Redraw this circuit as:
and apply knowledge about dynamic transfer functions and feedback:
Some of the account here, of time constants and velocity storage, depends on knowing a formula for feedback that will be developed in a separate lecture.
Lengthening the VOR time constant
The long time constant (for a primate) of the semicircular canal (SCC) is about 6 seconds. After 6 seconds of continuous constant-speed angular rotation the hair cells of the horizontal canal will be only 1/e of their maximum deflection. If a correct reflex is to be maintained, what is the eye movement control system to do? First of all it tries valiantly to lengthen the time constant, and manages, by positive feedback, to do so by a factor of 3-4. The time it takes for vestibular nystagmus to decay is much longer than would be predicted from the long time constant found on the VIII nerves leading into the CNS: about a factor of 4 longer!
In the "velocity storage" loop
Toptokinetic = Tcanal = 5 seconds;
TVOR = TOKAN = 20 seconds. OKAN = optokinetic after-nystagmus.
This difference, that Tcanal is smaller than TVOR, doesn't
seem to be much noted in RHSC2; see page 337ff. But on page 33 is a graphic
of 5 seconds vs 20 seconds!
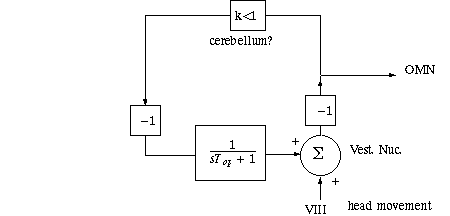
The diagram below has VIII as an input to vestibular nucleus, Vestib Nuc, a summation point. As part of the work of the Vestib Nuc, the sign of response is changed (-1) so the eyes move the opposite direction as the head.
The Vestib Nuc also projects to a "neural integrator"
loop, possibly in the cerebellum. On that feedback path an attenuation takes
place; Say k=0.75. On a pathway which we will shortly see has to do with optokinesis,
the sign is changed again (positive feedback now) and a low pass filter is encountered.
We tell you here that the time constant of the LP filter matches the low frequency
time constant of the canal.
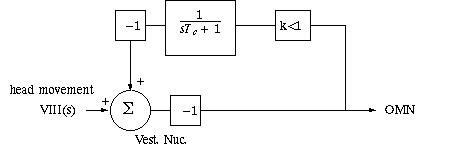
Redraw this circuit as:
and apply knowledge about dynamic transfer functions and feedback:
Expressed in the multiplicative form of Laplace transforms,
the output Vestibular Nucleus is
Vestib_Nuc_out(s) = T(s)*VIII(s). The transfer function T(s) is embedded in
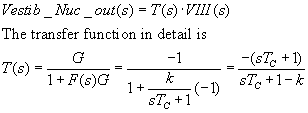
Now let's insert VIII(s), as calculated below:
A simplified formula for VIII(s), involving only the long
time constant, comes about by,
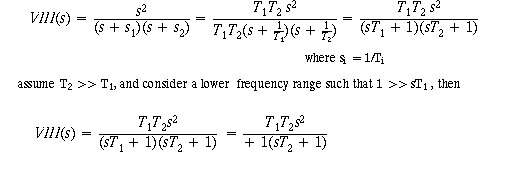
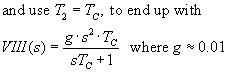
now multiply VIII(s) by T(s) to find the Vestib Nuc output = OMN input (assume
Tc = Top)
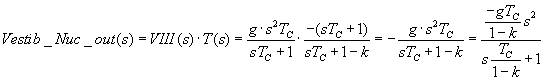
and if k = .67 or .75, we've increased the time constant by 3 or 4. And that's
what's recorded in the Vestib Nuc, and in eye movements: a 20 second time constant,
when the VIII is decaying after 5-6 seconds in primates. (During sleep or drowsiness,
some results indicate the VOR tau can decrease.)
At any rate, we have established the presence of a neural integator in the expression of the vestibuloocular reflex. This low-gain positive feedback loop is known to physiologists as the velocity storage loop.
Going into the OMN, the VOR system still presents the results a velocity-sensitive accelerometer, that needs to see another integration to be turned into eye position.
OPTOKINESIS & Optokinetic nystagmus
Optokinesis is the automatic response of the eyes to the world (a large
"target") moving around the observer. Optokinetic nystagmus
is the response to the world moving continuously around the observer "too
far" for a smooth eye movement to track the world. OKN includes a slow
tracking phase and a quick phase, as with vestibular nystagmus.
See first part of chapter 3, of RHSC2. In Fig. 3.4(a) we see how long it takes OK response to develop in rabbits, and how rapidly it develops in humans. And see the nystagmus after lights out...
OKN influenced by "attitude"...Look vs Stare: See Fig 3.1 and study Fig. 3.3. Why are the open circles (look) below the closed circles (stare)? good up to 30-60�/sec; See fig. 3.4 for 20 sec of OKN in rabbit.
Notes: 1996 episode of ER: rotating a vertically stripped drum in front
of a not-so-"blind" patient!"
Ballerinas spot, while ice skaters spin too fast for spotting, and are told
NOT to. Spotting means fix the head on ONE landmark in the distance, and crank
the head around for the next view, as the turn progresses.
Optokinetic and vestibular responses together
Notice that a person rotated in a chair, in the light, will, in addition
to having a vestibular experience, will see an OK stimulus . Just considering
OK alone, the eyes will try to keep the world stable on the retina and counter-rotate
to cancel the head movement. Therefore OK and VOR generate the same eye movement
for the rotating person. Do the two systems cooperate or compete? Seemingly,
they cooperate, with vestibular starting fast, and optokinetic being sustained.
In the cerebellum: p. 252 flocculus of cerebellum, properties of (Purkinje)
cells: "...showing most interest in large contrast-rich patterns moving
in particular directions."
"Floccular mossy fibers are mostly vestibularly driven, the majority of these
with visual input as well, and very few visual only." p.336ff ...in both cases
there is a strong sense of self-rotation in the absence of any signals from the
canal...
Example of the world moving around the observer? being in the middle of a tornado
funnel? See movie Twister?
If a person experiences OKN for many seconds, optokinetic-after nystagmus OKAN occurs as soon as the lights are turned out. What if the stimulus stops and the lights stay on? It is possible to suppress OKAN by fixation...easier than suppressing VAN, vestibular after nystagmus.
Further development of the velocity storage model
The time constant of optokinetic after nystagmus (OKAN, measured in
the dark) is created by the positive feedback loop with k<1. Without this
loop, there would be virtually no after nystagmus when the lights are turned
out. See diagram below, with switch OPEN in the dark. Notice also why the loop
is called a velocity storage loop!
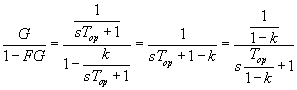
OKAN (in the dark) typically, k = 0.75 so 1/(1-k) = 4
The rise time of optokinesis (measured in the light, with
the world turning) can be less than the fall time of OKAN. See system
diagram below. By "movement" at 3 locations we means angular velocity:
world, head and eye rotation velocity. The retina is motion-sensitive and has
the equivalent of a differentiator or first-order HP filter.
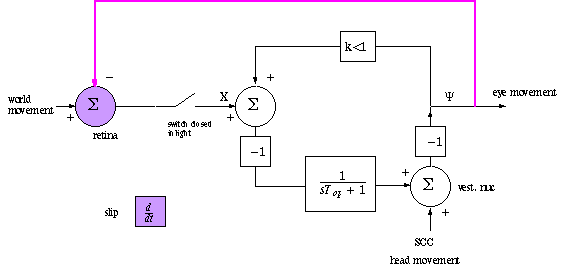
The outer negative feedback loop, in bold above, speeds up the response of the
system. The system as shown says that the retinal output compares world position
to eye position when in fact the slip detectors are zero when the velocities
of the world and the eye, in deg/sec, are equal. In SIMULINK you can examine
the differences in the two assumptions.
What is the transfer function of the system from world to eye movement, with head movement = 0 and k = 0.75, and the switch closed? Basically, the negative feedback of the outer loop will speed up the system when the switch is closed (start of OKN).The switch will be open when OKAN is measured.
What is the time constant associated with this overall transfer
function? Remember that "s" is the Laplace transform variable, and
represents frequency or derivation; thus T0�s is a dimensionless factor.

for the OK transfer function. The loop needs more gain, which we can apply at
the level of the retinal HP filter.
This derivation demonstrates that the outer negative feedback can alter the
system to create a smaller time constant and a faster response. However, when
the lights are turned out after optokinetic stimulation, then the opto loop
is opened and the time constant of OKAN in the dark reverts to TAU VOR, about
20 seconds.
Opening the loop
How can we open the loop on the feedback system, experimentally, and discover
what the forward pathway does? See Fig. 3.7 for increasing velocity. Note the
slow speed involved!
6 POSSIBLE ways
1. cut the internal pathway from slip to motor control-impossible
2. use after-image of a bright light just off fovea...huh? isn't this position error?
3. record from yoked eye while viewing eye is paralyzed, or muscle cut: rabbits!
...see Vis Res paper on one human subject
4. add eye movement signal electronically to moving image on CRT
5. stabilize image with optical feedback: Brown Univ Psyc Dept: Lorin Riggs
work
6. the first 100 msec on movement + delay - Lisberger
8. weaken eye muscle, record from OMN
Vis Res 22: 1197: Leigh et al. (in folder) ONE subject,
died two weeks later...
more complex than Ter Braak, 1936 results...
Q: what does fig. 3.9, RHSC1 imply? (part is on ps#7)
RHSC2: slip may try to choose models for best tracking, makes open-loop hard to
interpret. At least open loop gain seems "high" according to Leigh &
Zee
Does DELAY cause INSTABILITY? SEE FIG 3.25 IN RHSC2. When the loop is opened we see a neural integrator inside... match the plant?
Leaky integrator (see notes in separate file)
The figures in RNSC2 about 3.9 show a velocity that saturates, implying
the integrator can better be modelled by a leaky integrator, and we can use
the same e-dot = IN - K*e form to show a feedback system for leaking. Working
around Fig 3.10, 11...
Optokinesis vs smooth pursuit
RHSC2 p.39 Large vs small targets. "...optokinesis is
seen in all animals with moving eyes, smooth pursuit is only demonstrable in
animals with a small central retina." "The two mechanisms [SP and
OK] are at odds with each other." WHY?
See edited Ellen Barton ScM thesis, (1989)
Optokinesis complements VOR
There's a difference between the world not moving, and darkness...
QUESTION:
What happens when the head rotates while the eyes observe a textured background?
The VOR tends to rotate the eye opposite to head rotation.The background drifts opposite to head rotation, so optokinetic slow phase moves in agreement with vestibular, against head rotation. At the beginning, the vestibular response rise is rapid; the OK response rises slowly. After some seconds of rotation, the vestibular influence dies down, but the OK response builds up and continues.
Frequency response: When both OK and Vest. input are active, why don't we get a gain greater than 1?
DEMO INNISFREE image synthesizer, TO SHOW REALITY OF
OPTOKINETIC STIMULUS (sinewave grating)
Kathy E. Cullen (ex-student, now on faculty at McGill) papers
D. Guitton, "Control of eye-head coordination during orienting gaze
shifts," TINS May 1992, page 174. KEC mentor at McGill. Fig. 1b
a great example of head and eye rotation in the same direction for
a large gaze shift! VOR defeated in this case!
Kathy Cullen, T Belton & R.A. McCrea, "A non-visual mechanism for voluntary cancellation of the vestibulo-ocular reflex," Exp Brain Res 83: 237-252 (1991)
more recent J. Neurophysiol papers: 1998-99.
When should VOR be cancelled?
Why "non-visual" cancellation?
Paradigms: Monkey on a turntable viewing a head-centered moving target
paradigm 2: head acceleration steps
From KEC + McCrea, "Firing behavior of brain stem neurons
during voluntary cancellation of the horizontal vestibuloocular reflex I: Secondary
vestibular neurons." J. Neurophysiol. 70: 828 (1993)
INTRODUCTION: " Primates are capable of utilizing a variety of strategies
for tracking a moving visual target. An ocular saccade is usually made to bring
a moving visual target onto the fovea. Subsequent movements of the target with
respect to the fovea are tracked by generating additional saccades, by generating
a smooth pursuit eye movement, or by combining smooth eye and head movements,
i.e. gaze pursuit. For a smooth head movement to contribute to visual
tracking, the eye movements generated by the vestibuloocular reflex must be
suppressed or canceled in some way, because the VOR is designed to stabilize
gaze in space by generating an eye movement that is in the opposite direction
to an ongoing head movement."
KEC 1991 paper-- "The latency of VOR suppression
was significantly shorter than the latency of smooth prusuit while the monkey
was cancelling its VOR." What happens before smooth pursuit kicks in is
the "non-visual" part.
VOR capable of stabilizing gaze over a remarkable range of frequencies (0.05-14
Hz) and velocities (up to 350�/sec) of head rotation. VOR not needed during
large GAZE saccades. (head movement involved) or during combined head-eye
tracking
RESULTS: Fig 1a. While monkey fixates a target on the turntable, the turntable
rotates at a given velocity for 1 sec and eye movements are recorded. Below
60�/sec cancellation is complete. Fig. 1b. Smooth pursuit is good up to about
60�/sec. Fig 1c. Smooth pursuit aided by head rotation is goes up to 120�/sec.
"The results ... indicate squirrel monkeys were better able to fixate a
head stationary target during high velocity vestibular stimulation than they
were able to pursue targets that were moving at similar speeds." which
means?
Fig. 2. Latency of VOR is 100 msec? No. poorly presented figure. See H' trace
for actual stimulus. Almost no latency.
Fig. 3a. After 90 msec fixation could suppress VOR. Fig 3b. SP alone. What about
a saccade plus smooth pursuit?
Fig. 4. Unexpected change in head acceleration
KEC Fig 11:
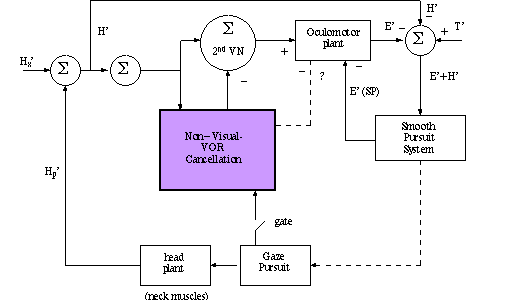
T' = target velocity; E' = eye velocity; E'(SP) = central smooth pursuit signal;
H' = head velocity; Hx = externally induced head velocity; Hp = head pursuit
velocity

--------------------------------- ------------------------------------
OTHER notes:
Discuss gaze, as a combination of head and eye movement.
The value of EB thesis lies not so much in the results, which are mostly negative, but in the exposition of a stable positive feedback model which explains lengthened time constants.
Need dynamics in the gain-less-than-one positive feedback velocity storage loop.
Should
say that s2H(s) is the acceleration term, so sH(s) is the velocity term.
How does the vestibular constant get lengthened by itself? It doesn't! It needs
to have dynamics in its positive feedback loop! Might as well bring in TOP before
the math...like Ellen did.
1990:
Georgopoulos at Brown, April 12: "Intentionality & Motor Control"
rough draft of term paper due
April 12, 1994: David Van Essen, "Mapping and modeling of the primate visual system."