Adjustment of VOR gain
You now know about interactions between the vestibular and the
visual (optokinetic) senses. Rotating around on a stool, with your eyes open, then
suddenly stopping, is the demo to keep in mind. Imagine such rotation while wearing
back-facing glasses, or binoculars. It will turn that with less than an hour of
such experience the gain of your VOR can change in a direction to compensation the
visual disturbance.
Slip detectors. The visual system, at the level
of retinal ganglion cells, activates slip detectors if the world image
is not stable on the retina. Slip represents an error that is to be corrected by
adaptive gain control in the VOR circuit. Slip is a vector, that has magnitude and
direction. Slip implies that not just a foveal-size target is moving, but the whole
patterned world. A large, vs a small, stimulus. Slip is reporting the rate of change
of position of the world: a high-pass filter.
OUTLINE
* Discuss motor adjustment in general, via parametric control
**neural network weight adjustment RHSC2, chpt 13.
**proprioception, in skeletal muscle
* Produce by optical means a mismatch between visual and vestibular error = retinal
slip (velocity detectors active)
* VOR after damage to ONE vestibular organ. 1 month of nystagmus
* Organization of the cerebellum for VOR gain control. flocculus.
* Robinson, '76: ablation of cat vestibulo-cerebellum after gain change.
* Lisberger theory for adaptive gain control
Gain of the VOR: Demonstration, measurement and modification
How do you measure gain of the VOR? in the dark as the VOR amplitude divided
by sinusoidal head rotation input what other output feature is there, besides gain?
Phase! How does gain depend on frequency?
DEMO WITH BINOCULARS & rear-view mirror.
with telescoping lenses or mirror to alter viewing, nothing is wrong with eye movement
control system (saccades and smooth pursuit are OK) until the head starts to move.
Viewing through binoculars while you rotate your head back and forth a few degrees
...Notice that MR isn't pulling enough...movement to the left for rotation to the
right means that head movement is "too much" for MR reaction...
with rear-view mirror demonstrate binocular rivalry, then close the unmirrored
eye! MIRROR deflection to defeat VOR: Why does it work? Doesn't the mirror just
turn you into a sort of side-viewing rabbit? And doesn't horizontal rotation of
a rabbit elicit a standard VOR? Hint: writing is reversed in a mirror...what about
up/down?! try it with two mirrors...
How long does the adjustment take? Hours? and days? or see RHSC2- try 15 minutes!
Fig 13.5, page 383!
VOR after damage to VIII nerve(s)
After unilateral damage to vestibular organs, nystagmus and turning disappear
in a month. Compensation by the remaining vestibular organ occurs over several months.
Balance and gain. See Fig. 13.3, and gain reduction of half.
See Adaptive Mechanisms in Gaze Control, page 251. Precht.
balance and habituation of VOR.Q: What does he mean by habituation? Decline in number
of nystagmus beats after (hours, days) of repeated rotation in the dark.
Arousal strongly affects vestibular nystagmus. Change of VOR--the long
time constant disappears during anesthesia...
Bottom line: after unilateral damage to vestibular system, the CNS can adjust
from push pull to just "push."
RHSC2 chpt 13
Adjustment of ballistic eye movements, like VOR. The controller must be the
inverse system of the plant. In the face of a S plant.
Need parametric feedback. Adaptive filter. Neural network. See Fig 13.1(a). Compare
desired with actual and use to adjust gain. What about the
sign at the comparator? IN-OUT is positive when goal is larger than output, and
therefore weight must increase.
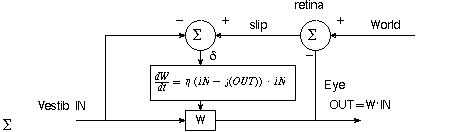
The effect is like rule learning in a neural network! If there's no vestibular change
(IN), no synaptic change should occur. Figure 13.2, examples of "parametric
feedback" in the oculomotor system.
RHSC2 suggests "stretch receptors" may monitor performance. Is it true?
a role for proprioception? Mention stretch reflex. [Habituation vs fatigue] Page
377 of RHSC2: fatigue. See page 181, & Fuchs & Binder.
Bottom of page 378: Transposition of medial and lateral rectus, and superior and
inferior rectus, in monkey, is compensated for in 8 days! Like the inverting prism
artist in the video (The Brain #2, at Athenaeum).
Even when the animal has stayed in the dark?! (Suggests a role for proprioception...)
terminology: Gaze-holding: Vestibular vs optokinetic: VOR is more "adjustable"
because it's ballistic. Optokinetic gaze holding can use immediate visual feedback.
Visual adjustment of VOR
In the light, a gain error of VOR is attacked by optokinetic following. DEMO on
LAB stool: rotation within a subjectively stationary visual field-follow a finger.
Decrease in vestibular nystagmus in ballet dancers! bottom of page 382. In man adjustments
to gain-changing spectacles are virtually complete in 30 minutes.
The extreme case: wear reversing prisms. Gain can be reversed in a few days. (Fig
13.6)
Optokinesis and strobe light
The adaptation of VOR can be frequency-selective (see Fig. 13.7). Does optokinetic
nystagmus gain change with VOR? The 2 are coupled through the velocity storage system...On
the other hand, can VOR gain change after a period of purely optokinetic stimulation?
Can VOR change during strobe illumination, when perception of motion is defeated?
(more later in perception of motion lecture). Frequency channels: Fig. 13.7. Strobe
viewing and slip. p. 386.
Development: Is optokinesis learned or inate? Appears to be inborn. Some
seen immediately at birth, but not with gain of adult. What about strobe rearing?
OKN gain is reduced.
From prior lecture: VOR and OK following both share the same velocity storage
loop. If adjustments of VOR take place in Vestib Nuc, then change in VOR may change
OK following.
VOR gain is cut in half by destruction of one labyrinth RHSC2 Fig 13.3. compensation
occurs over weeks. Balance and gain changed by elimination of one input for a push-pull
amplifier. Arousal can affect VOR gain. Habituation vs real gain change... Can VOR
be altered in strobe illumination? not as much...
looking ahead: Cerebellar poisoning: �13.5: "...many cerebellar
afflictions are remarkably similar to the effects of alcohol poisoning." Fig.
13.16 looks a lot like the first figure in Robinson 1976 reading from Nauta...Fibers
from the most medial of the deep cerebellar nuclei [fastigal] also leave the cerebellum.
They terminate largely in the vestibular nuclei.
RHSC2-Neural mechanisms
adjustments are NOT made at the level of the vestibular organ itself. Recordings
from VIII show responses unchanged before and after. "Functionally, there seems
little doubt that adjustments in gaze-holding are the result of alteration in the
gain of the velocity storage mechanism, and that whether or not the cerebellum itself
forms part of this mechanism, it is responsible for controlling it."
SEE IT ON THE SIMULINK DEMO SLvest.mdl model.
In general: Does the cerebellum do correlations, and discover "context"
for learning? Are synaptic connections from parallel fibers weakened during
learning because of the cerebellum's inhibition of Deep Cerebellar Nuclei? See Fig.
13.16 for cerebellar model, like Robinson '76. Input a is small when the reflex
is working. a is an error signal in the drawing. 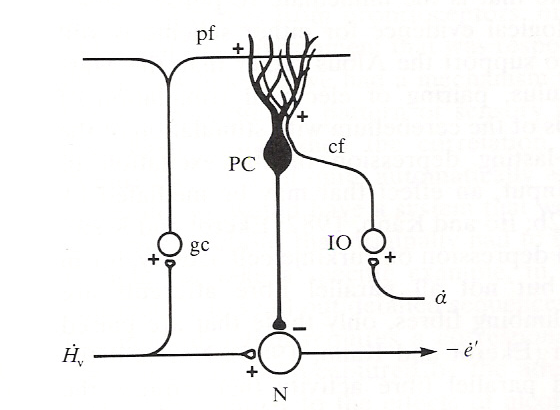
where N is a Vestib Nuc neuron, gc is cerebellar granule cell, IO stands for inferior
olive, cf for climbing fiber, pf for parallel fiber and PC for Purkinje cell (see
subsequent notes on cerebellum for details).
...from which we can form
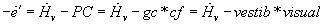
which implies that both vestibular and visual input must be nonzero for change of
gain.
RHSC2 says, "Under conditions of visual suppression, however,
[a-dot] will be greatly increased and will cause and immediate reduction in eye
velocity because of inhibition by the Purkinje cell." The term visual suppression
here may mean "visual disturbance due to optical interference" and not
just darkness.
Compare to Fig 1, Robinson '76, from which we hear later in his
paper:
"In the case of VOR gain control, bilateral destruction of the flocculus has
repeatedly been shown to prevent the occurrence of adjustment in response to reversing
prisms and magnification changes."
Q: What if flocculus ablated immediately after learning of VOR
adjustment? and see more citations from RHSC2, page 397. ...destruction of neonate
cerebellum.
Notes on cerebellum and its vestibular input ( and see other
webpage devoted to the cerebellum).
From Nauta & Fiertag, page 101-02: "The vestibular nuclei project in turn
to the cerebellum. There the projection distributes its fibers to part of the vermis
[the worm], the midline district of the cerebellum, and also the flocculonodular
lobe, the caudalmost part of the cerebellum, which consists of the caudalmost part
of the vermis, called the nodulus, and a slender wing that ends in a slight
swelling called the flocculus."
Nauta & F: page 101ff, the fibers from the fastigal nucleus
leave the cerebellum and terminate in the vestibular nuclei. "Some of the fibers
leaving the cortex of the flocculonodular lobe bypass the deep cerebellar nuclei
and project to the vestibular nuclei."
What about recordings from Purkinje cells during conditioning?
They fire more briskly during sensorimotor mismatch, but at this point, on to Lisberger
results...
REF: John Nolte, The Human Brain, 3rd Edition (1993)
pages 337-358 Nolte book: page 346 etc, flocculonodular region receives vestibular
input.
Cerebellar Purkinje cells in the flocculus respond to gaze
shift = head + eye movement. In VOR, headdot = -eyedot. Normally the PURKINJE
CELL response is zero, so how can it be site of VOR plasticity? They show corollary
discharge as input to cerebellum. Corollary discharge may be influenced by
pursuit movements which are trying to compensate for VOR mismatch. Where
does aot come in? they don't say.
David
A. Robinson, "Adaptive
gain control of vestibuloocular reflex by the cerebellum,'' J. Neurophysiol.
39: 954-969 (1976). [click
on highlighted link above and download PDF version of the paper]
Here we study a research paper, wondering
critically if his conclusions are justified by the results. As a first slice,
we should look at the figures and try to understand them. In general, when reading
a biologically-inspired research paper, you hope the author recapitulates the
results in the first paragraph of "Discussion." Probably not so in
this paper: Robinson seems to jump right into qualifications about the experiment.
1. Motivation: Demonstrate that the the
VOR gain of adult cats can be modified, then test the hypothesis that removal
of all or part of the cerebellum prevents the expected modification (plasticity),
or returns an altered system to the unmodified state. Note use of adults, not more
"plastic" kittens. The hypothesis is cast more precisely in the form of
a model:
2. Model * Study the Intro: Robinson's Fig.
1 model with gain = a - b; note the output/input is in terms of velocity! SLIP is
a deg/sec variable. And remember that the SCC transfer function for frequencies
in the range of 1 rad/sec acts like velocity transducer (since pole high pass filter).
The b pathway is feedforward inhibition and subtracts from the a path at the Vest.
Nuc. [from an engineering POV an example of automatic gain control]
The a.o.t. visual input, representing SLIP,
can alter the beta signal. Beta can be affected if the head is rotating and slip
is nonzero. If head velocity is clockwise and slip is clockwise, should the gain
increase or decrease? Another question: if the eyes don't move when the head rotates
CW, what direction is slip? ANS: It is CCW, if the world rotates CCW. In such a
case, the gain needs to increase. So in the case of both H and slip in the same
direction, the gain needs to decrease. These gains changes suggest what the a.o.t.
must do to change gain beta. At any rate, what does the model predict would be the
effect on VOR gain if the cerebellum were removed? Will the VOR gain go up or down?
In SIMULINK you can represent the gain change by a product, with one input to the
product being from an integrator that is (with the correct sign) integrating slip
gated by h(t).
3. Methods: How to measure VOR: in the dark!
See other lecture notes on methods of measuring eye movements...
Use search coil technique, implant coil
ahead of time, and a bolt in the skull for stabilizing the head on the turntable.
Gain of VOR in the dark about 0.9 < 1.0. In some cases (Fig 2A) the animal was
rotated at constant angular velocity and the slow phase of vestibular nystagmus
measured. Look at frequency dependence of the gain. Look at Fig. 2, and notice the
nystagmus. Was the input rotation or sinusoidal back and forth?
4. Cerebellectomy: recovery from and verification
of: How long after surgery can the test for VOR gain be done again? How can the
researcher document that the correct region of the brain was removed? Hint: not
by looking at the removed tissue! [Can you think of any way the experiment could
have been done reversibly, by just inactivating rather than removing the cerebellum?]
5. Results of reversing prism experience: More
gain change at lower frequency stimulation, but slightly higher gain at 1.2 vs 0.05
Hz rotation. Not as much chance for habituation.
*After 8 days / 2 hours per day of wearing left-right reversing prisms, the gain
was reduced by 93% at 0.05 Hz and 55% at 1.2 Hz. Later the gain could change sign!
Notice that, as an engineer, Robinson is going mainly for frequency response, although
in one case he did measure a "ramp response" to continuous rotation. slow
phase of Vestib Nystagmus Fig 2A.
* After removal of "vestibulocerebellum" the gain in the experimental
animals rose, to 1.17! abolishing the effect of VOR "learning."
* Fig. 4: Effects of 4 hours rotation with prisms, then removal of prisms. Why is
the gain greater than 1 for some of the animals?
* Fig 5: Gain at 0.05 Hz = .3 rad/s rotation frequency, over a period of weeks.
Curve C is VOR gain change for "free ranging" cats. On the right is shown
time course of recovery from prism wearing. Are any of the cats in the figure tested
with cerebellectomy? Yes! Curve X at the top of the figure.
* Fig 8: Removal of vestibulo-cerebellum while cat is in adapted state. Two different
frequencies, presumably for the same cat: open circles, higher frequency. On day
15 is surgery, then a day or so of recovery.
6. Cerebellar Lesions In Robinson 1976
CONTROL & Experimental: Fig. 5, recovery after prisms were removed. "After
total cerebellectomy the cats had the usual symptoms of head and forelimb extensor
rigidity, swaying, and oscillations." What is the effect of removal of the
cerebellum on eye movements? "Two animals never recovered above the crawling
stage." see page 962.
[4 CATS] "The gain of VOR always rose after surgery, usually above 1.0. The
gains remained fixed at this value for the remainder of the cat's life (3-9 mo.
) regardless of what was done to it." vestibulocerebellum removed in 7 animals.
Fig. 8. VOR gain of an adapted cat after removal of flocculus and nodulus. Gain
reverts to pre-conditioning levels and doesn't change with continued forced rotation
at 0.05 Hz.
7 Discussion: Where is the site of gain change?
the plastic synapses: in cerebellum or vestib nuc? Does Robinson imply the site
of plasticity is in the cerebellum? Lisberger, below, thinks the site in
the Vestib. Nuc.
Lisberger papers, another POV
S. G. Lisberger & T. J. Sejnowski, "Motor learning
in a recurrent model based on the vestibulo-ocular reflex," Nature
360: 159-161 (1992) And see News & Views by Stuart Judge, page
104
Lisberger Science 242: 728 (1988) - Is the path
through the cerebellum too long to account for short latency changes in VOR? Figure
7 shows a learning rule for the vestibular input onto the FTN. Vestibular
associated with flocculus input...
| vestibular nucleus |
flocculus |
vestib syn |
| above resting |
above resting |
decrease |
| below resting |
below resting |
decrease |
Ann Rev Neurosci. 1981 pp 273-299 "Plasticity in the
VOR: A new hypothesis," Miles & Lisberger.
Lisberger says several days (3) are needed for complete compensation for a 2X binocular
viewing adjustment. days, not hours.
Stuart Judge, News and Views: "There has been a long-standing
dispute about the nature of the brain circuitry that adjusts VOR gain, with Ito
championing the view that the VOR gain is adjusted by regulating the size of the
vestibular signal flowing from the canals through a particular part of the cerebellum
called the flocculus and back to the main pathway in the brainstem."
The S & L model, (in SIMULINK form): The time constants should
be the same for normal control.

Try to duplicate the waveforms of Fig. 2 in their paper.
More papers by Lisberger on VOR and cerebellum:
Lisberger Science Nov 4,
1988, p. 728 Q1/S35, 3rd floor south, SciLi. "Neural basis for
learning simple motor skills."
Lisberger Trends In Neuroscience, April
1988, "The neural basis for motor learning in the vestibulo-ocular reflex
in monkeys."
-------------- ----------------
Natural repetitive VOR: 0.16 Hz, 50� amplitude. 60�/sec for 1 hour. What is it?
Watching a tennis match! No habituation! Of course optokinetic response is also
involved.
Re-reading TINS April 1988 Lisberger. "The VOR undergoes
motor learning whenever errors in the VOR are signaled by the temporal coincidence
of retinal slip and head turns."
Consider retinal slip as detecting a movement (perceived movement
wrt to retina). When head rotates CCW with eyes not moving, the perceived movement
will be CW, opposite direction. This situation is "low gain" VOR, if it
were uncorrected, the VOR gain must increase.
Q: How will the gain need to be changed with binocular viewing?
Try it! the world slips in the opposite direction of head movement! gain too low!
Which way will gain of VOR change if head rotation and retinal
slip are in same direction? [Lisberger says gain should/will decrease. What
is "direction of retinal slip"? opposite of perceived movement]
Lisberger research, contrasted with Robinson '76, tries to find
the site of motor learning by recording from different neurons in the path, instead
of ablating the cerebellum. Figure 2A shows overshoot for low-gain modification.
Phasic VIII afferents are in the unmodified path, and do not diminish in size. Tonic
afferents have 4 ms longer latencies than phasic.
"Because Purkinje cells inhibit their target neurons, the
vestibular pathway through the flocculus is an inhibitory side loop of the brain
stem VOR pathways. Increases in the efficacy of vestibular through the flocculus
would cause decreases in the gain of the VOR." Watanabe recorded increased
vestibular responses from floccular Purkinje cells while monkeys (with lowered VOR
gain) were rotated in the dark, but did decreased eye movement influence in the
cerebellum cause the change? Miles saw small change in wrong direction.
Climbing fibers carry visual information into the cerebellum,
as do mossy fibers. Lisberger thinks the site of motor learning is at the FTN (Floccular
Target Neuron)-which receives monosynaptic input from floccular Purkinje cells and
is therefore "equivalent" to deep cerebellar nuclei cells.
Misc Notes:
Can gain change sign?
Gain in the dark within 5% of -1.00
1981 ref: Over hours gain can increase
to 1.7 (out of target of 2) or decrease to 0.7 (out of target of 0.5). If left in
the dark, or if head is immobilized in light, gain change seems permanent.
If it's tried again, the system takes the same amount of time to "re-calibrate"
... no higher order learning involved
After removal of cerebellum, gain rose (or
fell? see Lisberger 1984 JNP ) anyway
remove before or after prism adaptation...what happens?
CEREBELLAR LESIONS IN ROBINSON 1976:
Sign of mismatch determines the
direction of modification--to strengthen or to weaken
Can the plastic system be tricked by OK
stimulation? seemingly, in the rabbit...
Can lesion of olive stop adjustment of VOR?
yes RHSC2 page 397.
What happens, in general, to eye movements
after removal of cerebellum?
smooth pursuit is greatly affected (Leigh & Zee, p. 24)
RHSC p 209
spontaneous nystagmus
inability to hold fixation
repeated attempt to re-fix object
explain in terms of pulse-step model...
looks like pulse without step
"A loss of gain for slow components of movement"
slow phase of vestibular nystagmus affected
disable a neural integrator, or enable a decay term
Robinson: Cerebellum is the repair shop
of the brain
RHSC2: but it has competitors.
RHSC1 p 18-19: Can't find this
in RHSC2!..
"However, evidence is accumulating that the simple mechanical model discussed
so far cannot account for all dynamic properties of vestibular discharges in response
to angular rotations, at least at extremes of frequency. Reflex eye movements in
response to rotation of the head show an additional component of adaptation over
and above the adaptation component associated with the elastic time constant, and
intrinsic to the mechanism of the canal. The time constant of this component is
around 60-120 sec (compared with the 6 s to 16 s of the elastic time constant),
and so can hardly be said to contribute much to the reflex response under natural
conditions of stimulation. Although these observations by themselves cannot distinguish
between an adaptation process occurring in the occulomotor pathways from one in
the vestibular apparatus itself, Fernandez and Goldberg (1971) find a similar extra
component in their recording from squirrel monkey vestibular fibers, with a time
constant of some 80 s."
Centrally, both the vestibular and visual
systems can adapt or fatigue... 6-16 sec is the Tau of the slow acceleration-detector
component. Continued inertia of the canal fluid is responsible for PRN

~Summary
* Parametric feedback is like weight adjustment in a neural network.
* Vestibular and optokinetic are gaze-holding systems that may need adjustment
* Damage to one vestibular organ reveals some central compensation
* Gain of VOR can, after hours, adjust to cancel new optical arrangement
* Removal of cerebellum eliminates/prevents adjustment
* visual input gets to cerebellum via aot-inferior olive-climbing fibers;
cerebellum projects to brainstem (VN)
* A learning model can explain the gain changes
* recordings from flocculus P-cells suggest that gaze is primary, casts
doubt upon simple cerebellar hypothesis. (gaze is head plus eye movement)
* The site of synaptic plasticity is likely to be at the Floccular Target Neuron.
QUESTION--What will happen to the gain of
the vertical VOR after horizontal head rotation while wearing 2x binoculars? Not
addressed in chpt 13 of RHSC2.
--------------------
Ballistic movements need parametric gain control.
?inverse function for compensation of plant