 Laboratory Primate Newsletter Laboratory Primate Newsletter
Laboratory Primate Newsletter Laboratory Primate Newsletter VOLUME 41 NUMBER 3 JULY 2002
Articles and Notes
Effectiveness of Enrichment Devices during Brief Periods of Social Restriction in Singly Housed Baboons, by R. D. Hienz, A. Jones, D. A. Pyle, & J. Johnson ......1
Digestion by Trypsin Enhances Assessment of Sperm Parameters in the Black-handed Spider Monkey (Ateles geoffroyi), by L. Hernández-López, G. Cerezo Parra, A. L. Cerda-Molina, S. C. Pérez-Bolaños, & R. Mondragón-Ceballos ......4
Addressing the Social Needs of Macaques Used for Research, by V. Reinhardt ......7
Incidence and External Morphology of the Nematode Trypanoxyuris (Hapaloxyuris) callithricis, Isolated from Black-and-Gold Howler Monkeys (Alouatta caraya) in Corrientes, Argentina, by O. H. Prieto, A. M. Santa Cruz, N. Scheibler, J. T. Borda, & L. G. Gómez ......12
How Important Are Urinary Signals in Alouatta? by C. B. Jones ......15
Multiple Drug Resistance in Rhesus Macaques of Chinese Origin ......18
News, Information, and Announcements
Workshop Announcements ......3
. . .
Seventh EMRG Workshop; Uses of Nonhuman Primates
News Briefs ......10
. . .
Deters Murdered in Thailand Gibbon Sanctuary; Need for Nonhuman Primates in Biomedical Research; Hyde Director of Davis� Primate Center; Major New Primate Discoveries in Vietnam; Gunmen Kill Mountain Gorillas in Rwanda; $2.5 Million for Gorilla Health
Traveler�s Health Notes: International Assn for Medical Assistance to Travelers ......11
Announcements from Publications ......14
. . .
Developments in Primatology: Progress and Prospects; Handbook of Animal Behavior
Meeting Announcements ......17
Volunteer Opportunity: International Center for Gibbon Studies ......17
Certified Manager of Animal Resources Examination ......18
Information Requested or Available ......19
. . .
OLDMEDLINE; PrimateLit Update; Novel Cage Card Holder; Genomics and World Health; Field Studies Guide; More Interesting Websites
Research and Educational Opportunities ......20
. . .
Postdoctoral Training, Laboratory Animal Medicine; New Opportunities - NSF; Postdoc - Duke University; Postdoctoral Fellowship - Oregon NPRC; Captive Tarsiers - Indonesia
Resources Wanted and Available ......21
. . .
Enrichment Online; ARENA/OLAW IACUC Guidebook; Leaf Monkey Loud Calls
Grants Available ......25
. . .
Neuroscience Research on Drug Addiction; Veterinary Acupuncture
Departments
Positions Available ......22
. . .
Research Associates -Washington State; Laboratory Technicians - North Carolina; Assistant Supervisor - Maryland; Clinical Veterinarian - Tulane NPRC; Environmental Enrichment Coordinator - Tulane; Research Fellow in Reproductive Biology - Florida; Director, Center for Comparative Medicine - Texas; Research Assistant - Philadelphia; Quality Assurance Officer -Madison, Wisconsin; Animal Care/Lab Tech - Stanford; Veterinarian - SFBR, Texas
Recent Books and Articles ......28
* * *
Effectiveness of Enrichment Devices during Brief Periods of Social Restriction in Singly Housed Baboons
Robert D. Hienz, April Jones, Danielle A. Pyle, and Jennifer Johnson
The Johns Hopkins University School of Medicine
Introduction
Because it is nearly impossible to recreate a natural environment for nonhuman primates in a laboratory setting, the psychological well-being of laboratory primates has become an important topic, and many articles discuss the alternatives for allowing stimulation that will keep the animal dynamic, as well as allowing species-specific behaviors to flourish (Markowitz, 1982). As a means for promoting psychological well-being, environmental enrichment has experienced great exploration since the late 1980s (Bayne et al., 1991). The use of specific enrichment devices has been the main strategy for providing stimulation in laboratories, in particular to those animals who require individual housing (Fajzi et al., 1989).
Previous studies in our laboratory have documented the use of enrichment devices by singly caged baboons, and have shown various differences in the use of food-based and non-food-based devices. For example, adult male baboons readily interact with food-based devices, which can result in the reduction of stereotypical behaviors, and in the elicitation of species-typical behaviors such as foraging and grooming (Pyle et al., 1996). On the other hand, use of non-food-related devices (e.g., logs, Kong� toys, and swings) can vary considerably among individuals, with some baboons showing decreased, others increased, use over time (Hienz, et al., 1998). Further, within individual animals the use of the logs differs across the daily circadian cycle as well as across months of exposure (Hienz et al., 2000). Enrichment device use can also vary as a function of the frequency of availability of the device: log use, for example, is enhanced when the log is available every other day or every fourth day, while activity declines if the log is withheld longer than every fourth day or if it is continuously available. Finally, movement patterns of both logs and animals tend to vary together, with the patterns strongly influenced by the presence of humans.
The present report extends this research on enrichment device use by examining log use in singly caged adult male baboons during brief periods of social restriction. Data were collected on the animals� daily biscuit intake and activity levels as well as log activity prior to, during, and following social restriction, and also in the absence and presence of a log enrichment device under these conditions.
Method
Subjects: Three adult male baboons (Papio anubis, Charles River Research Primates) were individually housed in a single colony room in regulation cages. Each baboon could view others in the colony room but did not have a view of the baboon immediately adjacent to him, which was blocked with an opaque plastic panel to reduce both physical and social aggression between immediately adjacent males. The baboons had free access to water, and were maintained on a controlled feeding and husbandry schedule. Overhead lights in the colony room provided a 12-hr. light/dark cycle (lights on/off at 6 a.m./6 p.m.). The baboons were wild-caught, and had no previous experimental histories. An Institutional Review Committee for the use of animal subjects approved the experimental protocol for the study, and the procedures were in compliance with the National Institutes of Health Guide for Care and Use of Laboratory Animals.
Apparatus: As in our previous studies (e.g., Hienz et al., 2000), activity of the baboons and movements of the logs were measured via electronic activity monitors (Personal Activity Monitor, manufactured by Individual Monitoring Systems, Baltimore, MD). Briefly, the monitor is sensitive to acceleration changes exceeding 0.1 of a gravity force, and accumulates �activity counts� over successive .88-sec bins of time. The counts are then translated into movement durations because movements typically occur over a series of bins, and each count represents a movement over a brief time interval. For the present report these movement durations were translated into a percentage of the average (baseline) activities recorded from animals and logs prior to the experimental manipulations. Hand-cut cherry hardwood logs (9 cm diameter x 35 cm long) were employed as enrichment devices, and an activity monitor was placed inside each by splitting the log, hollowing out an area inside, and rejoining the log with recessed bolts. Additionally, each baboon wore a monitor attached to a handmade neck collar of soft leather. Activity data were retrieved every two weeks from the logs, and every 42 days from the baboons. The baboons were videotaped daily (M-F). Humans were not allowed in the colony room during videotaping.
Procedure: The effects of social restriction on animal activity and enrichment device use were assessed by measuring each animal�s activity and device use for two weeks in the normal laboratory environment, and then for four weeks in a restricted social environment without an enrichment device, for two weeks in the restricted environment with an enrichment device, and then again for two weeks upon return to the normal laboratory environment. The restricted environment consisted of a separate room in which no other animals were present. Each mid-morning a technician entered the room to take data, remove and weigh any remaining biscuits, give the animal fresh biscuits and fruit, and perform normal husbandry tasks. A daily record was kept of times when the logs were manipulated by a technician (e.g., cage washing, physical exams, retrieval of monitor data, etc.) as well as when any human entered the room. Periods of human activity that may have produced non-animal-related log movements were deleted from the activity data files.
Results
Figure 1 shows that the activity of the baboons decreased by an average of about 25% when they were placed in the restricted environment, compared to the prior two-week period in the normal environment. When given access to the hardwood logs during the last two weeks, however, the baboons� activity increased and approached their pre-isolation activity levels. These decreases in activity in the restricted environment and the subsequent increases in activity when the logs were available fell outside of the 95% confidence interval about the baseline performance, and were thus significant at the p = .05 level. Additionally, these same changes occurred individually for each of the three baboons, and were not simply due to an averaging of the data. Figure 2 shows the movements of the logs recorded during the same baseline and restricted environment periods. Surprisingly, the log movements did not show any consistent increases under the restricted environment, but actually decreased by 15-20% during this period. In spite of this decreased use of the logs relative to baseline conditions, however, the baboons� general activity increased (as shown in Figure 1). Thus the baboons did not necessarily handle the logs more in the restricted environment, but the presence of the logs was still correlated with the baboons� increased activity, suggesting that the mere presence of the logs had a significant effect on returning the baboons� activity levels to near-normal. Food and water intake were also monitored over these conditions, and no consistent changes in either were observed over these periods.
Discussion
While numerous studies have examined the benefits of enrichment programs in laboratory animals, research on the effects of enrichment during brief periods of social restriction has thus far been lacking. The current study concentrated on singly caged male baboons, and clearly showed that during these brief periods of social restriction, the presence or absence of an enrichment device can have a direct impact on an animal�s general activity level. All three baboons in the current study showed a marked decrease in activity during the brief periods of social restriction when the log enrichment devices were not available. However, once these devices were provided, general activity increased again, with two of the three baboons increasing their activity levels to near-normal. On the other hand, when the enrichment device movements were examined, the baboons appeared �less interested� in the logs during the restricted condition (i.e., moved the logs less) than during the baseline period. Furthermore, two of the three baboons failed to show recovery of prior log movement levels once they were returned to the normal colony room environment (data not shown). Finally, neither the restricted environment nor the presence or absence of enrichment devices seemed to have much effect upon either the food or water intake of the baboons under these conditions.
Figure 1: Mean bodily activity of all 3 baboons recorded during the last 14 days prior (�Baseline�), the first 14 days of the Restricted condition with no log present (�Restricted, No Log�), and the last 14 days of the Restricted condition during which a log was present (�Restricted + Log�). Shown is the percentage change from the average baseline activity level. The error bars for baseline activity represent a 95% confidence interval about the mean (95% CI).
Figure 2: Mean activity movements recorded from the logs under the same conditions described in Figure 1. Shown is the percentage change from the average baseline log movements.
These findings present further support for the importance of enrichment devices for laboratory primates, showing that in the presence of such devices, the behavior of the animal is positively influenced. While the devices themselves were not manipulated greatly in this study, their presence affected the activity of the baboons. This finding adds to the already consistent wealth of research declaring the importance of enrichment programs in influencing positive behavior in laboratory animals (e.g., Byrne & Suomi, 1991; Line & Houghton, 1987). Future research on enrichment devices should concentrate on the favorable influences produced by the devices, rather than simply how much the animals handle the devices. By looking at the positive side effects of the devices, a better view of the overall benefits of an enrichment program can be understood.
References
Bayne, K., Mainzer, H., Dexter, S., Campbell, G., Yamada, F., & Suomi, S. (1991). The reduction of abnormal behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology, 23, 23-33.
Byrne, G. D., & Suomi, S. J. (1991). Effects of woodchips and buried food on behavior patterns and psychological well-being of captive rhesus monkeys. American Journal of Primatology, 20, 23-30.
Fajzi, K., Reinhardt, V., & Smith, M. D. (1989). A review of environmental enrichment strategies for singly caged nonhuman primates. Lab Animal, 18 (3), 23-35.
Hienz, R. D., Pyle, D. A., Frey, J. J., Zarcone, T. J., Adams, R. J., & Turkkan, J. S. (2000). Enrichment device use by baboons during long-term vs. intermittent availability. Laboratory Primate Newsletter, 39 [2], 1-3.
Hienz, R. D., Zarcone, T. J., Turkkan, J. S., Pyle, D. A., & Adams, R. J. (1998). Measurement of enrichment device use and preference in singly caged baboons. Laboratory Primate Newsletter, 37 [3], 6-10.
Line, S. W., & Houghton, P. (1987). Influence of an environmental enrichment device on general behavior and appetite in rhesus macaques. Laboratory Animal Science, 37 (4), 508.
Markowitz, H. (1982). Behavioural Enrichment in the Zoo. New York: Van Nostrand Reinhold.
Pyle, D. A., Bennett, A. L., Zarcone, T. J., Turkkan, J. S., Adams, R. J., & Hienz, R. D. (1996). Use of two food foraging devices by singly housed baboons. Laboratory Primate Newsletter, 35 [2], 10-15.
----------
Authors� address: Behavioral Biology Res. Ctr, Suite 3000, Johns Hopkins Bayview Med. Ctr, 5510 Nathan Shock Dr., Baltimore, MD 21224-6823 [e-mail: [email protected]].
----------
* * *
Workshop Announcements
Seventh EMRG Workshop
The 7th Workshop of the European Marmoset Research Group will be held October 14-16th, 2002, in Paris. The workshop is funded by the European Commission as a �High-Level Scientific Conference� for up to 100 participants, and there will be an emphasis on graduate and postdoctoral training in callitrichid biology and biomedical science. Topics of discussion and presentations will include: Brain structure, function and disease; Development and aging; Genetics and colony management; and Ecology and social organization. A substantial number of European Commission and EMRG training grants are available. For details of abstract submission, grant application, and registration, contact Christopher Pryce [e-mail: [email protected]]; or see <www.dpz.gwdg.de/emrg/emrgcons.htm>.
Uses of Nonhuman Primates
The Office of Laboratory Animal Welfare, NIH (OLAW) will join the Association of Primate Veterinarians (APV) and the American Association of Laboratory Animal Practitioners in co-sponsoring a workshop entitled, �Emerging Uses of Nonhuman Primates in Biomedical Research.� It will be held on October 26, 2002, immediately preceding the annual meeting of the American Association for Laboratory Animal Science in San Antonio, Texas. It will bring together top researchers from a variety of specialties using nonhuman primates in cutting-edge science. Innovative uses of these animals can pose challenges for Institutional Animal Care and Use Committees (IACUCs) reviewing animal study proposals and for veterinarians providing clinical care. Emerging scientific uses of primates will be described as well as strategies for IACUCs and veterinarians to ensure appropriate oversight and animal well-being. The program and registration information will be posted at <www.aslap.org>. For questions or further information, contact Margaret C. Quinlan, Animal Welfare Program Specialist, OLAW, NIH, 6705 Rockledge Dr., Suite 360, Bethesda, MD 20892-7982 [301-402-4325; fax: 301-402-2803; e-mail: [email protected]].
* * *
Digestion by Trypsin Enhances Assessment of Sperm Parameters
in the Black-handed Spider Monkey (Ateles geoffroyi)
Leonor Hernández-López,1 Gerardo Cerezo Parra,2 Ana Lilia Cerda-Molina,1 Stephanella C. Pérez-Bolaños,1 and Ricardo Mondragón-Ceballos1
1 Instituto Nacional de Psiquiatría Ramón de la Fuente Muñíz and Universidad Nacional Autónoma de México; 2 Instituto Nacional de la Nutrición Salvador Zubirán
Introduction
In many nonhuman primate species, including the black-handed spider monkey (Ateles geoffroyi), sperm coagulates almost entirely shortly after ejaculation. (For a list of species in which this coagulation occurs, see Dixson, 1998; for A. geoffroyi: Goodman & Wislocki, 1935; Long et al., 1997; Hernández-López et al., 2002.) This relatively large and nearly solid clot presumably acts as a physical barrier that impedes or obstructs impregnation by other males. Another theory views the coagulum as a tightly packed sperm reservoir (Dixson, 1998, p. 236).
In some species the coagulum is mainly composed of seminal plasma proteins (semenogelin I, semenogelin II, beta-microseminoprotein, etc.) and sugars (fructose) (Lundwall, 1998; Dixson, 1998). Moreover, in species such as cotton-top tamarins (Saguinus oedipus), baboons (Papio cynocephalus), rhesus monkeys (Macaca mulatta), and marmosets (Callithrix jacchus), the genes that code for the proteins that form the seminal plasma as well as the coagulum are known (Marjaana, et al., 1999). In practice, the presence of the coagulum hinders assessment of sperm parameters. Analysis of the liquid portion yields poor sperm counts compared to species which are related or of equivalent body size (see Long et al., 1997); this makes interspecies comparisons, medical evaluations, and assessment of sperm quality in assisted reproduction techniques all difficult.
Digestion of the coagulum with proteases, such as trypsin, has been tried (Hendrickx & Kreamer, 1970), but the practice has been mostly abandoned because trypsin digests the membranes of spermatozoa (P. Nayudu, personal communication), which is very undesirable for either assisted reproduction or precise assessment of sperm quality. Nonetheless, lacking other nonharmful liquefying techniques, enzymatic digestion is still a tool to unravel the cellular composition of the coagulum.
As stated above, black-handed spider monkey ejaculate is characterized by the presence of a very large coagulum (ca. 90% of the volume). Coagulation starts almost as soon as the epididymal fluid joins the prostate gland and seminal fluids, occurring both naturally (e.g., when males masturbate) or during electroejaculation (Long et al., 1997; Hernández-López et al., 2002). Assessment of the remaining liquid yields low sperm concentration values, as well as low motility measures and a rather large number of dead sperm (personal observations). Therefore, the purpose of this study was to reevaluate the effects of trypsin digestion on sperm quality in the coagulum of the spider monkey.
Methods
The subjects were three adult male black-handed spider monkeys (LK, KI and AD). They form part of a social group (four females and three males at the time of the study) kept in a large outdoor facility at the Ethology Department of the Instituto Nacional de Psiquiatría, in Mexico City. All animals are unrelated to each other; details concerning their origins and housing conditions have been published elsewhere (Anaya-Huertas & Mondragón-Ceballos, 1998). Seventy-five semen samples were collected by electroejaculation from anesthetized spider monkeys (Long et al., 1997; Hernández-López et al., 2002) from January 1999 to August 2000. Of these, 44 samples included both the solid and liquid phases, unpolluted by urine. We tried two methods to retard coagulation. The first (8 samples from AD, 9 from KI, 7 from LK) was stirring the sperm manually and continually for 10 min, keeping it immersed in a 37° C water-bath. The second method (7 samples from AD, 6 from KI, 7 from LK) was to add a phosphate-buffered saline containing 0.25% trypsin (Sigma T4799; Sigma Inc., U.S.A.), at a 2:3 (trypsin solution: sperm) proportion, while keeping it immersed in a 37° C water-bath for 10 min. In both methods, samples were evaluated immediately afterwards.
Sperm analysis was done following the criteria of the World Health Organization (1992) for the assessment of human semen, because it has been successfully used in other nonhuman primate species (e.g., Macaca fascicularis: Gago, et al., 1999). Volume was measured directly from graduated Elkay tubes (+/- 0.5 ml of accuracy). Motility was divided into four categories: fast linear, slow linear, nonlinear, and immobile (definitions are given in Hernández-López et al., 2002). Abnormal spermatozoa were considered those having only the lower half of the head, two heads, a bent mid-piece, or a looped or coiled flagellum. Eosin B staining was used to estimate the percentages of live and dead spermatozoa (Martin & Gould, 1981). Each sperm measure was obtained by averaging the counts of five different fields from a 10microl. sample/animal. A different 10microl. sample was used to assess each of the seven sperm parameters analyzed. One person (G.C.P.) did all the microscope evaluations. Readings from the trypsin-digested samples were corrected for volume prior to analyses. The sperm treatment effects were assessed by means of split-plot ANOVAs, because the subjects sampled were too few for repeated-measures ANOVAs. Subjects (animals) were a random-effects factor; treatments per animal, a fixed-effects factor; and readings within treatments, the repeated measures.
| Animal | Spermatozoa (millions/ml of semen) Stir -- Tryp |
% Fast linear movement Stir -- Tryp |
% Slow linear movement Stir -- Tryp |
% Nonlinear movement Stir -- Tryp |
% Immobile Stir -- Tryp |
% Live sperm Stir -- Tryp |
% Abnormal sperm Stir -- Tryp |
| AD | |||||||
| Mean | 32.1 -- 39.8 | 6.3 -- 11.3 | 10.1 -- 44.1 | 2.5 -- 3.6 | 83.8 -- 41.3 | 61.7 -- 74.7 | 39.4 -- 38.3 |
| S.E. | 14.7 -- 13.6 | 4.1 -- 3.8 | 8.4 -- 7.8 | 1.6 -- 1.5 | 9.5 -- 8.8 | 9.9 -- 9.2 | 4.4 -- 4.1 |
| KI | |||||||
| Mean | 40.5 -- 52.6 | 15.9 -- 35.2 | 19.6 -- 32.5 | 3.1 -- 7.1 | 47.9 -- 27.9 | 68.9 -- 83.8 | 38.4 -- 38.0 |
| S.E. | 12.0 -- 15.3 | 3.4 -- 4.3 | 6.9 -- 8.7 | 1.3 -- 1.7 | 7.8 -- 9.9 | 8.1 -- 10.3 | 3.9 -- 4.6 |
| LK | |||||||
| Mean | 73.2 -- 92.8 | 11.9 -- 16.0 | 16.1 -- 42.0 | 12.6 -- 7.7 | 61.9 -- 35.0 | 50.7 -- 79.3 | 28.6 -- 39.4 |
| S.E. | 14.7 -- 13.6 | 4.1 -- 3.8 | 8.4 -- 7.8 | 1.6 -- 1.5 | 9.5 -- 8.5 | 9.9 -- 9.2 | 4.4 -- 4.1 |
| Total mean | 48.6 -- 61.7 | 11.3 -- 20.8 | 15.3 -- 39.5 | 6.1 -- 6.1 | 64.5 -- 34.7 | 60.4 -- 79.3 | 35.5 -- 38.6 |
| S.E. | 8.2 -- 8.2 | 2.3 -- 2.3 | 4.7 -- 4.7 | 0.9 -- 0.9 | 5.3 -- 5.3 | 5.5 -- 5.5 | 2.5 -- 2.5 |
| Fa | 9.908 | 7.059 | 27.391 | 0.000 | 35.049 | 22.306 | 1.212 |
| P | 0.034 | 0.056 | 0.006 | N.S. | 0.004 | 0.01 | N.S. |
Table I: Comparisons of manual stirring and trypsin digestion of sperm on concentration, motility, viability, and presence of abnormal spermatozoa in ejaculates from three Ateles geoffroyi monkeys. S.E. = Standard Error. a Between-treatments F-ratios; d.f. (calculated by Satterwaite�s method) ~= 4
Table I displays the sperm parameters and statistical analyses. Trypsin digestion significantly increased the concentration of spermatozoa, as well as the percentages of slow-moving and live spermatozoa, while significantly decreasing the percentage of immobile sperm. The percentage of fast-moving sperm showed a tendency to increase due to trypsin digestion. The proportion of nonlinear moving and abnormal sperm were unaffected by trypsin treatment.
Discussion
Despite within- and between-individual variability, readings from trypsin-digested samples showed an overall improvement over stirred samples. We assume that trypsin proved useful because we did not wait until the coagulum was completely digested, but only until the sample was liquefied enough to allow a convenient assessment. Therefore, most likely we did not give the trypsin time to start digesting the spermatozoa membranes.
The sperm concentration of hand-stirred samples was on average lower than values reported for other primate species (Hendrickx & Kraemer, 1970; Martin & Gould, 1981; Gould & Mann, 1988; Gould et al., 1993), including smaller animals like the capuchin and the squirrel monkey (Hendrickx & Kraemer, 1970). Trypsin digestion significantly increased the sperm concentrations (see Table I) to the ranges of species with naturally low sperm concentrations such as the capuchin, squirrel monkey, gibbon (Hendrickx & Kraemer, 1970), and gorilla (when urethral massage was added to the electroejaculation procedure: Schaffer et. al., 1991). The additional sperm cells were undoubtedly released from the coagulum by the trypsin, as occurs in other primate species (Wildt, 1986). Therefore, these values are viewed as more representative of the true sperm concentration for this species.
The coagulum most certainly is the vaginal plug reported by Goodman and Wislocki (1935), in their pioneering research on the sexual biology of the black-handed spider monkey. It most surely serves to prevent backflow of sperm and to obstruct sperm transport of another male (Dixson, 1998). Nonetheless, taking in account the peculiar anatomy of the spider monkey glans penis (Hill, 1962; Dixson, 1998) and the elongated cervix that protrudes into the vaginal lumen (Hill, 1962; Hernández-López & Aja, personal communication), the coagulum might also serve as a vehicle to deliver a tight, rich package of spermatozoids into the female�s cervix.
The improvements over stirring in other trypsin-treated sperm measures, such as the measures of motility and percentage of live cells, were possibly due to the capacitating properties of phosphate-buffered saline, but, as stated above, we ought not to discount the fact that these cells were released from the coagulum. The summed fast and slow linear motility values fall within motility measures reported in other primate species (Hendrickx & Kraemer, 1970; Martin & Gould, 1981; Gould & Mann, 1988; Gould, 1990; Gould et al., 1993; Schaffer et al., 1991), showing that neither the electroejaculation procedure nor trypsin digestion yield greater than expected adverse effects on the spider monkeys� ejaculates (Gould & Mann, 1988). Likewise, as is usual with rectal electroejaculation, our samples seemingly have a poorer viability than those obtained by penile electrical stimulation (Gould & Mann, 1988) or vibratory stimulation (Yeoman et al., 1998). However, the percentage of live spermatozoids ejaculated, whether motile or not, was similar to those of great apes (Yeoman et al., 1998) and reproductively healthy humans (World Health Organisation, 1992), and above required values for artificial insemination in nonhuman primates (Hendrickx & Kraemer, 1970) and humans (Karabinus & Gelety, 1997).
References
Anaya-Huertas, C., & Mondragón-Ceballos, R. (1998). Social behavior of black-handed spider monkeys (Ateles geoffroyi) reared as home pets. International Journal of Primatology, 19, 767-783.
Dixson, A. F. (1998). Primate sexuality: Comparative studies of the prosimans, monkeys, apes, and human beings. Oxford: Oxford University Press.
Gago, C., Pérez-Sánchez, F., Yeung, C. H., Tablado, L., Cooper, T. G., & Soler, C. (1999). Morphological characterization of ejaculated cynomolgous monkey (Macaca fascicularis) sperm. American Journal of Primatology, 47, 105-115.
Goodman, L., & Wislocki, G. (1935). Cyclical uterine bleeding in a New World monkey (Ateles geoffroyi). The Anatomical Record, 61, 379-387.
Gould, K. G. (1990). Techniques and significance of gamete collection and storage in the great apes. Journal of Medical Primatology, 19, 537-551.
Gould, K. G., & Mann, D. R. (1988). Comparison of electroestimulation methods for semen recovery in the rhesus monkey (Macaca mulatta). Journal of Medical Primatology, 17, 95-103.
Gould, K. G., Young, L. G., Smithwick, E. B., & Phythyon, S. R. (1993). Semen characteristics of the adult male chimpanzee (Pan troglodytes). American Journal of Primatology, 29, 221-232.
Hendrickx, A. G., & Kraemer, D. C. (1970). Primates. In E. S. E. Hafez (Ed.), Reproduction and breeding techniques for laboratory animals (pp. 317-335). Philadelphia: Lea & Febiger.
Hernández-López, L., Cerezo Parra, G., Cerda-Molina, A. L., Pérez-Bolaños, S. C., Díaz Sánchez, V., & Mondragón-Ceballos, R. (2002). Sperm quality differences between the rainy and dry seasons in captive black-handed spider monkeys (Ateles geoffroyi). American Journal of Primatology, 57, 35-41.
Hill, O. W. C. (1962). Ateles geoffroyi. In O. W. C. Hill (Ed.), Comparative anatomy and taxonomy. Vol. V. Cebidae, part B (pp.465-475). Edinburgh: Edinburgh University Press.
Karabinus, D. S., & Gelety, T. J. (1997). The impact of sperm morphology evaluated by strict criteria on intrauterine insemination success. Fertility and Sterility, 67, 536-541.
Long, J. A., Lambeski, N., & Shoemaker, A. H. (1997). Ejaculate characteristics of the black handed spider, southern black howler and Diana monkey. Journal of Andrology (Suppl. 41).
Lundwall, Å. (1998). The cotton-top tamarin carries an extended semenogelin I gene but no semenogelin II gene. European Journal of Biochemistry, 255, 45-51.
Marjaana, M., Valtonen-André, C., & Lundwall, Å. (1999). New World, but not Old World, monkeys carry several genes encoding beta-microseminoprotein. European Journal of Biochemistry, 264, 407-414.
Martin, D. E., & Gould, K. G. (1981). The male ape genital tract and its secretions. In C. E. Graham (Ed.), Reproductive biology of the great apes (pp. 127-162). New York: Academic Press.
Schaffer, N., Jeyendran, R. S., & Beehler, B. (1991). Improved sperm collection from the lowland gorilla: Recovery of sperm from bladder and urethra following electroejaculation. American Journal of Primatology, 24, 265-271.
Wildt, D. E. (1986). Spermatozoa: Collection, evaluation, metabolism, freezing, and artificial insemination. In: W. R. Dukelow & J. Erwin, (Eds.), Comparative Primate Biology (pp. 171-193). New York: Alan R. Liss.
Yeoman, R. R., Sonkesen, J., Gibson, S. V., Rizk, B. M., & Abee, C. R. (1998) Penile vibroestimulation yields increased spermatozoa and accessory gland production compared with rectal electroejaculation in a neurologically intact primate (Saimiri bolivensis). Human Reproduction, 13, 2527-2531.
----------
First author�s address: Instituto Nacional de Psiquiatr�a Ram�n de la Fuente, Subdirecci�n Neurociencias, Calzada M�xico-Xochimilco 101, Col. San Lorenzo Huipulco, Tlalpan 14370, M�xico, D.F., M�xico [e-mail: [email protected]].
----------
* * *
Addressing the Social Needs of Macaques Used for Research
Viktor Reinhardt
Animal Welfare Institute
Rhesus macaques are the predominant primate species found in research institutions. They are social animals. Federal rules, therefore, stipulate that the institution�s environmental enhancement plan must include specific provisions to address their �social needs� (United States Department of Agriculture, 1991, p. 6497). Unfortunately, this basic requirement is usually not met. �The common practice of housing rhesus monkeys singly calls for special attention� (National Research Council, 1998, p. 99) because solitary confinement is a serious stressor leading to apathy and depression (Erwin & Deni, 1979; Lilly et al., 1999), often manifested as behavioral pathologies such as self-biting (Jorgensen et al., 1998). Many scientists, veterinarians and colony managers, however, are resisting any change in traditional individual-caging practices, arguing that social housing would interfere with research and husbandry, jeopardize the animals� health and well-being, and cost too much money. To the knowledge of this author there is no published report on the practicability of large-scale social housing of macaques used for research. The present paper summarizes the author�s experiences transferring a colony of rhesus macaques (Macaca mulatta) from individual- to pair-caging.
Implementing Pair-Housing
Pair-caging was implemented as standard housing for rhesus macaques at the Wisconsin Regional Primate Research Center in 1987. Same-sex pairs of 4- to 34-year-old adults were formed by allowing partners first to establish dominance-subordinance relationships during a brief non-contact familiarization period (to avoid rank-determining fighting at the moment of introduction), and then introducing them in a different (to avoid territorial antagonism) double cage. Isosexual pairs were also formed by introducing 2- to 3-year-old juveniles to one another, or by introducing naturally weaned, 12- to 18-month-old infants to singly caged adults without prior familiarization (Reinhardt et al., 1987; Reinhardt, 1989; 1991). Pairs were considered to be compatible as long as: * no animal was seriously injured (requiring surgical treatment) during an aggressive dispute; * no animal showed signs of depression; and * both partners secured an adequate amount of the daily food ration and food treats.
Adult male pairs were kept in strictly male-only areas to forestall aggressive conflicts arising from sexual competition. Animals who permanently lost their partners for experimental, managerial or incompatibility reasons were re-paired with other conspecifics as soon as possible. Depending on the subjects� legal space requirements, most pairs were housed in horizontally arranged double cages, each provisioned with a privacy panel offering the option of visual seclusion during feeding time (Reinhardt et al., 1991a). All cages were equipped with such a privacy panel, two perches, and two gnawing sticks. At a census conducted in March, 1994, the Center�s caged rhesus colony comprised 728 animals. Of these, 644 (88.5%) lived in compatible pairs, 24 (3.3%) lived in kindergarten groups, and 60 (8.2%) lived alone (Reinhardt, 1994).
Between March, 1987, and March, 1993, a total of 726 pairs were formed involving 817 different animals. There were also several kindergarten groups, consisting of one or two adults and several weaned infants housed in gang cages (four interconnected standard cages). These latter groups are not included in the present report.
The following information was recorded and analyzed in retrospect: * Date of pair formation (six-year test period: March, 1987 to March, 1993); * Composition of pairs; * Compatibility of partners; * Physical and behavioral health status; � Research assignment; * Date of and reason for pair separation (seven-year test period: March, 1987 to March, 1994); * Number of days partners lived together; and � Number of days individual animal waited to be re-paired after having lost her/his partner.
Of the 726 pairs tested, 671 (92%) were compatible. Signs of incompatibility were observed in 33 (5%) cases shortly after pair formation, and in 22 (3%) cases 17-1168 days after the pair was originally formed. Of the 817 animals a total of 6 (0.7%) were inflicted with serious, but not life-threatening, lacerations resulting from aggressive disputes.
There was a trend for pairs with adult females to be more compatible than pairs with adult males, and juvenile pairs had a tendency to be more compatible than pairs with one or two adults. These differences were negligible and possibly related to different sample sizes (Table 1). Paired companions expressed their social disposition by engaging in affiliative social interactions (e.g., grooming, huddling, playing) approximately 20% of the day (Reinhardt, 1990a; Reinhardt & Hurwitz, 1993).
Seven individuals exhibited self-injurious stereotypies in the single-caging housing arrangement. Transfer to compatible pair-housing stopped this behavioral disorder (Reinhardt, 1999; cf. Line et al., 1990; Weed et al., 2002).
When compared with singly caged individuals, pair-housed animals did not differ in terms of body weight development (Reinhardt et al., 1988; cf., Eaton et al., 1994), serum cortisol concentration (Reinhardt et al., 1991b; cf., Schapiro et al., 1993), and frequency of veterinary treatments (Reinhardt, 1990b; cf., Schapiro & Bushong, 1994).
| Pair composition | Number of pairs tested -- compatible | Mean number of days partners lived together |
| Adult female/female | 333 -- 307 (92%) | 685 days |
| Adult male/male | 67 -- 58 (87%) | 513 days |
| Adult female/infant | 201 -- 189 (94%) | 531 days |
| Adult male/infant | 83 -- 75 (90%) | 436 days |
| Juvenile/juvenile | 42 -- 42 (100%) | 582 days |
| Total/Average | 726 -- 671(92%) | 602 days |
Table 1: Characteristics of 727 rhesus macaque pairs formed during an observation period of six years.
During the seven-year observation period, compatibility of pairs was confirmed for up to 7 years in adult female/female pairs, and up to 4.7 years in adult male/male pairs. Compatibility was ascertained up to 3.9 and 3.8 years in adult female/infant and adult male/infant pairs, respectively. The young were then about 5 years old and had reached the age of sexual maturity. This did not affect their compatibility with their adult partners. Compatibility of juvenile/juvenile pairs could be traced up to 3.5 years. By then the partners had reached full sexual maturity, which did not affect their compatibility.
Pair-housing did not interfere with common husbandry procedures such as visual inspection, capture, weighing, TB-testing, and intramuscular injection; managerial procedures such as timed breeding (Figure 1); or common research procedures such as blood collection, urine collection (partners separated by grated partition during collection period), oral and systemic drug administration (partners separated by grated partition during procedure), remote blood collection via tether (tested in adult-infant pairs only), food intake studies (partners are separated prior to food distribution and reunited after removal of left-over food), and neurophysiological studies requiring headcap implants (Reinhardt & Reinhardt, 2002).
Compatible companions lived together for an average of 602 days (Table 1). Companions lost their partners on 460 occasions (e.g., for segregation of SRV-positive and SRV-negative animals). In 183 instances re-pairing was precluded for IACUC-approved reasons. In 277 instances re-pairing with another companion was possible. These individuals were re-paired with other suitable partners on average 11.5 days after they had lost their companions. In 123 (44%) of these cases individuals received a new compatible cage mate within 24 hours; in 154 (56%) cases individuals had to wait up to 58 days because no suitable partner or no suitable cage was available.
Figure 1: Raising offspring does not affect compatibility of female pairs. A time-mated mother with her baby shares apples with the cage companion with whom she has lived for three years.
Expenses
The cost of the pair-housing program involved work time for the establishment and monitoring of new pairs and expenses for the double cages and privacy panels.
A cumulative total of about 30 minutes was invested to transfer each pair of singly caged animals to pair-housing and verify that the new pair was compatible (Reinhardt & Reinhardt, 2000). After compatibility was ascertained, pairs were checked during daily routine inspections.
Most of the Center�s monkey cages were conventional, horizontally arranged twin modules; a few were single hanging-cages. Double cages were created in over 300 instances at no cost by removing the cage dividers. The dividers were then converted into privacy panels for $5 to $10 each by cutting a passage hole close to the back (Reinhardt & Reinhardt, 2001, photos 47 & 48). Double cages were created in six instances by interconnecting two hanging-cages with short tunnels (Reinhardt & Reinhardt, 2001, photos 45 & 46). The expense for the preparation of one tunnel was $100-150. Each tunnel also served as passage �hole� for the privacy �panel� -the two parallel cage walls - at no extra cost.
Conclusions
The present report demonstrates that the social needs of rhesus macaques can readily be met in the research institution�s setting by allowing the animals to live in compatible pairs. The implementation of pair-housing as standard caging at the Wisconsin RPRC was not excessively expensive in terms of work time and material. It was not a hindrance to active research; rather it improved the animals� behavioral health and made them more species-representative models for research. A total of 726 pairs were formed involving 817 different animals, yet no indication was found that this rendered research data collected from them useless due to confounding variables such as stress or disease. On the contrary, there is evidence suggesting that living with a compatible companion rather than alone positively affects immune responses (Schapiro et al., 2000). This could be one reason why pair-housed rhesus macaques tend to require veterinary treatment less often than single-housed individuals (Reinhardt, 1990b; Schapiro & Bushong, 1994).
The findings of this report question the reservations against social caging of rhesus macaques in research but strongly endorse the National Research Council�s advice that �every effort should be made to house these animals socially (in groups or pairs), but when this is not possible, the need for single housing should be documented by investigators and approved by the IACAC. ... There are reasons for (temporary) single housing, that should not be accepted as the default situation� (National Research Council, 1998, p. 99).
References
Eaton, G. G., Kelley, S. T., Axthelm, M. K., Iliff-Sizemore, S. A., & Shiigi, S. M. (1994). Psychological well-being in paired adult female rhesus (Macaca mulatta). American Journal of Primatology, 33, 89-99.
Erwin, J., & Deni, R. (1979). Strangers in a strange land: Abnormal behavior or abnormal environments? In J. Erwin, T. Maple, & G. Mitchell (eds), Captivity and Behavior (pp. 1-28). New York, NY: Van Nostrand Reinhold.
Jorgensen, M. J., Kinsey, J. H., & Novak, M. A. (1998). Risk factors for self-injurious behavior in captive rhesus monkeys (Macaca mulatta). American Journal of Primatology, 45, 187.
Lilly, A. A., Mehlman, P. T., & Higley, J. (1999). Trait-like immunological and hematological measures in female rhesus across varied environmental conditions. American Journal of Primatology, 48, 197-223.
Line, S. W., Morgan, K. N., Markowitz, H. Roberts, J. A. & Riddell, M. (1990). Behavioral responses of female long-tailed macaques (Macaca fascicularis) to pair formation. Laboratory Primate Newsletter, 29 [4], 1-5.
National Research Council (1998). The Psychological Well-Being of Nonhuman Primates. Washington, DC: National Academy Press.
Reinhardt, V. (1989). Behavioral responses of unrelated adult male rhesus monkeys familiarized and paired for the purpose of environmental enrichment. American Journal of Primatology, 17, 243-248.
Reinhardt, V. (1990a). Time budget of caged rhesus monkeys exposed to a companion, a PVC perch and a piece of wood for an extended time. American Journal of Primatology, 20, 51-56.
Reinhardt, V. (1990b). Social enrichment for laboratory primates: A critical review. Laboratory Primate Newsletter, 29 [3], 7-11.
Reinhardt, V. (1991). Social enrichment for aged rhesus monkeys that have lived singly for many years. Animal Technology, 43, 173-177.
Reinhardt, V. (1994). Pair-housing rather than single-housing for laboratory rhesus macaques. Journal of Medical Primatology, 23, 426-431.
Reinhardt, V. (1999). Pair-housing overcomes self-biting behavior in macaques. Laboratory Primate Newsletter, 38 [1], 4.
Reinhardt, V., Houser, W. D., Eisele, S., & Champoux, M. (1987). Social enrichment with infants of the environment for singly caged adult rhesus monkeys. Zoo Biology, 6, 365-371.
Reinhardt, V., Cowley, D., Eisele, S., Vertein, R., & Houser, W. D. (1988). Pairing compatible female rhesus monkeys for the purpose of cage enrichment has no negative impact on body weight. Laboratory Primate Newsletter, 27 [1], 13-15
. Reinhardt, V., Pape, R., & Zweifel, D. (1991a). Multifunctional cage for macaques housed in pairs or in small groups. American Association for Laboratory Animal Science Bulletin [Contemporary Topics in Laboratory Animal Science], 30[5], 14-15.
Reinhardt, V., Cowley, D., & Eisele, S. (1991b). Serum cortisol concentrations of single-housed and isosexually pair-housed adult rhesus macaques. Journal of Experimental Animal Science, 34, 73-76.
Reinhardt, V., & Hurwitz, S. (1993). Evaluation of social enrichment for aged rhesus macaques. Animal Technology, 44, 53-57.
Reinhardt, V., & Reinhardt, A. (2001). Environmental Enrichment for Caged Rhesus Macaques (Macaca mulatta) - Photographic Documentation and Literature Review (Second Edition). Washington, DC: Animal Welfare Institute.
Schapiro, S. J., Bloomsmith, M. A., Kessel, A. L., & Shively, C. A. (1993). Effects of enrichment and housing on cortisol response in juvenile rhesus monkeys. Applied Animal Behaviour Science, 37, 251-263.
Schapiro, S. J., & Bushong, D. (1994). Effects of enrichment on veterinary treatment of laboratory rhesus macaques (Macaca mulatta). Animal Welfare, 3, 25-36.
Schapiro, S. J., Nehete, P. N., Perlman, J. E., & Sastry, K. J. (2000). A comparison of cell-mediated immune responses in rhesus macaques housed singly, in pairs, or in groups. Applied Animal Behaviour Science, 68, 67-84.
Weed, J. L., Byrum, R., Parrish, S., Knezevich, M., Powell, D. A., & O�Neill Wagner, P. L. (2002). Vasectomies as part of an environmental enrichment plan for primates. 25th Meeting of the American Society of Primatologists, Abstract #40.
----------
Author�s address: 6014 Palmer Drive, Weed, CA 96094; [e-mail: [email protected]].
I am very grateful for the excellent cooperation of the animal care personnel of the Wisconsin Regional Primate Research Center, in particular to Tammie Frost, Doug Cowley, Russell Vertein, Steve Eisele, and Harry Pape. Their genuine concern for the well-being of the animals in their charge was instrumental in the successful implementation of the new housing arrangement of the center�s caged rhesus macaque colony. This project was partly supported by NIH grant RR-00167.
----------
* * *
News Briefs
Deters Murdered in Thailand Gibbon Sanctuary
William Deters, 63, was shot dead on May 10, in the kitchen of his Highland Farm and Wildlife Refuge, 260 miles northwest of Bangkok. Bill and his Thai wife, Pharanee, opened the sanctuary in 1991, creating a refuge for abandoned, mistreated, or injured gibbons. The 35-acre farm is located about 25 miles south of Mae Sot, a major trading town on Thailand�s border with Myanmar. Deters� wife was not at the farm at the time of the murders.
Police said May 12 that they have arrested a man who allegedly killed Deters and four other people. Officers said Maung Htwe, 19, a former employee of the Refuge, had confessed to the killings, which were allegedly the result of a botched burglary attempt. He was arrested in a police raid the night of May 11.
The other victims were two men and a woman who worked at the farm, and the 3-year-old daughter of one of them. A Dutch volunteer worker at the sanctuary survived the attack by hiding in a bathroom. Police allege that Maung Htwe sneaked into a building on the farm and took six or seven guns, but encountered the 63-year-old Deters and the other victims as he was leaving, and shot them. - From an Associated Press report posted to Alloprimate
Need for Nonhuman Primates in Biomedical Research
The Scientific Steering Committee of the European Commission�s Health and Consumer Protection Directorate-General issued a Statement adopted at its meeting of April 4-5, 2002, on the implications of a complete ban on nonhuman primate research facilities. This statement concludes that �for certain experiments there are no alternatives to the use of nonhuman primates. Such experiments may be needed, for example, during the development of drugs and vaccines for prevention and cure of diseases� such as AIDS, malaria, transmissible spongiform encephalopathies, and influenza.
Hyde Director of UC Davis� Primate Center
Dallas Hyde, an authority on the biology of asthma and other lung diseases, has been named as Director of the California Regional Primate Research Center at the University of California at Davis. Hyde, a professor in the School of Veterinary Medicine, has served as interim director of the center since 2000, and as associate dean for research and graduate education for the veterinary school since 1997. - From the Woodland Daily Democrat, March 9, 2002
Major New Primate Discoveries in Vietnam
Two important primate discoveries in Vietnam have reinforced the importance of this region as a vital �hotspot� of primate species diversity, but have also confirmed that many of these species are staring straight into the face of extinction. Conservation biologists from Fauna & Flora International (FFI) and the government of Vietnam have discovered two new populations of the critically endangered Eastern black crested gibbon (Nomascus nasutus nasutus). Previously only known from Hainan Island in China, these discoveries double the known world population, estimated to no more than 30 gibbons - approximately half surviving in northern Vietnam. Fourteen remain on Hainan Island.
Sound recordings proved the gibbons� identity, and confirmed that at least eight individuals survive in the tiny forest fragment in Cao Bang on the Chinese border. An additional group of six gibbons has also been reported in Bac Kan by local hunters, but four months of intensive survey by FFI has not yet confirmed their presence. Local hunters claim the hunting pressure is so high that the gibbons have given up singing. With a global population of less then 30 individuals, the Eastern black crested gibbon is undoubtedly facing extinction within a few years without immediate action.
The Tonkin snub-nosed monkey (Rhinopithecus avunculus) is a critically endangered species, endemic to Vietnam, and found only in Tuyen Quang, Ha Giang, and Bac Can Provinces. A two-year study by FFI biologist Le Khac Quyet resulted in the discovery a new population of 50-60 Tonkin snub-nosed monkeys in Ha Giang. His discovery increases the known population from less then 200 to less than 260 individuals, and was verified by his photographs of these secretive endemic primates in the wild. The other 200 individuals are spread between the Nahang Nature Reserve and Cham Chu Nature Reserve, established in 2000-2001 by the Ministry of Agriculture and Rural Development specifically to protect Tonkin snub-nosed monkeys, with assistance from the Vietnamese Institute for Ecology and Biological Resources and FFI. - from a Flora and Fauna International news release, April 3rd, 2002
Gunmen Kill Mountain Gorillas in Rwanda
Gunmen killed two mountain gorillas as they nursed their young and stole one of the babies, Rwandan police said. Two suspects have been arrested and more were being sought. Poachers attacked the endangered animals at the Virunga National Park after rangers left the park, said Francois Bizimungu, senior conservation officer at the park.
The two suspects have admitted that they were seeking to grab baby gorillas to sell. �This cannot be an operation mounted by a couple of guys. We suspect there is a big ring behind the two men in custody,� said Solange Katarebe, director of the Rwandan National Parks and Tourism Office. It was the first known attack on the rare gorillas in a year. In June, Rwandan rebels killed two young mountain gorillas, apparently for food.
Bizimungu said by telephone from Ruhengeri, the base of the Virunga Mountains, 40 miles northeast of the Rwandan capital, Kigali, that the gunmen probably attacked when the two adult females were separated from the rest of the family group. �The killings were discovered by rangers the next day when routine checks revealed the body of the first gorilla with bullet wounds, minus her 2-year-old.� A second young gorilla was left behind, probably because the dominant male in the family moved in to help his mate.
Some 350 of the mountain gorillas remaining in the wild are found in the Virunga Mountains straddling Rwanda, Uganda and Congo. The remaining 320 of the subspecies live in a separate national park in Congo. Rwandan authorities and researchers closely monitor poaching and welfare of the rare gorillas on the park�s lush volcanic slopes.
�Because the population is so fragile and the two parks are not connected, the loss of every individual gorilla is significant in terms of genetic viability and long-term survival of the mountain gorilla,� said Katie Fawcett, director of a gorilla research center run by the Dian Fossey Gorilla Fund International. Gorilla tourism is Rwanda�s third highest hard currency earner after tea and coffee exports. After a four-year civil war and the 1994 genocide, tourists are just beginning to return to Rwanda. - By Rodrique Ngowi, Associated Press Writer, reported on Alloprimate May 13, 2002
$2.5 Million For Gorilla Health
Uganda, Rwanda, and the Democratic Republic of Congo have in the last one-and-half decade benefited from a grant of US$2.5m for treatment of the endangered mountain gorillas. Michael Cranfield of the Morris Animal Foundation, a U.S. donor agency, said they had been assisting the three countries, which have the 650 mountain gorillas in the world. He said the countries were earning a lot of money through gorilla tourism and there was an ethical responsibility to protect the animals. �That is why we consider their health a priority,� he said.
Dr. Joseph Okori of the Uganda Wildlife Authority (UWA) said they were working with the Makerere University�s Wildlife Department under the Faculty of Veterinary Medicine to improve gorilla health. - by Gerald Tenywa, from New Vision (Kampala), May 2, 2002
* * *
Travelers� Health Notes: International Assn for Medical Assistance to Travelers
The International Association for Medical Assistance to Travelers (IAMAT), a volunteer group, compiles an annual list of doctors around the world who meet the organization�s criteria, who speak English or another second language, and who agree to charge a specific fee. The 2002 Directory lists the current schedule of fees as US$55 for an office visit, US$75 for a house (or hotel) call, and US$95 for night, Sunday, and holiday calls. These fees do not include consultants, laboratory procedures, hospitalization, or other expenses. The current listing of doctors and centers includes 125 countries. IAMAT also publishes and provides to its members pamphlets on immunization and on health risks, including malaria. IAMAT has now established a scholarship program for physicians from developing countries to attend travel medicine training courses in North America. For information, contact IAMAT, 40 Regal Rd, Guelph, Ontario, N1K 1B5, Canada [519-836-0102]; 417 Center St, Lewiston, NY 14092, U.S.A. [716-754-4883]; P.O. Box 5049, Christchurch 5, New Zealand; or 57 Voirets, 1212 Grand-Lancy-Geneva, Switzerland [e-mail: [email protected]]; or see <www.iamat.org>.
* * *
Incidence and External Morphology of the Nematode Trypanoxyuris (Hapaloxyuris) callithricis, Isolated from Black-and-Gold Howler Monkeys (Alouatta caraya) in Corrientes, Argentina
O. H. Prieto1, A. M. Santa Cruz2, N. Scheibler1, J. T. Borda2, and L. G. Gómez2
Universidad Nacional de Buenos Aires1 and Universidad Nacional del Nordeste, Corrientes 2
The black-and-gold howler monkey, Alouatta caraya (in Spanish: mono carayá), is found in the rain forests of Argentina�s Paranaense and Chaqueña biogeographic regions (Cabrera & Willink, 1973). These primates are listed in Appendix II of the Convention on International Trade in Endangered Species (CITES), and their indiscriminate capture for the pet trade is causing serious problems for their conservation. Many of these animals end up in zoos when their owners tire of them or can�t deal with their increased size and aggressiveness at sexual maturity. The parasites affecting them have been little studied, but acquire greater importance when they are kept captive, thereby becoming reservoirs of parasitic zoonoses.
Oxyuriosis in primates is a parasitic disease caused by three genera of nematodes of the family Oxyuridae (Hugot, 1984, 1985; Inglis & Cosgrove, 1965; Masi Pallares, 1990; Petter & Quentin, 1974; Yamaguti, 1961; Yorke & Maplestone, 1926). These parasites have been little studied in Argentina (Moriena, et al., 1979; Pope, 1966) and have usually been found by necropsy. A similar study was carried out in London (Solomon, 1933). In 1925 Travassos revised the genus Enterobius Leach 1853.
These nematodes have a direct life cycle and lodge in the host�s large intestine and cecum. Symptomology is usually not apparent; one observes only pruritis in the perianal region, caused by the migration of females toward the anus to lay their eggs, which adhere to the perianal region or are eliminated with feces. Otherwise, this parasite is considered to have scant pathogenicity in most mammals.
The purpose of this study was to identify the species of oyxurid infecting Alouatta caraya, ascertain its incidence, and describe its external morphology by means of a light microscope and scanning electron microscopy (SEM).
Materials and Methods
This study was carried out in semideciduous rain forest in the basin of the Riachuelo River, San Cayetano, in Corrientes Province (27� 30� south; 58� 41� west); and at the Faculty of Veterinary Sciences, Universidad Nacional del Nordeste (UNNE), in Corrientes, Argentina. The sample consisted of 51 Alouatta caraya, captured by anesthetic darts containing 0.9 ml ketamine and 0.1 ml of xylazine. The captured animals belonged to different troops and were of both sexes, of various ages ranging from juvenile to adult, and of various weights.
After capture, the anesthetized monkeys were placed in individual, metal squeeze-back cages (80 cm long, 18 cm wide, 32 cm high) and transported to the laboratory in Corrientes. When the monkeys defecated in the cages, their feces were collected and labeled. When they had completely recovered from the anesthesia, they were transported back to the point of capture and released.
For all the monkeys, macro- and microscopic examinations were performed on the fecal matter within 6 hours. The samples were examined microscopically using direct observation and Willis�s flotation method (Basso, et al., 1987). Adult oxyurids that were eliminated spontaneously with feces were removed from the feces, washed with physiological saline, preserved in Raillet & Henry solution (NaCl: 0.8% = 920 ml; Formalin: 40% = 30 ml; glacial acetic acid: 50 ml [Zylber & Ostrowski de Nuñez, 1999]), and examined to ascertain their systematic classification. The specimens to be studied with the scanning electron microscope were fixed in a 3% glutaraldehyde solution, dehydrated, mounted, metalized, and observed and photographed by SEM. In addition, morphometric data and microphotographs were obtained from adult specimens and eggs observed through a light microscope.
Results
Of the 51 monkeys captured, 5 (9.8%) tested positive for nematodes using direct examination of fecal matter and Willis�s technique.
The nematodes extracted from the feces were very fragile and had a pearly white cuticle and pronounced sexual dimorphism. Females showed a tapering shape and a sharp, subulate (tapered) tail; males were smaller and comma-shaped. According to the morphologic and morphometric characteristics we observed (Table 1), the species was identified as Trypanoxyuris (Hapaloxyuris) callithricis (Solomon, 1933; Inglis & Cosgrove, 1975) (henceforth called T. callithricis).
Some of the morphological characteristics distinguishing T. callithricis are: the presence of a cephalic cuticular swelling at the anterior end, and three buccal lips surrounding the oral aperture (Figures 1, 2, and 3); a conspicuous esophageal bulb in both sexes; a simple spicule in the male�s tail; and the subulate form of the female�s tail (Figure 4).
Figure 1: General view of anterior end (ae) and body (b) with some details of the transversally striated cuticle and longitudinal cord (lc) of Tripanoxyuris (Hapaloxyuris) callithricis. (SEM: 190 X)
Figures 2 and 3: Anterior view and profile of anterior end of the body. oa = oral aperture; cex = cephalic cuticular swelling; l = length; w = width; l = lobulated lips. (SEM: 1300 and 900 X)
Figure 4: Female, caudal end. (SEM: 190 X)
Discussion
The nematodes of the family Oxyuridae have undergone many changes in their classification, especially in primates and rodents. The genera parasitizing primates are Enterobius, Trypanoxyuris, and Lemuricola (Inglis & Cosgrove, 1965; Masi Pallares, 1990; Petter & Quentin, 1974; Yamaguti, 1961; Yorke & Maplestone, 1926). In the nematodes we found in Alouatta caraya, the number of caudal papillae, the absence of sclerotized elements reinforcing the pre-anal papillae in the male, the absence of pharyngeal teeth, and the presence of three well developed buccal lips identify them as Trypanoxyuris (Hapaloxyuris) callithricis Solomon, 1933 (Hugot, 1984, 1985; Moriena et al., 1979; Travassos, 1925; Solomon, 1933).
| Characteristics | Male | Female |
| total length | 1887 to 2519 | 5000 to 7000 |
| width | 100 | 82.5 to 105 |
| length of cephalic cuticular swelling | 40.8 | 81.6 |
| width of cephalic cuticular swelling | 122.4 | 93 to 142 |
| teeth | - | - |
| number of buccal lips | 3 | 3 |
| esophageal length | 469.2 | 948.6 |
| esophageal width (last portion) | 40.8 | 81.6 |
| length of esophageal bulb | 91.8 | 122-127 |
| width of esophageal bulb | 71.4 | 122.4 |
| length of spicule | 42 | - |
| length of caudal spine | 10 | - |
| number of caudal papillae | 4 pairs | - |
| length of tail | - | 1315 to 1419 |
| distance between vulva and end of tail | - | 2397 |
| eggs | - | 46.2 to 57.5 x 20 to 22.5 |
Table 1: Morphometric characteristics (in �m) of Trypanoxyuris (Hapaloxyuris) callithricis.
This is the first report of T. callithricis in this host, Alouatta caraya, and in a new geographical distribution, northeastern Argentina. The SEM photographs presented here are the first for T. callithricis.
References
Basso, N., Calceta Resio, E., Dughetti, R. P., Jiménez, R. A., Perez Tort, G. B., Rosa, A. B., & Welch, E. L. (1987). Fundamentos de parasitología veterinaria. Buenos Aires: Editorial Hemisferio Sur.
Cabrera, A. L., & Willink, A. (1973). Biogeografía de América Latina. Serie de Biología, No. 13. Washington, DC: Organization of American States.
Hugot, J. P. (1984). Sur le genre Trypanoxyuris (Oxyuridae, Nematoda) II. Sous genre Hapaloxyuris parasite de primates Callithricidae. Bulletin du Museum (National) d�Histoire Naturelle, 6, 1007-1019.
Hugot, J. P. (1985). Sur le genre Trypanoxyuris (Oxyuridae, Nematoda) III. Sous genre Trypanoxyuris parasite de Cebidae et Atelidae. Bulletin du Museum (National) d�Histoire Naturelle, 7, 131-155.
Inglis, W. G., & Cosgrove, G. E. (1965). The pin-worm parasites (Nematoda: Oxyuridae) of the Hapalidae (Mammalia: Primates). Parasitology, 55, 731-737.
Masi Pallares, R. (1990). Nematodes de los vertebrados del Paraguay y otros parásitos del Nuevo Mundo. Asunción, Paraguay: EFACIM.
Moriena, R. A., Lombardero, O. J., & Coppo, J. A. (1979). Nuevos parásitos de primates para la Argentina. Acta Zoológica Lilloana, 35, 13-19.
Petter, A. J., & Quentin, C. (1976). Keys to genera of the Oxyuroidae. CIH keys to the Nematode parasites of vertebrates, No. 4. Farnham Royal, England: Commonwealth Agricultural Bureau.
Pope, B. L. (1966). Some parasites of the howler monkeys of Northern Argentina. Journal of Parasitology, 52, 166-168.
Solomon, G. S. (1933). On a new species of Enterobius from the marmoset (Callithrix jacchus). Journal of Helminthology, 11, 95-100.
Travassos, L. (1925). Fauna Brasiliense. Nematodes: Oxyuroidea-Oxyuridae. Revis�o do genero Enterobius Leach, 1853. Museu Nacional do Rio de Janeiro, 2, 5-13.
Yamaguti, S. (1961). Systema Helminthum 3. The nematodes of vertebrates. Parts I and II. New York: Interscience Publishers, Inc.
Yorke, W., & Maplestone, P. A. (1926). The nematode parasites of vertebrates. London: J. & A. Churchill.
Zylber, M. I., & Ostrowski de Nuñez, M. (1999). Some aspects of the development of Lobatostoma jungwirthi Kritscher, 1974 (Aspidogastrea) in snails and cichlid fishes from Buenos Aires, Argentina. Memórias do Instituto Oswaldo Cruz, 94[1], 31-35.
----------
Corresponding author: Dra. Antonia M. Santa Cruz, Manzana C, Casa 21, Barrio CODEPRO, Corrientes, C.P. 3400, Argentina [54 3783 444260; fax: 54 3783 425783; e-mail: [email protected]]. This article was translated by Elva Mathiesen and Juan T. Borda. We thank Dr. Morris Povar for his helpful comments on the manuscript.
----------
* * *
Announcements from Publications
Developments In Primatology: Progress and Prospects
Kluwer Academic/Plenum Publishers is pleased to introduce Developments in Primatology: Progress and Prospects (formerly Advances in Primatology), a new series under the direction of the Editor-in-Chief of the International Journal of Primatology, Dr. Russell H. Tuttle of the University of Chicago.
This peer-reviewed book series will meld the facts of organic diversity with the continuity of the evolutionary process. The volumes in this series will exemplify the diversity of theoretical perspectives and methodological approaches currently employed by primatologists and physical anthropologists. Specific coverage includes: primate behavior in natural habitats and captive settings; primate ecology and conservation; functional morphology and developmental biology of primates; primate systematics; genetic and phenotypic differences among living primates; and paleoprimatology.
Volume authors will be invited to participate based on their expertise in a given area and overall approval by the series editor. For further information on how to contribute an authored or edited volume to this series, contact Andrea Macaluso [212-620-8007; e-mail: [email protected]]. - From Neotropical Primates, 2001, 9[3]
Handbook of Animal Behavior
Marc Bekoff, of the University of Colorado, Boulder, has been asked by a publisher to consider editing a Handbook of Animal Behavior for middle- and high-school students. He knows this will be a very challenging task and asks how many scientists would consider writing a short piece for this three-volume set. �We envision that there will be 300 to 400 essays ranging in length from about 500 to 5000 words, depending on the topic. There likely will be no or very little remuneration for your efforts and there typically are no royalties for these sorts of volumes, but this is a great way to get word out about our wonderful field to students who really want to know more about animal behavior. In the end, a board of advisors will decide who will write on which topic, but I need to begin investigating whether there is enough interest at the current time. Essays will be due around December, 2003, so there is plenty of time to write them. But please respond as soon as possible, to Marc Bekoff, Dept of Biology, University of Colorado, Boulder, CO 80309 [e-mail: [email protected]].�
* * *
How Important Are Urinary Signals in Alouatta?
Clara B. Jones
Livingstone College, Community Conservation, Inc.
Introduction
Chemical communication, including the detection of pheromones, has proven to be a difficult undertaking in mammals, in part because analysis is complicated by the complex chemical structure of substances secreted by these animals, and in part because the causes and consequences of mammalian behavior are likely to be multifactorial, increasing the challenge of developing confident assays for specific chemical signals (Johnston, 2000). A further complication for the study of chemical communication in mammals, including primates, is the degree to which the olfactory system is specialized for the encoding and discrimination of chemicals (Johnston, 2000; Smith et al., 2001). It has generally been assumed that prosimians and New World primates are characterized by a vomeronasal system while Old World monkeys and apes are not (Johnston, 2000). Recent work, however, has reevaluated the vomeronasal system in Old World monkeys and apes, emphasizing the variability of the vomeronasal system in these animals and the need for further investigation (Smith et al., 2001). The objectives of the present technical comment are threefold: (1) to review briefly what is known of chemical communication in Alouatta, (2) to present the results of a chemical analysis of a single sample of urine for A. caraya (the black-and-gold howler monkey), and (3) to encourage further empirical work in both the laboratory and field on chemical communication in howler monkeys.
Objective 1
While I have been unable to locate publications demonstrating chemical communication in Alouatta, numerous behavioral studies suggest that chemical communication, especially urinary signals, may be important in this taxon. Although Milton (1985) claims that �an impressive number of New World primates...use urine in chemical communication� (p. 154), few primate studies provide experimental evidence for the role of urinary substances in intraspecific signaling (but see, for example, Laska & Hidson, 1995). Indirect and/or descriptive evidence that howler monkeys may employ urinary compounds in intraspecific communication derives primarily from two sources.
First, there is general agreement among students of Alouatta that their arboreal travel routes are often predictable, �linear�, and repetitious (A. palliata [the mantled howler monkey]: e.g., Carpenter, 1934; author�s personal observation (p.o.); A. pigra [the black howler monkey]: R. Brockett, personal communication (p.c.), May 2002; p.o.; A. caraya: G. E. Zunino, p.c., March 2002; J. C. Bicca-Marques, p.c., May 2002; A. seniculus [the red howler monkey]: C. Crockett, p.c., April 2002), raising the question of how this regularity is accomplished. Discussing �urine-rubbing behavior� (�urine-washing�) in mantled howlers, Milton (1975) suggests that group members may leave a �scent trail� through trees as a result of residues of urine deposited on hands, feet, and tails, facilitating efficient travel and group cohesion. Milton�s (1975) discussion of how a �scent trail� may function provides a possible explanation for the pattern of travel observed in Alouatta. Urine-washing, however, has not been reported in species of Alouatta other than A. palliata, possibly because other howlers are less studied or possibly because of differences in behavior and social structure. Mantled howlers are generally organized in multimale-multifemale groups whereas other howlers are generally polygynous.
A second body of evidence also indicates that urinary cues may be important in howlers. Males may actively investigate the urine of females by smell or taste, possibly to assess their reproductive condition (A. palliata: Glander, 1975, 1980, p.o.; A. pigra: K. Eckert, p.c., March 2002; see Horwich, 1983; A. seniculus: G. Agoramoorthy, p.c., April 2002). Responses by adult males to the urine of reproductive females have rarely been reported from this genus with the exception of A. palliata. If male inspection of female urine is rare in polygynous howlers, it is possible that there has been weaker selection on females in those species for signals to attract males by advertising reproductive condition and/or receptivity and/or to incite male-male competition.
Objective 2
A large body of literature exists on the behavioral responses to male urine by female rodents. Gosling & Roberts (2001), for example, propose that urinary and other scent marks by male mammals represent conditional �signals of quality�. In a preliminary attempt to assess the potential of urinary compounds to be employed as chemical signals by male howlers, a sample of urine was collected noninvasively from an adult male A. caraya at Riverbanks Zoological Park. Approximately 10 milliliters of urine were collected, frozen, and shipped in dry ice to S. A. Barker�s laboratory at Louisiana State University for analysis. The information in the following paragraph was sent to the author by Barker in March, 2002.
A gas chromatography-mass spectrometry (GC-MS) screening was conducted �of a base-neutral extract of the urine� and �the compounds were tentatively identified by mass spectral matching to a [National Institute of Standards and Technology] library data base�. Such matching does not permit positive identification of compounds nor does it provide a reliable assessment of the compounds comprising the urine of male A. caraya. �One of the major and most volatile components is quinolinone. A second major component appears to be an isomer of aminophenyl butanone (probably 1-[4-aminophenyl]-1-butanone]. A third component (minor) is hexahydro-3-[2-methylpropyl]-pyrrolo[1, 2-a]pyrazine-1, 4-dione. There also appears to be an isomer of this compound present. A related compound, 5, 10-diethoxy-2, 3, 7, 8-tetrahydro-1H, 6H-dipyrrolo[1, 2-a; 1�, 2�-d] pyrazine is a major component (but less than quinolinone). There are several fatty acid ester compounds present (low level). One steroid seems to be present. Currently, it is identified as � Adrenosterone or possibly a Deoxypregnenolone� There are a number of other trace compounds that are and will probably remain unidentified (insufficient data). The urine was not treated with sulfatase or glucuronidase to liberate conjugated amines, etc.� Neither head-space an-alysis nor acid extraction demonstrated significant peaks.
My primary justification for presenting these preliminary results is that all studies of chemical communication in animals begin with the results of a GC-MS screening (R. E. Johnston, p.c., March 2002). Since only one male was sampled, these results are also presented to demonstrate the potential for reliable analysis since the base-urine extraction revealed �several peaks that were identifiable�. However, R. E. Johnston (p.c., March 2002), a specialist, fails to recognize any of the tentatively identified compounds as pheromones.
Since chemical communication in mammals most likely involves mixtures of several compounds (�mosaic signals�: Johnston, 2000), it would be premature to speculate upon the possible role of the compounds tentatively identified in the urine of one male A. caraya. Such confident identification �would require much more effort and the availability of reference standards for each to obtain absolute confirmation� (S. A. Barker, p.c., February 2002). The present results, however, may indicate the potential for howlers, at least howler males, to employ chemical communication by urinary signals and may provide a rationale for future investigations.
Objective 3
It is hoped that the analysis of A. caraya urine combined with the observations from descriptive studies and anecdotal observations presented here may stimulate further research on the use of urinary compounds as chemical signals in Alouatta. Investigations of the vomeronasal system of howler monkeys, the development of assays to permit experimental tests of components of their urinary compounds, and the application of new research techniques (e.g., Toftegaard et al., 2002) in both the laboratory and field have the potential to advance knowledge of chemical communication in primates. In particular, methodological and technical advances by primatologists studying chemical communication in Alouatta may resolve important questions pertinent to group coordination and control, intrasexual competition, mate choice, the timing and frequency of abortions, estrous synchrony, and dominance rank, and may confirm the existence of pheromones in primates (see Epple, 1974).
References
Calegaro-Marques, C. (1992). Comportamento social de um grupo de Alouatta caraya (Primates, Cebidae) em Alegrete, RS, Brasil. Master�s Thesis. Universidade de Brasilia.
Carpenter, C. R. (1934). A field study of the behavior and social relations of howling monkeys. Comparative Psychology Monographs, 10, 1-167.
Epple, G. (1974). Pheromones in primate reproduction and social behavior. Advances in Behavioral Biology, 11, 131-155.
Glander, K. E. (1975). Habitat and resource utilization: An ecological view of social organization in mantled howling monkeys. Ph.D. Dissertation, University of Chicago.
Glander, K. E. (1980). Reproduction and population growth in free-ranging mantled howling monkeys. American Journal of Physical Anthropology, 53, 25-36.
Gosling, L. M. & Roberts, S. C. (2001). Scent-marking by male mammals: Cheat-proof signals to competitors and mates. Advances in the Study of Behavior, 30, 169-217.
Horwich, R. H. (1983). Breeding behaviors in the black howler monkeys (Alouatta pigra) of Belize. Primates, 24, 222-230.
Horwich, R. H., Brockett, R. C., James, R. A., & Jones, C. B. (2001). Population growth in the Belizean black howling monkey (Alouatta pigra). Neotropical Primates, 9, 1-7.
Johnston, R. E. (2000). Chemical communication and pheromones: The types of chemical signals and the role of the vomeronasal system. In T. E. Finger, W. L. Silver, & D. Restrepo (Eds). The neurobiology of taste and smell (2nd Ed.) (pp. 101-127). New York: Wiley-Liss.
Laska, M. & Hidson, R. (1995). Ability of female squirrel monkeys to discriminate between conspecific urine odours. Ethology, 99, 39-52.
Milton, K. (1975). Urine-rubbing behavior in the mantled howler monkey Alouatta palliata. Folia Primatologica, 23, 105-112.
Milton, K. (1985). Urine washing behavior in the woolly spider monkey (Brachyteles arachnoides). Zeitschrift fur Tierpsychologie, 67, 154-160.
Smith, T. D., Siegel, M. I., & Bhatnagar, K. P. (2001). Reappraisal of the vomeronasal system of catarrhine primates: Ontogeny, morphology, functionality, and persisting questions. Anatomical Record, 265, 176-192.
Toftegaard, C. L., McMahon, K. L., Galloway, G. J., & Bradley, A. J. (2002). Processing of urinary pheromones in Antechinus stuartii (Marsupialia: Dasyuridae): Functional magnetic resonance imaging of the brain. Journal of Mammalogy, 83, 71-80.
----------
Author�s address: Livingstone College, School of Liberal Arts, Dept of Psychology and Sociology, 701 W. Monroe St., Salisbury, NC 28144 [e-mail: [email protected]].
This research was supported by Riverbanks Zoological Park (Columbia, South Carolina). I thank Collection Manager A. Shoemaker and the staff at Riverbanks for approving the proposal, collecting the urine, and preparing it for shipping. Dr. S. A. Barker is gratefully acknowledged for conducting the urinalysis at the expense of his laboratory at Louisiana State University. Dr. R. E. Johnston provided rapid instruction on chemical communication in mammals, key references, and interpretations of the urinalysis. Dr. B. Fontenot generously assisted with logistics. I appreciate numerous communications received by personal request and in response to a query on Primate-Science [[email protected]]. Dr. G. Agoramoorthy, Dr. J. C. Bicca-Marques, Dr. C. M. Crockett, Dr. K. E. Glander, Dr. R. Horwich, Dr. A. Rylands, N. Shahnoor and, especially, Dr. L. Jacobsen kindly provided bibliographic references. Comments by Dr. J. C. Bicca-Marques, A. Shoemaker, Dr. R. Horwich, Dr. G. Agoramoorthy, Dr. G. Brooks, Dr. K. E. Glander, and the Editors of the Laboratory Primate Newsletter on prior versions of this technical comment significantly improved the manuscript. This communication is dedicated to Dr. G. Brooks for her personal and professional interest and advice.
----------
* * *
Meeting Announcements
The 19th Annual Conference of the European Association of Zoos and Aquaria (EAZA) will be held September 17-22, 2002, hosted by the Barcelona Zoo. The European Taxon Advisory Groups (TAGs) will meet on September 17, 2002. The main theme of the Conference will be Central and South America, with emphasis on EAZA�s current fund-raising and awareness campaign - the Atlantic forest of Brazil, Argentina, and Paraguay. See <www.eaza.net/index.html>. The deadline for early registration for the Conference is June 30, 2002. The registration and hotel booking forms can be downloaded from the Website or requested from the EAZA Executive Office: [e-mail: [email protected]]. As Barcelona is a popular tourist destination, hotel booking before 30 June 2002 is necessary. The meetings will be at Pompeu Fabra University, across the street from the Barcelona Zoo.
A conference, Infectious Diseases: Challenges, Threats and Responsibilities, will be held November 8-9, 2002, at the European Molecular Biology Laboratory (EMBL) in Heidelberg, Germany. This is the third annual Science and Society conference to be co-organized by EMBL and the European Molecular Biology Organization. As before, this year�s meeting will be interdisciplinary, its principal aim being to bring together life scientists, social scientists, decision-makers, and members of the public to discuss how best to generate and make available the scientific and economic means to protect humanity from infectious agents. For detailed information about the conference program, speakers and panelists, on-line registration, etc., please consult <www.embl-heidelberg.de/Conferences/SciSoc02/index.html>.
The 4th European Congress of Mammalogy will be held July 27 to August 1, 2003, in Brno, Czech Republic, hosted by the Institute of Vertebrate Biology, Academy of Sciences of the Czech Republic. Information and the pre-registration form are available at <www.ivb.cz>. Any questions should be directed to Jan Zima, Organizing Committee [e-mail: [email protected]].
* * *
Volunteer Opportunity: International Center for Gibbon Studies
The International Center for Gibbon Studies, directed by Alan Mootnick, is a nonprofit organization that has worked since 1977 to prevent the extinction of endangered gibbons and to educate the public, zoos, and scientists about them. �We are seeking volunteers to serve as primate keepers, who will prepare and distribute food to captive gibbons in 16 outdoor enclosures, clean enclosures, change water, clean the kitchen and bathrooms, and who might also do behavioral observation, medicate gibbons, enter data into a Macintosh computer, and do filing, fundraising, grounds maintenance (weeding, raking), and library research. �Volunteers must love animals, be at least 20 years old, be in good physical condition, be able to get along with a wide variety of people, and be able to work both independently and with others. Medical tests and vaccinations are required. Free lodging in older, austere travel trailers - with access to kitchen, bathroom, laundry facilities, Internet, and phone - is provided.
�While volunteers are needed all year, we generally have a shortage from September to April. We ask volunteers to commit to a minimum one-month stay. For more information, contact Patricia Dahle, Volunteer Coordinator, ICGS, P.O. Box 800249, Santa Clarita, CA 91380 [661-943-4915 (9 am-8 pm Pacific time); fax: 661-296-1237; e-mail: [email protected]].�
* * *
Multiple Drug Resistance in Rhesus Macaques of Chinese Origin
In May, Iris Bolton, DVM, of the Wisconsin Primate Research Center, posted the following notice to the Primate-Science and CompMed mailing lists. We thought it was worth printing here for more general distribution. Please send any comments to us, and to Dr. Bolton, Wisconsin PRC, 1220 Capitol Ct, Madison, WI 53715 [608-263-3571; e-mail: [email protected]].
The Wisconsin Primate Research Center recently received a shipment of Chinese origin rhesus macaques. Shortly after the animals� arrival, approximately 16% developed bloody diarrhea with subsequent loss of one animal within 72 hours. Rectal cultures from affected animals revealed Shigella flexneri, which was expected, and the following sensitivity pattern reflecting multiple drug resistance, which was not expected:
| cefotaxime | sensitive |
| cefazolin | sensitive |
| ceftriaxone | sensitive |
| naxcel | sensitive |
| enrofloxacin | resistant |
| tetracycline | resistant |
| chloramphenicol nos | resistant |
| gentamicin,gentamycin | resistant |
| ciprofloxacin | resistant |
| sulfamethoxazole/trimethoprim | resistant |
| cephalothin | resistant |
| amoxicillin/clavulanic acid | resistant |
We have since learned that antibacterial use in some Chinese facilities is less than optimal by common U.S. practices and that these animals had likely been treated with �whatever antibiotic they could get their hands on,� frequently in their water at varying doses and durations. This information prompted us to isolate additional bacteria from these animals to assess the extent of the drug resistance. From a representative subpopulation we isolated several cultures of Escherichia coli that consistently demonstrated the following sensitivity pattern:
| amoxicillin/clavulanic acid | resistant |
| enrofloxacin | resistant |
| tetracycline | resistant |
| chloramphenicol nos | resistant |
| naxcel | resistant |
| gentamicin,gentamycin | resistant |
| ciprofloxacin | resistant |
| sulfamethoxazole/trimethoprim | resistant |
| cefazolin | resistant |
| cephalothin | resistant |
| ceftriaxone | resistant |
| cefotaxime | resistant |
These results have warranted further questions with serious implications: What are the potential risks to our existing colony? What are the potential risks to our personnel? Do we want to immunosuppress these animals? Do we want to breed these animals?
Of the 7,708 rhesus monkeys imported to the United States from 1995-2001, many have come from China. I am curious whether any readers have had problems with multiple drug resistance in Chinese rhesus monkeys. If you haven�t, have you utilized these animals differently than other rhesus? If none of you have experienced problems, does that negate the risk of exposing our personnel and animals to these organisms and others they may be carrying? If none of you have experienced any problems, does that excuse us from taking this information seriously? I think not, but I would like to appeal to the community at large for information, suggestions, recommendations, and experiences.
Thank you for your help.
* * *
Certified Manager of Animal Resources Examination
The Certified Manager of Animal Resources (CMAR) examination will be administered as a paper-and-pencil exam at the AALAS National Meeting (October 27-31, 2002), and the AALAS Management & Technology Meeting (February 6-8, 2003). To apply, qualified candidates must submit a CMAR application form and fees to AALAS before the specified deadline for each meeting. There is a $25 application fee plus an $85 exam fee for a total of $110.
To learn about eligibility requirements and to apply for the exam, contact the AALAS office or download materials from <www.aalas.org>. Click on �Certification� and follow the link to �Manager Certification�. For any other questions, contact Marti White [e-mail: [email protected]] or Cristi Mann [[email protected]], or phone 901-754-8620.
* * *
Information Requested or Available
OLDMEDLINE
The National Library of Medicine�s OLDMEDLINE <www.nlm.nih.gov/databases/databases_oldmedline.html> contains citations published in the 1960-1965 Cumulated Index Medicus and the 1958-1959 Current List of Medical Literature. OLDMEDLINE covers the fields of medicine, preclinical sciences, and allied health sciences.
PrimateLit Update
PrimateLit <primatelit.library.wisc.edu/> is the key bibliographic database for searching the nonhuman primate research literature. Approximately 200,000 citations are indexed. Coverage is from 1940 to date. The National Center for Research Resources supports open access to this database for everyone in the national and international scientific communities. No fees or passwords are required. Everyone with an interest in primates is encouraged to use this resource.
Features include: * Allows subject, author and key word searches * Can limit results by date, species and country * Search results can be saved, e-mailed or printed. Recently links have been added from citations in PrimateLit to corresponding abstracts in the National Library of Medicine�s PUBMED database. A project is underway to match all candidate records to their equivalent PUBMED abstracts. Links to full text of many electronically available publications (including the LPN) are being established.
Comments and suggestions on how PrimateLit can be modified and improved are welcome. Contact Larry Jacobsen, Director, Wisconsin Primate Research Center Library and Information Service, 1223 Capitol Ct, Madison, WI 53715 [e-mail: [email protected]].
PrimateLit is supported by NCRR Grant RR15311: Coordinated Information Services to Support Primate Research. The database is indexed by the Primate Information Center at the Washington PRC in Seattle and managed by the Wisconsin PRC and the Libraries at the University of Wisconsin-Madison.
Novel Cage Card Holder
Scott R. Klipstein of the Institute for Human Gene Therapy, University of Pennsylvania, together with Jeffrey A. House and Stuart E. Leland, has created and is patenting a �Novel Nonhuman Primate Cage Card Holder�. He writes, �Maintaining intact cage cards on nonhuman primate cages was a challenge at our facility. The integrated modular cage-card holders on commercially available nonhuman primate cages were inadequate for the purpose of retaining cage cards for any significant amount of time. Applying tape to the cage or the card itself did not significantly improve retention time.�
At <mail.med.upenn.edu/~scott2/cagecard.html>, he gives clear instructions for creating a polycarbonate plastic and stainless steel gadget that is water-resistant, chemical-resistant, and autoclavable. He claims that the devices cost about $5.00 each in materials to produce, pay for themselves in reduced labor and increased compliance, are highly visible and indestructible, and serve as enrichment manipulanda. For more information, contact Scott [e-mail: [email protected]].
Genomics and World Health
The World Health Organization�s recently published report, Genomics and World Health, is now available in pdf format at <www.who.int/genomics>. The report is intended to highlight the relevance of genomics for health care world-wide, with a particular focus on its potential for improving health in developing countries. The report notes that genomics is a double-edged sword, as it can lead to new interventions that can improve the health of all but can also widen existing inequalities and the health and technology gaps between rich and poor countries. Chinese, Arabic, Russian, Spanish and French editions, the other official languages of WHO, will be available later in the year.
Field Studies Guide
The Primate Society of Great Britain and the Wisconsin Primate Research Center are compiling a database of field studies. If you are carrying out field studies of any sort, see <www.primate.wisc.edu/pin/idp/scope.html> for a form (or write to Eluned Price, 2 La Grange, La Rue de Cambrai, Trinity, Jersey JE3 5AL, Channel Islands, G.B. [e-mail: [email protected]] for a copy), fill it out, and send it to Eluned.
More Interesting Web Sites
* BioAcoustics Team, animal acoustic communication: <www.cb.u-psud.fr/cb/index.html>
* Bushmeat Exhibition: <www.bushmeat-campaign.net>
* Conservation database for lorises and pottos: <www.ruhr-uni-bochum.de/neuroanatomie/people/helga_schulze/loris/index.html>
* The Extinction Crisis Face to Face (IUCN Red Book): <www.iucn.org/redlist/redbook/>
* International Society for Comparative Psychology: <www.er.uqam.ca/nobel/iscp/>
* IUCN Species Survival Commission: <www.iucn.org/>
* Veterinary Emergency and Critical Care Society: <www.veccs.org/>
* Life Spans of Nonhuman Primates: <www.primate.wisc.edu/pin/lifespan.html>
* Loris Homepage: <www.nocturnalprimate.org>
* National Association for Biomedical Research: <www.nabr.org>
* National Library of Medicine�s Databases and Electronic Information Sources: <www.nlm.nih.gov/databases/databases.html>
* Macaca nemestrina Brain Atlas: <www.loni.ucla.edu/Research_Loni/atlases/nemestrina.html>
* Species Information Network: <www.species.net>
* State of the World�s Forests, 2001: <www.fao.org/docrep/003/y0900e/y0900e00.htm>
* * *
Research and Educational Opportunities
Postdoctoral Training, Laboratory Animal Medicine
The Scripps Research Institute�s Department of Animal Resources (DAR) invites applications for a post-doctoral veterinary training position in laboratory animal medicine. Review of applications is ongoing and will continue until the position is filled. The three-year program is designed to support preparation toward ACLAM board certification, to provide a good foundation for ability to manage a program of laboratory animal care and research support, and to develop and/or increase research aptitude. Areas of training include laboratory animal clinical medicine, comparative pathology, methods and practice of biomedical research, and animal resource and facilities management. The position furnishes opportunities to work with a wide variety of animal species in an AAALAC-accredited, respected animal care and use program. It also includes opportunities to work with established research scientists in a sophisticated research environment to attain the necessary skills to plan and conduct research and to contribute to the scientific literature.
Candidates for this position should have a DVM/ VMD or equivalent degree. The starting salary is $28,416 plus benefits, increasing to $33,516 in the second year.
Interested applicants should forward a CV, statement of goals and interests, and three letters of recommendation to the DAR Office c/o Director of Comparative Medicine, Dept of Animal Resources, MB-18, Scripps Research Inst., 10550 N. Torrey Pines Rd, La Jolla, CA 92037. The Scripps Research Institute is an AA/EOE.
New Opportunities - NSF
The National Science Foundation�s (NSF) Undergraduate Mentoring in Environmental Biology (UMEB) program, which seeks to provide research opportunities for students from groups underrepresented in science, now offers three types of grants. In addition to Research-Mentoring grants of up to $500,000 total for five years, the expanded UMEB Program can now support Planning Activities aimed at strengthening partnerships between institutions that plan to submit a Research-Mentoring proposal. Travel grants can also be made that enable professional societies to enrich their meetings with undergraduate students from underrepresented groups.
The expanded UMEB Program is described, along with descriptions of current awards, at <www.nsf.gov/bio/progdes/umeb.htm>. If you have questions after reading the program solicitation, you may write to the general UMEB e-mail address <[email protected]> or to any of the following program officers: � Elizabeth E. Lyons, Division of Environmental Biology [703-292-7188; e-mail: [email protected]]; � Fred Stollnitz, Division of Integrative Biology and Neuroscience [703-292-7868; e-mail: [email protected]]; � Ronald M. Weiner, Division of Molecular and Cellular Biosciences [703-292-7144; e-mail: [email protected]]; � Sally E. O�Connor, Division of Biological Infrastructure [703-292-4552; e-mail: [email protected]]; all are at NSF, 4201 Wilson Blvd, Arlington, VA 22230.
Postdoc - Duke University
A postdoctoral position is available immediately in the laboratory of Dr. Roger Madison at Duke University and the Durham Veterans Affairs Medical Center. The salary will be $24,000-$30,000, commensurate with experience. They are seeking a self-motivated and independent worker to supervise behavioral and physiological experiments using an animal model to study recovery of normal hand function following experimental median nerve repair. Duties will include animal handling, observation of behavioral sessions, physiological recording, data collection and analysis, manuscript preparation, and direct supervision of a dedicated behavioral technician. Previous work with large animals, whole animal electromyography or related physiology work, and any computer programming experience with LabView are considered assets. Send a recent CV and contact information for three references to: Roger Madison, Research Career Scientist, Veterans Affairs Medical Center, Bldg 16, Rm 38, 508 Fulton St, Durham, NC 27705 [e-mail: [email protected]].
Postdoctoral Fellowship - Oregon NPRC
An NIH-funded postdoctoral position is available at the Oregon Health and Science University (Oregon National Primate Research Center) to study postnatal and pubertal development of neuroendocrine circuits in the brain, and to examine the influence of sex steroids and photoperiod on brain plasticity. The research will involve the use of nonhuman primates and employ a wide range of techniques, including laser-capture microdissection, gene microarray screening, confocal microscopy, and in situ hybridization histochemistry. Experience in reproductive neuroendocrinology, molecular biology, chronobiology, hormone assay, and immunohistochemistry would be considered an advantage. Candidate must be a U.S. citizen or hold a visa for permanent residence. Send a CV, a copy of graduate school transcripts, two letters of recommendation, and a one-page research plan to: Dr. Henryk Urbanski, Div. of Neuroscience, ONPRC/OHSU, 505 N.W. 185th Ave, Beaverton, OR 97006 [503-690-5306; fax: 503-690-5384; e-mail: [email protected]].
Captive Tarsiers - Indonesia
The Indonesian Institute of Sciences, Center for Zoological Research (better known as the Museum Zoologicum Bogoriense, or MZB), and the Center for Biodiversity and Conservation Studies, University of Indonesia (CBCS-UI), are looking for a volunteer/trainee to work at the MZB with captive tarsiers. In line with a larger goal to protect the biodiversity of Sulawesi, MZB and CBCS-UI have a small number of captive Eastern tarsiers, captured and housed with permission from the Indonesian Department of Forestry�s Directorate for the Preservation of Forests and Biodiversity. The colony includes a mated pair from Pattanuang, South Sulawesi (Tarsius tarsier Erxleben 1777), an adult and subadult male from Selayar Island, South Sulawesi (Tarsius sp? Groves 1998), and a subadult male from Gimpu, Central Sulawesi (the �Palu Acoustic form� of MacKinnon and MacKinnon). The colony is being used for a variety of research projects such as captive behavior, parasitology, taxonomy, acoustic behavior, etc. The trainee will be responsible for daily care of the tarsiers as well as assisting the scientists working on the project.
Dedicated, hard-working applicants with a keen interest in primate research and conservation should be prepared to live in a foreign country, and must request a letter reference from an advisor or sponsor. Knowledge of the Bahasa Indonesian language is not necessary.
Trainees must provide their own support. Meals and lodging (with air-conditioning and private bath) cost about $12 per day. Transportation to and from the airport is about $20 each way.
This is a good opportunity for anyone interested in pursuing research on tarsiers, or primatological research of any kind in Indonesia. The trainee will live and work at the compound. The guesthouse, the tarsier enclosure, and the MZB are all within a short walk of each other. The trainee will have ample opportunity to tour the MZB and all its facilities, as well as to interact with the MZB staff. Interested persons are strongly encouraged to apply via e-mail. Contact Myron Shekelle, Center for Biodiversity Studies and Conservation, Faculty of Mathematics and Science, University of Indonesia, Depok, West Java, Indonesia [62-21-786-3431; fax: 62-21-786-3431; e-mail: [email protected]].
* * *
Resources Wanted and Available
Enrichment Online
The Fort Worth Zoo, in conjunction with the American Institute of Biological Sciences, has created an interactive Web-based enrichment resource at <www.enrichmentonline.org>, to provide professional animal managers, both in zoos and laboratories, a comprehensive resource for incorporating enrichment into the care and husbandry of captive animals. A search engine accommodates detailed searches for enrichment ideas. The database allows users to input their own ideas and to provide comments on items already there. Supplemental materials on the site include a list of hot links to other enrichment-related Websites, a list of periodicals and published materials, and a detailed help section.
The Editors, Tarren Wagener and Meg Bommarito, encourage you to assist them in their efforts to make this the best Web-based enrichment resource available. They look forward to hearing from you. Contact information: Tarren K. Wagener, Director of Conservation and Science [817/759-7175; e-mail: [email protected]]; and Meg Bommarito, Conservation Biologist [817-759-7178; e-mail: [email protected]]. For both: Fort Worth Zoo, 1989 Colonial Pkwy, Fort Worth, TX 76110 [fax: 817/759-7501; e-mail: [email protected]].
ARENA/OLAW IACUC Guidebook
The Office of Laboratory Animal Welfare (OLAW) and the Applied Research Ethics National Association (ARENA) announce the second edition of the ARENA/ OLAW Institutional Animal Care and Use Committee (IACUC) Guidebook. The Guidebook is not a regulatory document and neither establishes nor reflects a change in Public Health Service (PHS) Policy on Humane Care and Use of Laboratory Animals. Following the recommendations of the Guidebook will facilitate implementation of institutional animal care and use programs, but is not required. PHS Policy does require compliance with the Institute for Laboratory Animal Research Guide for the Care and Use of Laboratory Animals and Animal Welfare Act regulations.
The Guidebook is available at <grants.nih.gov/grants/olaw/GuideBook.pdf>. Printed copies will soon be mailed to the IACUC chairperson of each institution holding a PHS Animal Welfare Assurance. For additional copies send e-mail to: <[email protected]>. For questions or further information, contact Margaret C. Quinlan, Animal Welfare Program Specialist, OLAW, NIH, 6705 Rockledge Dr., Suite 1050, Bethesda, MD 20892-7982 [301/402-4325; fax: 301-402-2803; e-mail: [email protected]].
Leaf Monkey Loud Calls
Thomas Geissmann (Inst. of Zoology, Tieraerztliche Hochschule Hannover, B�nteweg 17, D - 30559 Hannover, Germany [+49-511-953 84 31; fax: +49-511 953 85 86; e-mail: [email protected]]) has been studying Asian leaf monkey vocalizations. He is trying to collect male loud calls from as many different leaf monkey populations as possible in order to reconstruct leaf monkey phylogeny (as he has been doing for many years for gibbons). He has carried out a preliminary analysis of some leaf monkey calls from Sumatra and the Malayan peninsula, and the results looked very promising. Now he is seeking material from some key taxa, such as any Presbytis from eastern Sumatra south of Medan, P. hosei (Sabah), P. chrysomelas (Sarawak) and Simias concolor (Mentawai islands). Please contact him if you have any material. He will gladly acknowledge your contribution in any publication that uses your material.
* * *
Positions Available
Research Associates - Washington State
A number of positions are available as Research Associates/Specialists in Scientific Services at SNBL USA, a rapidly growing bio-medical research organization located in Everett, Washington (see <www.snblusa.com>). Due to rapid growth, they are seeking qualified candidates for the position of Research Associate. Following standard operating procedures, successful candidates will assist with and independently perform technical, preclinical procedures on study animals, including dosing, drawing blood, collecting urine samples, placing catheters, preparing for surgery, administering anesthesia, and collecting tissue. Other responsibilities include monitoring and reporting animal behavior, setting up equipment, restraining animals, performing other laboratory protocols, and assisting with or performing surgeries. May perform necropsies, as needed. May supervise the work of others. Higher-level positions will act as Study Supervisor in a Good Laboratory Practices facility. These positions require a high school diploma (or equivalent) and one to three years of veterinary clinic and/or farm animal experience, OR a bachelor�s degree in a scientific discipline with related laboratory training/experience. Equivalent education and experience may substitute for stated requirements. SNBL offers a competitive salary and benefit package along with a dynamic, team-oriented work environment. Please send a resume to: SNBL USA, Ltd, 6605 Merrill Creek Pkwy, Everett, WA 98203, Attn: Human Resources [fax: 425-407-8601; e-mail: [email protected]]. EOE.
Laboratory Technicians - North Carolina
Full-time laboratory technicians are needed to work on a team investigating early environmental influences on physiological and behavioral development in rhesus monkeys, as part of ongoing pharmacological and behavioral genetic studies. Duties include daily behavioral observations, cognitive testing, physiological measurements, data entry, training animals for various behavioral tasks, maintaining animal protocol forms, ordering supplies, and other duties necessary for functioning of the laboratory.
The ideal applicant will have previous research experience or coursework, as well as strong communication skills and willingness to participate in multiple research projects as part of a collaborative team. A bachelor�s degree in psychology, physiology, biology, biochemistry, or other related field with a minimum of one year of laboratory experience is preferred. Strong organizational abilities, flexibility, and enthusiasm for working closely with rhesus monkeys are necessary. Research experience with nonhuman primates or other animals is an asset, but not required. The suitable candidate should be able to participate in the day-to-day operations of studies, analyze research data using appropriate computer software applications, use internet bioinformatic resources, and contribute to the preparation of technical procedures and reports.
Contact Allyson J. Bennett, Depts of Physiology/Pharmacology and Pediatrics, Wake Forest Univ. School of Med., Winston-Salem, NC 27157 [336-716-1529; fax: 336-716-1515; e-mail: [email protected]].
Assistant Supervisor - Maryland
Priority One Services, a government contracting company based in Alexandria, Virginia, with multiple sites on the east coast, is currently seeking candidates for the position of Animal Facility Assistant Supervisor in Poolesville, Maryland. Candidates must be certified at the LAT level, with at least four years� experience in an animal facility and experience with nonhuman primates. EOE; Drug Free Workplace. Mail or fax cover letters and resumes to Priority One Services, Inc., Attn: Recruiter, 6600 Fleet Dr., Alexandria, VA 22310 [fax: 703-971-0117; e-mail: [email protected]].
Clinical Veterinarian - Tulane NPRC
The Tulane National Primate Research Center (TNPRC) is seeking applications for the position of clinical veterinarian within the Department of Veterinary Medicine. The TNPRC is an AAALAC International-accredited facility, housing approximately 5,000 nonhuman primates of nine different species. The largest research program involves infectious disease research, concentrating on the study of AIDS. Other areas of research include gene therapy, reproduction, vaccine studies, malaria, Lyme disease, tuberculosis, antiviral therapy, and clinical nonhuman primate medicine and surgery.
Responsibilities include general medical and surgical care of breeding colony and research animals, and provision of research support and training of investigators, veterinary students and technicians. The successful candidate will be provided opportunities to participate in the conduct of independent or collaborative clinical research directed toward nonhuman primate medicine and surgery.
The candidate must hold a DVM/VMD degree from an AVMA-accredited college of veterinary medicine and be licensed to practice veterinary medicine in one of the 50 states. ACLAM, ACVIM, or ACVECC board certification or eligibility is desirable. The candidate should have good verbal and written communication skills and the ability to interact positively with others. This position will remain open until a qualified candidate is selected.
Interested persons should submit a letter of interest, CV, and a list of three references to: Rudolf P. Bohm, Jr., DVM, Chair, Div. of Veterinary Medicine, Tulane NPRC, 18703 Three Rivers Rd, Covington, LA 70433 [985-871-6266; e-mail: [email protected]].
Environmental Enrichment Coordinator - Tulane
The Tulane National Primate Research Center, Division of Veterinary Medicine, is seeking an Environmental Enrichment Program and Research Coordinator, who will oversee the environmental enrichment program, ensure compliance of the facility�s enrichment plan with regulatory agencies, and serve as a liaison to research and other departments. Responsibilities include maintaining appropriate documentation and monitoring nonhuman primate behavior, collecting behavioral data, planning and evaluating enrichment strategies, and assisting with personnel training programs. (This individual will work with environmental enrichment, veterinary, animal care, and research staff to facilitate the implementation of behavioral management techniques.) The successful applicant will seek independent funding, conduct independent and collaborative research projects, prepare manuscripts for publication, and participate in national meetings.
Qualifications include � PhD in anthropology, psychology, zoology, or a related field; � at least 3-5 years of directly-related experience, including supervisory experience; � demonstrated experience in implementing or planning environmental enrichment techniques for animals to improve their well-being; � familiarity with animal welfare legislation and policies; � excellent time management and organizational skills; � computer skills in word processing, data management and analysis, and development of training material; � excellent oral and written communication skills, and the ability to work independently.
Applicants should submit a CV and representative publications, and have three letters of reference sent, to Rudolf P. Bohm, Jr., at the address above.
Research Fellow in Reproductive Biology - Florida
The Wildlife Tracking Center at Disney�s Animal Kingdom has a research fellowship available in animal reproduction. This position will focus on monitoring the reproductive functioning and efficacy of contraceptives in exotic animals with a primary emphasis on hippos, giraffes and New World primates. The individual will be responsible for analysis of hormonal data and will be actively involved in sharing this information with our staff and professional colleagues. Given Disney�s Animal Kingdom�s commitment to sharing scientific information as it relates to long-term conservation efforts, this position requires the ability to interact and share information with guests of all ages and enjoy working in a laboratory that is visible to the public.
Other qualifications include � a bachelor�s or master�s degree; � experience with enzyme immunoassay techniques (essential); � ability to work with others in a fast-paced, team-oriented atmosphere; � effective public speaking skills and strong publication record (preferred). The salary will be $25,000-30,000, dependent upon experience. Applications should be received by June 30, to begin work in September. This is a one-year appointment, renewable for a second year.
Send a copy of CV and three letters of recommendation to Dr. Anne Savage, Disney�s Animal Kingdom, P.O. Box 10000, Lake Buena Vista, FL 32830. For more information, contact Dr. Savage [407-938-2837; fax: 407-939-6391; e-mail: [email protected]].
Director, Center for Comparative Medicine - Texas
Baylor College of Medicine seeks a Laboratory Animal Veterinarian to assume academic and administrative leadership of the Center for Comparative Medicine (CCM), a large, AAALAC-accredited, state-of-the-art program located in Houston, Texas. The CCM oversees all animal research facilities at Baylor College of Medicine. The combined facilities house species that range from rodents and rabbits to nonhuman primates and farm animals, and include extensive SPF barrier housing for rodents. Baylor College of Medicine is a leading academic center in mouse genetics, phenotyping, behavior, and imaging. The position offers outstanding opportunities to participate in collaborative research programs in molecular genetics, functional genomics, infectious diseases, cancer research, and neuroscience, and carries an associated academic title appropriate to the candidate�s experience.
The Director of the CCM provides leadership to a large staff of veterinarians, pathologists, veterinary technicians, and animal husbandry and administrative support staff. The position involves considerable interaction with academic faculty in the design and execution of research projects, and facilitation of communication between academic research staff and CCM staff. The Director of the CCM must take a leading role in planning and designing animal research facilities, grant writing to support institutional animal needs, and articulation of an exciting vision for the role of the Center and animal research in a large and growing academic research enterprise. Establishment of an ACLAM-certified laboratory animal medicine training program and PhD program in Comparative Medicine must be a priority.
Qualifications include a DVM or equivalent from an AVMA-accredited school of veterinary medicine, Diplomate status in the American College of Laboratory Animal Medicine, and at least five years� experience in laboratory animal science. Also expected are knowledge of the technical aspects of animal research and animal housing facilities for rodents and large animals and of regulations pertaining to animal research and facility operations, to provide assistance to the IACUC and other College personnel. Strong administrative, communication and management skills are essential, as are prior budget management and supervisory experience.
To apply, send a letter, CV, and list of three references to: Dr. James Patrick, Vice President and Dean of Research, Baylor College of Medicine, Houston, TX 77030 [713-798-6970; fax: 713-798-5902; e-mail: [email protected]].
Research Assistant - Philadelphia
Three positions are available at Thomas Jefferson University for individuals interested in being part of a research team studying cognitive and neurochemical/neuropathological aspects of chronic low level exposure to manganese. The positions involve the training and testing of macaques on a variety of computerized and non-computerized attention, memory, and motor function tasks and assessing behavioral consequences of heavy metal exposure. In addition to the training and testing of animals, these positions also involve administering enrichment paradigms, administering routine medications and treatments, and maintaining all appropriate study records. The research assistants will also assist in conducting functional brain imaging studies. Opportunities also exist for applicants with expertise in quantitative autoradiography, immunohistochemistry, and/or in situ hybridization histochemistry to be involved in neurobiological studies of receptor binding and localization and gene expression studies in post-mortem brain tissue.
Qualifications include a BS, MS or PhD in neuroscience, biology, or psychology; experience with or desire to work with nonhuman primates; and familiarity with cognitive assessment and behavioral observation methods. Ability to work independently yet be part of a research team is essential, as are good interpersonal and communication skills and ability to work with computers.
Contact J. S. Schneider, Thomas Jefferson University, Dept of Pathology, Anatomy, and Cell Biology, 1020 Locust St, Philadelphia, PA 19107 [fax: 215-923-3808; e-mail: [email protected]].
Quality Assurance Officer - Madison, Wisconsin
The Wisconsin Primate Research Center is seeking to hire a Quality Assurance Officer. A bachelor�s degree in biological sciences or a related field is required, but a master�s degree is preferred. Other requirements include one to two years� experience, including administrative and research responsibilities, preferably in a biological, medical education, or biomedical research setting; a clear understanding of the Animal Welfare Act of 1984 and PHS policies related to research animal use; and previous training experience.
Principal duties include, but are not limited to the following: � overseeing the review process for animal research protocols; � interpreting, analyzing, and applying federal, state and university guidelines to promote ethical practices and ensure compliance with regulations; � analyzing research protocol submissions to determine compliance with regulations; � managing a filing system for active animal protocols; � developing a system for on-line protocol submission; � using professional interpersonal and communication skills to effectively gain understanding and resolve problems; � applying an understanding of research regulations and guidelines to provide training for Wisconsin PRC investigators and staff.
Send a resume and cover letter referring to Listing #41062 to Dr. Joseph Kemnitz, c/o Susan Carlson, Wisconsin PRC, 1220 Capitol Ct, Madison WI 53715-1299 [fax: 608-263-1984; e-mail: [email protected]]. UW-Madison is an EO/AAE.
Animal Care/Lab Tech - Stanford
An animal care/lab technician is sought for a neurophysiology lab that investigates the neural mechanisms of sensory-motor coordination and designs neural prosthetic systems. Duties include daily monitoring of animal health; maintenance of health records; performing or assisting in routine veterinary procedures; surgery preparation and assistance; monitoring animals during post-operative recovery; administering routine medications and treatments; assisting with the program for psychological enrichment for nonhuman primates; assisting in ensuring that the laboratory meets all federal, state and local regulations; training incoming lab members on proper health, safety and lab procedures; ordering supplies and equipment; training nonhuman primates on behavioral task requirements; becoming adept at working with animals and with the computer software used in the lab to run training sessions; and performing various other duties as necessary for the functioning of the laboratory.
Qualifications include a bachelor�s degree in psychology, biology, neuroscience, or a related discipline; excellent interpersonal skills; strong communication and organizational skills; and working proficiency with PC computer systems and spreadsheet programs (e.g., Excel). Experience working with nonhuman primates is preferred.
Contact: Prof. Krishna Shenoy, 319 CISX, 330 Serra Mall, Dept of Elec. Engineering and Neurosciences Program, Stanford Univ., Stanford, CA 94305 [650-723-4789; fax: 650-723-4659; e-mail: [email protected]].
Veterinarian - SFBR, Texas
The Southwest Foundation for Biomedical Research (SFBR) is seeking a veterinarian to provide clinical and surgical support for a colony of about 4,000 nonhuman primates (baboons, chimpanzees, and macaques) and over 2,000 other animals; provide support to research programs in the areas of hepatitis, AIDS, genetic-related diseases, reproductive diseases, and blood substitutes; assist in training programs for animal care personnel; and conduct collaborative research. There will be other duties as designated by supervisor. This veterinarian will report to the Chair of Laboratory Animal Medicine.
A DVM or VMD from an accredited veterinary school and veterinary work experience are required. Residency training and experience with laboratory animals are preferred. Candidates must be licensed in the U.S. to practice veterinary medicine; have a valid driver�s license; and meet SFBR driving standards.
This work may involve exposure to radioactive materials, toxic materials, and animals infected with infectious agents such as HIV-1, HIV-2, HBV, HCV, HEV, HXV, herpes B, and influenza. Employee may drive a motor vehicle on SFBR business, and must be able to lift 20 pound sedated animals and carry them for short distances without assistance. Participation in a medical monitoring and surveillance program is required. Immunization for hepatitis B is required unless waived by the employee.
Please mail a CV, along with names and contact details of three references, to Director of Human Resources (02-025), SFBR, P.O. Box 760549, San Antonio, TX 78245-0549. Please provide expendable copies of any documents you submit.
* * *
Grants Available
Neuroscience Research on Drug Addiction
The National Institute on Drug Abuse (NIDA) announces a program intended to encourage investigator interest in the wide range of neuroscience research relevant to drug abuse, drug dependence, and drug addiction. Of particular interest are new areas of neuroscience that may be applied to questions of drug abuse and addiction.
Specific research topics emphasized in the program announcement include: * Animal models to study the neurobiology of addiction; * Vulnerability to drug addiction; * Neuroadaptation and neurotoxicity; * Developmental (ontogenetic) neurobiology; * Drugs and learning, memory, and cognition; * Pain and analgesia; * Drug abuse effects on sensory and perceptual systems; * Drugs of abuse, sleep and circadian rhythms; * Neuropsychopharmacology of drugs of abuse; * Drugs of abuse and CNS interaction with other systems; * Neural cell biology; * Prescription drug abuse; and * NeuroAIDS.
For information about the program, see <grants.nih.gov/grants/guide/pa-files/PA-02-085.html>; or contact Roger M. Brown, Assoc. Dir. for Neuroscience Coordination, Div. of Neuroscience and Behavior Research, 600 Executive Blvd, MSC 9555, Bethesda, MD 20892-9555 [301-443-1887; fax: 301-594-6043; e-mail: [email protected]]. Applications must be received by or mailed on or before the receipt dates described at <grants.nih.gov/grants/funding/submissionschedule.htm>.
Veterinary Acupuncture
The International Veterinary Acupuncture Society (IVAS) wishes to support high-quality research utilizing acupuncture. Accordingly IVAS is soliciting research proposals for funding with a budget of up to $20,000. Funding period is November 1, 2002, to October 31, 2003. Proposals evaluating the effects of acupuncture, the mechanisms of action of acupuncture, or where acupuncture is utilized in animals of any species to provide treatment for a specific problem, syndrome, or disease will be considered. In the proposed studies, the use of objective measures of outcome is preferred.
A pre-proposal will be due August 26, 2002. Following review, successful applicants will be asked to submit a longer, more detailed application. For information, and details about the pre-proposal, contact IVAS, P.O. Box 271395, Ft. Collins, CO 80527-1395 [970-266-0666; fax: 970-266-0777; e-mail: [email protected]].
* * *
Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
Books
* The Human Fossil Record. Volume 1. Terminology and Craniodental Morphology of Genus Homo (Europe). J. H. Schwartz & I. Tattersall. Somerset, NJ: Wiley-Liss, 2002. [Price: $125.00]
. . .
Contents: Part 1: Terminology and craniodental morphology of genus Homo. Part 2: Site-by-site atlas of European hominid fossils.
* All Apes Great and Small. Volume 1: African Apes. B. M. F. Galdikas, N. E. Briggs, L. K. Sheeran, G. L. Shapiro, & J. Goodall (Eds.). Hingham, MA: Kluwer, 2001. [Price: $75.00]
. . .
Contents: I - Issues in ape and human evolution: African apes as time machines, by R. Wrangham & D. Pilbeam; Primate divergence times, by A. Janke & U. Arnason; The cerebellum: An asset to hominoid cognition, by C. E. MacLeod, K. Zilles, A. Schleicher, & K. R. Gibson.
. . .
II - Bonobos, the �forgotten ape�? The status of the bonobo (Pan paniscus) in the Democratic Republic of Congo, by J. Dupain & L. Van Elsacker; The status of bonobos in their southernmost geographic range, by J. A. Myers Thompson; Current situation of the bonobos in the Luo Reserve, Equateur, Democratic Republic of Congo, by C. Hashimoto & T. Furuichi.
. . .
III - Chimpanzees, the best-known ape: Pan in pandemonium, by S. T. Boysen & T. Butynski; Predation of mammals by the chimpanzees of the Mahale Mountains, Tanzania, by K. Hosaka, T. Nishida, M. Hamai, A. Matsumoto-Oda, & S. Uehara; Representational capacities in the chimpanzees: Numerical and spatial reasoning, by S. T. Boysen & V. A. Kuhlmeier.
. . .
IV - Gorillas, the greatest of the apes: The status of gorillas worldwide, by F. G. P. Patterson & M. L. Matevia; Twenty-seven years of Project Koko and Michael, by F. G. P. Patterson & W. Gordon; Who�s in charge? Observations of social behavior in a captive group of western lowland gorillas, by T. A. Stein.
. . .
V - Physiological bases for behavior and aging: Great apes and humans: The Great Ape Aging Project: A resource for the comparative study of behavior, cognition, health, and neurobiology, by J. M. Erwin, M. Bloomsmith, S. T. Boysen, P. R. Hof, R. Holloway, L. Lowenstine, R. McManamon, D. P. Perl, W. Young, & A. Zihlman; An international database for the study of diabetes, obesity, and aging in great apes and other nonhuman primates, by R. Martens, R. Couch, B. Hansen, C. Howard, J. Kemnitz, D. P. Perl, & J. M. Erwin; Studies of age-related neuronal pathology in great apes, by D. P. Perl, P. R. Hof, E. A. Nimchinsky, & J. M. Erwin; Metabolites of ovarian hormones and behavioral correlates in captive female bonobos (Pan paniscus), by M. H. Jurke, L. R. Hagey, N. M. Czekala, & N. C. Harvey; Sexual motivation of male chimpanzees during the female cycle, including preliminary data on age effects, by R. D. Nadler.
. . .
VI - The bushmeat crisis: African apes at risk: Bushmeat, primate kinship, and the global conservation movement, by A. L. Rose; Status of the proposed Lomako Forest bonobo reserve: A case study of the bushmeat trade, by J. Dupain & L. Van Elsacker; What happened to Gorilla gorilla uellensis? A preliminary investigation, by K. Ammann & N. Briggs; Apes, persons, and bioethics, by P. Singer & P. Cavalieri.
* The Evolution of Neogene Terrestrial Ecosystems in Europe. Volume 1 of Hominoid Evolution and Climatic Change in Europe. J. Agusti, L. Rook, & P. Andrews (Eds.). New York: Cambridge University Press, 1999. [Price: $100.00]
* Almost Human: A Journey Into the World of Baboons (with a new introduction and epilogue). S. C. Strum. Chicago: University of Chicago Press, 2001. [Price: $18.00]
* Dynamics in Human and Primate Societies: Agent-Based Modeling of Social and Spatial Processes. T. A. Kohler & G. J. Gumerman (Eds.). Cary, NC: Oxford University Press, 2000. [Price: $65.00 (cloth); $40.00 (paper)]
* Primates Face to Face: Conservation Implications of Human-Nonhuman Primate Interconnections. A. Fuentes & L. D. Wolfe (Eds.). New York: Cambridge University Press, 2002. [Price: $90.00]
. . .
Contents: Foreword, by K. B. Strier. I: Science and nonhuman primates: Anthropology and primatology, by P. Dolhinow; Resistance to the cross-species perspective in anthropology, by M. M. Pavelka; The ethics and efficacy of biomedical research in chimpanzees with special regard to HIV research, by R. S. Fouts, D. H. Fouts, & G. S. Waters.
. . .
II: Cultural views of nonhuman primates: Monkey as food, monkey as child: Guaja symbolic cannibalism, by L. Ann Cormier; Ethnoecology of monkeys among the Bari of Venezuela: Perception, use and conservation, by M. Lizarralde; Primates in Matsigenka subsistence and world view, by G. H. Shepard; Monkey King in China: Basis for a conservation policy? by F. D. Burton; Reflections on the concept of nature and gorillas in Rwanda: Implications for conservation, by P. Sicotte & P. Uwengeli.
. . .
III: Conservation of nonhuman primates: Monkeys, humans and politics in the Mentawai Islands: No simple solutions in a complex world, by A. Fuentes; Conservation must pursue human-nature biosynergy in the era of social chaos and bushmeat commerce, by A. L. Rose; A cultural primatological study of Macaca fascicularis on Ngeaur Island, Republic of Palau, by B. Wheatley, R. Stephenson, H. Kurashina, & K. Marsh-Kautz; Monkeys in the backyard: Encroaching wildlife and rural communities in Japan, by D. S. Sprague.
. . .
IV: Government actions, local economies and nonhuman primates: The primatologist as minority advocate, by A. A. Eudey; Monkey business? The conservation implications of macaque ethnoprimatology in southern Thailand, by L. E. Sponsel, N. Ruttanadakul, & P. Natadecha-Sponsel; Rhesus macaques: A comparative study of two sites, Jaipur, India, and Silver Springs, Florida, by L. D. Wolfe.
* Nouragues: Dynamics and Plant-Animal Interactions in a Neotropical Rainforest. F. Bongers, P. C. Dominique, P.-M. Forget, & M. Thery (Eds.). Hingham, MA: Kluwer Academic Publ., 2001. [Price: $115.00]
. . .
Contents include: Diet and population densities of the primate community in relation to fruit supplies, by B. Simmen, C. Julliot, F. Bayart, & E. Pages-Feuillade; Comparative positional behaviour of five primates, by D. Youlatos & J.-P. Gasc; and Frugivory and seed dispersal by three neotropical primates: Impact on plant regeneration, by C. Julliot, B. Simmen, & S. Zhang.
* Primate Dentition: An Introduction to the Teeth of Non-Human Primates. D. R. Swindler. Illustrated by R. M. George. New York: Cambridge University Press, 2002. [Price: $80.00]
* Towards New Horizons in Primate Toxicology: Perspectives for the New Millennium. R. Korte & G. F. Weinbauer (Eds.). M�nster, Germany: Waxmann, 2000. [Price: EUR 50.10 ($44.70)]
* Aging in Nonhuman Primates. Interdisciplinary Topics in Gerontology, Vol. 31. J. M. Erwin & P. R. Hof (Eds). Basel: Karger, 2002. [Price: $172.25]
. . .
Contents: One gerontology: Advancing understanding of aging through studies of great apes and other primates, by J. M. Erwin, P. R. Hof, J. J. Ely, & D. P. Perl; Skeletal and dental evidence of aging in captive western lowland gorillas: A preliminary report, by K. A. Nichols & A. L. Zihlman; Skeletal aging in macaque monkeys, by R. J. Colman & N. Binkley; Getting old at Gombe: Skeletal aging in wild-ranging chimpanzees, by M. E. Morbeck, A. Galloway, & D. R. Sumner; Nutritional modulation of aging by caloric restriction, by J. A. Mattison, D. K. Ingram, G. S. Roth, & M. A. Lane; Methodological considerations when studying the aging process in the nonhuman primate brain, by J. I. H. Keuker, T. Michaelis, G. de Biurrun, P. G. M. Luiten, M. P. Witter, & E. Fuchs; Nigrostriatal function in aged nonhuman primates, by M. E. Emborg & J. H. Kordower; The brain of the aging baboon: A nonhuman primate model for neuronal and glial tau pathology, by C. Schultz, K. del Tredici, U. R�b, E. Braak, G. B. Hubbard, & H. Braak; Comparative neuropathology of brain aging in primates, by P. R. Hof, E. P. Gilissen, C. C. Sherwood, H. Duan, P. W. H. Lee, B. N. Delman, T. P. Naidich, P. J. Gannon, D. P. Perl, & J. M. Erwin; The aged rhesus macaque in neuroscience research: Importance of the nonhuman primate model, by J. A. Roberts; The rhesus monkey model as a heuristic resource in cognitive aging research, by J. G. Herndon & A. Lacreuse; Comparative models of cognitive decline in aging great apes, by J. A. Corr, L. J. Martin, & S. T. Boysen; and The behavior of aged great apes, by L. R. Tarou, M. A. Bloomsmith, M. P. Hoff, J. M. Erwin, & T. L. Maple.
* 25th Anniversary Album. Oklahoma City: American Society of Primatologists, 2002. 90 pp. (Price: $15, including postage, from Janette Wallis, Dept of Psychiatry & Behavioral Sci, Univ. of Oklahoma Health Sci. Ctr, P.O. Box 26901, Oklahoma City, OK 73190 [405-627-8820; fax: 405-271-3808; e-mail: [email protected]])
Handbooks
* Veterinary Drug Handbook. Fourth Edition. D. C. Plumb. Ames: Iowa State Univ. Press, 2002. [Price: $59.99]
* Methods and Welfare Considerations in Behavioral Research with Animals. Bethesda: NIH, 2002. [Available at <www.nimh.nih.gov/research/animals.cfm>; or phone 301-443-4513]
. . .
A handbook to assist Institutional Animal Care and Use Committees in the evaluation of protocols that employ various means to manipulate the behavior and health of laboratory animals.
Magazines and Newsletters
* ASP Bulletin, March/April, 2002, 26[1]. [Janette Wallis, Dept of Psychiatry and Behavioral Sciences, Univ. of Oklahoma Health Sciences Center, P.O. Box 26901, Oklahoma City, OK 73104-5020]
. . .
Includes reports by recipients of ASP Conservation Awards and Grants: �Littoral forest primate fauna in the Tolagnaro (Fort Dauphin) region of southeastern Madagascar�, by M. Banks; �Monitoring the behavioral ecology and viability of forest fragment chimpanzees, Masindi district, Uganda�, by J. Wallis; �Primates status survey and conservation priorities in Namdapha National Park, Arunachal Pradesh, India�, by D. Chetry; and �Survey of primates in West Kameng District, Arunachal Pradesh, India�, by A. Choudhury.
* Connection, Winter/Spring 2002. [AAALAC International, 11300 Rockville Pike, Suite 1211, Rockville, MD 20852-3035]
. . .
Includes articles on IACUC training, and on the semiannual review of animal care and use programs.
* CPCSEA Newsletter, October-December 2001 (Committee for the Control and Supervision of Experiments on Animals, Dept of Animal Welfare, Government of India, 3rd Seaward Rd, Valmiki Nagar, Thiruvanimiyur, Chennai - 600 041, India [e-mail: [email protected]])
* Going Ape at Ohio State! (Comparative Cognition Project, OSU Chimpanzee Center, 242 Townshend, 1885 Neil Ave, Columbus, OH 43210-1222)
* The Newsletter, 2002, 13[2]. [Primate Foundation of Arizona, P.O. Box 20027, Mesa, AZ 85277-0027]
. . .
Includes �The Semliki Chimpanzee Project�, by T. O�Mara & K. Hunt; and �On the incidence and prevalence of diabetes in captive chimpanzees (Pan troglodytes): An update on a call for information�, by J. Fritz.
* Neotropical Primates: A Newsletter of the Neotropical Section of the IUCN/SSC Primate Specialist Group, August, 2001, 9[2]. [Conservation International, 1919 M St, NW, Suite 600, Washington, DC 20036]
. . .
Contents include: Standard errors of survey estimates: What do they mean? by W. E. Magnusson; Primatas da Regi�o do Rio Tapajós, Pará, Brasil, by S. M. Vaz; Body weights before and after first pregnancies of immigrant adult female mantled howling monkeys (Alouatta palliata) in Costa Rica, by E. L. Zucker, M. R. Clarke, & K. E. Glander; Food resources and the survival of a group of howler monkeys (Alouatta palliata mexicana) in disturbed and restricted habitat at Los Tuxlas, Veracruz, Mexico, by F. Gómez-Marin, J. J. Veá, E. Rodriguez-Luna, F. Garcia-Orduña, D. Canales-Espinosa, M. EScobar, & N. Asensio; Multiple breeding females and allo-nursing in a wild group of moustached tamarins (Saguinus mystax), by A. C. Smith, E. R. Tirado Herrera, H. M. Buchanan-Smith, & E. W. Heymann; and Densidade e conserva��o do bugio (Alouatta fusca) no Parque Estadual Intervales, by S. Steinmetz.
* Neotropical Primates: A Newsletter of the Neotropical Section of the IUCN/SSC Primate Specialist Group, December, 2001, 9[3]. [Conservation International, 1919 M St, NW, Suite 600, Washington, DC 20036]
. . .
Contents include: Body weights of adult female Alouatta palliata in Costa Rica increase with age, by E. L. Zucker, M. R. Clarke, & K. E. Glander; Variations in group size in white-faced sakis (Pithecia pithecia): Evidence for monogamy or seasonal congregations? by S. M. Lehman, W. Prince, & M. Mayor; Diurnal activity budgets of black spider monkeys, Ateles chamek, in a southern Amazonian tropical forest, by R. B. Wallace; Distribución parcial del tití gris (Saguinus leucopus, Callitrichidae) en el departamento de Antioquia, Colombia, by C. A. Cuartas-Calle; Drinking by howler monkeys (Alouatta fusca) and its seasonality at the Intervales State Park, S�o Paulo, Brazil, by S. Steinmetz; Uso de plantas como alimento por monos aulladores, Alouatta palliata, en el Parque Yumká, Tabasco, México, by Y. García del Valle, D. Muñoz, M. Magaña-Alejandro, A. Estrada, & B. Franco; Alouatta guariba clamitans (Cabrera, 1940): A new southern limit for the species and for neotropical primates, by R. C. Printes, M. V. A. Liesenfeld, & L. Jerusalinsky; and Two taxonomies of the New World Primates - A comparison of Rylands et al. (2000) and Groves (2001), by A. B. Rylands.
* Reaching Out, 2002, 4[1]. (Primate Rescue Center, Inc., 5087 Danville Rd, Nicholasville, KY 40356 [e-mail: [email protected]; <www.primaterescue.org>])
Manuals
* Management of Lorises in Captivity: A Husbandry Manual for Asian Lorisines (Nycticebus & Loris ssp.) . H. Fitch-Snyder and H. Schulze (Eds.). San Diego: Zoological Society of San Diego, 2001. (A limited number of copies are available from the publisher, but copies may be borrowed from the WiPRC Library, 1223 Capitol Ct,, Madison, WI 53715 [fax: 1-608-263-3512; e-mail: [email protected]])
. . .
The information in this manual has been compiled from numerous articles, reports, and personal experiences by the authors and editors. Because new information is continuously being discovered, it is tempting to keep delaying publication until the husbandry manual is even more complete. However, as an increasing number of institutions become involved with the management of these specialized prosimians, it has become essential to have this information available in a usable format. The editors plan to periodically distribute updated and new information to SSP Institution Representatives and other appropriate recipients. Users of this publication are encouraged to contribute additional material for future updates.
Reports
* Orangutan Reintroduction and Protection Workshop, 15-18 June, 2001, Wanariset-Samboja and Balikpapan, E. Kalimantan, Indonesia, Final Report. [Price: $35.00, from IUCN/SSC CBSG, 12101 Johnny Cake Ridge Rd, Apple Valley, MN 55124]
. . .
Report of a workshop sponsored by the Gibbon Foundation, the Wanariset Orangutan Reintroduction Project, the Balikpapan Orangutan Survival Foundation, and the Balikpapan Orangutan Society, in collaboration with the Primate Specialist Group (SSC/IUCN PSG) and the Conservation Breeding Specialist Group (SSC/IUCN CBSG).
Special Journal Issues
* Malaria. Nature Insight, 2002, 415[6872]. <www.nature.com/nature/insights/6872.html>
* American Genetic Association Symposium Issue: Primate Evolutionary Genetics, May 19-20, 2001. Journal of Heredity, 2001, 92[6].
. . .
Contents: Primate evolutionary genetics, by Oliver A. Ryder; Analysis of the complete human mtDNA genome: Methodology and inferences for human evolution, by M. Ingman & U. Gyllensten; Divergent origins and concerted expansion of two segmental duplications on chromosome 16, by E. E. Eichler, M. E. Johnson, C. Alkan, E. Tuzun, C. Sahinalp, D. Misceo, N. Archidiacono, & M. Rocchi; Human and ape molecular clocks and constraints on paleontological hypotheses, by R. L. Stauffer, A. Walker, O. A. Ryder, M. Lyons-Weiler, & S. Blair Hedges; Modern African ape populations as genetic and demographic models of the last common ancestor of humans, chimpanzees, and gorillas, by M. I. Jensen-Seaman, A. S. Deinard, & K. K. Kidd; Genomic divergence between human and chimpanzee estimated from large-scale alignments of genomic sequences, by F.-C. Chen, E. J. Vallender, H. Wang, C.-S. Tzeng, & W.-H. Li; Mitochondrial 16S rRNA sequence diversity of hominoids, by R. Noda, C. G. Kim, O. Takenaka, R. E. Ferrell, T. Tanoue, I. Hayasaka, S. Ueda, T. Ishida, & N. Saitou; and Determinants of effective population size for loci with different modes of inheritance, by J. F. Storz, U. Ramakrishnan, & S. C. Alberts.
* Program and abstracts of the 25th annual meeting of the American Society of Primatologists, June 1-4, 2002. American Journal of Primatology, 2001, 57[Suppl. 1].
Studbooks
* Allen�s Swamp Monkey (Allenopithecus nigroviridis). 2000 North America Regional Studbook. S. Averill. San Diego: San Diego Zoo, 2000. (This document may be borrowed from the WiPRC. Contact Joanne Brown [608-263-3512; e-mail: [email protected]])
* De Brazza�s Monkey (Cercopithecus neglectus). European Studbook. Number 1. M. Janiszewska. Lodz, Poland: Lodz-Miejski Ogrod Zoologiczny, 2001. (See information above)
* Pied Tamarin (Saguinus bicolor). 2000 International Studbook. A. J. Baker & A. Pissinatti. Philadelphia Zoo, 2001. (See information above)
* 2000 North American Regional Studbook of the Pygmy Marmoset (Callithrix pygmaea). D. D. Aden. Denver Zoological Gardens, 2001. (See information above)
* European Studbook for the: Sooty Mangabey (Cercocebus atys atys),White-Crowned Mangabey (Cercocebus a. lunulatus), Cherry-Crowned Mangabey (Cercocebus torquatus). M. T. Abello, Parc Zoologic de Barcelona, 2001. (See information above)
* Owl Monkey (Aotus Spp.) North American Region Studbook Update. H. Colahan. Disney�s Animal Kingdom, 2001. (See information above)
Anatomy and Physiology
* Social intelligence, innovation, and enhanced brain size in primates. Reader, S. M., & Laland, K. N. (Dept of Biology, McGill Univ., 1205 Ave Docteur Penfield, Montréal, P.Q., Canada H3A 1B1 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, USA, 2002, 99, 4436-4441.
. . .
Despite interest in the evolution of intelligence, the intuitively appealing notion that brain volume and �intelligence� are linked remains untested. Here, ecologically relevant measures of cognitive ability, reported incidences of behavioral innovation, social learning, and tool use, are used to show that brain size and cognitive capacity are indeed correlated. A comparative analysis of 533 instances of innovation, 445 observations of social learning, and 607 episodes of tool use established that social learning, innovation, and tool use frequencies are positively correlated with species� relative and absolute �executive� brain volumes, after controlling for phylogeny and research effort. Moreover, innovation and social learning frequencies covary across species, in conflict with the view that there is an evolutionary tradeoff between reliance on individual experience and social cues. These findings provide an empirical link between behavioral innovation, social learning capacities, and brain size in mammals. The ability to learn from others, invent new behaviors, and use tools may have played pivotal roles in primate brain evolution.
* Synaptic distribution of ionotropic glutamate receptors in the inner plexiform layer of the primate retina. Gr�nert, U., Haverkamp, S., Fletcher, E. L., & W�ssle, H. (H. W., Max-Planck-Inst. f�r Hirnforschung, Deutschordenstr. 46, D-60528 Frankfurt, Germany). Journal of Comparative Neurology, 2002, 447, 138-151.
. . .
The distribution and synaptic clustering of glutamate receptors (GluRs) were studied in the inner plexiform layer (IPL) of the macaque monkey retina by using subunit specific antisera. A punctate immunofluorescence pattern was observed in the IPL for all subunits tested, and electron microscopy confirmed that the immunoreactive puncta represent clustering of receptors at sites postsynaptic to the bipolar cell ribbon synapses (dyads). Usually only one of the two postsynaptic processes at the dyads expressed a given subunit. Immunoreactive GluR2, GluR2/3, and GluR4 puncta were found at high density throughout the IPL and are probably expressed at every dyad. The GluR1 subunit was expressed at lower density. The N-methyl-D-aspartate (NMDA) receptor subunits NR2A and NR1C2' were restricted to synapses localized in two broad bands in the center of the IPL. They were often colocalized with GluR2/3 and GluR4 subunits. The orphan receptor subunits d1/2 predominated in three horizontal bands. The kainate receptor subunits GluR6/7 were clustered in large postsynaptic densities adjacent to bipolar cell axon terminals but lacking a synaptic ribbon on the presynaptic side. This might represent a conventional synapse made by a bipolar axon terminal. The results suggest that GluR2/3 and GluR4, together with NMDA receptors, are preferentially expressed on ganglion cell dendrites, whereas kainate receptors and the d1/2 subunits are mostly localized on amacrine cell processes.
Animal Models
* Brain-machine interface: Instant neural control of a movement signal. Serruya, M. D., Hatsopoulos, N. G., Paninski, L., Fellows, M. R., & Donoghue, J. P. (Dept of Neuroscience, Box 1953, Brown Univ., Providence, RI 02912 [e-mail: [email protected]]). Nature, 2002, 416, 141-142.
. . .
The activity of motor cortex (MI) neurons conveys movement intent sufficiently well to be used as a control signal to operate artificial devices, but until now this has called for extensive training or has been confined to a limited movement repertoire. Here it is shown how activity from a few (7-30) MI neurons can be decoded into a signal that a monkey is able to use immediately to move a computer cursor to any new position in its workspace (14° x 14° visual angle). The results, which are based on recordings made by an electrode array that is suitable for human use, indicate that neurally based control of movement may eventually be feasible in paralyzed humans.
* Outcome of simian-human immunodeficiency virus strain 89.6p challenge following vaccination of rhesus macaques with human immunodeficiency virus Tat protein. Silvera, P., Richardson, M. W., Greenhouse, J., Yalley-Ogunro, J., Shaw, N., Mirchandani, J., Khalili, K., Zagury, J.-F., Lewis, M. G., & Rappaport, J. (Infectious Disease Research, Southern Research Institute, 431 Aviation Way, Frederick, MD 21701 [e-mail: [email protected]]). Journal of Virology, 2002, 76, 3800-3809.
. . .
The regulatory proteins Nef, Rev, and Tat of human immunodeficiency virus type 1 (HIV-1) are attractive targets for vaccine development, since induction of effective immune responses targeting these early proteins may best control virus replication. The authors investigated whether vaccination with biologically active Tat or inactive Tat toxoid derived from HIV-1IIIB and simian-human immunodeficiency virus (SHIV) strain 89.6p would induce protective immunity in rhesus macaques. Vaccination induced high titers of anti-Tat immunoglobulin G in all immunized animals by week 7, but titers were somewhat lower in the 89.6p Tat group. Dominant B-cell epitopes mapped to the amino terminus, the basic domain, and the carboxy-terminal region. Tat-specific T-helper responses were detected in 50% of immunized animals. T-cell epitopes appeared to map within amino acids 1 to 24 and 37 to 66. In addition, Tat-specific gamma interferon responses were detected in CD4+ and/or CD8+ T lymphocytes in 11 of 16 immunized animals on the day of challenge. However, all animals became infected upon intravenous challenge with 30 50% minimal infective doses of SHIV 89.6p, and there were no significant differences in viral loads or CD4+ T-cell counts between immunized and control animals. Thus, vaccination with HIV-1IIIB or SHIV 89.6p Tat or with Tat toxoid preparations failed to confer protection against SHIV 89.6p infection despite robust Tat-specific humoral and cellular immune responses in some animals. Given its apparent immunogenicity, Tat may be more effective as a component of a cocktail vaccine in combination with other regulatory and/or structural proteins of HIV-1.
* Tat-vaccinated macaques do not control simian immunodeficiency virus SIVmac239 replication. Allen, T. M., Mortara, L., Mothé, B. R., Liebl, M., Jing, P., Calore, B., Piekarczyk, M., Ruddersdorf, R., O�Connor, D. H., Wang, X., Wang, C., Allison, D. B., Altman, J. D., Sette, A., Desrosiers, R. C., Sutter, G., & Watkins, D. I. (D. I. W., WiPRC, 1220 Capitol Ct., Madison, WI 53715 [e-mail: [email protected]]). Journal of Virology, 2002, 76, 4108-4112.
. . .
The regulatory proteins of human immunodeficiency virus may represent important vaccine targets. Here the role of Tat-specific cytotoxic T lymphocytes (CTL) in controlling pathogenic simian immunodeficiency virus SIVmac239 replication after using a DNA-prime, vaccinia virus Ankara-boost vaccine regimen is assessed. Despite the induction of Tat-specific CTL, there was no significant reduction in either peak or viral set point compared to that of controls.
* Rhesus offspring produced by intracytoplasmic injection of testicular sperm and elongated spermatids. Hewitson, L., Martinovich, C., Simerly, C., Takahashi, D., & Schatten, G. (G. S., 204 Craft Ave, Pittsburgh, PA 15213 [e-mail: [email protected]]). Fertility and Sterility, 2002, 77, 794-801.
. . .
Oocytes collected by laparoscopy from gonadotropin-stimulated female rhesus monkeys were injected with testicular sperm or elongated spermatids obtained from the testes of males. Cleavage stage embryos were transferred to surrogate females. Fertilization failures were fixed and processed for detection of microtubules and chromatin configurations. Fertilization, assessed by the presence of two pronuclei within 15 hours after injection, was 60% for intracytoplasmic sperm injection with testicular sperm and 47% for elongated spermatid injection. Fertilized zygotes co-cultured in Connaughts Medical Research Labs medium on a Buffalo Rat Liver cell monolayer resulted in hatched blastocysts after testicular sperm extraction-intracytoplasmic sperm injection and elongated spermatids. Embryos transferred at the 4- to 8-cell stage gave rise to three pregnancies: 2/3 from testicular sperm and 1/1 from an elongated spermatid. Three healthy infants were delivered by caesarean section. Oocytes that failed to fertilize typically remained arrested in metaphase of meiosis.
* Acute phase cytotoxic T lymphocyte escape is a hallmark of simian immunodeficiency virus infection. O�Connor, D. H., Allen, T. M., Vogel, T. U., Jing, P., DeSouza, I. P., Dodds, E., Dunphy, E. J., Melsaether, C., Mothé, B., Yamamoto, H., Horton H., Wilson, N., Hughes, A. L., & Watkins, D. I. (D. I. W., WiPRC, Univ. of Wisconsin, Madison, WI 53715 [e-mail: [email protected]]). Nature Medicine, 2002, 8, 493-499.
. . .
Cytotoxic T-lymphocyte (CTL) responses peak coincident with the decline in acute HIV viremia. Despite two reports of CTL-resistant HIV variants emerging during acute infection, the contribution of acute CTL escape to HIV pathogenesis remains unclear. Difficulties inherent in studying acute HIV infection can be overcome by modeling virus-host interactions in SIV-infected rhesus macaques. Twenty-one complete simian immunodeficiency virus (SIV)mac239 genomes were sequenced at four weeks post-infection to determine the extent of acute CTL escape. Here it is shown that viruses from 19 of 21 macaques escaped from CTLs during acute infection and that these escape-selecting CTLs were responsive to lower concentrations of peptide than other SIV-specific CTLs. Interestingly, CTLs that require low peptide concentrations for stimulation (high �functional avidity�) are particularly effective at controlling other viral infections. The results suggest that acute viral escape from CTLs is a hallmark of SIV infection and that CTLs with high functional avidity can rapidly select for escape variants.
* Evaluation in nonhuman primates of vaccines against Ebola virus. Geisbert, T. W., Pushko, P., Anderson, K., Smith, J., Davis, K. J., &. Jahrling, P. B. (USAMRIID, Attn: MCMR-UIP-D, 1425 Porter St, Fort Detrick, MD 21702-5011 [e-mail: [email protected]]). Emerging Infectious Diseases, 2002, <www.cdc.gov/ncidod/eid/vol8no5/01-0284.htm>.
. . .
Ebola virus (EBOV) causes acute hemorrhagic fever that is fatal in up to 90% of cases in both humans and nonhuman primates. No vaccines or treatments are available for human use. We evaluated the effects in nonhuman primates of vaccine strategies that had protected mice or guinea pigs from lethal EBOV infection. The following immunogens were used: RNA replicon particles derived from an attenuated strain of Venezuelan equine encephalitis virus (VEEV) expressing EBOV glycoprotein and nucleoprotein; recombinant Vaccinia virus expressing EBOV glycoprotein; liposomes containing lipid A and inactivated EBOV; and a concentrated, inactivated whole-virion preparation. None of these strategies successfully protected nonhuman primates from robust challenge with EBOV. The disease observed in primates differed from that in rodents, suggesting that rodent models of EBOV may not predict the efficacy of candidate vaccines in primates and that protection of primates may require different mechanisms.
Animal Welfare
* Ethics and animal welfare evaluations in South East Asian Zoos: A case study of Thailand. Agoramoorthy, G., & Harrison, B. (Sun Yat-sen Univ., P.O. Box 59-157, Kaohsiung 80424, Taiwan [e-mail: [email protected]]). Journal of Applied Animal Welfare Science, 2002, 5, 1-13.
. . .
With a case study of three zoos in Thailand, this article presents results for the first time on how ethics and welfare evaluations are conducted in southeast Asian zoos. The study identified several major and minor welfare problems and provided constructive suggestions to zoo authorities, which in turn significantly improved the standards of animal welfare. This article could serve as a model for other zoos to follow for animal welfare evaluations.
Behavior
* How cross-fostered chimpanzees (Pan troglodytes) initiate and maintain conversations. Bodamar, M. D., & Gardner, R. A. (R. A. G., Dept of Psychology, Univ. of Nevada, Reno, NV 89557-0062 [e-mail: [email protected]]). Journal of Comparative Psychology, 2002, 116, 12-26.
. . .
This study systematically sampled typical attention-getting sounds and sign language conversations between each of four originally cross-fostered chimpanzees, still living freely, but now in a laboratory setting, and a familiar human interlocutor. Videotape records showed that when they encountered a human interlocutor sitting alone at his desk with his back turned to them, the cross-fosterlings either left the scene or made attention-getting sounds. The only signs they made to the interlocutor�s back were noisy signs. When the human turned and faced them, the chimpanzees promptly signed to him (98% of the time) and rarely made any sounds during the ensuing signed conversations. Under systematic experimental conditions, the signed responses of the chimpanzees were appropriate to the conversational styles of the human interlocutor, confirming daily field observations.
* The relationship between problem solving and inhibitory control: Cotton-top tamarins (Saguinus oedipus) performance on a reversed contingency task. Kralik, J. D., Hauser, M. D., & Zimlicki, R. (Dept of Neurobiology, Duke Univ. Med. Ctr, DUMC-Box 3209, Durham, NC 27710 [e-mail: [email protected]]). Journal of Comparative Psychology, 2002, 116, 39-50.
. . .
To explore the relationship between problem solving and inhibitory control, the authors present four experiments on cotton-top tamarins using a reverse-reward contingency task. In Experiment 1, a group of tamarins was given a choice between a small and a large quantity of food. Whichever quantity the tamarins reached for first, they received the alternative. The tamarins consistently picked the larger quantity, thereby receiving the smaller. A second group of tamarins was given the same task, except that if they reached for the larger quantity of food, they received nothing. The tamarins continued to pick the larger quantity, even though this resulted in no food. In addition, most of the tamarins continued to pick the larger quantity even when the food payoff for choosing the smaller quantity was increased (Experiment 2) or when the visual salience of the food was reduced (Experiment 3). Experiment 4 was based on the finding that chimpanzees (Pan troglodytes) that have been trained on the concept of number can solve the reversed contingency task if the food is replaced by Arabic numerals. With the help of a color association, and a higher cost incurred by picking the color associated with three food items, the tamarins learned to pick the color associated with one food item. These results are compared with those obtained from studies of other primate species, highlighting the importance of comparative studies of problem solving that use comparable methods.
* Monitoring spatial transpositions by bonobos (Pan paniscus) and chimpanzees (P. troglodytes). Beran, M. J., & Minahan, M. F. (GSU Language Research Center, 3401 Panthersville Rd., Decatur, GA 30034 [e-mail: [email protected]]. International Journal of Comparative Psychology, 2002, 13, 1-15.
. . .
Two bonobos and three chimpanzees monitored spatial transpositions, or the simultaneous movement of multiple items in an array, so as to select a specific item from the array. In the initial condition of Experiment 1, a food reward was hidden beneath one of four cups, and the apes were required to select the cup containing the reward in order to receive it. In the second condition, the test board on which the cups were located was rotated 180 degrees after placement of the food reward. In the third condition, two of the three cups switched locations with one another after placement of the food reward. All five apes performed at very high levels for these conditions. Experiment 2 was a computerized simulation of the tasks with the cups in which the apes had to track one of four simultaneously moving stimuli on a computer monitor. Two of the three apes that were tested performed at a very high level for this computerized task. Therefore, members of the genus Pan can perform complex feats of spatial monitoring such as transpositions both in real-world contexts and in computerized tasks.
* Emergents and rational behaviorism. Rumbaugh, D. M. (Address same as above [e-mail: [email protected]]). Eye on Psi Chi, 2002, 6[2], 8-14 [and <www.psichi.org/content/publications/eye>].
. . .
The rational behavioristic framework here advanced allows for cognitive operations and emergent phenomena that are, in measure, based on the syntheses of general life experiences and observations and also upon the learning induced by traditional respondent and operant conditioning. The framework is based on the assumption that notably the primate brains, and particularly those of the apes and humans, have evolved so as to be acutely sensitive to the detection and storage of predictive relationships. Memory systems store the knowledge thereby derived in a logical set of �files,� which allows the natural operations of the brain to formulate creative behavioral patterns and solutions to novel challenges. Metaphorically, their storage might be thought of in terms of chords which, in their interaction with other memory chords, can be synergistic through the natural physics known otherwise to be characteristic of chords (e.g., fundamentals, resonance, harmonics, overtones, etc.).
* A nonhuman primate�s perception of object relations: Experiments on cottontop tamarins, Saguinus oedipus. Kralik, J. D., & Hauser, M. D. (Dept of Neurobiology, Duke Univ. Med. Center, Box 3209, Durham, NC, 27710 [e-mail: [email protected]]). Animal Behaviour, 2002, 63, 419-435.
. . .
Objects in nature often have spatial and functional relationships with other objects. For example, fruit may be connected to tree branches, bushes sometimes function as landmarks for home, and stones are functionally associated with nuts when they are used to crack the nuts open. Although animals may use the spatial and functional relationships between specific objects, ethologists interested in cognitive mechanisms should ask whether animals understand the spatial relationship between objects in a more general and abstract way. In this experiment, it is asked whether a small New World monkey, the cottontop tamarin, is capable of perceiving the abstract relational concept of �connectedness�. Using a traditional operant paradigm, subjects were required to press one button to images with connected objects, and another button to images with separated objects. In Phase 1 of the experiment, subjects received training with only one connected and one separated image. Following training, probe images were presented in which features such as color, texture, and shape were systematically manipulated to determine which features were more important in stimulus classification. Accuracies and reaction times of responses were recorded. On the basis of their performance, the tamarins appeared to recognize that changes in the color or texture of two objects play no functional role in determining whether such objects are connected or separated. In contrast, changes in the shape and distance between two objects does play an important functional role, and the tamarins appeared to be sensitive to such changes. In Phase 2, subjects received training with a larger set of images. After this training, classification accuracy remained significantly above chance with most probes, suggesting that the tamarins had acquired a more general connectedness concept. In Phase 3, novel images were presented. Classification accuracy was significantly above chance for many of the novel images. Overall, results suggest that tamarins perceive spatial relationships between stimuli to some extent and can classify images based in part on their spatial relationship. Other recent experiments have shown that cottontop tamarins respond to spatial relationships even better than in the current experiment when they have to act directly on the objects. The difference between these results suggests that tamarins perceive the spatial relationships between objects more readily in the context of an ecologically valid problem.
Development and Aging
* Male orangutan subadulthood: A new twist on the relationship between chronic stress and developmental arrest. Maggioncalda, A. N., Czekala, N. M., & Sapolsky, R. M. (Dept of Anthropological Sciences, Bldg 360, Stanford University, Stanford, CA 94305-2117 [e-mail: [email protected]]). American Journal of Physical Anthropology, 2002, 118, 25-32.
. . .
Both in the wild and in captivity, a marked and enduring arrest of secondary sexual development occurs in some male orangutans (Pongo pygmaeus). Researchers have hypothesized that chronic stress, perhaps related to aggression from mature males, causes endocrine changes altering growth and maturation rates in these males. In this study, urine samples were collected over a 3-year period from 23 captive male orangutans to test the hypothesis that developmentally arrested male orangutans have an endocrine profile consistent with chronic stress. Three study males were juveniles, seven were arrested adolescents, six were developing adolescents, and seven were mature adults. Morning samples were analyzed by radioimmunoassay for levels of the stress hormones cortisol and prolactin, and group hormone profiles were compared by analysis of variance. Results indicate that developing adolescent male orangutans have a significantly higher stress hormone profile than juvenile, developmentally arrested adolescent, or adult males. These results imply that the arrest of secondary sexual development in some male orangutans is not stress-induced, but perhaps is an adaptation for stress avoidance during the adolescent or subadult period. These data, together with previously reported data on levels of gonadotropins, testicular steroids, and growth-related hormones, define endocrine profiles associated with alternative reproductive strategies for males with and males without secondary sexual features.
Disease
* Risk to human health from a plethora of simian immunodeficiency viruses in primate bushmeat. Peeters, M., Courgnaud, V., Abela, B., Auzel, P., Pourrut, X., Bibollet-Ruche, F., Loul, S., Liegeois, F., Butel, C., Koulagna, D., Mpoudi-Ngole, E., Shaw, G. M., Hahn, B. H., & Delaporte, E. (Lab. Retrovirus, UMR036, IRD, 911 Ave Agropollis, BP5045, 34032 Montpellier Cdx 1, Montpellier, France [e-mail: [email protected]]). Emerging Infectious Diseases, 2002, <www.cdc.gov/ncidod/EID/vol8no5/01-0522.htm>.
. . .
To assess human exposure to simian immunodeficiency virus (SIV) in west central Africa, 788 monkeys that were hunted in the rainforests of Cameroon for bushmeat or kept as pets were tested for SIV. Serologic reactivity suggesting SIV infection was found in 13 of 16 primate species, including four not previously known to harbor SIV. Overall, 131 sera (16.6%) reacted strongly and an additional 34 (4.3%) reacted weakly with HIV antigens. Molecular analysis identified five new phylogenetic SIV lineages. These data document for the first time that a substantial proportion of wild monkeys in Cameroon are SIV infected and that humans who hunt and handle bushmeat are exposed to a plethora of genetically highly divergent viruses.
* Association between simian virus 40 and non-Hodgkin lymphoma. Vilchez, R. A., Madden, C. R., Kozinetz, C. A., Halvorson, S. J., White, Z. S., Jorgensen, J. L., Finch, C. J., & Butel, J. S. (Baylor College of Med., Dept of Med., Section of Infectious Diseases, BCM 286, Rm N1319, 1 Baylor Plaza, Houston, TX 77030 [e-mail: [email protected]]). Lancet, 2002, 359, 817-823.
. . .
�We analyzed systemic non-Hodgkin lymphoma from 76 HIV-1-infected and 78 HIV-1-uninfected patients, and non-malignant lymphoid samples from 79 HIV-1-positive and 107 HIV-1-negative patients without tumors; 54 colon and breast carcinoma samples served as cancer controls. We used PCR followed by Southern blot hybridization and DNA sequence analysis to detect DNAs of polyomaviruses and herpesviruses. Polyomavirus T antigen sequences, all of which were SV40-specific, were detected in 64 (42%) of 154 non-Hodgkin lymphomas, none of 186 non-malignant lymphoid samples, and none of 54 control cancers. This difference was similar for HIV-1-infected patients and HIV-1-uninfected patients alike. Few tumors were positive for both SV40 and Epstein-Barr virus. Human herpesvirus type 8 was not detected. SV40 sequences were found most frequently in diffuse large B-cell and follicular-type lymphomas. These results add lymphomas to the types of human cancers associated with SV40.�
* Resistance to antifolates in Plasmodium falciparum, the causative agent of tropical malaria. Warhurst, D. C. (London School of Hygiene & Tropical Med., London WC1E 7HT, England [e-mail: [email protected]]). Science Progress, 2002, 85, 89-111.
. . .
Chloroquine resistance has led to reliance on antimalarial antifolates, in particular the synergistic combination sulfadoxine/pyrimethamine (S/P), which targets enzymatic synthesis of folate co-factors through dihydropteroate synthase (DHPS) and dihydrofolate reductase (DHFR). Resistance to S/P is now increasing and replacement antimalarials are needed. This review focuses on the activity of drugs on DHFR in malaria parasites. It attempts to interpret differences in activity of pyrimethamine and its related drugs, and to clarify how residue changes due to point mutations determine the development of resistance.
* Transgenic anopheline mosquitoes impaired in transmission of a malaria parasite. Ito, J., Ghosh, A., Moreira, L. A., Wimmer, E. A., & Jacobs-Lorena, M. (M. J.-L., Case Western Reserve Univ., Dept of Genetics, 10900 Euclid Ave, Cleveland, OH 44106-4955 [e-mail: [email protected]]). Nature, 2002, 417, 452-455.
. . .
Malaria is estimated to cause 0.7 to 2.7 million deaths per year, but the actual figures could be substantially higher, owing to under-reporting and difficulties in diagnosis. If no new control measures are developed, the malaria death toll is projected to double in the next 20 years. Efforts to control the disease are hampered by drug resistance in the Plasmodium parasites, insecticide resistance in mosquitoes, and the lack of an effective vaccine. Because mosquitoes are obligatory vectors for malaria transmission, the spread of malaria could be curtailed by rendering them incapable of transmitting parasites. Many of the tools required for the genetic manipulation of mosquito competence for malaria transmission have been developed. Foreign genes can now be introduced into the germ line of both culicine and anopheline mosquitoes, and these transgenes can be expressed in a tissue-specific manner. This is a report on the use of such tools to generate transgenic mosquitoes that express antiparasitic genes in their midgut epithelium, thus rendering them inefficient vectors for the disease. These findings have significant implications for the development of new strategies for malaria control.
* Fatal infection with human pinworm, Enterobius vermicularis, in a captive chimpanzee. Murata, K., Hasegawa, H., Nakano, T., Noda, A., & Yanai, T. (H.H., Dept of Biology, Oita Med. Univ., Hasama, Oita 879-5593, Japan [e-mail: [email protected]]). Journal of Medical Primatology, 2002, 31, 104-108.
. . .
A fatal infection with human pinworms was found in a 26-year-old chimpanzee kept in a zoo. The animal was highly emaciated, and had severe enteritis with cecal multifocal nodules and severe cholelithiasis. Histopathologically, a large number of human pinworms were observed in the nodular lesions in the cecum and intestinal wall. These migrating worms were surrounded by an inflammatory cell infiltration which lacked eosinophils. There were areas of mulitfocal hyperemia and/or hemorrhages in various organs including the entire gastrointestinal tract. Pinworms were also observed in the portal venule and parenchyma of the liver. A light infection with Strongyloides cf. stercoralis was also observed.
* Fatal infection of a pet monkey with human herpesvirus. Huemer, H. P., Larcher, C., Czedik-Eysenberg, T., Nowotny, N., & Reifinger, M. (P.O. Box 151, A-6010 Innsbruck, Austria [e-mail: [email protected]]). Emerging Infectious Diseases, 2002, <www.cdc.gov/ncidod/eid/vol8no6/01-0341-G.htm>.
. . .
Concerns have been raised about pet monkeys as a potential threat to humans. The opposite situation is reported here, a danger to pets that arises from humans. Similar to herpesvirus B (Cercopithecine herpesvirus 1), which endangers humans but not its host species, Human herpesvirus 1 can act as a �killer virus� when crossing the species barrier to New World monkeys.
Evolution, Genetics, and Taxonomy
* Strong male-driven evolution of DNA sequences in humans and apes. Makova, K. D., & Li, W.-H. (W.-H.L., Dept of Ecology & Evolution, Univ. of Chicago, 1101 East 57th St, Chicago, IL 60637 [e-mail: [email protected]]). Nature, 2002, 416, 624-626.
. . .
Studies of human genetic diseases have suggested a higher mutation rate in males than in females and the male-to-female ratio (alpha) of mutation rate has been estimated from DNA sequence and microsatellite data to be about 4-6 in higher primates. Two recent studies, however, claim that alpha is only about 2 in humans. This is smaller than the estimates (a > 4) for carnivores and birds; humans should have a higher alpha than carnivores and birds because of a longer generation time and a larger sex difference in the number of germ cell cycles. To resolve this issue, noncoding fragments on Y of about 10.4 kilobases (kb) and a homologous region on chromosome 3 in humans, greater apes, and lesser apes were sequenced. An estimate of alpha from the internal branches of the phylogeny is 5.25 (95% confidence interval [CI]: 2.44 to infinity), similar to the previous estimates, but significantly higher than the two recent ones. In contrast, for the external (short, species-specific) branches, alpha is only 2.23 (95% CI: 1.47-3.84). It is suggested that closely related species are not suitable for estimating alpha, because of ancient polymorphism and other factors. Moreover, an explanation for the small estimate of alpha in a previous study is given. This study reinstates a high alpha in hominoids and supports the view that DNA replication errors are the primary source of germline mutation.
* Intra- and interspecific variation in primate gene expression patterns. Enard, W., Khaitovich, P., Klose, J., Z�llner, S., Heissig, F., Giavalisco, P., Nieselt-Struwe, K., Muchmore, E., Varki, A., Ravid, R., Doxiadis, G. M., Bontrop, R. E., & P��bo, S. (S. P., Max-Planck-Inst. for Evolutionary Anthropology, Inselstrasse 22, D-04103 Leipzig, Germany [e-mail: [email protected]]. Science, 2002, 296, 340-343.
. . .
Although humans and their closest evolutionary relatives, the chimpanzees, are 98.7% identical in their genomic DNA sequences, they differ in many morphological, behavioral, and cognitive aspects. The underlying genetic basis of many of these differences may be altered gene expression. The transcriptome in blood leukocytes, liver, and brain of humans, chimpanzees, orangutans, and macaques have been compared using microarrays, as well as protein expression patterns of humans and chimpanzees using two-dimensional gel electrophoresis. Three mouse species that are approximately as related to each other as are humans, chimpanzees, and orangutans have been studied. Species-specific gene expression patterns were identified, indicating that changes in protein and gene expression have been particularly pronounced in the human brain.
* Using the fossil record to estimate the age of the last common ancestor of extant primates. Tavaré, S., Marshall, C. R., Will, O., Soligo, C., & Martin, R. D. (Depts of Biological Sciences, Mathematics, and Preventive Medicine, USC, Los Angeles, CA 90089-1340 [e-mail: [email protected]]). Nature, 2002, 416, 726-729.
. . .
Divergence times estimated from molecular data often considerably predate the earliest known fossil representatives of the groups studied. For the order Primates, molecular data calibrated with various external fossil dates uniformly suggest a mid-Cretaceous divergence from other placental mammals, some 90 million years (Myr) ago, whereas the oldest known fossil primates are from the basal Eocene epoch (54-55 Myr ago). The common ancestor of primates should be earlier than the oldest known fossils, but adequate quantification is needed to interpret possible discrepancies between molecular and paleontological estimates. Here a new statistical method is presented, based on an estimate of species preservation derived from a model of the diversification pattern, that suggests a Cretaceous last common ancestor of primates, approximately 81.5 Myr ago, close to the initial divergence time inferred from molecular data. It also suggests that no more than 7% of all primate species that have ever existed are known from fossils. The approach unites all the available paleontological methods of timing evolutionary events: the fossil record, extant species, and clade diversification models.
* A radiation hybrid mapping panel for the rhesus macaque. Murphy, W. J., Page, J. E., Smith, C., Jr., Desrosiers, R. C., & O�Brien, S. J. (Laboratory of Genomic Diversity, Bldg. 560, Rm. 11-10, Frederick, MD 21702 [e-mail: [email protected]]). Journal of Heredity, 2001, 92, 516-519.
. . .
The genomes of nonhuman primates have recently become highly visible candidates for full genome analysis, as they provide powerful models of human disease and a better understanding of the evolution of the human genome. A 5000 rad radiation hybrid (RH) mapping panel for the rhesus macaque was created. Duplicate genotypes of 84 microsatellite and coding gene sequence tagged sites from six macaque chromosomes produced an estimated whole genome retention frequency of 0.33. To test the mapping ability of the panel, RH maps for macaque chromosomes 7 and 9 were constructed and compared to orthologous locus orders in existing human and baboon maps derived from different methodologies. Concordant marker order between all three species maps suggests that the current panel represents a powerful mapping resource for generating high-density comparative maps of the rhesus macaque and other species genomes.
* Excavation of a chimpanzee stone tool site in the African rainforest. Mercader, J., Panger, M., & Boesch, C. (Dept of Anthropology, George Washington Univ., 2110 G Street NW, Washington, DC 20052 [e-mail: [email protected]]). Science, 2002, 296, 1452-1455.
. . .
Chimpanzees from the Ta� forest of C�te d�Ivoire produce unintentional flaked stone assemblages at nut-cracking sites, leaving behind a record of tool use and plant consumption that is recoverable with archeological methods. About 40 kilograms of nutshell and 4 kilograms of stone were excavated at the Panda 100 site. The data unearthed show that chimpanzees transported stones from outcrops and soils to focal points, where they used them as hammers to process foodstuff. The repeated use of activity areas led to refuse accumulation and site formation. The implications of these data for the interpretation of the earliest hominin archeological record are explored.
Instruments and Techniques
* Scan and focal sampling: Reliability in the laterality for maternal cradling and infant nipple preferences in olive baboons, Papio anubis. Damerose, E., & Hopkins, W. D. (Centre PsyCL&EACUTE;, Univ. de Provence, 29 Ave Schuman, 13621 Aix-en-Provence Cedex 1, France [email: [email protected]]). Animal Behaviour, 2002, 63, 511-518.
. . .
A comparison of measures of laterality obtained by two observational sampling procedures (a 15-min focal-dyad sampling with continuous recording, and a scan and instantaneous sampling), using 10 mother-infant dyads of captive olive baboons. The two measures of lateral biases for maternal cradling, infant nipple preference, infant head position and maternal carrying, but not those for infant retrieval, were positively and significantly correlated. The results clearly show that the two sampling procedures produce equally sensitive measures of lateral bias for both the maternal and the infant behaviors. They also provide evidence of asymmetries in mean bout length and therefore suggest that recording bouts is not necessarily the best measure of lateral bias. Taken together, these results show empirically that the scan and instantaneous sampling procedure does not lead to a lack of independence of data points, as previously assumed.
Reproduction
* Sperm competition: Motility and the midpiece in primates. Anderson, M. J., & Dixson, A. F. (San Diego Zoo, P.O. Box 120551, San Diego, CA 92112-0551 [e-mail: [email protected]]. Nature, 2002, 416, 496.
. . .
In animals with multiple-partner mating systems, the gametes of two or more males must compete to fertilize a given set of ova. Here it is shown that the volume of the midpiece in individual sperm is significantly greater in primate species in which the females mate with multiple partners, and in which males have larger testes in relation to their body weight, than in those species that mate with only one partner and have relatively small testes. Results indicate that sexual selection by sperm competition has influenced the evolution of a specific component of male-gamete morphology, the volume of the sperm midpiece.
* Life in the slow lane? Demography and life histories of male and female sifaka (Propithecus verreauxi verreauxi). Richard, A. F., Dewar, R. E., Schwartz, M., & Ratsirarson, J. (Dept of Anthropology, Yale Univ., P.O. Box 208277, New Haven, CT 056520). Journal of Zoology, 2002, 256, 421-436.
. . .
A 16-year study of wild, unprovisioned sifaka at Beza Mahafaly in SW Madagascar provides estimates of age-specific fertility, mortality, and dispersal in a population of 426 marked animals, and longitudinal records of individual life histories. Sifaka females give birth for the first time later and live longer, for their size, than mammals in other orders; they also give birth later and continue reproducing longer, for their size, than other primates. Theory postulates that these features, commonly referred to as bet-hedging, evolve in unpredictable environments in association with widely varying infant survival and a trade-off between reproductive effort and adult survival. The climate of SW Madagascar is highly unpredictable compared to almost all other regions in the tropics with similar average rainfall, and it is argued that sifaka females are bet-hedgers par excellence. Male sifaka, in contrast, become reproductively active at an earlier age than females, and are less likely to have long lives than females. The atypical direction of this asymmetry between males and females reflects a �slowing down� of female life histories rather than a �speeding up� of male ones. Two other unusual features of sifaka biology and behavior may be linked to the unpredictability of the climate: intense local competition between females, and a sex ratio at birth strongly biased in favor of males in most years. In drought years, reproductive females must cope with suddenly intensified resource constraints. This, in turn, may strongly limit the number of �breeding slots� available over the long-term for females.
* Sperm quality differences between the rainy and dry seasons in captive black-handed spider monkeys (Ateles geoffroyi). Hernández-López, L., Cerezo Parra, G., Cerda-Molina, A. L., Pérez-Bolaños, S. C., Díaz Sánchez, V., & Mondragón-Ceballos, R. (Inst. Nac. de Psiquiatría Ramón de la Fuente, Calzada México-Xochimilco 101, Col. San Lorenzo Huipulco, Tlalpan 14370, México, D.F., México [e-mail: [email protected]]). American Journal of Primatology, 2002, 57, 35-41.
. . .
An assessment of sperm measures (concentration, motility, viability, etc.) of three black-handed spider monkeys (Ateles geoffroyi) during the rainy and dry seasons in Mexico City, as well as an evaluation of the between-subjects variability of sperm quality. Twenty samples obtained by rectal electroejaculation and digested with trypsin were evaluated. The results showed that during the dry season (n = 9) the semen samples were of better quality than those obtained during the rainy season. The individual animals showed differences in sperm concentration, although there were no differences in sperm quality.
* Changes of urinary steroid conjugates and gonadotropin excretion in the menstrual cycle and pregnancy in the Yunnan snub-nosed monkey (Rhinopithecus bieti). He, Y., Pei, Y., Zou, R., & Ji, W. (W. J., Kunming Inst. of Zoology, Chinese Acad. of Sci., 32 Jiaochang Donglu, Kunming, Yunnan 650223, China [email: [email protected]]). American Journal of Primatology, 2001, 55, 223-232.
. . .
The Yunnan snub-nosed monkey, one of the most endangered species in the world, is endemic to China. The authors know of no information on reproduction for this species. The present study was designed to understand the characteristics of reproductive hormone secretion during the menstrual cycle and pregnancy of this species by monitoring urinary estrone conjugate, pregnanediol-3-glucuronide, bioactive follicle-stimulating hormone (FSH), and luteinizing hormone (LH). Urine samples were collected each day from four adult females for eight menstrual cycles, and once in 3 days during pregnancy (three full-term pregnancies, one mid-term abortion). The steroid conjugate was tested by radioimmunoassays, and bioactive FSH and LH levels were measured in vitro by the sensitive bioassays granulosa cell aromatize bioassay and rat interstitial cell testosterone, respectively. The results suggested that the pattern of reproductive hormones for R. bieti is similar to that of other Old World monkeys, but the concentrations of hormones is different from that of other species. This species has a longer progestation period, which may be related to its classification status.
* Relationship between platelet-activating factor concentration in rhesus monkey (Macaca mulatta) spermatozoa and sperm motility. Roudebush, W. E., Gerald, M. S., Cano, J. A., Lussier, I. D., Westergaard, G., & Higley, J. D. (Reproductive Biology Associates, 1150 Lake Hearn Dr., Suite 400, Atlanta, GA 30342 [e-mail: [email protected]]). American Journal of Primatology, 2002, 56, 1-7.
. . .
Platelet-activating factor (PAF) is a potent signaling phospholipid that has been implicated in a number of biological activities. PAF concentration in primate spermatozoa has a positive correlation with fertility. While PAF is present in rhesus spermatozoa, there are no relational reports on its concentration and the cells motility. The study objective was to determine if PAF concentration in rhesus spermatozoa was correlated with motility indices (percent motility and forward progression). Semen was collected from sexually mature males and cell counts, and percent motilities and forward progressions were recorded prior to PAF measurement by radioimmunoassay. The data demonstrate that PAF concentration in rhesus spermatozoa has a significant relationship with percent motility and the cells forward progression. Determining PAF concentration in spermatozoa may be a significant predictor of fertility in the primate. Additional studies will elucidate the role of PAF in spermatozoa function and the significance PAF plays in primate fertility.
* * *
----------
We would like to acknowledge Primate-Science as a source for information about new books.
----------
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; FAX: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
Current and back issues of the Newsletter are available on the World Wide Web at
http://www.brown.edu/primate
The Newsletter is supported by U. S. Public Health Service Grant RR-00419 from the Comparative Medicine Program, National Center for Research Resources, N.I.H.
Cover illustration of a gibbon (Hylobates lar) by Anne M. Richardson
Copyright (c) 2002 by Brown University
Assistant Editor: Elva Mathiesen
Last updated: June 19, 2002
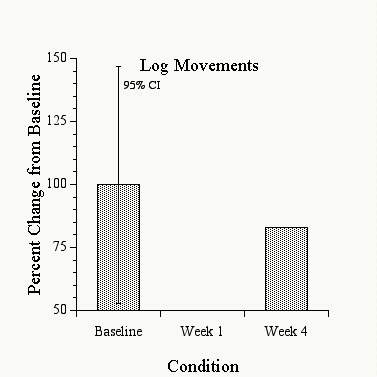{kind=link}
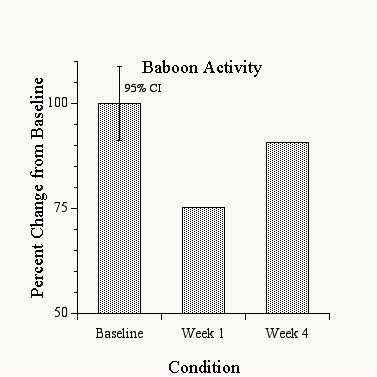{kind=link}
{kind=link}
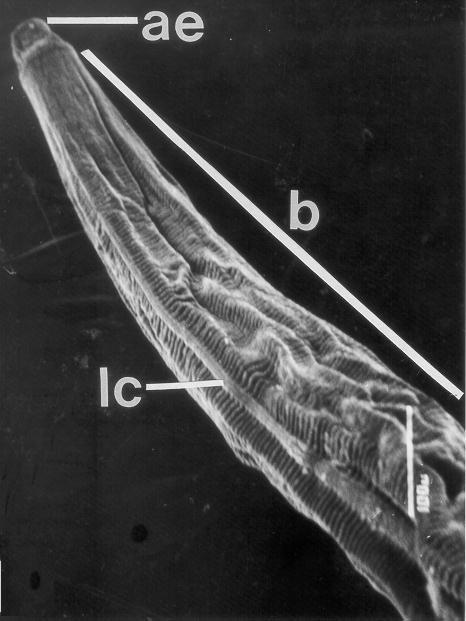{kind=link}
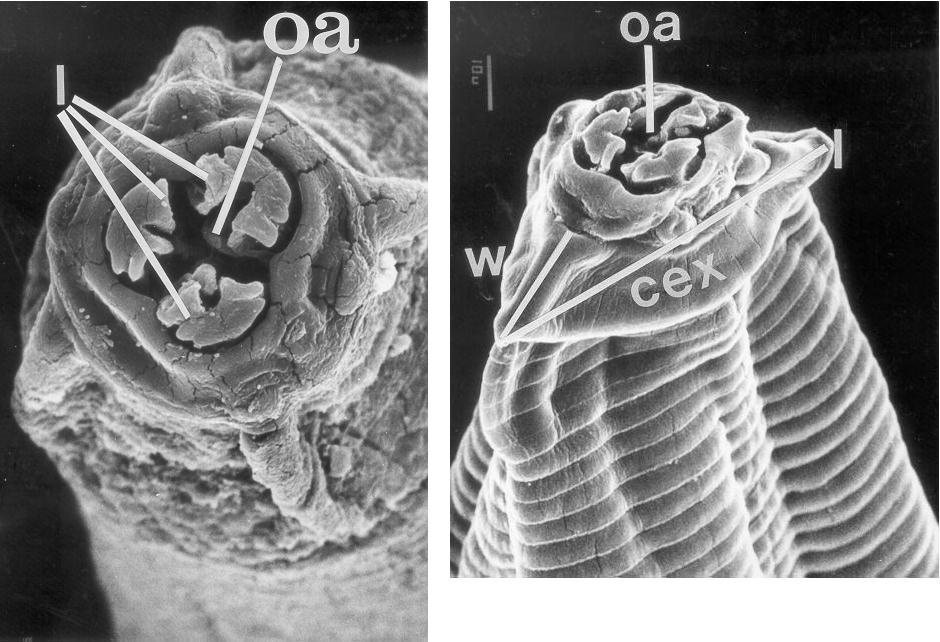{kind=link}
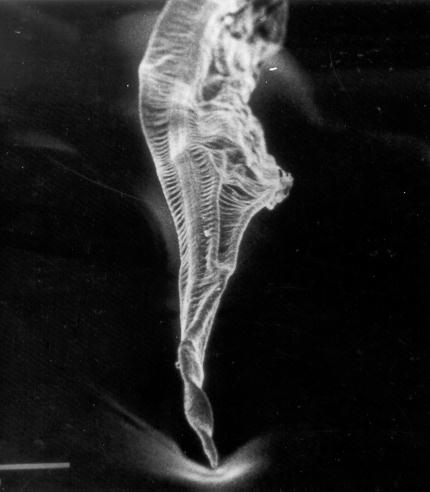{kind=link}