van Couvering, & A. S. Brooks (Eds.). New York & London: Garland Publishing, Inc., 2000. [Price: $175]
* The Orangutans: Their Evolution, Behavior, and Future. G. Kaplan and L. Rogers. Cambridge, MA: Perseus Publishing, 2000. [Price: $23]
* Biomedical Imaging, Visualization, and Analysis. R. A. Robb. Somerset, NJ: Wiley-Liss, 2000. [Price: $125]
Audiovisual Material
* Working Safely with Nonhuman Primates. NIH Office of Animal Care and Use, 1999. [May be viewed free at <grants.nih.gov/grants/olaw/PrimateVideo.cfm> or contact Office of Laboratory Animal Welfare, NIH, RKL1, Suite 1050, MSC 7982, 6705 Rockledge Dr., Bethesda, MD 20892-7982]
Booklets
* NCRR Highlights, 1998-1999. Bethesda, MD: National Center for Research Resources. NIH Publ. No. 97-2309, 2000. [NCRR, NIH, 1601 Research Blvd, Rockville, MD 20850-3173; e-mail: [email protected]]
Magazines and Newsletters
* Primate Eye, October, 2000, No. 72. [Bill Sellers, Primate Society of Great Britain, Dept of Biomed. Science (Anatomy Sect.), Univ. of Edinburgh, Hugh Robson Bldg, George Sq., Edinburgh EH8 9XD, Scotland]
Reports
* Primate Report, July, 2000, 57. [German Primate Center (DPZ), Kellnerweg 4, 37077 Göttingen, Germany]
Supplements
* Current Primate Field Studies. Primate Eye, October 2000, No. 72 (supplement). [Bill Sellers, Primate Soc. of Great Britain, Dept of Anatomy, Univ. of Edinburgh, Med. School, Teviot Pl., Edinburgh EH8 9AG, Scotland]
Anatomy and Physiology
* Promiscuity and the primate immune system. Nunn, C. L., Gittleman, J. L., & Antonovics, J. (Department of Biology, Gilmer Hall, University of Virginia, Charlottesville, VA 22904-4328 [e-mail: [email protected]]). Science, 2000, 290, 1168-1170.
* Preparental hormone levels and parenting experience in male cotton-top tamarins, Saguinus oedipus. Ziegler, T. E. & Snowdon, C. T. (Wisconsin RPRC, Univ. of Wisconsin, 1220 Capitol Ct, Madison, WI 53715 [e-mail: [email protected]]). Hormones and Behavior, 2000, 38, 159-167.
Animal Models
* Vaccination with Tat toxoid attenuates disease in simian/HIV-challenged macaques. Pauza, C. D., Trivedi, P., Wallace, M., Ruckwardt, T. J., Le Buanec, H., Lu, W., Bizzini, B., Burny, A., Zagury, D., & Gallo, R. C. (Univ. of Wisconsin, Dept of Pathology, 1300 University Ave, Madison, WI 53706 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, USA, 2000, 97, 3515-3519.
* Lymphoid organs as a major reservoir for human T-cell leukemia virus type 1 in experimentally infected squirrel monkeys (Saimiri sciureus): Provirus expression, persistence, and humoral and cellular immune responses. Kazanji, M., Ureta-Vidal, A., Ozden, S., Tangy, F., de Thoisy, B., Fiette, L., Talarmin, A., Gessain, A., & de Thél, G. (Lab. de Rétrovirologie, Inst Pasteur de la Guyane, B.P. 6010, 23, Av. Pasteur, 97300 Cayenne, French Guiana [e-mail: [email protected]]). Journal of Virology, 2000, 74, 4860-4867.
* Viremia control following antiretroviral treatment and therapeutic immunization during primary SIV251 infection of macaques. Hel, Z., Venzon, D., Poudyal, M., Tsai, W.-P., Giuliani, L., Woodward, R., Chougnet, C., Shearer, G., Altman, J. D., Watkins, D., Bischofberger, N., Abimiku, A., Markham, P., Tartaglia, J. & Franchini, G. (G. F., NCI, Basic Research Lab., 41/D804, Bethesda, MD 20892 [e-mail: [email protected]]). Nature Medicine, 2000, 6, 1140-1146.
* Control of viremia and prevention of clinical AIDS in rhesus monkeys by cytokine-augmented DNA vaccination. Barouch, D. H., Santra, S. Schmitz, J. E., Kuroda, M. J., Fu, T.-M., Wagner, W., Bilska, M., Craiu, A., Zheng, X. X., Krivulka, G. R., Beaudry, K., Lifton, M. A., Nickerson, C. E., Trigona, W. L., Punt, K., Freed, D. C., Guan, L., Dubey, S., Casimiro, D., Simon, A., Davies, M.-E., Chastain, M., Strom, T. B., Gelman, R. S., Montefiori, D. C., Lewis, M. G., Emini, E. A., Shiver, J. W., & Letvin, N. L. [Dept of Med., Beth Israel Deaconess Medical Center, Harvard Medical School, 330 Brookline Ave, Boston, MA 02215 [e-mail: [email protected]]. Science, 2000, 290, 486-492.
* Control of SIV rebound through structured treatment interruptions during early infection. Lori, F., Lewis, M. G., Xu, J., Varga, G., Zinn, D. E. Jr., Crabbs, C., Wagner, W., Greenhouse, J., Silvera, P. Yalley-Ogunro, J., Tinelli, C., & Lisziewicz, J. (J. L., RIGHT, Medical-Dental Bldg SW307, 3900 Reservoir Rd, NW, Washington, DC 20007 [e-mail: [email protected]].). Science, 2000, 290, 1591-1593.
* Protection against Plasmodium falciparum malaria in chimpanzees by immunization with the conserved pre-erythrocytic liver-stage antigen 3. Daubersies, P., Thomas, A. W., Millet, P., Brahimi, K., Langermans, J. A. M., Ollomo, B., Ben Mohamed, L., Slierendregt, B., Eling, W., Van Belkum, A., Dubreuil, G., Meis, J. F. G. M., Guérin-Marchand, C., Cayphas, S., Cohen, J., Gras-Masse, H., & Druilhe, P. (P. D., Unité de Parasitologie Biomédicale, Institut Pasteur, 28 rue du Dr Roux, 75015 Paris, France [e-mail: [email protected]]. Nature Medicine, 2000, 6, 1258-1263.
* Neurodegeneration prevented by lentiviral vector delivery of GDNF in primate models of Parkinson's disease. Kordower, J. H., Emborg, M. E., Bloch, J., Ma, S. Y., Chu,Y., Leventhal, L., McBride, J., Chen, E.-Y., Palfi, S., Roitberg, B. Z., Brown, W. D., Holden, J. E., Pyzalski, R., Taylor, M. D., Carvey, P., Ling, Z.-D. Trono, D., Hantraye, P., Déglon, N., & Aebischer, P. (Dept of Neurological Sciences, Rush Presbyterian-St. Luke's Medical Center, Chicago, IL 60612 [e-mail: [email protected]]). Science, 2000, 290, 767-773.
* Real-time prediction of hand trajectory by ensembles of cortical neurons in primates. Wessberg, J., Stambaugh, C. R., Kralik, J. D., Beck, P. D., Laubach, M., Chapin, J. K., Kim, J., Biggs, S. J.,M. A., & Nicolelis, M. A. L. (M. A. L. N., Dept of Neurobiology, Duke Univ., Durham, NC 22710 [e-mail: [email protected]]). Nature, 2000, 408, 361-365.
* Integration of target and body-part information in the premotor cortex when planning action. Hoshi, E. & Tanji, J. (J. T., Dept of Physiology, Tohoku Univ. School of Med., 2-1 Seiryo-cho, Aoba-ku, Sendai 980-8575, Japan [e-mail: [email protected]]). Nature, 2000, 408, 466-470.
Behavior
* Spontaneous use of tools by wedge-capped capuchin monkeys (Cebus olivaceus). Urbani, B. (Apto 47.028, Caracas 1041-1, Venezuela [e-mail: [email protected]]). Folia Primatologica, 1999, 70, 172-174.
* Intra-troop affiliative relationships of females with newborn infants in wild ring-tailed lemurs (Lemur catta). Nakamichi, M. & Koyama, N. (Lab. of Ethological Studies, Fac. of Human Sciences, Osaka Univ., Suita, Osaka 565-0871, Japan [e-mail: [email protected]]). American Journal of Primatology, 2000, 50, 187-203.
* Pituitary-adrenocortical responses to the first dyadic encounters in male rhesus monkeys: Effect of dominance relationship. Kimura, K., Shimizu, K., Hayashi, M., Ishikawa, T., & Ago, Y. (Toukatsu Clinic, 4-136 Shinmatsudo, Matsudo City, Chiba, 270, Japan). American Journal of Primatology, 2000, 50, 247-256.
* Feeding tactics in juvenile rhesus monkeys (Macaca mulatta). Peláez, F., Gil-Burmann, C., & Sánchez, S. (Area de Psicobiología, Fac. De Psicología, Univ. Autónoma de Madrid, 28049-Madrid, Spain [e-mail: [email protected]]). American Journal of Primatology, 2000, 51, 147-152.
* Vertebrate predation by sympatric tamarins, Saguinus mystax and Saguinus fuscicollis. Heymann, E. W., Knogge, C., & Tirado Herrera, E. R. (Dept of Behavioral Research and Ecology, German Primate Center, Kellnerweg 4, D-37077 Göttingen, Germany [e-mail: [email protected]]). American Journal of Primatology, 2000, 51, 153-158.
* Observations on the behavior of gibbons (Hylobates leucogenys, H. gabriellae, and H. lar) in the presence of mirrors. Ujhelyi, M. Merker, B., Buk, P., & Geissmann, T. (Semmelweis Univ. of Med., Inst of Behavioural Sciences, H-1089 Budapest, VIII. Nagyvárad tér 4, Hungary [e-mail: [email protected]]). Journal of Comparative Psychology, 2000, 114, 253-262.
* Development of hand preferences in marmosets (Callithrix jacchus) and effects of aging. Hook, M. A. & Rogers, L. J. (Dept of Vet. Sciences, UTMDACC, Rt 2, Box 151-B1, Bastrop, TX 78602 [e-mail: [email protected]]). Journal of Comparative Psychology, 2000, 114, 263-271.
* Naive chimpanzees' (Pan troglodytes) observation of experienced conspecifics in a tool-using task. Hirata, S. & Morimura, N. (Primate Research Inst., Kyoto Univ., Inuyama, Aichi, 484-8506, Japan [e-mail: [email protected]]). Journal of Comparative Psychology, 2000, 114, 291-296.
* Solving a cooperation task without taking into account the partner's behavior: The case of capuchin monkeys (Cebus apella). Visalberghi, E., Quarantotti, B. P., & Tranchida, F. (Ist. Di Psicologia del Consiglio Naz. Delle Ricerche, Reparto di Psicologia Comparata, Via Aldrovandi 16 b, 00197 Rome, Italy [e-mail: [email protected]]). Journal of Comparative Psychology, 2000, 114, 297-301.
* Birth order and hand preference in chimpanzees (Pan troglodytes): Implications for pathological models of handedness in humans. Hopkins, W. D. & Dahl, J. F. (Div. of Psychobiology, Yerkes RPRC, Emory Univ., Atlanta, GA 30322 [e-mail: [email protected]]). Journal of Comparative Psychology, 2000, 114, 302-306.
* Capuchins do cooperate: The advantage of an intuitive task. Mendres, K. A. & de Waal, F. B. M. (F. B. M. W., Living Links Center, Yerkes RPRC, Emory Univ., 954 N. Gatewood Rd, Atlanta, GA 30329 [e-mail: [email protected]]). Animal Behaviour, 2000, 60, 523-529.
* Grooming, social bonding, and agonistic aiding in rhesus monkeys. Matheson, M. D. & Bernstein, I. S. (I. S. B., Dept of Psychology, Univ. of Georgia, Athens, GA 30602-3013 [e-mail: [email protected]]). American Journal of Primatology, 2000, 51, 177-186.
Care
* Injury risks among chimpanzees in three housing conditions. Baker, K. C., Seres, M., Aureli, F., & de Waal, F. B. M. (Living Links Center, Yerkes RPRC, Emory Univ., Atlanta, GA 30322 [e-mail: [email protected]]). American Journal of Primatology, 2000, 51, 161-175, 2000. Conservation
* Extinction of a West African red colobus monkey. Oates, J. F., Abedi-Lartey, M., McGraw, W. S., Struhsaker, T. T., & Whitesides, G. H. (Dept of Anthro., Hunter College, 695 Park Ave, New York, NY 10021 [e-mail: [email protected]]). Conservation Biology, 2000, 14, 1526-1532.
* Habitat alteration and the conservation of African primates: Case study of Kibale National Park, Uganda. Chapman, C. A. & Lambert, J. E. (Dept of Zoology, Univ. of Florida, Gainesville, FL 32610). American Journal of Primatology, 2000, 50, 169-185.
* Habitat use by Chiropotes satanas and syntopic platyrrhines in eastern Amazonia. Bobadilla, U. L. & Ferrari, S. F. (Dpto de Biologia, Centro de Ciencias Naturais e Exatas, Univ. Luterana do Brasil, Rua Miguel Tostes, 101, 92.420-380 Canoas, RS, Brazil [e-mail: [email protected]]). American Journal of Primatology, 2000, 50, 215-224.
* Mass differences among male mangabey monkeys inhabiting logged and unlogged forest compartments. Olupot, W. (Institute for Tropical Forest Ecology, Mbarara University of Science and Technology, P.O. Box 44, Kabale, Uganda [e-mail [email protected]]). Conservation Biology, 2000, 14, 833-843.
* Genome resource banking for wildlife research, management, and conservation. Wildt, D. E. (Dept of Reproductive Sciences, Conservation and Research Center, Nat. Zool. Park, Washington, DC 20008). ILAR Journal, 2000, 41, 228-234.
Development and Aging
* Effects of aging on bone mineral content and bone biomarkers in female cynomolgus monkeys. Chen, Y., Shimizu, M., Sato, K., Koto, M., Tsunemi, K., Yoshida, T., & Yoshikawa, Y. (Tsukuba Primate Center, NIID, Hachimandai 1, Tsukuba-shi, Ibaraki 305-0843, Japan). Experimental Animals, 2000, 49, 163-170.
* Placental expression of the nonclassical MHC class I molecule Mamu-AG at implantation in the rhesus monkey. Slukvin, I. I., Lunn, D. P., Watkins, D. I., & Golos, T. G. (T. G. G., Wisconsin RPRC, Univ. of Wisconsin, 1223 Capitol Ct, Madison, WI 53715-1299 [e-mail: [email protected]]. Proceedings of the National Academy of Sciences, USA, 2000, 97, 9104-9109.
Disease
* Adrenal gland tumours in two cotton-top tamarins (Saguinus oedipus oedipus). M. Brack. (German Primate Center (DPZ), Kellnerweg 4, 3400 Göttingen, Germany). Laboratory Animals, 2000, 34, 106-110.
* KSHV-like herpesviruses in chimps and gorillas. Lacoste, V., Mauclre, P., Dubreuil, G., Lewis, J., Georges-Courbot, M.-C., & Gessain, A. (A. G., Unité d' Oncologie Virale, Dépt du SIDA et des Rétrovirus, Inst. Pasteur, 25-28 rue du Dr. Roux, 75724 Paris, CEDEX 15, France [e-mail: [email protected]]). Nature, 2000, 407, 151-152.
* Squamous cell carcinoma of the oral cavity in an infant cynomolgus monkey. Nakamura, S., Sakakibara, I., Ono, F., Shibata, S., Michishita, M., Ishii, Y., Kobayashi, R., Takahashi, K., & Yoshikawa, Y. (Dept of Vet. Pathology, Nippon Veterinary & Animal Science Univ., 1-7-1 Kyonan-cho, Musashino, Tokyo 180-8602, Japan). Experimental Animals, 2000, 49, 225-228.
* A new group of hepadnaviruses naturally infecting orangutans (Pongo pygmaeus). Warren, K. S., Heeney, J. L., Swan, R. A., Heriyanto, & Verschoor, E. J. (J. L. H., Biomedical Primate Research Center (BPRC), Department of Virology, P.O. Box 3306, 2280 GH Rijswijk, The Netherlands [e-mail: [email protected]]. Journal of Virology, 1999, 73, 7860-7865.
* Detection of hepatitis B virus infection in wild-born chimpanzees (Pan troglodytes verus): Phylogenetic relationships with human and other primate genotypes. MacDonald, D. M., Holmes, E. C., Lewis, J. C. M., & Simmonds P. (P.S., Laboratory for Clinical and Molecular Virology, University of Edinburgh, Summerhall, Edinburgh EH9 1QH, U.K. [e-mail: [email protected]]). Journal of Virology, 2000, 74, 4253-4257.
Evolution, Genetics, and Taxonomy
* A primate genome project deserves high priority. McConkey, E. H. & Varki, A. [Dept of Molecular, Cellular, and Developmental Biology, Univ. of Colorado, Boulder, CO 80309]. Science, 2000, 289, 1295-1296.
* Neanderthal diet at Vindija and Neanderthal predation: The evidence from stable isotopes. Richards, M. P., Pettitt, P. B., Trinkaus, E., Smith, F. H., Paunovi, M., & Karavani, I. (E. T., Unité Mixte de Recherche, 5809 du Centre National de la Recherche Scientifique, Lab. d'Anthropologie, Univ. de Bordeaux, 33405 Talence, France [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, USA, 2000, 97, 7663-7666.
* Pleistocene milestones on the Out-of-Africa corridor at Gesher Benot Ya'aqov, Israel. Goren-Inbar, N., Feibel, C. S., Verosub, K. L., Melamed, Y., Kislev, M. E., Tchernov, E., & Saragusti, I. (Institute of Archaeology, Hebrew University, Mt. Scopus, Jerusalem 91905, Israel [e-mail: [email protected]]). Science, 2000, 289, 944-947.
* A genetic linkage map of the baboon (Papio hamadryas) genome based on human microsatellite polymorphisms. Rogers, J., Mahaney, M. C., Witte, S. M., Nair, S. Newman, D., Wedel, S., Rodriguez, L. A., Rice, K. S., Slifer, S. H., Perelygin, A., Slifer, M., Palladino-Negro, P., Newman, T., Chambers, K., Joslyn, G., Parry, P., & Morin, P. A. (Dept of Genetics, Southwest Foundation for Biomedical Research, P.O. Box 760549, San Antonio, TX 78227 [e-mail: [email protected]]). Genomics, 2000, 67, 237-247.
* Remarkable species diversity in Malagasy mouse lemurs (primates, Microcebus). Yoder, A. D., Rasoloarison, R. M., Goodman, S. M., Irwin, J. A., Atsalis, S., Ravosa, M. J., & Ganzhorn, J. U. (Dept of Cell & Molecular Biology, Northwestern Univ. Med. School, Chicago, IL 60611 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, USA, 2000, 97, 11325-11330.
* Mitochondrial genome variation and the origin of modern humans. Ingman, M., Kaessmann, H., Pääbo, S. & Gyllensten, U. (U. G., Dept of Genetics & Pathol., Rudbeck Lab., Univ. of Uppsala, S-75185 Uppsala, Sweden [e-mail: [email protected]]). Nature, 2000, 408, 708-713.
Field Studies
* Foraging ecology of the mountain monkey (Cercopithecus l'hoesti): Implications for its evolutionary history and use of disturbed forest. Kaplin, B. A. & Moermond, T. C. (Dept of Environmental Studies, Antioch New England Grad. School, 40 Avon St, Keene, NH 03431-3516). American Journal of Primatology, 2000, 50, 227-246.
Reproduction
* Effect of Cetrorelix on sperm morphology during migration through the epididymis in the cynomolgus macaque (Macaca fascicularis). Gago, C., Soler, C., Pérez-Sánchez, F., Yeung, C. H., & Cooper, T. G. (T. G. C., Inst. of Reproductive Med. of the University, Domagkstr. 11, D-48129 Münster, Germany [e-mail: [email protected]]). American Journal of Primatology, 2000, 51, 103-117.
* Hormonal correlates of reproductive seasonality in wild female Hanuman langurs (Presbytis entellus). Ziegler, T., Hodges, K., Winkler, P., & Heistermann, M. (M. H., Dept of Reproductive Biology, German Primate Center, Kellnerweg 4, D-37077 Göttingen, Germany [e-mail: [email protected]]). American Journal of Primatology, 2000, 51, 119-134.
* Phylogenetic analysis of twinning in Callitrichinae. Ah-King, M. & Tullberg, B. S. (Dept of Zoology, Stockholm Univ., SE-106 91, Stockholm, Sweden [e-mail: [email protected]]). American Journal of Primatology, 2000, 51, 135-146.
----------
* * *
The International Journal of Comparative Psychology (IJCP) invites your submissions. The IJCP publishes studies on the evolution and development of animal behavior, including humans. Articles based on experimental or descriptive research procedures, carried out in laboratory, semi-natural, or natural settings, and involving mechanistic, developmental, functional-adaptive, or evolutionary theoretical approaches are welcome. Articles may include a single study or a series of related studies. Papers developing original theoretical models of behavior and brief reports not exceeding 4000 words (including references) will also be considered for publication. Articles reviewing the literature in a particular area of research will be occasionally accepted for consideration when the author has demonstrated significant contributions to the topic under review.
Some of the relevant areas include animal communication, animal learning and cognition, behavior analysis, behavior genetics, behavioral ecology, comparative behavioral neuroscience, developmental psychobiology, evolutionary psychology, migratory behavior, models of animal behavior, motivation, sensory-perceptual processes, and social behavior.
Send four copies of your manuscript, prepared according to APA format (see last edition of the APA Publication Manual for guidelines), and a cover letter indicating the name and address (including e-mail address) of three potential reviewers to: Mauricio R. Papini, Editor, IJCP, Dept of Psychology, Texas Christian Univ., Box 298920, Fort Worth, TX 76129. - From ABSnet, 6[29], 2000
ILAR Journal
The Institute for Laboratory Animal Research (ILAR) is pleased to announce that Gerald F. Gebhart, PhD, is the new ILAR Journal Editor-in-Chief. Dr. Gebhart is a member of the ILAR Council and Chairman of the Department of Pharmacology at the University of Iowa.
APE Boletín on the Web
The Spanish language Boletín de la Asociación Primatológica Espańola is now available on the Web at <www.uam.es/ape>. All of Volume 7 (2000) is available there.
* * *
All correspondence concerning the Newsletter should be addressed to: Current and back issues of the Newsletter are available on the World Wide Web at The Newsletter is supported by U. S. Public Health Service Grant RR-00419 from the Comparative Medicine Program, National Center for Research Resources, N.I.H.
Cover illustration of a chimpanzee (Pan troglodytes) by Margaret Dannemiller (Warner Lambert/Parke Davis Research)
Copyright (c) 2001 by Brown University
Assistant Editor: Elva Mathiesen
Last updated: December 18, 2000Research and Educational Opportunities
Continuing Education for Lab Animal Technicians
* Primate Behavior;
* Approaches to the Study of Behavior; and
* Laboratory for Approaches to the Study of Behavior.
The Primate Behavior and Ecology Program will provide the tools for understanding behavioral and ecological processes and problems, while immersing students in a field research site focused on primate social ecology and conservation. Information Requested or Available
International Veterinary Information Service Grants Available
ACLAM Foundation Request For Proposals Resources Wanted and Available
Species Information Service Launched Recent Books and Articles
(Addresses are those of first authors)
. . .
Sections are: Diversity; Behavioral Ecology; Community Ecology; Distribution, Abundance, and Rarity; Population Biology; Extinction Processes; Habitat Disturbance; Hunting; Conservation Strategies; Conservation Tactics; and Conclusions.
. . .
Sections are: History; Controlling aggression; Repairing the damage; Triadic affairs; Ecological and cultural contexts; and Conclusion.
. . .
Sections are: Animal biology; Animal health and husbandry; Nursing; and an appendix of anatomy and physiology terminology.
. . .
Contents: Comparative perspectives on male-female association; Variation in male numbers: Taxon-level analyses; Variation in adult sex ratios of red colobus monkey social groups; Behavioral aspects of male coexistence; Evolutionary determinants and consequences; Conclusions: Understanding male primates.
. . .
Chapters include: Comparative developmental evolutionary biology, anthropology, and psychology: Convergences in the study of human behavioral ontogeny, by S. T. Parker; The developmental timing of primate play: A neural selection model, by L. A. Fairbanks; The heterochronic evolution of primate cognitive development, by J. Langer; and Cultural apprenticeship and culture change: Tool learning and imitation in chimpanzees and humans, by P. Greenfield, A. Maynard, C. Boehm, & E. Yut Schmidtling.
. . .
Contents: Historical introduction: Evolution and theories of mind and behaviour, Darwin and after; Darwin's legacy; The selfish gene; Mating behaviour: From systems to strategies; Sexual selection; The evolution of brain size; Language and the modular mind; Understanding human sexual behaviour: Anthropological approaches; Human mate choice: The evolution logic of sexual desire; Conflict within families and other groups; Altruism, cooperation and the foundations of culture; Epilogue: The use and abuse of evolutionary theory.
. . .
A revision and expansion of the first edition, including more than 800 entries by 49 authors.
. . .
A sequel to, and updating of, the 1995 Three Dimensional Biomedical Imaging: Principles and Practice.
. . .
This video has a strong emphasis on proper use of personal protective equipment combined with understanding nonhuman primate behavior. The general principles are applicable to any program that houses nonhuman primates.
. . .
Projects are listed by continent. There are tables and graphs giving the distributions of studies by geographic region, taxonomy, subject, and duration.
. . .
The behavioral and ecological factors involved in the evolution of the immune system remain poorly explored. We present a phylogenetic analysis of white blood cell counts in primates to test three hypotheses related to disease risk: increases in risk are expected with group size or population density, exposure to soil-borne pathogens, and mating promiscuity. White blood cell counts were significantly greater in species where females have more mating partners, indicating that the risk of sexually transmitted disease is likely to be a major factor leading to systematic differences in the primate immune system.
. . .
Male cotton-top tamarins, Saguinus oedipus, display hormonal changes associated with parenting prior to the birth of their infants. Here we examined the hormonal changes that occurred in experienced and inexperienced fathers during the postconception period, prior to the birth of infants. Noninvasive techniques were used to collect urine from 10 male cotton-top tamarins (5 experienced and 5 inexperienced breeders) three times weekly during the 6-month gestation period. Samples were analyzed for prolactin, testosterone, dihydrotestosterone (DHT), and cortisol, averaged by gestational month. Experienced males showed elevated prolactin during the mate's third gestational month, and the elevation correlated with the number of infants surviving from the previous birth (0, 1, and 2) but not with outcome of the current pregnancy. However, an experienced male with no infants present still showed elevated prolactin and some inexperienced males showed elevated prolactin just before parturition, suggesting non-infant cues are also important. While prolactin levels were influenced by the male's prior infant experience, testosterone levels did not differ between experienced and inexperienced males. Testosterone levels were significantly elevated for all males during the third, fourth and fifth months but had no relationship with number of infants present or with outcome of current pregnancy. DHT decreased during the second half of pregnancy compared with testosterone but this finding was not consistent for every male. No significant changes occurred in cortisol levels. These results suggest that infant-rearing experience affected the hormonal responsiveness of the male to his mate's current pregnancy.
. . .
The Tat protein is essential for HIV type 1 (HIV-1) replication and may be an important virulence factor in vivo. We studied the role of Tat in viral pathogenesis by immunizing rhesus macaques with chemically inactivated Tat toxoid and challenging these animals by intrarectal inoculation with the simian/human immunodeficiency virus 89.6PD. Immune animals had significantly attenuated disease with lowered viral RNA, interferon- [alpha ], and chemokine receptor expression (CXCR4 and CCR5) on CD4+ T cells; these features of infection have been linked to in vitro effects of Tat and respond similarly to extracellular Tat protein produced during infection. Immunization with Tat toxoid inhibits key steps in viral pathogenesis and should be included in therapeutic or preventive HIV-1 vaccines.
. . .
The aim of this study was to investigate the distribution of human T-cell leukemia virus type 1 (HTLV-1) in various organs of serially sacrificed squirrel monkeys in order to localize the reservoir of the virus and to evaluate the relationship between viral expression and the humoral or cellular immune response during infection. Six squirrel monkeys infected with HTLV-1 were sacrificed 6, 12, and 35 days and 3, 6, and 26 months after inoculation, and 20 organs and tissues were collected from each animal. PCR and reverse transcription-PCR (RT-PCR) were performed with gag and tax primers. Proviral DNA was detected by PCR in peripheral blood mononuclear cells (PBMCs) of monkeys sacrificed 6 days after inoculation and in PBMCs, spleens, and lymph nodes of monkeys sacrificed 12 and 35 days and 3, 6, and 26 months after inoculation. Furthermore, tax/rex mRNA was detected by RT-PCR in the PBMCs of two monkeys 8 to 12 days after inoculation and in the spleens and lymph nodes of the monkey sacrificed on day 12. In this animal, scattered HTLV-1 tax/rex mRNA-positive lymphocytes were detected by in situ hybridization in frozen sections of the spleen, around the germinal centers and close to the arterial capillaries. Anti-HTLV-1 cell-mediated immunity was evaluated at various times after inoculation. Anti-p40Tax and anti-Env cytolytic T-cell responses were detected 2 months after infection and remained detectable thereafter. When Tax peptides were used, this response appeared to be directed against various Tax epitopes. Our results indicate that squirrel monkeys represent a promising animal model for studying the early events of HTLV-1 infection and for evaluating candidate vaccines against HTLV-1.
. . .
Prolonged antiretroviral therapy (ART) is not likely to eradicate human immunodeficiency virus type I (HIV-I) infection. Here we explore the effect of therapeutic immunization in the context of ART during primary infection using the simian immunodeficiency virus (SIV251) macaque model. Vaccination of rhesus macaques with the highly attenuated poxvirus-based NYVAC-SIV vaccine expressing structural genes elicited vigorous virus-specific CD4+ and CD8+ T cell responses in macaques that responded effectively to ART. Following discontinuation of a six-month ART regimen, viral rebound occurred in most animals, but was transient in six of eight vaccinated animals. Viral rebound was also transient in four of seven mock-vaccinated control animals. These data establish the importance of antiretroviral treatment during primary infection and demonstrate that virus-specific immune responses in the infected host can be expanded by therapeutic immunization.
. . .
With accumulating evidence indicating the importance of cytotoxic T lymphocytes (CTLs) in containing human immunodeficiency virus-1 (HIV-1) replication in infected individuals, strategies are being pursued to elicit virus-specific CTLs with prototype HIV-1 vaccines. Here, we report the protective efficacy of vaccine-elicited immune responses against a pathogenic SHIV-89.6P challenge in rhesus monkeys. Immune responses were elicited by DNA vaccines expressing SIVmac239 Gag and HIV-1 89.6P Env, augmented by the administration of the purified fusion protein IL-2/Ig, consisting of interleukin-2 (IL-2) and the Fc portion of immunoglobulin G (IgG), or a plasmid encoding IL-2/Ig. After SHIV-89.6P infection, sham-vaccinated monkeys developed weak CTL responses, rapid loss of CD4+ T cells, no virus-specific CD4+ T cell responses, high setpoint viral loads, significant clinical disease progression, and death in half of the animals by day 140 after challenge. In contrast, all monkeys that received the DNA vaccines augmented with IL-2/Ig were infected, but demonstrated potent secondary CTL responses, stable CD4+ T cell counts, preserved virus-specific CD4+ T cell responses, low to undetectable setpoint viral loads, and no evidence of clinical disease or mortality by day 140 after challenge.
. . .
In a randomized controlled trial with acute simian immunodeficiency virus (SIV)-infected macaques, both highly active antiretroviral therapy (HAART) and HAART with fixed-schedule structured treatment interruption (STI-HAART; alternating 3 weeks on and 3 weeks off therapy) suppressed viral load. In the STI-HAART group, T cell virus-specific immune response (VIR) and control of viral rebound increased concurrently during subsequent interruptions. In contrast, VIR did not increase and SIV rebounded after permanent treatment withdrawal in all animals on continuous HAART. Fixed-schedule STI-HAART appears to be an effective alternative to continuous HAART for the early treatment of retroviral infection.
. . .
In humans, sterile immunity against malaria can be consistently induced through exposure to the bites of thousands of irradiated infected mosquitoes. The same level of protection has yet to be achieved using subunit vaccines. Recent studies have indicated an essential function for intrahepatic parasites, the stage after the mosquito bite, and thus for antigens expressed during this stage. We report here the identification of liver-stage antigen 3, which is expressed both in the mosquito and liver-stage parasites. This Plasmodium falciparum 200-kilodalton protein is highly conserved, and showed promising antigenic and immunogenic properties. In chimpanzees (Pan troglodytes), the primates most closely related to humans and that share a similar susceptibility to P. falciparum liver-stage infection, immunization with LSA-3 induced protection against successive heterologous challenges with large numbers of P. falciparum sporozoites.
. . .
Lentiviral delivery of glial cell line-derived neurotrophic factor (lenti-GDNF) was tested for its trophic effects upon degenerating nigrostriatal neurons in nonhuman primate models of Parkinson's disease (PD). We injected lenti-GDNF into the striatum and substantia nigra of nonlesioned aged rhesus monkeys or young adult rhesus monkeys treated 1 week prior with 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP). Extensive GDNF expression with anterograde and retrograde transport was seen in all animals. In aged monkeys, lenti-GDNF augmented dopaminergic function. In MPTP-treated monkeys, lenti-GDNF reversed functional deficits and completely prevented nigrostriatal degeneration. Additionally, lenti-GDNF injections to intact rhesus monkeys revealed long-term gene expression (8 months). In MPTP-treated monkeys, lenti-GDNF treatment reversed motor deficits in a hand-reach task. These data indicate that GDNF delivery using a lentiviral vector system can prevent nigrostriatal degeneration and induce regeneration in primate models of PD and might be a viable therapeutic strategy for PD patients.
. . .
Signals derived from the rat motor cortex can be used for controlling one-dimensional movements of a robot arm. It remains unknown, however, whether real-time processing of cortical signals can be employed to reproduce, in a robotic device, the kind of complex arm movements used by primates to reach objects in space. Here we recorded the simultaneous activity of large populations of neurons, distributed in the premotor, primary motor and posterior parietal cortical areas, as nonhuman primates performed two distinct motor tasks. Accurate real-time predictions of one- and three-dimensional arm movement trajectories were obtained by applying both linear and nonlinear algorithms to cortical neuronal ensemble activity recorded from each animal. In addition, cortically derived signals were successfully used for real-time control of robotic devices, both locally and through the Internet. These results suggest that long-term control of complex prosthetic robot arm movements can be achieved by simple real-time transformations of neuronal population signals derived from multiple cortical areas in primates.
. . .
To plan an action, we must first select an object to act on and the body part (or parts) to use to accomplish our intention. To plan the motor task of reaching, we specify both the target to reach for and the arm to use. In the process of planning and preparing a motor task, information about the motor target and the arm to use must be integrated before a motor program can be formulated to generate the appropriate limb movement. One of the structures in the brain that is probably involved in integrating these two sets of information is the premotor area in the cerebral cortex of primates. The lateral sector of the dorsal premotor cortex is known to receive both visual and somatosensory input, and we show here that neurons in this area gather information about both the target and the body part, while subsequent activity specifies the planned action.
. . .
The use of tools by New World primates has only been observed in capuchin monkeys, genus Cebus, mainly C. apella in captivity. In this note we report three cases of tool use by C. olivaceus in captivity. The study group of 11 individuals has been systematically observed in the Parque del Este, Caracas, where they live on an island in an artificial lake: therefore the monkeys have close contact with water resources. The observations are 1) use of half an orange peel as a water container; 2) use of leaves as a sponge; and 3) use of a branch as a cane for balance.
. . .
To determine how the birth and development of infants influence their mothers' social relationships with other adult troop members, we observed two free-ranging troops of ring-tailed lemurs at the Berenty Reserve, Madagascar. The number of acts of affiliative contact that the mothers received from other adult troop members during the first and second months of infant life were significantly higher than those before they gave birth, and the values during the third month were as low as that before giving birth. Two mothers received acts of affiliative contact less frequently after their infants died, compared with the values while the infants were alive. On the other hand, more than 95% of all acts of licking of infants by adult troop members other than their mothers occurred with the infants were in contact with their mothers. These findings suggest that infants per se and mothers per se were not attractive, but rather the infant-mother pair was attractive to other troop members. Frequency of infant-licking was not affected by female parity, female and male dominance rank, or infant sex. Acts of infant-licking, which are widespread among troop members, may function to make or maintain stable social relationships.
. . .
Male rhesus monkeys unfamiliar with each other were paired in a cage, and blood samples were collected before and a few hours after pair formation. Adrenocorticotrophic hormone (ACTH) and cortisol levels in each blood sample were measured. The dominance relationship was determined in seven pairs formed from five animals, by observing food competition and agonistic behavior immediately after pair formation; and the differences in ACTH and cortisol values between the dominant and subordinate animal in these pairs were compared statistically. The day after the first encounter, a second encounter was conducted in randomly selected pairs of monkeys. In the first encounters, higher levels of both ACTH and cortisol were detected in dominant animals in comparison to subordinates. Changing the animal's partner altered the stress responses whenever the animal's status changed. Elevated levels of ACTH and cortisol in dominant animals disappeared on the day after the first encounter.
. . .
In a group of rhesus monkeys, feeding tactics of juveniles were studied in a competitive situation in which food presentation had been modified, and where food was supplied in a feeding trough six times a day. Juvenile offspring of high-ranking mothers remained longer in the feeding area after gathering food. In contrast, juvenile offspring of low-ranking mothers went less often to the feeding area, primarily to collect food. Low-ranking juveniles gathered food faster than did dominants and used longer feeding bouts. High-ranking juveniles interrupted their feeding more spontaneously than did subordinates. Juvenile males entered the feeding area to satisfy their food requirements more often than females, but were also aggressively expelled more than females. No relation was found between age in months of juveniles and any feeding parameters or causes of interruption; nor were significant differences found between old and young juveniles.
. . .
Both species, living in the Amazon rainforest of northeastern Peru, prey on frogs and lizards, and very rarely on nestling birds. As a result of divergent foraging strategies, S. mystax primarily exploited frogs at higher strata of the forest, while S. fuscicollis predominantly preyed on reptiles in the lower strata and on the ground. This difference may strengthen the niche differentiation between these two tamarin species that exists with regard to other prey.
. . .
Three captive gibbons were videotaped in the course of longitudinal exposure to mirrors introduced into their familiar cage or island housing situation. The gibbons, which differed in age, sex, species, and rearing condition, exhibited great individual differences in their behavioral reactions to mirrors, spanning from a minimal reaction dominated by social responses to a dramatic sequence of progressive behavioral change that featured a variety of contingency testing behaviors and included mirror-mediated, self-directed behavior. Additional information on the mirror competence of gibbons was provided by modified mark tests and a hidden object task. The results are discussed in relation to current criteria for self-recognition in primates as well as individual and species differences in reactions to mirror exposure.
. . .
The authors investigated the development of unimanual hand use and hand preferences during feeding in 15 marmosets (Callithrix jacchus), ages birth to 51-70 months. Bimanual hand use was common at 1-2 months, but by 5-8 months unimanual holding had developed and so had significant hand preferences. Half of the marmosets preferred to pick up and take food to the mouth with the left hand, and half preferred the right hand. Individuals maintained the same hand preference at all ages examined. Significant relationships were also found between the postures adopted during feeding and the direction of hand preferences displayed by juvenile marmosets. There was a positive correlation between increased suspension and increased left-hand preference, and a negative correlation between increased feeding in a tripedal posture and increased left-hand preference. These results are discussed in terms of motor development and hemispheric specialization.
. . .
The authors investigated the occurrence of naive chimpanzees' spontaneous observation of experienced conspecifics during a tool-use task entailing honey fishing. The chimpanzees were presented with 20 kinds of "tools" of which 12 kinds were usable. Six pairs of naive and experienced chimpanzees were brought to this honey-fishing situation. A total of 40 observation episodes occurred between the naive and experienced groups, 34 of which were from naive toward experienced individuals. Naive chimpanzees never observed their partners after their own success but did so after their own failure or before their first attempts. In addition, there were 10 cases in which naive individuals used the discarded tools of the experienced ones. Two factors for the transmission of tool use were clearly evident in this study: (a) spontaneous observation of an appropriate behavioral sequence and (b) enhanced environmental cues made by skilled individuals.
. . .
Four pairs of tufted capuchin monkeys (Cebus apella) were tested in a task requiring that both partners pull a handle simultaneously in order for both to be rewarded. The experimental design, an improved version of that of R. Chalmeau, E. Visalberghi, and A. Gallo (1997), aimed at assessing the extent to which a monkey that is pulling takes account of the behavior and spatial position of its partner, that is, whether the monkey understands what cooperation involves. Although all pairs succeeded, pulling was not affected by the partner's behavior, and it was affected only to a certain extent by the partner's spatial position. In addition, more experienced capuchins did not outperform naive individuals. The finding that capuchins were successful without understanding the role of the partner suggests that their cooperation is not cognitively grounded, as has been argued in descriptions of the hunting behavior of wild capuchins.
. . .
The effect of birth order on hand preference was assessed in a sample of 154 captive-born chimpanzees. Subjects were classified as first, middle, or latter born. Hand preference was measured using a task that elicited coordinated bimanual actions. Significant birth-order effects were found for both classification criteria, with first- and latter-born subjects exhibiting a lesser degree of right-handedness compared with middle-born subjects. These data suggest that biological rather than sociological factors play a greater role in explaining the observed birth-order effects on hand preference in humans.
. . .
We used a cooperative pulling task to examine proximate aspects of cooperation in captive brown capuchin monkeys, Cebus apella. Specifically, our goal was to determine whether capuchins can learn the contingency between their partner's participation in a task and its successful completion. We examined whether the monkeys visually monitored their partners and adjusted pulling behavior according to their partner's presence. Results on five same-sex pairs of adults indicate that (1) elimination of visual contact between partners significantly decreased success; (2) subjects glanced at their partners significantly more in cooperative tests than in control tests in which no partner-assistance was needed; and (3) they pulled at significantly higher rates when their partner was present rather than absent. Therefore, in contrast to a previous report, cooperating capuchins do seem able to take the role of their partner into account. However, the type of task used may be an important factor affecting the level of coordination achieved.
. . .
An analysis of simultaneous grooming bouts in a captive group of rhesus monkeys (Macaca mulatta) failed to provide evidence of competition to groom high ranking partners. Not only were grooming supplantations rare, but the highest ranking individuals performing grooming did not groom the highest ranking animals receiving grooming. Lower ranking partners, however, did more grooming in nonkin dyads. Grooming partners aided one another in agonistic episodes, but the individual receiving the aid did not groom the individual providing the aid more than vice versa. Kin dyads did aid and groom one another at greater than expected rates, but the aider did not receive the greater proportion of grooming in the dyad. Males participated in more grooming than expected, but their grooming was not related to aiding either with regard to one another or female partners. Animals that were targeted in joint aggression, or aided against, received significantly less grooming from their opponents. A general social relationship expressed in partner preferences, social grooming, and agonistic aiding better explained the observed pattern than any model based on the exchange of services for favors in different currencies.
. . .
Meeting the psychological needs of chimpanzees (Pan troglodytes) can be a challenge given their aggressiveness on the one hand and the complexity of their social lives on the other. It is unclear how to balance the need to provide opportunities for species-appropriate behavior against potential risks of injury chimpanzees may inflict on each other. This study evaluates the suggestion that simpler social environments protect chimpanzees from wounding. Over a two-year period all visible injuries to 46 adult males, 64 adult females, and 25 immature chimpanzees were recorded at the Yerkes Regional Primate Research Center. Approximately half of the subjects were mother-reared, and the rest were nursery-reared. Housing included compounds containing about 20 chimpanzees, interconnected indoor-outdoor runs for groups of up to 12 individuals, and smaller indoor-outdoor runs for pairs and trios. Annual wounding rates were calculated for serious wounds (extensive injuries and all those requiring veterinary intervention) as well as for minor wounds. Compound-housed chimpanzees incurred the highest level of minor wounding, but serious wounding levels were not affected by housing condition. Even with a period of dominance instability and elevated levels of wounding in one compound, compound chimpanzees were not injured more than those in smaller social groups over the long term. Nursery-reared females in moderate-sized groups were wounded more than mother-reared females. Also, nursery-reared males and females were wounded less often when paired with mother-reared companions. Overall, this study indicates that maintaining chimpanzees in pairs and trios would not be an effective means for reducing injuries. The management of wounding in chimpanzee colonies is influenced more by the sex and rearing composition of a colony.
. . .
The first documented case of the extinction in the 20th century of a widely recognized primate taxon. During surveys in Ghana and Côte d'Ivoire in 1993-1999, no surviving populations of Miss Waldron's red colobus monkey (Procolobus badius waldroni), a primate taxon endemic to the forests of this part of West Africa, were found. It is concluded that this omnkey, which at least one authority considers worthy of species status, is probably extinct. Hunting by humans appears to be the ultimate cause of the extinction. The extinction of other large animals in the Upper Guinea rainforest region is likely to follow soon unless more attention is paid to the full range of endangered forms and more resources are devoted to their rigorous protection.
. . .
A case study of patterns of primate abundance in areas of Kibale National Park (766 km2) that have been modified by different types and intensities of human activities, primarily commercial logging and agricultural clearing. Distributional surveys at 24 sites and detailed line-transect censuses at six sites demonstrate that primate populations in Kibale are often high and suggest that patterns of population change associated with disturbance are complex.
. . .
Primates were surveyed at two sites in the Xingu-Tocantins interfluvium, where at least eight platyrrhines are known to occur, including the endemic Chiropotes satanas utahicki, vulnerable to extinction. Data on habitat use were collected in standard line transect surveys and analyzed with relation to the availability of forest types, as well as between sites and species. It remains unclear whether and to what extent observed patterns are determined by differences between taxa, populations, or ecosystems, but the data indicate that C. s. utahicki is relatively tolerant of habitat disturbance.
. . .
Remedies for reduction in primate numbers following rainforest habitat disturbance must incorporate understanding of how disturbance affects population biology. Between July, 1996, and July, 1998, I captured 31 adult male grey cheeked mangabeys (Lophocebus albigena) from groups occupying logged and unlogged forest in Kibale National Park, Uganda. Captured males were weighed, measured, fitted with radio-collars, and radio-tracked three to five times a week. Males from groups in the unlogged forest were significantly heavier than males from the logged forest. Males that immigrated into the study groups were significantly lighter than resident males in unlogged forest, but they did not differ in mass from males in logged forest. I tested hypotheses that these differences were due to differences in age, skeletal size, and tissue mass. Results support the hypothesis that differences are due to differences in tissue mass. Results of further tests showed that differences in tissue mass were explained more by differences in habitat type than by rates of male turnover. There was no evidence to suggest that differences in tissue mass were related to differences in costs of travel, crowding, or seasonal variation in habitat quality. Results suggest that selective logging may affect primate biomass, although these changes may or may not be reflected in estimates of primate density.
. . .
The benefits of developing "genome resource banks" for wildlife are profound, perhaps more so than for traditional uses in terms of livestock and human fertility. In addition to preserving heterozygosity and assisting in the genetic management of rare populations held in captivity, frozen respositories help insure wild populations against natural and human-induced catastrophes. It is crucial that genome resource banks for wildlife species be developed in a coordinated fashion that first benefits the conservation of biodiversity. Areas requiring priority attention are reviewed, including the need for more basic research, advocacy, and support for developing organized repositories of biomaterials representing the world's diverse biota.
. . .
Age-related changes in bone mineral content and bone biomarkers were assessed over the complete lifespan in female cynomolgus monkeys. The bone mass of the lumbar spine increased linearly from birth to about 2.5 years of age, and this increase gradually slowed thereafter until a peak bone mass was achieved at 9 years. The bone mass stabilized after 9 years of age, showing no sign of further reducation with age. In contrast with the significant increase in bone mass before 2.5 years, significant decreases occurred in the serum concentrations of the following bone formation markers: intact osteocalcin, bone-specific alkaline phosphatase, and amino-terminal propeptide of type I procollagen, but the serum concentration of carboxy-terminal propeptide of type I procollagen did not change significantly throughout the entire lifespan. Concerning the bone resorption markers, the levels of tartrate-resistant acid phosphatase fluctuated throughout the entire lifespan. The skeleton of an aging female monkey undergoes changes similar to those observed in senescent humans, but did not undergo the menopausal changes seen in women. The use of female cynomolgus monkeys to model human skeletal interventions should therefore be undertaken with consideration of the similarities and differences between the monkeys and humans.
. . .
During human implantation, trophoblasts mediate attachment of the embryo to the uterine epithelium and invade and reorganize vessels of the maternal endometrium to initiate blood flow to the intervillous space. Expression of the nonclassical MHC class I molecule HLA-G by invading trophoblasts may play a central role in their protection from recognition by the maternal immune system; however, the ontogeny of trophoblast HLA-G expression during the earliest stages of implantation is difficult to evaluate in human pregnancy. We previously identified a novel nonclassical MHC class I molecule, Mamu-AG, which is expressed in the rhesus monkey placenta and shares many unique characteristics of HLA-G. Immunocytochemical analysis with a Mamu-AG-specific mAb and locus-specific in situ hybridization of rhesus implantation sites 7-12 days after embryo attachment (days 14-19 of pregnancy) demonstrated that Mamu-AG molecules are expressed predominantly in cytotrophoblasts invading the maternal vessels and endometrium, whereas syncytiotrophoblasts covering trophoblastic lacunae or newly formed chorionic villi remained largely Mamu-AG-negative. By day 36 of pregnancy, Mamu-AG glycoprotein also was expressed in villous syncytiotrophoblasts, and accumulation of Mamu-AG glycoprotein was noted at the border between maternal decidua and fetal trophoblasts. The ontogeny of a nonclassical MHC class I molecule at the implantation site supports the hypothesis that its expression is important for the establishment of maternal-fetal immune tolerance.
. . .
Two adrenal gland tumors in captive-born cotton-top tamarins (Saguinus oedipus oedipus) are described. One was a pheochromocytoma in a 14-year-old male, the other one a cortical adenoma in a 7.5-year-old female. Both were associated with morphological signs of myocardial damage and circulatory problems. The findings are discussed and compared to adrenal gland tumors in man.
. . .
Among the herpesviruses, KSHV (Kaposi's-sarcoma-associated herpesvirus) is the human prototype of the rhadinovirus genus. Rhadinoviruses (or gamma-2-herpesviruses) are found in several animal species, including New and Old World monkeys, but not in the great apes. Here we describe the detection and sequencing of a polymerase gene fragment from three new rhadinoviruses discovered in chimpanzees and in a gorilla, which are more closely related to KSHV than to any other virus of this genus described so far. Our results indicate that the great apes from central Africa could provide a reservoir of new ?2-herpesviruses that are potentially transmissible to humans.
. . .
Squamous cell carcinoma was observed in the oral cavity of a one-year-old male cynomolgus monkey. The tumor consisted of various shaped cells; its assemblies infiltrated into the surrounding connective tissues. Although no obvious metaplastic keratinized cancer pearls were found in the tumor cells, intercellular bridges were observed. Tumor cells were stained with anti-keratin, but not with anti-vimentin. No papilloma virus antigen or Epstein-Barr Virus small mRNA could be detected. Under the electron microscope, incomplete tonofibrils and desmosomes in the cytoplasm and microvillus of the cell membrane were observed, suggesting a malignancy or low differentiation of the tumor cells.
. . .
A high prevalence (42.6%) of hepatitis B virus (HBV) infection was suspected in 195 formerly captive orangutans due to a large number of serum samples which cross-reacted with human HBV antigens. It was assumed that such viral infections were contracted from humans during captivity. However, two wild orangutans were identified which were HBV surface antigen positive, indicating that HBV or related viruses may be occurring naturally in the orangutan populations. Sequence analyses of seven isolates revealed that orangutans were infected with hepadnaviruses but that these were clearly divergent from the six known human HBV genotypes and those of other nonhuman hepadnaviruses reported. Phylogenetic analyses revealed geographic clustering with Southeast Asian genotype C viruses and gibbon ape HBV. This implies a common origin of infection within this geographic region, with cross-species transmission of hepadnaviruses among hominoids.
. . .
Infection with hepatitis B virus (HBV) was detected by serological testing for HBV surface antigen and by PCR assay for HBV DNA in serum samples from two common chimpanzees born in West Africa. The complete genome sequences obtained by nucleotide sequencing of overlapping DNA fragments amplified by PCR were compared with HBV variants recovered from other primates and with human genotypes A to F. Both chimpanzee sequences were 3,182 nucleotides in length, and the surface gene sequence predicted the existence of a, d, and w serological determinants. Neither sequence contained stop codons in the precore region. On phylogenetic analysis, the HBV variants infecting the chimpanzees clustered together with a third chimpanzee HBV isolate independently obtained from an infected captive animal, with an overall sequence similarity of >94%. This provides strong evidence for a chimpanzee-specific genotype of HBV which circulates in nature. These findings add to the recent evidence for infection in the wild of other Old and New World primates (gibbon, orangutan, and woolly monkey) with species-specific variants of HBV. There is no evidence for close phylogenetic clustering of variants found so far in primates with any of the established HBV genotypes from humans. With the new evidence for the widespread distribution of HBV in primates, hypotheses for the origins of human infection are reviewed.
. . .
A letter, with numerous distinguished cosignatories, urging that the chimpanzee and at least one of the commonly studied Old World primates should be given high priority for genome sequencing.
. . .
Archeological analysis of faunal remains and of stone and bone tools has suggested that hunting of medium to large mammals was a major element of Neanderthal subsistence. Plant foods are almost invisible in the archeological record, and it is impossible to estimate accurately their dietary importance. However, stable isotope (delta-13C and delta-15N) analysis of mammal bone collagen provides a direct measure of diet and has been applied to two Neanderthals and various faunal species from Vindija Cave, Croatia. The isotope evidence overwhelmingly points to the Neanderthals behaving as top-level carnivores, obtaining almost all of their dietary protein from animal sources. Earlier Neanderthals in France and Belgium have yielded similar results, and a pattern of European Neanderthal adaptation as carnivores is emerging. These data reinforce current taphonomic assessments of associated faunal elements and make it unlikely that the Neanderthals were acquiring animal protein principally through scavenging. Instead, these findings portray them as effective predators.
. . .
The Acheulean site of Gesher Benot Ya'aqov in the Dead Sea Rift of Israel documents hominid movements and technological development on a corridor between Africa and Eurasia. New data place the site at 780,000 years ago (oxygen isotope stage 19), considerably older than previous estimates. The archeological data from the site portray strong affinities with African stone tool traditions. The findings also reflect adroit technical skills and in-depth planning abilities, more advanced and complex than those of earlier archeological occurrences in the Levant.
. . .
A first-generation genetic linkage map of the baboon (Papio hamadryas) genome was developed for use in biomedical and evolutionary genetics. Pedigreed baboons (n = 694) were selected from the breeding colony maintained by the Southwest Foundation for Biomedical Research. To facilitate comparison with the human genome, the baboon linkage map consists primarily of human microsatellite loci amplified using published human PCR primers. Genotypes for 325 human microsatellites and 6 novel baboon microsatellites were used in linkage analyses performed with the MultiMap expert system. The resulting sex-averaged meiotic recombination map covers all 20 baboon autosomes, with average spacing among loci of 7.2 cM. Direct comparison among homologous (orthologous) loci reveals that, for 7 human autosomes, locus order is conserved between humans and baboons. For the other 15 autosomes, one or more rearrangements distinguish the two genomes. The total centimorgan distances among homologous markers are 28.0% longer in the human genome than in the baboon, suggesting that rates of recombination may be higher in humans. This baboon linkage map is the first reported for any nonhuman primate species and creates opportunities for mapping quantitative trait loci in baboons, as well as for comparative evolutionary analyses of genome structure.
. . .
Phylogenetic analysis of mtDNA sequence data confirms the observation that species diversity in the world's smallest living primate (genus Microcebus) has been greatly underestimated. The description of three species new to science, and the resurrection of two others from synonymy, has been justified on morphological grounds and is supported by evidence of reproductive isolation in sympatry. This taxonomic revision doubles the number of recognized mouse lemur species. The molecular data and phylogenetic analyses presented here verify the revision and add a historical framework for understanding mouse lemur species diversity. Phylogenetic analysis revises established hypotheses of ecogeographic constraint for the maintenance of species boundaries in these endemic Malagasy primates. Mouse lemur clades also show conspicuous patterns of regional endemism, thereby emphasizing the threat of local deforestation to Madagascar's unique biodiversity.
. . .
The analysis of mitochondrial DNA (mtDNA) has been a potent tool in our understanding of human evolution, owing to characteristics such as high copy number, apparent lack of recombination, high substitution rate and maternal mode of inheritance. However, almost all studies of human evolution based on mtDNA sequencing have been confined to the control region, which constitutes less than 7% of the mitochondrial genome. These studies are complicated by the extreme variation in substitution rate between sites, and the consequence of parallel mutations causing difficulties in the estimation of genetic distance and making phylogenetic inferences questionable. Most comprehensive studies of the human mitochondrial molecule have been carried out through restriction-fragment length polymorphism analysis, providing data that are ill suited to estimations of mutation rate and therefore the timing of evolutionary events. Here, to improve the information obtained from the mitochondrial molecule for studies of human evolution, we describe the global mtDNA diversity in humans based on analyses of the complete mtDNA sequence of 53 humans of diverse origins. Our mtDNA data, in comparison with those of a parallel study of the Xq13.3 region in the same individuals, provide a concurrent view on human evolution with respect to the age of modern humans.
. . .
C. l'hoesti spent 35% of observation time feeding on terrestrial herbaceous vegetation. Thirty-five fruit and seed species comprised 42% of their diet, and invertebrates composed 9%. They spent 38% of observation time on the ground and 27% in canopy trees. Although they primarily foraged in undisturbed open areas, they also foraged in disturbed open habitats. Synthesizing these results with Pleistocene forest history and C. l'hoesti evolutionary history, the authors suggest that, although these monkeys readily use disturbed forest for herb foraging, they are a forest-adapted species which has adopted a terrestrial lifestyle to exploit the abundant herb layer common to montane forests.
. . .
Computer-aided sperm morphological analysis was used to provide morphometric data on sperm heads as they traversed the epididymal duct of M. fascicularis. The duct was divided into six regions, starting close to the testis (proximal) and ending close to the vas deferens (distal). To determine the androgen dependence of the changes, one group of animals received a GnRH-antagonist (Cetrorelix, Asta Medica, Frankfurt) to induce testicular regression and lower epididymal androgens, while a control group received only vehicle. Epididymides were removed 16 and 25 days after treatment. Cluster analysis revealed swollen sperm heads in proximal regions 1 and 2 of the epididymis, but fewer such forms distally. Normal head shapes became the majority in region 4 and these underwent a gradual but statistically significant decrease in size (area, perimeter, length, width) and shape as they reached the distal regions. In the animals given Cetrorelix, sperm with swollen heads were found more distally than in the controls, although they were never present in region 6. Normal heads still became predominant in region 4 after 16 days treatment, and in region 6 after 15 days. The normal forms in the cauda epididymidis of treated animals were significantly larger than cells from control animals.
. . .
This study used noninvasive fecal hormone analysis in combination with behavioral observations to * investigate and characterize seasonal patterns of ovarian cyclicity and timing of conception in wild langur females living in a highly seasonal habitat; and * examine the relationship between seasonal patterns of ovarian cyclicity, behavioral estrus, and female physical condition. Hormone profiles in individual females revealed a clearly seasonal distribution in the occurrence of ovulatory cycles, which were restricted to the period from July to October. Distribution of female estrus behavior showed a similar seasonal pattern, and 88.2% of all estrus periods observed in the nine focal females were accompanied by ovulation. The data suggest that in these animals ecological conditions (rainfall, and food availability and quality) influence the onset of ovulations and timing of conceptions.
. . .
In this study predictions from earlier hypotheses are tested on the basis of two alternative phylogenetic trees. From this analysis we infer that helping behavior and male care preceded the origin of twinning, and that these traits did not coevolve with, but might have been important prerequisites for, twinning in callitrichines. Small body size does not necessarily result in twinning, although it might still have been a prerequisite for its evolution. Gum feeding was an ecological change which evolved along with twinning. If nutrition was a limiting factor in the number of offspring produced, then the use of a new feeding resource could have been crucial for the origin of twinning. According to one of the two alternative solutions inferred by the total evidence tree, and in accordance with the morphological tree, semi-annual breeding appears in the marmosets together with specialization in gum feeding. The fact that gums are available for these monkeys all year may have facilitated semi-annual breeding. We suggest that the exploitation of gums as a feeding resource could have been the decisive factor in the increase of the reproductive rate by twinning and by semi-annual breeding.
We would like to acknowledge Primate-Science as a source for information about new books.
---------- Announcements from Publications
International Journal of Comparative Psychology
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; FAX: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
http://www.brown.edu/Research/Primate
ACKNOWLEDGMENTS
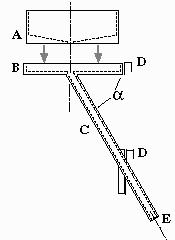{kind=link}
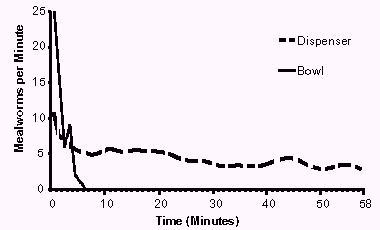{kind=link}
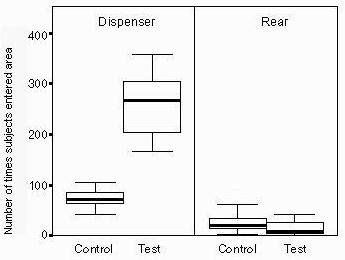{kind=link}
{kind=link}
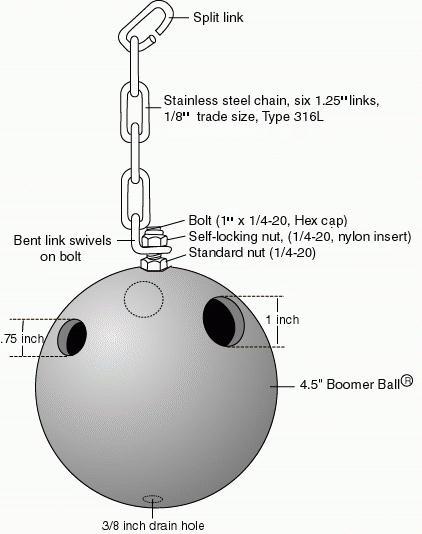{kind=link}
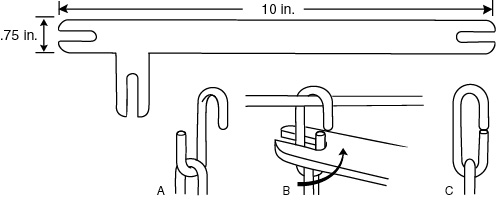{kind=link}
{kind=link}
{kind=link}