 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 50 NUMBER 1 JANUARY 2011
Printable (PDF) Version of this issue
Dead Infant Carrying in the Hanuman Langur (Semnopithecus entellus) Around Jodhpur (Rajasthan), by G. Sharma, B. Swami, C. Ram, & L. S. Rajpurohit......1
Rita the Chimpanzee (Pan troglodytes ellioti): The Start of an Enrichment Program at the Doha Zoo, Qatar, by H. Tresz......6
Separating Cage-Mates Temporarily – A LAREF Discussion......10
News, Information, and Announcements
Meeting Announcements......11
Information Requested or Available......11
. . . Great Apes in Japan; New ASP Website; More Interesting Websites
Resources Available......12
. . .Special OLAW Online Seminar; CANTAB Cognitive Testing System; 2011 Bonobo Calendar; Hominoid Carrying Archive Online; Procedures with Care; Lincoln Park Zoo’s Blood Type Database for Apes
Resources Wanted: Videos of the Birthing Process......13
ASLAP Veterinary Student Award Program......13
Research and Educational Opportunities......14
. . .Veterinary Residency, New York; Laboratory Animal Science Management Courses; Wildlife Studies and Primatology – Japan; Education and Training in Field Primatology; Husbandry for Zoo and Aquarium Animals – Mexico
News Briefs......15
. . .Devra G. Kleiman: Conservation Biologist; Will Aging Research Chimpanzees Be Relocated? NY Must Release Records on Monkey Research; AVMA Revises Veterinarian’s Oath; Gorilla Tested for Possible Human Disease; Census: Mountain Gorilla Population Grows
President Obama Honors Nation’s Top Scientists and Innovators......17
Announcements from Publications: The Journal of Evolutionary Biology Research......17
Departments
Recent Books and Articles......18
* * *
Dead Infant Carrying in the Hanuman Langur (Semnopithecus entellus) Around Jodhpur (Rajasthan)
Goutam Sharma, Bharti Swami, Chena Ram, and L. S. Rajpurohit
Animal Behaviour Unit, Department of Zoology, Jai Narain Vyas University, Jodhpur
Introduction
The young of many mammalian species provide a variety of stimuli which influence vital parental responses. Bell & Harper (1977) state that primate and other mammalian young “must emit the appropriate cues and possess the necessary response capabilities to ensure that care giving is released and functional.” Ethologists and comparative psychologists have long recognized that proper care of dependent young requires that parent-offspring interactions be regulated by reciprocally exchanged signals, and it is becoming increasingly clear that both parent and young are endogenously tuned to detect and react to each other’s signals. In neonate mammals, behavior is typified from birth by reciprocal stimulative relationships between parents and young, whereby the neonate attracts the female, and the female presents a variety of primarily approach-inducing stimuli (Schneirla & Rosenblatt, 1961). Alley (1980) states that the distinctive markings and coloration found in young infants of many primate species function as signals which tend to “release” caregiving responses.
Jay (1962) suggests that three important elements (coat color, vocalizations, and quality of movements) in the infant’s appearance and behavior are essential in releasing maternal behavior of the mother (or other adult females) in langurs. Thus the body of a dead infant may continue to elicit maternal behavior (such as carrying and grooming) from its mother and other adult females for several days after death. The carrying of dead infants has been reported for several primate species in the wild: Van Lawick-Goodall (1967) in the chimpanzee (Pan troglodites); Schaller (1963) in the gorilla (G. gorilla gorilla); DeVore (1963) in savannah baboons (Papio cynocephalus); Kummer (1968) in hamadrayas baboons (Papio h. hamadryas); Prakash (1962) and Alley (1980) in rhesus macaques (Macaca mulatta); Rahaman & Parthasarathy (1969) in bonnet macaques (M. radiata); Merz (1978) in barbary macaques (M. sylvanus); Bowden et al. (1967), Cleve (1969), and Kaplan (1973) in squirrel monkeys (Saimiri sciureus); and Jay (1962, 1963), Mohnot (1974, 1977), Hrdy (1976), Rajpurohit (1987, 1992), and Winkler, Sommer, Bornes, and Paul (pers. communications) in the Hanuman langur (Semnopithecus entellus).
The Hanuman langur (Dufresne, 1797) is the best studied and the most adaptable south Asian colobine. They live in a wide range of habitats from the Himalayas and peninsular forests to semiarid lands, in villages and towns, and on cultivated land (Roonwal & Mohnot, 1977).
Data presented here are from a continuing seven-year-long study of the free-ranging langurs around Jodhpur (India). Jodhpur is located in Rajasthan at the eastern edge of the Great Indian Desert. This town is surrounded by semi-desert plateau, which is inhabited by a geographically isolated population of about 1850 langurs, comprising 32–34 one-male multi-female troops and 12–14 all-male bands. The climate is dry, with maximum temperatures about 48°C in May/June and minimum temperatures around 0°C in December and January. Jodhpur receives 90% of its scanty rainfall (annual average: 360 mm) during the monsoon (July to September).
The natural open scrub vegetation is dominated by xerophytic plants, including Prosopis juliflora, Prosopis cinararia, Acacia senegal, and Euphorbia caducifolia. The langurs feed on about 190 wild and cultivated plant species (for ecology, see Mohnot, 1974; Winkler, 1981; and Srivastava, 1989). For religious reasons local people provision most of the groups with vegetables, fruits, nuts, and cooked or uncooked food. In some groups the provisioning accounts for about two thirds of the total feeding time. Some groups raid crops and orchards but, because they are considered to be sacred, these langurs are never hunted. Apart from feral dogs, there are no natural predators in this area. The animals are easy to observe since they are not shy and spend most of the day on the ground.
The reproductive units are one-male multi-female troops with a single adult resident male. As a rule in this species, males never carry or feed infants. Each troop occupies its own home range of about 0.5–1.5 km². With few exceptions, females remain for life in their natal troops. Males emigrate, usually as juveniles, to all-male bands, whose home ranges can be as large as 20 km². According to censuses carried out in 1983–1986 (Mohnot et al., 1987), 1990 (Rajpurohit & Sommer, 1991), 1997–2001 (Rajpurohit, 1992), and 2005 (Rajpurohit et al., 2006), the number of one-male multi-female troops varied between 32 and 34. The mean troop size was 38.5 members (range, 7–124 animals). The number of all-male bands was about 13, and averaged 11.8 members (range, 2–47 members). The mean annual population consisted of 101.4 adult males (age > 7–8 years) and 500.5 adult females (ages > 3–4 years), yielding a mean of 17.4 reproductive females per troop. Male–female sex ratio at birth was 1.1 : 1, which among adults is female biased: 1 : 4.1 (Rajpurohit & Sommer, 1991). All-male bands invade home ranges of one-male multi-female troops in an unpredictable pattern, sometimes resulting in rapid or gradual replacement of the resident male (Sommer & Rajpurohit, 1989).
The Hanuman langur infant stage I (“black coat”) lasts from birth to 5–6 months; stage II (white or brown coat) lasts until weaning ends at about 12–15 months, when it is almost independent of its mother (Rajpurohit & Mohnot, 1991).
Theoretically, the age of mother, age of infant (at death), cause of death, and, to some extent type of habitat, should be responsible factors affecting the duration of keeping infants after their death. Predictions are (1) mothers should carry a dead infant longer if it died at an early age; (2) if death is natural or due to some unknown cause the mother should carry the corpse longer than in cases of accidents (e.g., electrocution or fall) or “mishandling” (see below); and (3) old mothers should carry their dead infants longer than young females do. All these predictions are tested in this paper.
Whenever a dead infant was seen being carried by its mother in this study site, further observation was continued every day until the carrier finally abandoned the corpse. Ten cases were observed from August, 2008, to July, 2010 (see Table 1).
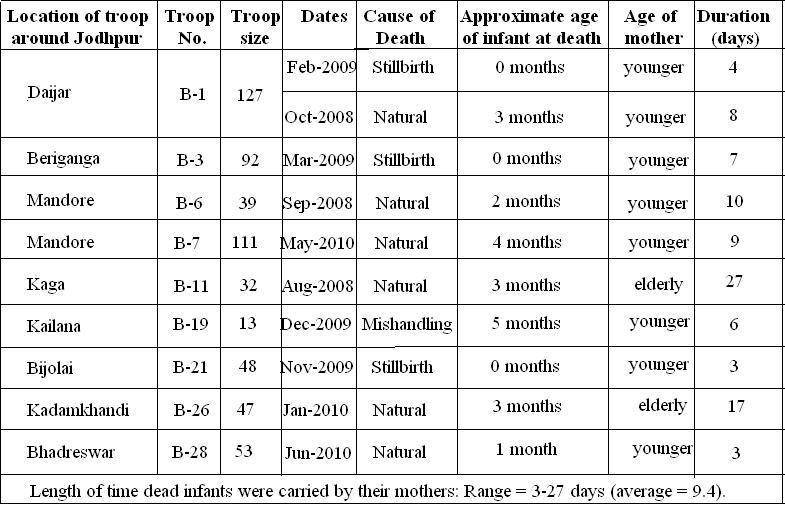
Table 1: Keeping of dead infants by mothers in Hanuman langurs, Semnopithecus entellus (Jodhpur 2008–2010).
Langurs at Jodhpur breed throughout the year: births occur year round but are not uniformly distributed. During this study, ten cases of dead infant carrying by their mothers were observed. In one case, in Kailana (B-19), an infant was killed by mishandling. We observed that this mother carried her infant for 6 days, after which the corpse, completely decomposed, was somehow hung on a tree branch. For the next week, the mother was frequently seen sitting near the decomposed body.
Of the 10 infant deaths, 3 were newborns found dead (considered stillbirths), 6 were natural deaths due to unknown causes, and one was due to mishandling, i.e., suffocation or choking – inexperienced mothers press infants hard during nursing and “kissing” and during infant transfer from one female to another.
In Kaga (troop B-11), an old female was noted carrying her infant for 27 days. It was a natural death of a 3-month-old. The dead infant became completely dry and decomposed. In another case, in Kadamkhandi (troop B-26), a female was observed carrying her dead infant for 17 days. In five cases, the ages of infants on their deaths were less than three months old, two were 4 and 5 months old respectively, and the remaining three were stillbirths. The causes of death we saw are categorized as (1) mishandling, (2) stillbirths, and (3) “natural” (unknown causes). In this study it was noted that after the change of coat color (stage I to stage II), dead infants were not carried.

Figure 1: Dead infant being carried by its mother.
However, the other factors, such as the age of the mother and cause of death, may also affect the duration of carrying. We observed the range of carrying dead infants to be 3–27 days (average: 9.4 days).
In troop Kaga in August, 2008, an old female was observed carrying a dead infant for 27 days. It was the longest period that a mother langur carried a dead infant during this study. She kept the infant’s body under her legs at feeding time, and was very careful of outside interaction. In troop Daijar-I in February, 2009, a female carried her newborn dead baby with the placenta for four days. In January, 2010, in Kadamkhandi, an old female carried a dead infant for 17 days.
Behavior of Other Troop Members The resident males ignored dead infants in the same manner as they do live ones. But other adult females approached the body, touched it, and sometimes groomed it. They responded this way for only a few minutes, and at feeding time they carelessly left it anywhere. The female juveniles and older infants of the troop were also observed smelling, touching, and watching the corpses. Sometimes juveniles picked up a dead infant, but in a few minutes they left it on the ground.

Figure 2: Troop members examining a dead infant.
The mothers of dead infants kept some distance from other troop members. In the first 2–3 days other females of a troop approached these mothers and spent some time with them. Many times they tried to take dead infants and groom them. We interpret these activities as “sympathy” towards the mothers.
Jay (1962) in the Hanuman langur, and Prakash (1962) in the rhesus monkey (Macaca mulatta), observed mothers carrying dead infants. Jay (1962) suggested that mothers should carry dead infants longer when they died at an early age. As an infant’s coat color changes, the interest of adult females declines and they no longer seek out the infant to hold and groom it as often. It is her speculation that the natal coat color of a dead infant is sufficient to initiate approach and grooming by adult females.
Van Lawick-Goodall (1967) reports that a female chimpanzee (Pan troglodytes) carried her dead infant for three days. Schaller (1963) saw a gorilla (G. gorilla beringei) mother carry a dead infant (which died two days after birth) four days before leaving it. A case of a free-ranging group of rhesus macaques (Macaca mulatta) in the Jackoo Forest, Simla, has been reported by Camperio Ciani (1984), in which a one-year-old male infant died due to an adult male’s attack. The supposed mother took the dead body and carried it 400 m into the forest and sat there with it, without following the rest of the troop. Severely injured infants (due to infanticidal attacks) in Hanuman langurs at Jodhpur were also observed being carried by their mothers or an allomother (Agoramoorthy & Mohnot, 1988; Rajpurohit, 1987; Sommer; 1987). Angst & Thommen (1977) also report other females carrying a newly injured infant in a semi-free-ranging colony of Barbary macaques (M. sylvanus). Rahaman & Parthasarathy (1969, p. 157) report that “a dead baby is carried for several days by its mother in bonnet macaques” (M. radiata). On the other hand, Merz (1978), in a study of a free-ranging population of Barbary macaques, reports that all ages (juveniles and up) and sex classes will carry dead infants.
Rumbaugh (1965) reported a case in which a mother squirrel monkey (Saimiri sciureus) continued to respond to her stillborn infant for more than six weeks while the infant remained in the cage with the mother and three other monkeys. Clewe (1969, p. 154) has suggested that the presence of hair may be the crucial stimulus in this species, since squirrel monkeys born without hair are dropped to the floor, while those born with it are held.
Kaplan (1973) found that the response of the male squirrel monkey to a dead infant depends partly on the age at which the infant dies. This kind of behavior has also been reported in savannah (Papio cynocephalus) and hamadryas (P. hamadryas hamadryas) baboons by DeVore (1963) and Kummer et al. (1970), respectively. A second hypothesis is that if infant death is due to some accident (e.g., electrocution or fall), mothers would not carry a corpse for as long as in the case of a natural death. This would be because, in an accident, she has seen the incident and probably could make out that there is something wrong and nothing can be done; but with natural death the mother may not understand what is wrong with the infant and therefore may carry it for some time, as if there werre a chance for recovery. It may be, as Zuckerman (1932) believed for the baboons, that langurs do not recognize the phenomenon of death, and this would seem to be supported by the continued attachment of mothers to their dead infants for long periods.
We predicted that old females should carry a dead infant (believing it to be unconscious) longer than younger females do. This might be because they are more worried about their reproductive success, as the fertile period left for them is shorter than that of young females. In the present study it was observed that two old females carried dead infants for the longest periods. Jay (1963), however, reports, without mentioning the age of langur females, that dead infants are carried until they are decomposed. Other adult and juvenile females approach a dead infant less frequently than a live infant, and stay in contact with it for a shorter time (or not at all) than with moving, vocalizing live infants. As in many primate species, langur mothers carry and protect dead infants for days after their death, suggesting that a reason or factor other than lack of vocalization or movement is responsible for this kind of maternal behavior. This factor might be natal coat color, as the coat color of a newborn infant of all species of Old World monkey is different from that of an adult of the same species. But then it is not clear to us why other females than the mother do not carry or groom a dead infant with its natal coat color, which otherwise they do with live infants.
Agoramoorthy, G., & Mohnot, S. M. (1988). Infanticide and juvenilicide in Hanuman langurs (Presbytis entellus) around Jodhpur, India. Human Evolution, 3, 279-296.
Alley, T. R. (1980). Infantile colouration as an elicitor of caretaking behaviour in Old World primates. Primates, 21, 416-429.
Angst, W., & Thommen, D. (1977). New data and a discussion of infant killing in Old World monkeys and apes. Folia Primatalogica, 27, 198-229.
Bell, R. Q., & Harper, L. V. (1977). Child effect on adults. Hillsdale. N J: Lawrence Erlbaum Associates.
Bowden, D. P., & Ploog, D. (1967). Pregnancy and delivery behavior in the squirrel monkey (Saimiri sciureus). Folia Primatologica, 5, 1-42.
Camperio Ciani, A. (1984). A case of infanticide in a free-ranging group of rhesus monkeys (Macaca mulatta) in the Jackoo Forest, Simla, India. Primates, 25, 372-377.
Clewe, T. H. (1969). Observations on reproduction of squirrel monkeys in captivity. Journal of Reproduction and Fertility, Suppl. 6, S151-S156.
DeVore, I. (1963). Mother-infant relations in free ranging baboons. In H. L. Rheingold (Ed.), Maternal behavior in mammals (pp. 305-335). New York: Wiley.
Dufresne, P. (1797). Sur une nouvelle espčce de singe. Bulletin des Sciences par la Société Philomatique de Paris, 2 [7], 49.
Hrdy, S. B. (1976). Care and exploitation of non-human primate infants by conspecifics other than the mother. In J. S. Rosenblatt, R. A. Hinde, E. Shaw, & C. Beer (Eds.), Advances in the study of behavior, VI (pp. 101-158). New York: Academic Press.
Jay, P. C. (1962). Aspects of maternal behavior among langurs. Annals of the New York Academy of Sciences, 102, 468-476.
Jay, P. C. (1963). The social behavior of the langur monkey. Ph.D. thesis, University of Chicago.
Kaplan, J. (1972). Differences in the mother-infant relations of squirrel monkeys housed in social and restricted environments. Developmental Psychobiology, 5, 43-52.
Kummer, H., (1968). Social organization of Hamadryas baboons: A field study. University of Chicago Press.
Kummer, H., Goetz, W. & Angst, W. (1970). Cross-species modifications of social behaviour in baboons. In J. R. Napier & P. H. Napier (Eds.), Old World monkeys (pp. 351-363). New York: Academic Press.
Merz, E. (1978). Male-male interactions with dead infants in Macaca sylvanus. Primates, 19, 749-754.
Mohnot, S. M. (1974). Ecology and behaviour of the common Indian langur, Presbytis entellus. Ph.D. thesis, University of Jodhpur.
Mohnot, S. M., Agoramoorthy, G., Rajpurohit, L. S., & Srivastava, A. (1987). Ecobehavioural studies of Hanuman langur, Presbytis entellus. Technical Report (1983-86) (pp. 1-89). MAB Project, Department of Environment, Govt. of India, New Delhi.
Prakash, I. (1962). Group organization, sexual behaviour and breeding season of certain Indian monkeys. Japanese Journal of Ecology, 12, 83-86.
Rahaman, H. & Parthasarathy, M. D. (1969). Studies on the social behaviour of bonnet monkeys. Primates, 10, 149-162.
Rajpurohit, L. S. (1992). Origin and composition of the unisexual unit: An all male band in Hanuman langurs, Presbytis entellus, around Jodhpur, India. Primate Report, 34, 47-52.
Rajpurohit, L. S. (1992). Mother-infant bond and dead infant carrying behaviour of female langurs, Presbytis entellus. Abstracts–XIVth Congress of the International Primatological Society (p. 216). August, 1992, Strasbourg.
Rajpurohit, L. S. (1987). Male social organization in Hanuman langurs (Presbytis entellus). Ph.D. Thesis, University of Jodhpur.
Rajpurohit, L. S., & Mohnot, S. M. (1991). The process of weaning in Hanuman langurs, Presbytis entellus. Primates, 32, 213-218.
Roonwal, L., & Mohnot, S. M. 1977. Primates of South Asia: Ecology, sociobiology and behaviour. Cambridge: Harvard University Press.
Rumbaugh, D. H. (1965). Maternal care in relation to infant behavior in the squirrel monkey. Psychological Report, 16, 171-176.
Schneirla, T. C., & Rosenblatt, J. S., (1961). Animal Research. I. Behavioral organization and genesis of the social bond in insects and mammals. American Journal of Orthopsychiatry, 31, 223-253.
Schaller, G. B. (1963). The mountain gorilla: Ecology and behavior. University of Chicago Press.
Sommer, V., & Rajpurohit, L. S. (1989). Male reproductive success in harem troops of Hanuman langur, Presbytis entellus. International Journal of Primatology, 100, 293-317.
Sommer, V. (I987). Infanticide among free-ranging langurs (Presbytis entellus) at Jodhpur (Rajasthan / India): Recent observations and reconsideration of hypothesis. Primates, 28, 163-197.
Srivastava, A. (1992). The honorary uncle. BBC Wildlife, 10, 25-32.
Srivastava, A. (1989). Feeding ecology and behaviour of Hanuman 1angur, Presbytis entellus. Ph.D. Thesis, University of Jodhpur.
Van Lawick-Goodall, J. (1967). Mother-offspring relationship in free-ranging chimpanzees. In: D. Morris (Ed.)., Primate ethology (pp. 287-346). London: Weidenfeld and Nielson.
Winkler, P. (1981). Zur oko-ethologe freilehender Hanuman langur (Presbytis entellus entellus) Dufresen [sic], 1797) in Jodhpur (Rajasthan), Indien. Ph.D. Thesis, Göttingen: George-August Universität.
Zuckerman, S. (1932). The social life of monkeys and apes. London: Kegan Paul, Trench, Trubner & Co.
-------------------------------------------
Author’s address: Dr. Goutam Sharma, Animal Behaviour Unit, Dept of Zoology, Jai Narain Vyas University, Jodhpur–342005 (Rajasthan), India [e-mail: [email protected]].
-------------------------------------------
* * *
Rita the Chimpanzee (Pan troglodytes ellioti):
The Start of an Enrichment Program at the Doha Zoo, Qatar
Hilda Tresz
Phoenix Zoo
When the word “success” is mentioned in this article, readers should look for the real meaning behind it. Success in this story is not simply improving the life of a solitary chimpanzee somewhere in a zoo. This article shows the year-and-a-half-long, incredible teamwork of a group of people from several different countries, who made something wonderful happen. This program was achieved despite monetary problems, cultural and religious differences, diverse life and work styles, different levels of knowledge and training, different ages (from 8 to 60), language barriers, computer problems, and time zone differences. When people were ready to learn and work, and wanted to make a difference for the greater good, they did so.
This article outlines the coordinated efforts of a group of selfless American teachers and diverse animal care professionals who started a program that made a difference, not only in the lives of the animals, but in the lives of the Doha Zoo staff and the local Qatari zoo visitors.
For an “animal person”, helping to improve an animal’s life is always rewarding. In Qatar the idea is supported by Islam, which encourages people to do good by their animals at all times. “People once asked Prophet Muhammad … ‘does God reward us when we treat animals well?’ Muhammad (Peace be upon Him) answered: ‘There’s a reward for acts of charity to every beast alive.’” (Mishkat al Masabih, Book 6, Chapter 6). Therefore, starting a program that would improve animals’ lives would be a very desirable task. Coupled with the wisdom of Hamad Salih Al-yazeedi, the Head of Doha Zoo (clearly a man of vision who thinks outside the box), it became a successful program that was praised on several occasions even by Gulf News.
Jenny Stands Wilson and Barbara Wilson (not related) were teachers at the Learning Center School and the American School of Doha in Qatar. Jenny and Barb’s students were part of Jane Goodall’s Roots & Shoots program, which teaches children all over the world to show care and concern for animals, people, and the environment. As part of this program the students began manufacturing behavioral enrichment items for the chimpanzees at Doha Zoo. During this time Jenny noticed that a female chimpanzee, Rita, was kept separated from the zoo’s pair, Timmy and Tina. Rita and the pair had no visual or tactile contact, and limited olfactory and acoustic contact, since their exhibits were far apart.

Figure 1: Jenny and Barb with Head Keeper of chimpanzees, Mohammed Tariq. Photo by Chris Wilson.
On March 21, 2009, Jenny contacted Dr. Jane Goodall, asking for instructions regarding potential improvements for Rita’s life. On March 23, Dr. Virginia Landau, Director of ChimpanZoo, a program of the Jane Goodall Institute, directed the assignment to me, and preparations began immediately. The essential first step was to get support from the management of the Doha Zoo. Jenny, already familiar with the Zoo and acquainted with Hamad Salih al-Yazeedi, became the “middle man” between me and the Zoo.
Jenny’s role was to create a report of the current situation, find resources to accomplish plans, involve teachers and students of the Learning Center School and the American School of Doha in developing enrichment ideas, coordinate the manufacturing of enrichment items, conduct and record behavioral observations, provide constant communication between me (the ChimpanZoo coordinator) and Doha Zoo management, and document proceedings by written reports (accompanied with pictures and videos) to Doha Zoo management and me.
On Monday, April 13, 2009, Jenny learned that the Doha Zoo had agreed to work with ChimpanZoo.
Rita’s origin was unclear. She was approximately 10 years old when the enrichment program started and had been housed alone all her life due to an unsuccessful introduction to Timmy and Tina, when they were all babies. Also, it was believed that she was a different subspecies from the pair. She was kept in a concrete cage with metal bars, some wooden furniture, and a tire, but no other objects to manipulate.

Figure 2: Rita alone. Photo by Barbara Wilson.
The primary goal was to develop a program which would elicit species-appropriate behaviors, using the limited available supplies. Following the daily activity budget of wild chimpanzees (Figure 3), it was decided to start by immediately providing Rita with sleeping and resting opportunities, as well as by helping her to spend most of her awake time foraging and playing.
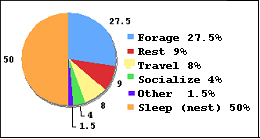
Figure 3: Average 24-hour cycle of a wild chimpanzee (percentages of 24 hours), from L. Brent: The care and management of captive chimpanzees (p. 18).
Short-Term Goals: To provide immediate relief. Enrichment ideas needed to be simple, free, delivered quickly, easy to supply, safe, non-threatening, and introduced one step at a time. Due to Rita’s fairly empty concrete cage, the choice of enrichments that first came to mind was: 1. Substrate to increase foraging time and for nest building. 2. Browse to increase foraging time and for nest building. 3. Variation of current diet presentation to increase foraging time (as in scattering and hiding chopped basic diet, and introducing novelty food items). 4. Manipulative items (toys, paper boxes, etc.) for playing and to increase foraging time.
Substrate was the first choice of enrichment because it was multi-purpose. Besides helping with foraging and nest building, substrate also was free (dry leaves, donated paper and clothing, soil, etc.) or cheap (hay, straw, etc.) and helped prevent health problems caused by wet or hot concrete.
However, in Qatar there were some cultural problems that prevented the use of substrates in exhibits. When visitors saw keepers scattering food in the outside cage, they demanded to be able to do the same. People also threw cigarettes inside exhibits, threatening fire within the cages. This is still an issue. Although the security guards were trained to watch visitor activities, it never became truly safe. The Zoo is working on educating the public on this issue. Rita also contributed to the problem herself, by refusing to go inside when outside substrate was available. Rita felt too comfortable to leave her outside nest in the evenings. Dry substrates such as shredded paper or hay were taken out of the exhibit and are now provided only in the night house. Outside, only sand is used.

Figure 4: Rita receives paper the first time in her life, April 28, 2009. Photo by Barbara Wilson.
Browse was the next logical step. Browse is one of the most important elements of foraging. It extends foraging time up to several hours, provides fiber, and has practically no nutrition value. It is the best material for nest building for chimpanzees. It is also free if cultivated and harvested on the grounds.
However, there were obstacles to providing browse as well. Green, leafy branches are difficult to come by in the desert. Most of the plants available were eucalyptus and date palm species. Eucalyptus is inedible by chimpanzees, and the date palm caused some blood in the stool. Finding other sources was a challenge, due to lack of plant knowledge and local language differences. A request for information about edible plants specific to that region was sent all over the world, and was answered by the Tel-Aviv Zoo and Haifa University of Israel. Pictures of local trees and bushes were taken by Jenny Wilson and sent to the Phoenix Zoo and to UNESCO for identification. I also sent a list of edible plants suitable for browse (by scientific and common names) to the Doha Zoo veterinarian, who is successfully giving the chimpanzees acacia now.
Variable presentation of the current diet was a high priority, since it is an important part of foraging. Initially, only those methods were chosen that were fast, free, and could be immediately implemented. Feedings were increased from once to twice per day. Food was chopped into very small pieces (but not diced so small that Rita would not bother gathering them) and scattered on the floor, and hidden in substrates or on furniture. However, scattering food could not be continued, due to both keeper workload and problems with visitors seeing food in the cage.

Figure 5: Doha Zoo staff hiding food in substrate on exhibit. Photo by Barbara Wilson.
Manipulative items. Rita received many non-food-related enrichments that she could play with. Items included paper products such as cardboard tubes and boxes, wrapping paper, books, ribbons, etc. Other items included a basketball, tennis balls, logs, clothing, plastic tubing, a mortar and pestle, a necklace, mirrors, cleaning brushes, a toothbrush and toothpaste, plastic dishes, and wicker baskets.
Long-term plans: Once the basic enrichment program was in place, it was time to work on long-term goals: 1. Creating reusable/permanent fixtures that elicit species-appropriate behaviors. 2. Finding resources to fund these projects. 3. Documenting enrichment. 4. Companionship for Rita. 5. Educating staff to continue with enrichment.
Creating reusable/permanent fixtures: Keeping in mind the goals of stimulating basic chimpanzee behaviors (Figure 3), the features designed first were to support resting, sleeping, foraging, and manipulating behaviors. These were a metal sleeping basket and a metal puzzle feeder. The basket (Figure 6) allowed Rita to sleep high in the “canopy”, in a stable, permanent nest, feeling safe and secure for the first time in her life – just as she would have felt in the wild. The puzzle feeder was a locked, simple mesh cage which was easy to clean and refill without taking too much of the keepers’ time. Originally it was placed in a fairly high area, but it turned out to be hard to reach for the keepers to fill. It has not yet been moved and is currently not being used.
Finding resources. What can be done, and what should be done, as well as issues of time, money and manpower, were resolved by organizing a support system. All enrichment items were made by Qatari school children and their teachers and then distributed during the weekends. Three groups belonged to Jane Goodall’s Roots & Shoots program and as such, spent most of the weekends observing and documenting that enrichment at the Doha Zoo. Besides their time they also provided almost all of the financial support.
Documenting enrichment. Experience shows that keeping up with documentation is one of the hardest tasks in any program. When it comes to inventing and implementing new ideas, creativity almost always outpaces resources, such as time, money, and staffing. The collection of data by students, even as young as 8 to 12 years, served several very important purposes. When the children could observe their enrichment items being used with their own eyes, they could also prove the usefulness of their enrichments by documenting the chimpanzees’ behavior with them and then evaluating the items during classes – certainly a successful teaching/learning process. Having the students and teachers helping out also freed up some time for the Doha Zoo staff; however, sometimes the overwhelming amount of incoming enrichment items created extra work for them. In gratitude the kids make thank-you cards and gift boxes for the staff.
Companionship for Rita. Naturally, Rita’s new enrichment program also affected the other two chimpanzees’ lives, as Timmy and Tina received items similar to hers.
It is well known that of all enrichment categories, social enrichment is the highest priority for any species that do not lead solitary lives. The integration of the three chimpanzees was imperative. With the help of the Institute of Vertebrate Biology, Academy of Sciences of the Czech Republic, it was ascertained that all three chimpanzees were the same subspecies, matched on the matrilineal side.
The three chimpanzees were successfully introduced to each other in June of 2010. The details of the introduction will be described in a separate article.
Educating Staff. New programs and new ideas cannot be successfully implemented without the staff fully understanding the reasons behind them. The ideas need to be plausible, and staff need to understand the final outcomes as well as all the details that will lead to accomplishing them. The Doha Zoo management worked continuously with its staff and provided help whenever possible to make the new program happen. Everybody was on board! In June, 2010, because of the increasing enrichment programs, and to ensure continuing education of staff, The Doha Zoo organized its first five-day enrichment workshop. Beyond a doubt this opened people’s eyes to the endless possibilities which can improve animal welfare.
“He is not a perfect believer, who goes to bed full and knows that his neighbor is hungry” (Mishkat al Masabih, Book 2, Chapter 424) is truly a phrase that should guide us all regardless of our beliefs. No one should walk by any unwanted situation and not do something about it, whether the motivation is from religious principles or one’s own conscience – or any other reason. And no one needs to feel alone in doing so, either. The story of Rita illustrates this. She went from being a lonely chimpanzee to a well-adjusted animal living in a small but functioning social group, receiving enrichment daily and exhibiting species-appropriate behaviors. All thanks are due to a group of people and their continuous efforts to do better. In return Rita touched their souls, and by doing so encouraged them to do more, not just for her but for all animals. As a result, an enrichment program that first focused on one individual chimpanzee and then on two others, has spread out to more species that needed immediate help, and from there has developed into a zoo-wide program. What more can one want? Is this not the real meaning of success?

Figure 6: Rita in new basket nest. Photo by Barbara Wilson..
Special thanks to all the people who participated in this project: Hamad Salih Al-Yazeedi, Mohammed Hussein Hassan, Moza Ahmed Al-Qahtani, Abubaker Hamuda, DVM, Selma Al-Shaik, Mohammed Abdulraheem, Mohammed Nurnobi Chowdhory, Mohammed Tariq, Jenny Stands Wilson, Chris Wilson, Barbara Wilson, Jody Kruger, Tawney Kruger, Andra Vizante, Joyce Liberto, Peter Vallo, PhD, Klara J. Petrzelkova, PhD, Mwanahamissi Issa Mapua, BSc, Josef Bryja, PhD, Benno Boer, PhD, Itai Roffman, Ori Pomerantz, Julie Swenson, DVM, the Learning Center School of Qatar Foundation, the American School of Doha, Qatar Academy School, and the American Women’s Association of Doha.
The Arabic phrase “Bismillah ir-Rahman ir-Rahim” is often said when one is embarking on any significant endeavor. It is a beautifully poetic expression which offers both insight and inspiration. It is almost impossible to capture either the true depth of meaning or the inspirational message of this beautiful phrase by simply translating it. One poetic translation is: “Every moment of this life is filled with your eternal radiance, my Beloved, You are the Beneficent One who endlessly showers all of creation with nourishment and blessings, and the One who generously rewards those who live in harmony with Your Divine Will.” For further information see: <wahiduddin.net/words/bismillah.htm>.
-------------------------------------------
Author’s address: 455 N. Galvin Pkwy, Phoenix, AZ 85008 [e-mail: [email protected]].
-------------------------------------------
* * *
On November 15, Viktor Reinhardt < viktor@ snowcrest.net> posted the following to the LAREF (Laboratory Animal Refinement & Enrichment Forum) e-mail discussion list: “Research protocols sometimes require that compatible macaque pairs be physically separated for a limited time period (e.g., controlled food-intake studies; sample collection from chair-restrained subjects).
“Obviously – and as documented in the literature – partner separation is a stressful event that not only has animal welfare implications but can also skew subsequently collected research data.
“If pair-housed macaques in your care have to be separated, (a) what do you do to minimize the stress for the animals; or, if you don’t have the authority, (b) what would you do to minimize the stress for the animals?”
Thomas Ferrell < [email protected]> responded: “We use either a single-wire divider panel or a solid Plexiglass panel, depending on the situation. In either case, separation is for the shortest time possible.”
Melissa Truelove < [email protected]>, of the Behavioral Management Unit at Yerkes NRPC, wrote: “Allow maximum visual contact (e.g., lexan [transparent] solid panel if monkeys can tolerate it) during separation period or maximum limited physical contact (e.g., perforated panels) with their partner(s). I’ve worked with several pairs who have remained separated for projects for more than two months with a solid lexan panel in place and successfully returned to pair housing following completion of the project. From a behavioral management perspective, it made the reintroductions easier as the animals maintained visual (and thus social) contact for the duration of the required separation. The same can be said for separation using perforated panels. These types of separation should be less disruptive than total separation from a partner, thus should minimize stress. Also a privacy panel/visual barrier can be helpful during times when a partner isn’t available.”
Erik Moreau < [email protected]>, an animal health technician from a contract research organization in Montreal, wrote: “The animals are separated with visual, olfactory, and limited physical contact through a wire-mesh separator.”
Later that same day, Viktor posted another query: “Are any of you aware of published studies assessing stress parameters of macaques who maintained visual contact during physical, research-related separation versus those who were separated by moving one animal out of the room?”
Marcie Donnelly < [email protected]> responded: “I am not aware of any published studies, but I would guess there are some out there.
“We do on occasion take a partner out for a pharmacokinetic study – but they would only be removed from the room for about nine hours (the day pretty much), then returned and fed. Many times they remain in visual contact overnight to allow the monkeys to eat. If they finish eating everything before we go home they are returned to pair housing for overnight.
“In the past we have had to separate monkeys for feces / urine collection, as well as food consumption, and we have kept the partners on visual access via mesh. They do well and are able to be re-paired. This would be for one week generally – we did have a pair of girls and one would not eat well while separated from her partner – makes you sad for sure. It also tells you there is some stress related to the separation. They have visual, tactile (but the screens are small), and olfactory contact, but they can’t be next to one another and groom, or eat next to each other. I think that can be really hard for them.”
* * *
The Third East Mediterranean ICLAS Symposium, organized by the Laboratory Animal Science Association (LASA)–Turkey in association with International Council for Laboratory Animal Science (ICLAS), will be held in the city of Istanbul, Turkey, June 13–15, 2011. The symposium is open to everyone interested in Laboratory Animal Science and experimental studies. Many topics will be under discussion, including ethical evaluation, experimental design, new technologies in laboratory animal science, animal models, methods for reduction, etc. Siyami Karahan, President of (LASA)–Turkey, invites you to participate in the meeting and welcomes you to submit abstracts for oral or poster presentations. For further information, see <www.iclas2011istanbul.org/default.asp>.
The American College of Veterinary Internal Medicine (ACVIM), the mission of which is to enhance animal and human health by advancing veterinary internal medicine through training, education, and discovery, will hold its 2011 Forum in Denver, Colorado, June 15–18. See <www.acvim.org/websites/forum2011/index.php?p=460>.
In association with BioMed Central, the Society of Chinese Bioscientists in America’s 13th International Symposium ‘Bioscience for the 21st century: Emerging Frontiers and Evolving Concepts’, takes place in Guangzhou, China, July 25–29, 2011. Visit the conference Website, <www.scbameeting2011.org>, for further information. Then, register at <www.scbameeting2011.org/Channel/register.aspx>; and submit your abstract at <www.scbameeting2011.org/channel/submission.aspx>.
The 13th Congress of the European Society for Evolutionary Biology will be held in Tübingen, Germany, from August 20–25, 2011. See <www.eseb2011.de>
* * *
Information Requested or Available
Great Apes in Japan
The Great Ape Information Network has created an English homepage and database on great apes living in Japan: <www.shigen.nig.ac.jp/gain/index.jsp>. The Network writes, “Though some parts are still only in Japanese, we will complete it as soon as possible.”
New ASP Website
The American Society of Primatologists has a new Website, at <www.asp.org>, with a complete redesign by a professional designer. There is still content to be incorporated into the new Website; it is not fully complete as of this writing. Some pages may redirect to the old Website, and a few may redirect to a broken link, as those pages have not been entered yet. The programming is complex because all content will now be in a database (instead of as static html), and an interface has been created that will allow specific people (such as committee chairs) to enter content directly. Be patient; it should be worth it!
Interesting Websites
* * *
Special OLAW Online Seminar
“AC and OLAW Perspective on Nonhuman Primate Enrichment and Social Housing” was recorded by the National Institutes of Health (NIH), Office of Laboratory Animal Welfare (OLAW); U.S. Department of Agriculture (USDA), Animal and Plant Health Inspection Service (APHIS), Animal Care (AC); and USDA, National Agricultural Library (NAL), Animal Welfare Information Center (AWIC) on June 11, 2010. The seminar can be viewed as Power Point at <https://webmeeting.nih.gov/p13739483/>, or as a PDF file at <grants.nih.gov/grants/olaw/NHP_Enrichment_transcript.pdf>.
Speakers at the Seminar were Dr. Robert Willems, Assistant Regional Director, Eastern Region, USDA, APHIS, AC; Dr. Axel Wolff, OLAW, Director, Division of Compliance Oversight; and Ms. Kristina Adams, Technical Information Specialist, USDA, NAL, AWIC.
Moderators were Dr. Jerry Collins, OLAW, Division of Policy and Education and Yale University; and Ms. Susan Silk, OLAW, Director, Division of Policy and Education.
The subject of the presentation is social housing of nonhuman primates in a research setting. Social housing is a part of the enrichment program for nonhuman primates required by the Animal Welfare Act regulations. Dr. Willems presented the regulatory requirements for social housing of nonhuman primates and covered the allowable exceptions to that requirement.
Questions may be submitted to OLAW [e-mail: [email protected]].
CANTAB Cognitive Testing System
The IntelliStation T is the latest development in the CANTAB Cognitive Testing system for nonhuman primates. It retains all the test-specific criteria, stimulus presentation, and animal response/reward hardware of all past systems. The application portfolio comprises a battery of tests performed via a touch screen. Test protocols are flexible and can be configured by the user. Multiple tests may be linked in the same subject session.
Monkey CANTAB is comprised of the following:
If you have questions, you can contact Vern E. Davidson, Neuroscience Product Manager, Lafayette Instrument Co. [765-423-1505, Ext. 206; Fax: 765-423-4111; e-mail: [email protected]].
2011 Bonobo Calendar
Marian Brickner, some of whose bonobo photographs have appeared in the LPN (including the current cover!), has produced a beautiful 2011 calendar, which is available on a CD for $7, including U.S. postage. See <marianbricknerphotography.com/Bonobos.aspx>, or contact her (766 Harvard Ave, St. Louis, MO 63130 [314-680-4333; e-mail: insect1 @swbell.net]).
Hominoid Carrying Archive Online
Dr. Bill Sellers, of the University of Manchester, announces the availability of the Hominoid Carrying Archive online. This is a large collection of video data collected as part of a Natural Environment Research Council project into the carrying behavior of great apes. The video is in standard digital format with an Audio Video Interleave wrapper that should be readable by almost any video software. Basic joint center motion capture was performed by hand for almost all the clips presented and this data is available too. Hopefully you will find some of this information useful for teaching or research and you can access the library through the Animal Simulation Laboratory Website at <www.animalsimulation.org>.
For more information, contact Dr. Sellers at the Fac. of Life Sciences, Univ. of Manchester, 3.614 Stopford Bldg, Oxford Rd, Manchester, M13 9PT, U.K. [e-mail: [email protected]].
Procedures with Care
Paul Flecknell, of the Comparative Biology Centre at the Medical School in Newcastle, U.K., announces the launching of a new Website, the development of which was co-funded by the IAT and NC3Rs: <www.procedureswithcare.org.uk>. He writes, “We will be adding more material to the site over the next month or so. At present it covers a series of injection techniques with rat and mouse, and on each ‘procedure’ page there is a ‘trainer’s pack’ that lets you download the video and still images for use in your own training activities.”
We look forward to the addition of nonhuman primates to the animals covered on this site.
Lincoln Park Zoo’s Blood Type Database for Apes
At present, no standard blood type data has been determined for the great ape (gorillas, chimpanzees, bonobos and orangutans) population in zoos. The knowledge of blood types would provide better acute veterinary medical care, and reduce transfusion reactions by knowing compatibility of donor and patient blood.
In March of 2006 Veterinarian Dr. Kathryn Gamble and Master Keeper Jill Moyse started the project “Cataloging blood types for SSP populations of great apes”, which aims to gather and test blood samples from zoo apes as they undergo their routine physicals. All four great ape species are being tested and the results are entered into a database coordinated at the the Zoo’s Lester E. Fisher Center for the Study and Conservation of Apes
. Eldon Biologicals donated the bloodtyping cards that are used and validation of the cards is being performed at the University of Chicago, since the cards were designed for human blood types. The project has grown significantly, to include not only all accredited zoos that house great apes in North America, but also populations in European zoos (EEP), PASA (Pan African Sanctuary Alliance) sanctuaries, and the Nyaru Menteng Orangutan Rehabilitation Center. As of 2009, they had received 539 completed blood typing cards. They now have typed blood from about 700 animals around the world. – from the Lincoln Park Zoo Webpage, and a December 5, 2010, article in the Chicago Sun-Times
* * *
* * *
The American Society of Laboratory Animal Practitioners (ASLAP) Veterinary Student Liaison Committee (VSLC) is seeking nominations of veterinary students for the Society’s annual Veterinary Student Award Program (VSAP). The VSLC, with approval of the Board of Directors, has recently expanded the Veterinary Student Award Program to allow for one award to be given at each veterinary school. Last year 25 awards were given.
The VSAP is intended to increase awareness of the practice of laboratory animal medicine by recognizing students who have demonstrated significant interest and potential in the field. Awards (a certificate, one year’s membership in ASLAP, and a cash sum of $300) will be presented at each school during their awards program or another suitable public event involving a large proportion of the student body.
Nominations are welcomed and encouraged from any current national ASLAP member and should be accompanied by a brief statement from the member that explains why the student should be recognized. Please note that students who wish to nominate themselves or another student must be members of the national ASLAP organization. Nominations must be received by January 14, 2011, and must include contact information (including e-mail) for the nominee.
Nominees will be asked to submit a current CV and an essay of up to 500 words describing their participation in activities related to laboratory animal medicine and how these activities have influenced their interests and career goals. Information will be evaluated by the Veterinary Student Liaison Committee.
Please send nominations to Judy Daviau [e-mail: [email protected]].
Please support and encourage a veterinary student you know who is interested in laboratory animal medicine by nominating them for this award.
* * *
Veterinary Residency, New York
The Institute of Comparative Medicine at Columbia University has a postdoctoral research scientist position available for a highly motivated veterinarian with a background in veterinary practice and comparative medicine (Requisition # 0001696). The residency in Comparative Medicine focuses on phenomena basic to disease in all species, including humans, and provides a challenging intellectual opportunity to work with medical, biomedical and veterinary specialists. The residency prepares the candidate for board certification by the American College of Laboratory Animal Medicine (ACLAM) and a career in biomedical research. See <www.cumc.columbia.edu/dept/icm/rtp.html> for details.
There are two options: 1) Residency – Main Track: This 3-year program rotates through medicine, surgery, pathology, and research, with substantial didactic and experiential components. 2) Residency – Two-Year Option: Like 1), without the research project if the applicant has already satisfied the ACLAM requirement for research training and manuscript publication.
Columbia University offers a competitive package, including a family health program, and four weeks of vacation. Interested individuals should apply at <academicjobs.columbia.edu/applicants/jsp/ shared/frameset/Frameset.jsp?time=1289849645830>.
If you have any questions, please contact Rivka Shoulson, DVM, MPH, Inst. of Comp. Med., Columbia Univ. Med. Ctr, 650 W. 168th St, BB 1810, New York, NY 10032 [[email protected]].
Laboratory Animal Science Management Courses
The Guido Bernardini Foundation (FGB) is a not-for-profit organization established to promote biomedical research by focusing on providing training and continuous professional development for laboratory animal science professionals. The FGB is pleased to announce the upcoming Laboratory Animal Science Management Courses to be held in Milan, Italy, in 2011:
Wildlife Studies and Primatology – Japan
The Primate Research Institute and the Wildlife Research Center, both affiliated with Kyoto University, have launched a new graduate program, the Center for International Collaboration and Advanced Studies in Primatology, aimed at facilitating international collaboration and advanced studies in conducting multidisciplinary studies of wildlife in either field or captive settings. The details are available at <www.pri.kyoto-u.ac.jp/sections/CICASP>.
Education and Training in Field Primatology
The American Society of Primatologists (ASP) will offer two scholarships annually to outstanding undergraduate and graduate students for research and career training in a field course at La Suerte Biological Research Station, Costa Rica, or Ometepe Biological Research Station, Nicaragua, funded through an educational grant from the Maderas Rainforest Conservancy (MRC). The goals of the MRC are to advance research, education and conservation of primates and tropical forests. Interested students from anywhere in the world are encouraged to apply.
The scholarship provides full tuition for an approximately one-month course, including room and board (3 meals/day), instruction, access to all facilities at the field site, and in-country transportation to and from the field site. The scholarship does not include airfare to Costa Rica or Nicaragua, books, supplies, field equipment, medical insurance, or hotel/meals the first and last night in San Jose (Costa Rica) or Managua (Nicaragua). Field courses are offered during December–January and from May through August.
Students awarded scholarships will be selected by the ASP Awards Committee based on the strength of the student’s academic record, responses provided in the application materials, and a letter of recommendation.
Additional information and application instructions and forms can be found at: <www.asp.org/grants/scholarship/index.cfm>. Online submission will begin January 1, 2011. The student must submit his/her completed application materials to the ASP Awards Committee by March 1, 2011.
For questions concerning the scholarship application, contact Peter Judge [e-mail: [email protected]]. For questions concerning course offerings, course content, and field site facilities, contact Paul A. Garber [e-mail: [email protected]].
Husbandry for Zoo and Aquarium Animals – Mexico
The Guadalajara Zoo and The Mexican Institute for Wildlife and Companion Animals invite you to participate in the course: “Behavioral Husbandry (Enrichment and Training) for Zoo and Aquarium Animals (Terrestrial and Marine Mammals, Birds, Reptiles, Fishes)”, which will be held at the Guadalajara Zoo, Jalisco, Mexico, May 17–21, 2011. This course is designed for zookeepers, aquarists, trainers, curators, veterinarians, related career students, and anyone interested in the topics.
The registration fee is US$150 before February 17, US$200 before April 17, and US$250 after April 17. Daily rate is US$65. The fee includes: lectures, practices and visits around the Zoo, translation if needed (English – Spanish – English), Proceedings, a Certificate of Attendance, transportation (Hotel – Zoo – Hotel), coffee breaks, and closing lunch.
For information about the city, transportation, and attractions, please visit: <www.guadalajaramidestino .com/index.php?lang=en>. For more information about and registration for this course, contact the Educational Dept, Guadalajara Zoo, Paseo del Zoológico #600, A. P. 1-1494 C. P. 44390, Guadalajara, Jalisco, México [Tel. +52 (33) 3674 4488 / 01 800 000 8000; Fax +52 (33) 3674 3848; e-mail: [email protected] and [email protected]].
* * *
Devra G. Kleiman: Conservation Biologist
Devra G. Kleiman, 67, a biologist whose groundbreaking research on giant pandas and South American monkeys showed how zoos can play a critical role in preserving endangered species, died April 29 at George Washington University Hospital. She had cancer. In a career spanning more than 40 years, much of it at the National Zoo, Dr. Kleiman helped create and define the new field of conservation biology.
She was perhaps best known for spearheading an unprecedented international effort to save golden lion tamarins – small, reddish-orange monkeys that live in Brazil's Atlantic coastal forests – from extinction. In the early 1970s, Dr. Kleiman responded to an alarm sounded by Brazilian biologist Adelmar Coimbra Filho. Golden lion tamarins were in trouble: research showed there were only several hundred of the animals remaining in the wild and fewer than 75 in captivity. Kleiman and Coimbra helped persuade officials at more than a dozen zoos not to sell their golden lion tamarins for profit. Instead, the zoos would lend the animals to one another for breeding. Eventually they gave up title to the tamarins altogether, ceding ownership to the Brazilian government. Dr. Kleiman played monkey matchmaker, using genetic data to determine which animals should mate to create strong offspring.
Those offspring were reintroduced to Brazil, where Kleiman and Coimbra helped preserve and restore wide swaths of the animals' habitat. Today, about 1,600 golden lion tamarins live in the wild. Another 500 live in 145 zoos around the world. The species' status has been changed from critically endangered to endangered, and a Brazilian organization that Kleiman helped found is coordinating efforts to ensure the species' long-term survival.
The cooperative model Dr. Kleiman pioneered with the golden lion tamarin project has since been widely adopted as the most effective way to manage the genetics of rare species. It has been crucial to the successful reintroduction to the wild of species including the black-footed ferret and the California condor. – by Emma Brown, Washington Post Staff Writer, May 4, 2010
Will Aging Research Chimpanzees Be Relocated?
“Most of the chimpanzees at the Alamogordo Primate Facility are well into their twenties and have not been research subjects for more than a decade. After a ten-year hiatus, the chimpanzees of the Alamogordo Primate Facility in New Mexico are being called back to duty. The 186 chimps, already grizzled veterans of medical research, will be pulled from an unofficial retirement and sent back into the lab by the end of 2011, the National Institutes of Health (NIH) announced in August. But the decision has brought to a head a simmering debate about the use of chimpanzees for medical research in the United States – a practice banned by the European Union this September. The chimps would rejoin a dwindling cadre of research primates. In 1995, the NIH established a moratorium on chimpanzee breeding in federally supported laboratories, and scientists have developed alternative ways to study the basic biology of diseases. But the roughly 700 chimps remaining still have a key role in vaccine testing for viruses such as hepatitis C and HIV, which don’t infect other laboratory animals.” – by H. Ledford, from Nature, October, 2010, 467, 507-508, <www.nature.com/news/2010/100927/full/467507a.html>
NY Must Release Records on Monkey Research
A judge in Albany, New York, says New York must release records on substance abuse experiments on monkeys and other animals. The state’s Office of Mental Health tried to block the Freedom of Information Act request from the Physicians Committee for Responsible Medicine (PCRM), saying research scientists may be targeted by animal-rights terrorists. In a ruling PCRM received this week, state Supreme Court Justice Richard Platkin rejected the state’s argument. Many of the experiments conditioned rhesus monkeys to have drug and alcohol addictions, then tested them to see if medications broke the addictions. – PCRM press release, November 12, 2010
AVMA Revises Veterinarian’s Oath
The American Veterinary Medical Association (AVMA) has revised the Veterinarian’s Oath — which all graduates of U.S. veterinary schools take — to stress the importance of animal welfare. The revised oath, approved by the AVMA Executive Board at its meeting this month, is as follows: “Being admitted to the profession of veterinary medicine, I solemnly swear to use my scientific knowledge and skills for the benefit of society through the protection of animal health and welfare, the prevention and relief of animal suffering, the conservation of animal resources, the promotion of public health, and the advancement of medical knowledge.”
“The Veterinarian’s Oath reflects every veterinarian’s aspirations for themselves and the veterinary profession,” says Dr. Bruce Nixon, Chair-Elect of the Animal Welfare Committee. “It’s a promise that each veterinarian makes at graduation, so these words have tremendous meaning. The Animal Welfare Committee recommended these changes to emphasize that veterinarians have responsibilities not only to animal health but also to animal welfare.”
“These changes make it clear that the scope of veterinarians’ efforts toward improving animal welfare include not only treatment, but also prevention of suffering and promotion of good welfare, which is consistent with today’s approach to veterinary practice,” adds Dr. Gail Golab, Director of the AVMA’s Animal Welfare Division.
For more information, see <www.avma.org>. – AVMA Press Release, December 2, 2010, Schaumburg, Illinois
Gorilla Tested for Possible Human Disease
A female gorilla at the San Francisco Zoo is recovering after a –unique medical procedure. This gorilla has undergone a special kind of surgery, and a local doctor is helping to diagnose what appears to be a human disease.
At the gorilla enclosure, dominant silverback Oscar Jonesy watched intently, as technicians anesthetized his sister, 29-year-old Zura, and took her to the zoo hospital.
“She has had chronic medical problems with her gastrointestinal tract, mainly diarrhea, constipation, bloat,” said zoo veterinarian Dr. Graham Crawford. Unlike the other gorillas, Zura will eat cups, purses, and even cell phones dropped within her reach.
“She’s had some GI issues,” said zoo primate curator Corinne MacDonald, “and we’re wondering whether or not it is because of that, or if there something else going on.”
As with any other patient, doctors needed to get a closer look, and so this first-of-its-kind here procedure called a capsule endoscopy, in which a tiny video camera is actually inserted into Zura’s intestines. “We think that probably she has Crohn’s disease, based on a lot of serological markers, blood tests that we’ve taken on her,” said gastroenterologist Dr. David Shields.
Gorillas carry the gene for Crohn’s, a sometimes fatal autoimmune disease. Powerful drugs can treat it. “All these drugs have risks,” said Shields. “Unless we’re sure that this is the case with her, we’d like not to treat her.”
Results from the implanted camera should be available in a few days. Zoo veterinarians will then decide what to do. – NECN/KTVU, December 6, 2010
Census: Mountain Gorilla Population Grows
The population of mountain gorillas in their main Central African habitat has increased by a quarter in seven years, regional authorities say. Most of the world’s mountain gorillas are found in the Virunga massif, which includes three contiguous national parks in Rwanda, the Democratic Republic of Congo (DRC) and Uganda. The population of the iconic but endangered animal in that area increased from 380 individuals in 2003 to 480, according to a census carried out earlier this year and funded by a number of local and foreign wildlife organizations.
“The increase in mountain gorilla numbers is a testament that we in the Virunga massif are all reaping from the conservation efforts sowed on a daily basis,” Rica Rwigamba, of the Rwanda Development Board said.
The only place outside of the Virunga massif where mountain gorillas are found is Uganda’s Bwindi Impenetrable National Park. Along with 302 individuals believed to dwell in Bwindi and four orphans living in a sanctuary in DRC, the census put the known world population of mountain gorillas at 786.
Mountain gorillas, who have fallen prey to conflict and poaching over the years, were brought to the world’s attention by the late Dian Fossey and are one of the region’s main tourist attractions. – AFP©, a news agency, December 7, 2010
* * *
President Obama Honors Nation’s Top Scientists and Innovators
On October 15, President Obama named ten eminent researchers as recipients of the National Medal of Science, and three individuals and one team as recipients of the National Medal of Technology and Innovation, the highest honors bestowed by the United States government on scientists, engineers, and inventors.
Mortimer Mishkin, Chief of the Section on Cognitive Neuroscience in the Laboratory of Neuropsychology, National Institute of Mental Health, is one of the Medal of Science recipients. The Section he leads uses a multidisciplinary approach to investigate the neurobiological mechanisms underlying learning and memory in primates. In monkeys, the approach involves:
* * *
Announcements from Publications: The Journal of Evolutionary Biology Research
The Journal of Evolutionary Biology Research (JEBR) is a multidisciplinary, peer-reviewed, open access journal, published monthly by Academic Journals, <www.academicjournals.org/JEBR>. JEBR is dedicated to increasing the depth of evolutionary biology research across disciplines, with the ultimate aim of improving evolutionary research. JEBR welcomes the submission of manuscripts that meet the general criteria of significance and scientific excellence in this subject area, and will publish:
* * *
Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
Books
* Ape House. S. Gruen. New York: Spiegel & Grau, 2010. 320 pp. [Price: Hard-cover: $26]
. . .This novel was inspired by the author’s visits to the Great Ape Trust, in Des Moines, Iowa. It is available as an unabridged audio compact disc, an unabridged audio-book download, an E-Book, and a large-print trade paperback, as well as a hard-cover.
* Biologie Moleculaire et Phylogenese des Lemuriens (Madagascar): Les Lemuriens, Primates de Madagascar. D. Montagnon. Sarrebruck, Germany: Editions Universitaires Europeennes, 2010.
* The Encyclopedia of Applied Animal Behaviour and Welfare. D. S. Mills, J. N. Marchant-Forde, P. D. McGreevy, D. B. Morton, & C. J. Nicol (Eds.). Wallingford, U.K.: CABI, 2010.
* The Infanticide Controversy: Primatology and the Art of Field Science. A. Rees. University of Chicago Press, 2009. 288 pp. [Price: $40]
* The Mind of the Chimpanzee: Ecological and Experimental Perspectives. E. Lonsdorf, S. R. Ross, & T. Matsuzawa (Eds.). University of Chicago Press, 2010. [Price: Cloth: $125; Paper: $49; E-book: $7 to $49]
. . .Contents: Foreword, by Jane Goodall; The chimpanzee mind: Bridging fieldwork and laboratory work, by T. Matsuzawa. Part I. Cognitive Mechanisms. Early social cognition in chimpanzees, by M. Myowa-Yamakoshi; Using an object manipulation task as a scale for comparing cognitive development in chimpanzees and humans, by M. Hayashi; Do the chimpanzee eyes have it? by M. Tomonaga; Understanding the expression and classification of chimpanzee facial expressions, by L. A. Parr; Behavioral and brain asymmetries in chimpanzees, by W. D. Hopkins, J. Taglialatela, D. A. Leavens, J. I. Russell, & S. J. Shapiro; and Trapping the minds of apes: Causal knowledge and inferential reasoning about object-object interactions, by J. Call. Part II. Tool Use and Culture. A coming of age for cultural panthropology, by A. Whiten; The cultural mind of chimpanzees: How social tolerance can shape the transmission of culture, by V. Horner; How are army ants shedding new light on culture in chimpanzees? by T. Humle; The complexity of chimpanzee tool-use behaviors, by C. M. Sanz & D. B. Morgan; Tools, traditions, and technologies: Interdisciplinary approaches to chimpanzee nut cracking, by D. Biro, S. Carvalho, & T. Matsuzawa; Ubiquity of culture and possible social inheritance of sociality among wild chimpanzees, by M. Nakamura; and New theaters of conflict in the animal culture wars: Recent findings from chimpanzees, by W. C. McGrew. Part III. Social Minds: Ecological Perspectives. Chimpanzee minds in nature, by J. C. Mitani, S. J. Amsler, & M. E. Sobolewski; Vocal communication in chimpanzees, by K. Slocombe & K. Zuberbühler; The function and cognitive underpinnings of post-conflict affiliation in wild chimpanzees, by R. M. Wittig; and The role of intelligence in group hunting: Are chimpanzees different from other social predators? by I. C. Gilby & R. C. Connor. Part IV. Social Minds: Empirical Perspectives. Chimpanzee social cognition, by M. Tomasello & J. Call; Intentional communication and comprehension of the partner’s role in experimental cooperative tasks, by S. Hirata, N. Morimura, & K. Fuwa; Collaboration and helping in chimpanzees, by A. P. Melis, F. Warneken, & B. Hare; Inequity and prosocial behavior in chimpanzees, by S. F. Brosnan; and The need for a bottom-up approach to chimpanzee cognition, by F. B. M. de Waal. Part V. Ethics, Care, and Conservation. How cognitive studies help shape our obligation for the ethical care of chimpanzees, by S. R. Ross; Positive reinforcement training, social learning, and chimpanzee welfare, by J. E. Perlman, V. Horner, M. A. Bloomsmith, S. P. Lambeth, & S. J. Shapiro; Chimpanzee orphans: Sanctuaries, reintroduction, and cognition, by B. B. Beck; Human-chimpanzee competition and conflict in Africa: A case study of coexistence in Bossou, Republic of Guinea, by K. J. Hockings; and Chimpanzee mind, behavior, and conservation, by E. V. Lonsdorf. Afterword: Meanings of chimpanzee mind, by R. Wrangham. Appendix: Major chimpanzee research sites.
* Multisensory Object Perception in the Primate Brain. M. J. Naumer & J. Kaiser (Eds.). New York: Springer, 2010.
* Neurobiology of Grooming Behavior. A. V. Kalueff, J. L. LaPorte, & C. L. Bergner (Eds.). Cambridge, U.K.: Cambridge University Press, 2010.
* The Noisy Brain: Stochastic Dynamics as a Principle of Brain Function. E. T. Rolls & G. Deco. Oxford, U.K.: Oxford University Press, 2010. 310 pp. [Price: Ł37.95]
* Quirks of Human Anatomy: An Evo-Devo Look at the Human Body. L. I. Held, Jr. Cambridge, U.K.: Cambridge University Press, 2009. 272 pp. [Price: Ł65.00]
* Saraguatos: Voces de la Selva. J. C. Serio Silva. Secretaría de Educación de Veracruz, 2009. <www.sev.gob.mx/servicios/publicaciones/serie_paradocencia/saraguatos.pdf>.
* Wild Mammals in Captivity: Principles and Techniques for Zoo Management, 2nd ed. D. G. Kleiman, K. V. Thompson, & C. Kirk Baer (Eds.). University of Chicago Press, 2010.
* American Journal of Primatology, 2010, 72[8], <onlinelibrary.wiley.com/doi/10.1002/ajp.v72:8/issuetoc>.
. . .Contents: Personality in nonhuman primates: A review and evaluation of past research, by H. D. Freeman & S. D. Gosling; Ovarian reserve tests and their utility in predicting response to controlled ovarian stimulation in rhesus monkeys, by J. M. Wu, D. L. Takahashi, D. K. Ingram, J. A. Mattison, G. Roth, M. A. Ottinger, & M. B. Zelinski; Time course of vocal modulation during isolation in common marmosets (Callithrix jacchus), by C. Yamaguchi, A. Izumi, & K. Nakamura; A longitudinal study of urinary dipstick parameters in wild chimpanzees (Pan troglodytes verus) in Côte d’Ivoire, by S. A. J. Leendertz, S. Metzger, E. Skjerve, T. Deschner, C. Boesch, J. Riedel, & F. H. Leendertz; Phylogenetic analysis of the fecal flora of the wild pygmy loris, by X. Bo, H. Zun-xi, W. Xiao-yan, G. Run-chi, T. Xiang-hua, M. Yue-lin, Y. Yun-Juan, S. Hui, & Z. Li-da; Flexibility in the use of requesting gestures in squirrel monkeys (Saimiri sciureus), by J. R. Anderson, H. Kuroshima, Y. Hattori, & K. Fujita; Phylogeography of the Angolan black and white colobus monkey, Colobus angolesnsis palliatus, in Kenya and Tanzania, by M. M. McDonald & H. Hamilton; Floating limb behaviors and self-biting are associated in laboratory monkeys, by K. L. Bentson, C. M. Crockett, K. L. Wahl, E. P. Runeson, R. U. Bellanca, G. H. Lee, J. P. Thom, H. B. Montgomery, M. H. Yi, J. G. S. McComas, & J. C. Ha; and Mating promiscuity and reproductive tactics in female black and gold howler monkeys (Alouatta caraya) inhabiting an island on the Parana river, Argentina, by M. M. Kowalewski & P. A. Garber.
* American Journal of Primatology, 2010, 72[11], <onlinelibrary.wiley.com/doi/10.1002/ajp.v72:11/issuetoc>.
. . .Contents: Testing extraction and storage parameters for a fecal hormone method, by D. J. Pappano, E. K. Roberts, & J. C. Beehner; Mated pairs of owl monkeys (Aotus nancymaae) exhibit sex differences in response to unfamiliar male and female conspecifics, by C. K. Wolovich, S. Evans, & S. M. Green; Ape behavior in two alternating environments: Comparing exhibit and short-term holding areas, by S. R. Ross, K. E. Wagner, S. J. Schapiro, & J. Hau; The ontogeny of handling hard-to-process food in wild brown capuchins (Cebus apella apella): Evidence from foraging on the fruit of Maximiliana maripa, by N. Gunst, J.-B. Leca, S. Boinski, & D. Fragaszy; The communicative content of the common marmoset phee call during antiphonal calling, by C. T., Miller, K. Mandel, & X. Wang; Effects of food, proximity, and kinship on social behavior in ringtailed lemurs, by G. C. Sbeglia, Z. Tang-Martinez, & R. W. Sussman; Nutrient transport within and between habitats through seed dispersal processes by woolly monkeys in north-western Amazonia, by P. R. Stevenson & D. C. Guzmán-Caro; Pattern of maternal circulating CRH in laboratory-housed squirrel and owl monkeys, by M. L. Power, L. E. Williams, S. V. Gibson, J. Schulkin, J. Helfers, & E. P. Zorrilla; Variation in physiological health of diademed sifakas across intact and fragmented forest at Tsinjoarivo, eastern Madagascar, by M. T. Irwin, R. E. Junge, J.-L. Raharison, & K. E. Samonds; and Variation in dental wear and tooth loss among known-aged, older ring-tailed lemurs (Lemur catta): A comparison between wild and captive individuals, by F. P. Cuozzo, M. L. Sauther, L. Gould, R. W. Sussman, L. M. Villers, & C. Lent.
* American Journal of Primatology, 2010, 72[12], <onlinelibrary.wiley.com/doi/10.1002/ajp.v72:12/issuetoc>.
. . .Contents: Hunting strategies in wild common marmosets are prey and age dependent, by N. Schiel, A. Souto, L. Huber, & B. M. Bezerra; Phylogeny and distribution of crested gibbons (genus Nomascus) based on mitochondrial cytochrome b gene sequence data, by Van Ngoc Thinh, B. Rawson, C. Hallam, M. Kenyon, T. Nadler, L. Walter, & C. Roos; The use of new world primates for biomedical research: An overview of the last four decades, by L. Brito Torres, B. H. Silva Araujo, P. H. Gomes de Castro, F. Romero Cabral, K. Sarges Marruaz, M. Silva Araujo, S. Gomes da Silva, J. A. Pereira Carneiro Muniz, & E. Abrăo Cavalheiro; Does nonnutritive tree gouging in a rainforest-dwelling lemur convey resource ownership as does loud calling in a dry forest-dwelling lemur? by S. Rasoloharijaona, B. Randrianambinina, & M. Joly-Radko; Reproductive characteristics of wild female Phayre’s leaf monkeys, by A. Lu, C. Borries, N. M. Czekala, & J. C. Beehner; Specific gravity and creatinine as corrections for variation in urine concentration in humans, gorillas, and woolly monkeys, by B. C. White, K. M. Jamison, C. Grieb, D. Lally, C. Luckett, K. S. Kramer, & J. Phillips; Nocturnal sleeping habits of the Yunnan snub-nosed monkey in Xiangguqing, China, by D. Li, B. Ren, C. C. Grueter, B. Li, & M. Li; Line transect surveying of arboreal monkeys: Problems of group size and spread in a highly fragmented landscape, by S. F. Ferrari, R. R. D. Chagas, & J. Pedro Souza-Alves; Density and microhabitat use of Bengal slow loris in primary forest and non-native plantation forest, by M. Pliosungnoen, G. Gale, & T. Savini; Male dominance and reproductive success in wild white-faced capuchins (Cebus capucinus) at Lomas Barbudal, Costa Rica, by L. Muniz, S. Perry, J. H. Manson, H. Gilkenson, J. Gros-Louis, & L. Vigilant; and How many species of woolly monkeys inhabit Colombian forests? by S. Botero, L. Y. Rengifo, M. L. Bueno, & P. R. Stevenson.
* Animal Behaviour, 2010, 80[1], <www.sciencedirect.com/science/journal/00033472>.
. . .Contents include: The central importance of information in studies of animal communication, by R. M. Seyfarth, D. L. Cheney, T. Bergman, J. Fischer, K. Zuberbühler, & K. Hammerschmidt; Ranging patterns of hamadryas baboons: Random walk analyses, by A. L. Schreier & M. Grove; Message ‘scent’: Lemurs detect the genetic relatedness and quality of conspecifics via olfactory cues, by M. J. E. Charpentier, J. C. Crawford, M. Boulet, & C. M. Drea; What time is it? Coping with expected feeding time in capuchin monkeys, by E. Polizzi di Sorrentino, G. Schino, E. Visalberghi, & F. Aureli; and Spatial reference of black capuchin monkeys in Brazilian Atlantic Forest: Egocentric or allocentric? By A. Presotto & P. Izar.
* Animal Behaviour, 2010, 80[2], <www.sciencedirect.com/science/journal/00033472>.
. . .Contents include: Conflicts induce affiliative interactions among bystanders in a tolerant species of macaque (Macaca tonkeana), by A. De Marco, R. Cozzolino, F. Dessě-Fulgheri, & B. Thierry; How wild bearded capuchin monkeys select stones and nuts to minimize the number of strikes per nut cracked, by D. M. Fragaszy, R. Greenberg, E. Visalberghi, E. B. Ottoni, P. Izar, & Q. Liu; and Statistical analysis of animal observations and associated marks distributed in time using Ripley’s functions, by M. Marcoux, G. Larocque, M. Auger-Méthé, P. Dutilleul, & M. M. Humphries.
* Animal Behaviour, 2010, 80[3], <www.sciencedirect.com/science/journal/00033472>.
. . .Contents include: Vocal greeting behaviour in wild chimpanzee females, by M. N. C. Laporte & K. Zuberbühler; Does watching a monkey change its behaviour? Quantifying observer effects in habituated wild primates using automated radiotelemetry, by M. C. Crofoot, T. D. Lambert, R. Kays, & M. C. Wikelski; A note on reconstructing animal social networks from independent small-group observations, by C. Perreault; Recognizing and respecting claims over resources in free-ranging rhesus monkeys, Macaca mulatta, by B. E. Russ, J. A. Comins, R. Smith, & M. D. Hauser; and Repeated measures analysis of contests and other dyadic interactions: Problems of semantics, not statistical validity, by M. Briffa & R. W. Elwood.
* Animal Behaviour, 2010, 80[4], <www.sciencedirect.com/science/journal/00033472>.
. . .Contents include: Coalition formation among Barbary macaque males: The influence of scramble competition, by A. Berghänel, O. Schülke, & J. Ostner; and Should risk allocation strategies facilitate or hinder habituation to nonlethal disturbance in wildlife? By E. Sirot.
* Animal Behaviour, 2010, 80[5], <www.sciencedirect.com/science/journal/00033472>.
. . .Contents include: Predator-deterring alarm call sequences in Guereza colobus monkeys are meaningful to conspecifics, by A. M. Schel, A. Candiotti, & K. Zuberbühler; No evidence for ovarian synchrony or asynchrony in hamadryas baboons, by R. Tobler, S. Pledger, & W. Linklater; Chimpanzee helping in collaborative and noncollaborative contexts, by J. R. Greenberg, K. Hamann, F. Warneken, & M. Tomasello; Individual variation in temporal activity patterns in open-field tests, by P.-O. Montiglio, D. Garant, D. Thomas, & D. Réale; and Comparing samples with large numbers of zeros, by G. D. Ruxton, D. Rey, & M. Neuhäuser.
* Animal Behaviour, 2010, 80[6], <www.sciencedirect.com/science/journal/00033472>.
. . .Contents include: Inducing traditions in captive capuchin monkeys (Cebus apella), by J. Crast, J. M. Hardy, & D. Fragaszy; Men’s oxidative stress, fluctuating asymmetry and physical attractiveness, by S. W. Gangestad, L. A. Merriman, & M. E. Thompson; and A comparison of bonobo and chimpanzee tool use: Evidence for a female bias in the Pan lineage, by T. Gruber, Z. Clay, & K. Zuberbühler.
* Brain, Behavior and Evolution, 2010, 76[2], <content.karger.com/produktedb/produkte.asp?doi=320214>.
. . .Contents include: Comparative brain collections are an indispensable resource for evolutionary neurobiology, by A. N. Iwaniuk; A voxel-based morphometry analysis of white matter asymmetries in chimpanzees (Pan troglodytes), by W. D. Hopkins, J. P. Taglialatela, T. Nir, N. M. Schenker, & C. C. Sherwood; and Extensive changes in the expression of the opioid genes between humans and chimpanzees, by P. Cruz-Gordillo, O. Fedrigo, G. A. Wray, & C. C. Babbitt.
* Comparative Medicine, 2010, 60[5], <tinyurl.com/2ucxakf>.
. . .Contents include: Experimental induction of reduced ovarian reserve in a nonhuman primate model (Macaca fascicularis), by S. E. Appt, T. B. Clarkson, P. B. Hoyer, N. D. Kock, A. K. Goode, C. M. May, J. T. Persyn, N. K. Vail, K. F. Ethun, H. Chen, N. Sen, & J. R. Kaplan; Thoracic radiography as a refinement methodology for the study of H1N1 influenza in cynomologus macaques (Macaca fascicularis), by D. L. Brining, J. S. Mattoon, L. Kercher, R. A. LaCasse, D. Safronetz, H. Feldmann, & M. J. Parnell; and Radiographic incidence of spinal osteopathologies in captive rhesus monkeys (Macaca mulatta), by B. Hernández-Godínez, A. Ibáńez-Contreras, G. Perdigón-Castańeda, A. Galván-Montańo, G. García-Montes de Oca, C. Zapata-Valdez, & E. Tena-Betancourt.
* Folia Primatologica, 2010, 81[3], <www.karger.com/fpr>.
. . .Contents: Molecular systematics and phylogeography of the genus Lagothrix (Atelidae, Primates) by means of the mitochondrial COII gene, by M. Ruiz-Garcia & M. O. Pinedo-Castro; Feeding ecology, food availability and ranging patterns of wild Hamadryas baboons at Filoha, by A. L. Schreier; Evaluation of male inter-troop transfer as a mating strategy among ring-tailed lemurs on St. Catherines Island, USA, by J. A. Parga; and Play behavior of red colobus monkeys in Kibale National Park, Uganda, by E. A. Worch.
* IDA–Africa – Fall eNews, Oct 5, 2010, <www.ida-africa.org>.
* International Journal of Primatology, 2010, 31[5], <www.springerlink.com/content/104389>.
. . .Contents: Functions of intermittent locomotion in mustached tamarins (Saguinus mystax), by M. Stojan-Dolar & E. W. Heymann; Rondon’s marmoset, Mico rondoni sp. n., from southwestern Brazilian Amazonia, by S. F. Ferrari, L. Sena, M. P. C. Schneider, & J. S. Silva, Jr.; It’s all in the timing: Birth seasonality and infant survival in Eulemur rubriventer, by S. R. Tecot; Asymmetry and dimensions of relationship quality in the Japanese macaque (Macaca fuscata yakui), by B. Majolo, R. Ventura, & G. Schino; Peruvian red uakaris (Cacajao calvus ucayalii) are not flooded-forest specialists, by E. W. Heymann & R. Aquino; Vocal repertoire of golden-backed uakaris (Cacajao melanocephalus): Call structure and context, by B. Martins Bezerra, A. S. Souto, & G. Jones; Levels of intraspecific variation within the catarrhine skeleton, by L. T. Buck, J. T. Stock, & R. A. Foley; Effects of habitat structure and fragmentation on diversity and abundance of primates in tropical deciduous forests in Bolivia, by L. W. Pyritz, A. B. S. Büntge, S. K. Herzog, & M. Kessler; Genetic consequences of habitat fragmentation in black-and-gold howler (Alouatta caraya) populations from northern Argentina, by L. I. Oklander, M. M. Kowalewski, & D. Corach; Design and analysis of line transect surveys for primates, by S. T. Buckland, A. J. Plumptre, L. Thomas, & E. A. Rexstad; Food sharing in lion tamarins (Leontopithecus chrysomelas): Does foraging difficulty affect investment in young by breeders and helpers? by A. C. de A. Moura, H. G. Nunes, & A. Langguth; Exploring new areas: How important is long-term spatial memory for mangabey (Lophocebus albigena johnstonii) foraging efficiency? by K. R. L. Janmaat & R. L. Chancellor; Seasonal variations in the diet and feeding effort of two groups of howlers in different sized forest fragments, by J. C. Dunn, J. Cristóbal-Azkarate, & J. J. Veŕ; Abundance and morphology of Japanese mulberry trees in response to the distribution of Japanese macaques in snowy areas, by H. Enari & H. Sakamaki; and Gastrointestinal parasites of indigenous and introduced primate species of Rubondo Island National Park, Tanzania, by J. Petrášová, D. Modrý, M. A. Huffman, M. I. Mapua, L. Bobáková, V. Mazoch, J. Singh, T. Kaur, and K. J. Petrželková.
* IPPL News, September, 2010, 37[2], <www.ippl.org/news.php>. (Intl Primate Protection League, P.O. Box 766, Summerville, SC 29484 [e-mail: [email protected]]).
. . .Contents include: The colobus monkeys of Kenya; Meet Rosie, the newest IPPL gibbon; Remembering Marjorie Doggestt; and announcements and requests for help.
* Journal of the American Association for Laboratory Animal Science, 2010, 49[5], <tinyurl.com/24d8ze3>.
. . .Contents include: Measurement of fetal biparietal diameter in owl monkeys (Aotus nancymaae), by A. M. Schuler, A. G., Brady, G. W. Tustin, V. L. Parks, C. G. Morris, & C. R. Abee; Multidimensional cost-benefit analysis to guide evidence-based environmental enrichment: Providing bedding and foraging substrate to pen-housed monkeys, by A. J. Bennett, C. A. Corcoran, V. A. Hardy, L. R. Miller, & P. J. Pierre; Population genetic statistics from rhesus macaques (Macaca mulatta) in three different housing configurations at the California National Primate Research Center, by S. Kanthaswamy, A. Kou, & D. G. Smith; A comprehensive laboratory animal facility pandemic response plan, by G. S. Roble, N. M. Lingenhol, B. Baker, A. Wilkerson, & R. J. Tolwani; and Nonsurgical repair of a pseudoaneurysm in a cynomolgus macaque (Macaca fascicularis), by J. S. Daviau & D. A. Merton.
* Journal of the American Association for Laboratory Animal Science, 2010, 49[6], <tinyurl.com/2fljr3s>.
. . .Contents include: Pharmacokinetics of cefovecin in squirrel monkey (Saimiri sciureus), rhesus macaques (Macaca mulatta), and cynomolgus macaques (Macaca fascicularis), by R. Papp, A. Popovic, N. Kelly, & R. Tschirret-Guth; Generation of a specific-pathogen-free baboon colony, by R. F. Wolf, R. Eberle, & G. L. White; Recurrent idiopathic gingival enlargement in an olive baboon (Papio anubis), by K. Kolappaswamy, S. Shipley, M. A. Reynolds, C. McLeod, & L. Louis; Camptomelia in a rhesus macaque (Macaca mulatta), by K. Hopper, P. Morales, A. Garcia, & J. Wagner; Giant thoracic schwannoma in a rhesus macaque (Macaca mulatta), by D. A. Alves, T. M. Bell, C. Benton, E. J. Rushing, & E. L. Stevens; and Successful treatment of cryptosporidiosis in 2 common marmosets (Callithrix jacchus) by using paromomycin, by N. E. Hahn & S. V. Capuano, III.
* Journal of Medical Primatology, 2010, 39[5], <onlinelibrary.wiley.com/doi/10.1111/jmp.2010.39.issue-5/issuetoc>.
. . .An updated review of simian betaretrovirus (SRV) in macaque hosts, by N. A. Montiel; Epithelioid hemangioendothelioma in the right auricle of an adult, male rhesus macaque (Macaca mulatta), by E. D. Lombardini, R. Virmani, T. W. Blanchard, J. F. Lafond, S. Ménard, & M. Doré; Evaluation of miosis, behavior and cholinesterase inhibition from low-level, whole-body vapor exposure to soman in African green monkeys (Chlorocebus sabeus), by R. F. Genovese, B. J. Benton, J. L. Oubre, P. J. Fleming, E. M. Jakubowski, & R. J. Mioduszewski; Medical and surgical management of reproductive neoplasia in two western lowland gorillas (Gorilla gorilla gorilla), by E. M. Stringer, R. S. De Voe, F. Valea, S. Toma, G. Mulvaney, A. Pruitt, B. Troan, & M. R. Loomis; Tetraparesis resembling acute transverse myelitis in a captive chimpanzee (Pan troglodytes): Long-term care and recovery, by T. Miyabe-Nishiwaki, A. Kaneko, K. Nishiwaki, A. Watanabe, S. Watanabe, N. Maeda, K. Kumazaki, M. Morimoto, R. Hirokawa, J. Suzuki, Y. Ito, M. Hayashi, M. Tanaka, M. Tomonaga, & T. Matsuzawa; Leptin associations with age, weight, and sex among chimpanzees (Pan troglodytes), by R. G. Bribiescas & S. F. Anestis; and AIDS and optic neuritis in a rhesus monkey infected with the R5 clade C SHIV-1157ipd3N4, by A. García, N. B. Siddappa, Q. Li, A. T. Haase, K. Paul, F. Stroud, X. Zhang, J. A. Fountain, F. Villinger, F. J. Novembre, J. G. Else, W. E. Secor, & R. M. Ruprecht.
* Journal of Medical Primatology, 2010, 39[6], <onlinelibrary.wiley.com/doi/10.1111/jmp.2010.39.issue-6/issuetoc>.
. . .Estimates of milk constituents from lactating bonnet macaque (Macaca radiata) mothers between two and seven months post-partum, by M. L. Laudenslager, C. Natvig, H. Cantwell, M. C. Neville, & M. L. Reite; Frequency of the major histocompatibility complex Mamu-A*01 allele in experimental rhesus macaques in China, by W. Wang, Z. Cong, X. Liu, W. Tong, H. Qiao, H. Jiang, Q. Wei, & C. Qin; Urinary estrogens, progesterone, and LH changes during normal menstrual cycles of a captive female pileated gibbon (Hylobates pileatus), by Y. Aramaki, H. Oae, Y. Mouri, K. Takizawa, K. Nakanishi, Y. Hyoudou, M. Omasa, & H. Kusunoki; Splenic angioleiomyoma in an owl monkey (Aotus nancymae), by A. S. Gozalo, P. M. Zerfas, M. F. Starost, W. R. Elkins, & C. L. Clarke; Clinical chemistry and hematology values in a Caribbean population of African green monkeys, by S. Liddie, R. J. Goody, R. Valles, & M. S. Lawrence; Real-time telemetric monitoring in whole-body &sup60;Co gamma-photon irradiated rhesus macaques (Macaca mulatta), by C. A. Carrier, T. B. Elliott, & G. D. Ledney; Retrospective case-control study of hyperglycemia in group-housed, mature female cynomolgus macaques (Macaca fascicularis), by S. A. Bauer, K. E. Leslie, D. L. Pearl, J. Fournier, & P. V. Turner; Isolation, identification of Streptococcus pneumoniae from infected rhesus monkeys and control efficacy, by S. Zou, Q. Luo, Z. Chen, A. Cheng, M. Wang, D. Zhu, R. Jia, F. Liu, X. Chen, Y. Zhou, F. Bi, & Z. Yang.
* Lemur News: The Newsletter of the Madagascar Section of the IUCN/SSC Primate Specialist Group, December, 2009, No. 14, <dpz.eu/index.php?id=53>.
. . .Contents: Rapid lemur survey in northern Menabe, by M. Dammhahn, L. Schäffler, C. Fichtel, & P. M. Kappeler; Grazing lemurs: Exhibition of terrestrial feeding by the southern gentle lemur, Hapalemur meridionalis, in the Mandena littoral forest, southeast Madagascar, by T. M. Eppley & G. Donati; Note on lemurs of the Réserve Spéciale d’Ambohitantely, Madagascar, by J. M. Ralison; Population genetic parameter estimates for six populations of crowned lemurs, Eulemur coronatus (Gray, 1842), from northern Madagascar, by R. Ramanamahefa, S. M. McGuire, E. E. Louis, Jr., & R. A. Brenneman; Population genetic parameter estimates for five populations of Sanford’s lemur, Eulemur sanfordi (Archbold, 1932), from northern Madagascar, by R. Ramanamahefa, S. M. McGuire, E. E. Louis, Jr., & R. A. Brenneman; Re-introduction of diademed sifaka (Propithecus diadema) and black and white ruffed lemurs (Varecia variegata editorum) at Analamazaotra Special Reserve, Eastern Madagascar, by S. R. Day, R. E. A. F. Ramarokoto, B. D. Sitzmann, R. Randriamboahanginjatovo, H. Ramanankirija, V. R. A. Randrianindrina, G. Ravololonarivo, & E. E. Louis, Jr.; Distribution of a conservation-based activity book at two primary schools near Analamazaotra Special Reserve, Madagascar, by S. M. McGuire, B. D. Sitzmann, K. Herrington, S. R. Day, R. E. A. F. Ramarokoto, & E. E. Louis, Jr.; Population survey of the Greater bamboo lemur (Prolemur simus) at Kianjavato Classified Forest, by S. M. McGuire, C. A. Bailey, J.-N. Rakotonirina, L. G. Razanajatovo, J. F. Ranaivoarisoa, L. M. Kimmel, & E. E. Louis, Jr.; Impact du tourisme sur Prolemur simus ŕ Talatakely, dans le Parc National de Ranomafana, by J. A. Norosoarinaivo, C. Tan, L. Rabetafika, & D. Rakotondravony; Possible new Mirza taxon from the Fiherenana River valley, Atsimo-Andrefana Region, by C. J. Gardner & L. Jasper; Rapid primatological surveys of the Andringitra forest corridors: Direct observation of the greater bamboo lemur (Prolemur simus), by K. E. Delmore, M. F. Keller, E. E. Louis, Jr., & S. E. Johnson; Allomothering and allonursing behaviour in collared lemurs (Eulemur collaris), by K. Kesch; Costs of alarm calling: Lemur alarm calls attract fossas, by C. Fichtel; Involving local communities in conservation: An example involving blue-eyed black lemurs (Eulemur flavifrons), by M. S. N. Volampeno & C. T. Downs; Funding and training; and Theses completed.
* Primate Conservation, Inc. UPDATE, 2010, <www.primate.org/news.htm>, (1411 Shannock Rd, Charlestown, RI 02813-3726).
* Primates, 2010, 51[4], <www.springerlink.com/content/0032-8332/51/4>.
. . .Contents: Observed infanticides following a male immigration event in black howler monkeys, Alouatta pigra, at Palenque National Park, Mexico, by S. Van Belle, A. E. Kulp, R. Thiessen-Bock, M. García, & A. Estrada; Identification of non-taster Japanese macaques for a specific bitter taste, by N. Suzuki, T. Sugawara, A. Matsui, Y. Go, & H. Hirai; Theropithecus gelada distribution and variations related to taxonomy: History, challenges and implications for conservation, by S. Gippoliti; Aggression does not increase friendly contacts among bystanders in geladas (Theropithecus gelada), by A. Leone, M. Mignini, G. Mancini, and E. Palagi; Polyspecific associations of Cercopithecus campbelli and C. petaurista with C. diana: What are the costs and benefits? by P. J. Buzzard; Maternal care and infant development in Callimico goeldii and Callithrix jacchus, by A. C. Ross, L. M. Porter, M. L. Power, & V. Sodaro; Responses of golden-backed uakaris, Cacajao melanocephalus, to call playback: Implications for surveys in the flooded Igapó forest, by B. M. Bezerra, A. S. Souto, & G. Jones; Two-year-old children copy more reliably and more often than nonhuman great apes in multiple observational learning tasks, by C. Tennie, K. Greve, H. Gretscher, & J. Call; Host age, sex, and reproductive seasonality affect nematode parasitism in wild Japanese macaques, by A. J. J. MacIntosh, A. D. Hernandez, & M. A. Huffman; and The Primates 2010 Most-Cited Paper Award is conferred upon Dr. Satoshi Hirata.
* Research Saves, 2010, Issue 3, <www.fbresearch.org/WorkArea/DownloadAsset.aspx?id= 1067&libID=1074>.
* Zoo Biology, 2010, 29[4], <onlinelibrary.wiley.com/doi/10.1002/zoo.v29:4/issuetoc>.
. . .Contents include: Observational methods to measure behaviors of captive cotton-top tamarins (Saguinus oedipus), by L. N. Edwards, A. E. Sorkin, R. C. Rhodes, III, & K. H. Petersson; and Female reproductive parameters in the Javan gibbon (Hylobates moloch), by S. Hodgkiss, E. Thetford, C. D. Waitt, & V. Nijman.
* Zoo Biology, 2010, 29[5], <onlinelibrary.wiley.com/doi/10.1002/zoo.v29:5/issuetoc>.
. . .Contents include: Factors influencing weight changes in callitrichids at the Bronx Zoo, by E. Kaplan & N. Shelmidine; and The potential for microchip-automated technology to improve enrichment practices, by J. M. Hoy, P. J. Murray, & A. Tribe.
* Program and Abstracts of the 33rd Annual Meeting of the American Society of Primatologists, June 16–19, 2010. American Journal of Primatology, 2010, 72[S1], <onlinelibrary.wiley.com/doi/10.1002/ajp.v72:1%2B/issuetoc>.
* Special Section on Ethical Issues in Field Primatology. American Journal of Primatology, 2010, 72[9], <onlinelibrary.wiley.com/doi/10.1002/ajp.v72:9/issuetoc>.
. . .Contents: Field primatology of today: Current ethical issues, by K. C. MacKinnon & E. P. Riley; Ethical issues faced by field primatologists: Asking the relevant questions, by L. M. Fedigan; Long-term field studies: Positive impacts and unintended consequences, by K. B. Strier; Ethics commentary: Subjects of knowledge and control in field primatology, by N. M. Malone, A. Fuentes, & F. J. White; Putting the community back in community ecology and education: The role of field schools and private reserves in the ethical training of primatologists, by P. A. Garber, A. Molina, & R. L. Molina; Male mating tactics in spider monkeys: Sneaking to compete, by K. N. Gibson; Preliminary evidence of accumulation of stress during translocation in mantled howlers, by M. A. Socorro Aguilar-Cucurachi, P. A. D. Dias, A. Rangel-Negrín, R. Chavira, L. Boeck, & D. Canales-Espinosa; Estimates of heritability for reproductive traits in captive rhesus macaque females, by C. Gagliardi, K. P. Falkenstein, D. E. Franke, & H. M. Kubisch; and Feeding ecology of Bornean orangutans (Pongo pygmaeus morio) in Danum Valley, Sabah, Malaysia: A 3-year record including two mast fruitings, by T. Kanamori, N. Kuze, H. Bernard, T. P. Malim, & S. Kohshima.
* Special Issue on Ethnoprimatology. American Journal of Primatology, 2010, 27[10], <onlinelibrary.wiley.com/doi/10.1002/ajp.v72:10/issuetoc>.
. . .Contents: The ethnoprimatological approach in primatology, by A. Fuentes & K. J. Hockings; Macaques in farms and folklore: Exploring the human–nonhuman primate interface in Sulawesi, Indonesia, by E. P. Riley & N. E. C. Priston; Human–nonhuman primate interactions amongst Tikuna people: Perceptions and local initiatives for resource management in Amacayacu in the Colombian Amazon, by H. E. Parathian & A. M. Maldonado; Local attitudes and perceptions toward crop-raiding by orangutans (Pongo abelii) and other nonhuman primates in northern Sumatra, Indonesia, by G. Campbell-Smith, H. V. P. Simanjorang, N. Leader-Williams, & M. Linkie; Exploring cultural drivers for wildlife trade via an ethnoprimatological approach: A case study of slender and slow lorises (Loris and Nycticebus) in South and Southeast Asia, by K. A. I. Nekaris, C. R. Shepherd, C. R. Starr, & V. Nijman; Attacks on local persons by chimpanzees in Bossou, Republic of Guinea: Long-term perspectives, by K. J. Hockings, G. Yamakoshi, A. Kabasawa, & T. Matsuzawa; Behavioral responses of one western lowland gorilla (Gorilla gorilla gorilla) group at Bai Hokou, Central African Republic, to tourists, researchers and trackers, by M. Klailova, C. Hodgkinson, & P. C. Lee; Chimpanzee responses to researchers in a disturbed forest–farm mosaic at Bulindi, western Uganda, by M. R. McLennan & C. M. Hill; and Perceptions of nonhuman primates in human–wildlife conflict scenarios, by C. M. Hill & A. D. Webber.
* Animal Research. Chronicle of Higher Education, Nov. 9, 2010.
. . .Contents: Animal research: Why we need alternatives, by H. Ferdowsian; Animal research: Groupthink in both camps, by L. A. Hansen; and Animal Research: Activists’ wishful thinking, primitive reasoning, by E. Holland.
* One Health: The Intersection of Humans, Animals, and the Environment. ILAR Journal: Institute for Laboratory Animal Research, 2010, 51[3], <dels-old.nas.edu/ilar_n/ilarjournal/51_3/html>.
* Building Bridges Toward a New Understanding of Tarsier Diversity. M. Shekelle & S. Gursky (Eds.). International Journal of Primatology, 2010, 31[6], <www.springerlink.com/content/104389>.
. . .Contents: Why tarsiers? Why now? An introduction to the special edition on tarsiers, by M. Shekelle & S. Gursky; Progreditur ordinara saltando et retrorsum...Normally proceeds in a leaping fashion, and backwards..., by C. T. Niemitz; Vertical clinging and leaping revisited: Locomotion and habitat use in the western tarsier, Tarsius bancanus explored via loglinear modeling, by R. H. Crompton, M. L. Blanchard, S. Coward, R. McNeill Alexander, & S. K. Thorpe; Influence of orbit size on aspects of the tarsier postorbital septum, by D. G. Savakova & M. Dagosto; Overview of sensory systems of Tarsius, by P. Wong, C. E. Collins & J. H. Kaas; The skull of Tarsius: Functional morphology, eyeballs, and the nonpursuit predatory lifestyle, by A. L. Rosenberger; The rediscovery of Buffon’s tarsier, by C. Callou, J. Cuisin, & C. Groves; The genera and species of Tarsiidae, by C. Groves & M. Shekelle; Molecular phylogenetics and chronometrics of Tarsiidae based on 12S mtDNA haplotypes: Evidence for Miocene origins of crown tarsiers and numerous species within the Sulawesian clade, by M. Shekelle, R. Meier, I. Wahyu, Wirdateti, & N. Ting; Tarsius wallacei: A new tarsier species from central Sulawesi occupies a discontinuous range, by S. Merker, C. Driller, H. Dahruddin, Wirdateti, W. Sinaga, D. Perwitasari-Farajallah, & M. Shekelle; Geographical variation in duet songs of Sulawesi tarsiers: Evidence for new cryptic species in south and southeast Sulawesi, by J. A. Burton & A. Nietsch; Checkerboard patterns, interspecific competition, and extinction: Lessons from distribution patterns of tarsiers (Tarsius) and slow lorises (Nycticebus) in insular Southeast Asia, by V. Nijman & K. A. I. Nekaris; Intraspecific variation in the mating system of spectral tarsiers, by S. Gursky-Doyen; preliminary data on the behavior, ecology, and morphology of pygmy tarsiers (Tarsius pumilus), by N. Grow & S. Gursky-Doyen; and Strengths and weaknesses of a population viability analysis for Philippine tarsiers (Tarsius syrichta), by I. Neri-Arboleda.
* Connectivity-driven white matter scaling and folding in primate cerebral cortex. Herculano-Houzel, S., Mota, B., Wong, P., & Kaas, J. H. (J. H. K., Dept of Psych., Vanderbilt Univ., Nashville, TN 37204 [e-mail: jon.h.kaas @vanderbilt.edu]). Proceedings of the National Academy of Sciences, U.S.A. , 2010, 107, 19008-19013, <www.pnas.org/content/107/44/19008.full>.
. . . “Larger brains have an increasingly folded cerebral cortex whose white matter scales up faster than the gray matter. Here we analyze the cellular composition of the subcortical white matter in 11 primate species, including humans, and one Scandentia [tree shrew], and show that the mass of the white matter scales linearly across species with its number of nonneuronal cells, which is expected to be proportional to the total length of myelinated axons in the white matter. This result implies that the average axonal cross-section area in the white matter, a, does not scale significantly with the number of neurons in the gray matter, N. The surface area of the white matter increases with N0.87, not N1.0. Because this surface can be defined as the product of N, a, and the fraction n of cortical neurons connected through the white matter, we deduce that connectivity decreases in larger cerebral cortices as a slowly diminishing fraction of neurons (which varies with N-0.16 sends myelinated axons into the white matter. Decreased connectivity is compatible with previous suggestions that neurons in the cerebral cortex are connected as a small-world network and should slow down the increase in global conduction delay in cortices with larger numbers of neurons. Further, a simple model shows that connectivity and cortical folding are directly related across species. We offer a white matter-based mechanism to account for increased cortical folding across species, which we propose to be driven by connectivity-related tension in the white matter, pulling down on the gray matter.”
* Functional organization for color and orientation in macaque V4. Tanigawa, H., Lu, H. D., & Roe, A. W. (Dept of Psychology, Vanderbilt Univ., Nashville, TN 37203 [e-mail: [email protected]]). Nature Neuroscience, 2010, 13, 1542-1548, <www.nature.com/neuro/journal/v13/n12/full/nn.2676.html>.
. . . “Visual area V4 in the macaque monkey is a cortical area that is strongly involved in color and shape perception. However, fundamental questions about V4 are still debated. V4 was initially characterized as a color-processing area, but subsequent studies revealed that it contains a diverse complement of cells, including those with preference for color, orientation, disparity and higher-order feature preferences. This has led to disputes and uncertainty about the role of V4 in vision. Using intrinsic signal optical imaging methods in awake, behaving monkeys, we found that different feature preferences are functionally organized in V4. Optical images revealed that regions with preferential response to color were largely separate from orientation-selective regions. Our results help to resolve long-standing controversies regarding functional diversity and retinotopy in V4 and indicate the presence of spatially biased distribution of featural representation in V4 in the ventral visual pathway.”
* Separate value comparison and learning mechanisms in macaque medial and lateral orbitofrontal cortex. Noonan, M. P., Walton, M. E., Behrens, T. E. J., Sallet, J., Buckley, M. J., & Rushworth, M. F. S. (Dept of Exp. Psych., Univ. of Oxford, Oxford OX1 3UD, U.K. [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2010, 107, 20547-20552, <www.pnas.org/content/107/47/20547.full>.
. . . “Uncertainty about the function of orbitofrontal cortex (OFC) in guiding decision-making may be a result of its medial (mOFC) and lateral (lOFC) divisions having distinct functions. Here we test the hypothesis that the mOFC is more concerned with reward-guided decision making, in contrast with the lOFC’s role in reward-guided learning. Macaques performed three-armed bandit tasks and the effects of selective mOFC lesions were contrasted against lOFC lesions. First, we present analyses that make it possible to measure reward-credit assignment—a crucial component of reward-value learning—independently of the decisions animals make. The mOFC lesions do not lead to impairments in reward-credit assignment that are seen after lOFC lesions. Second, we examined how the reward values of choice options were compared. We present three analyses, one of which examines reward-guided decision making independently of reward-value learning. Lesions of the mOFC, but not the lOFC, disrupted reward-guided decision making. Impairments after mOFC lesions were a function of the multiple option contexts in which decisions were made. Contrary to axiomatic assumptions of decision theory, the mOFC-lesioned animals’ value comparisons were no longer independent of irrelevant alternatives.”
* PIB binding in aged primate brain: Enrichment of high-affinity sites in humans with Alzheimer’s disease. Rosen, R. F., Walker, L. C., & LeVine, H., III. (L. C. W., Div. of Neuroscience, Yerkes NPRC, Emory Univ., Atlanta, GA 30322 [e-mail: [email protected]]). Neurobiology of Aging, 2009, 32, 223-234, <www.ncbi.nlm.nih.gov/pubmed/19329226>.
. . . “Aged nonhuman primates accumulate large amounts of human-sequence amyloid β (Aβ) in the brain, yet they do not manifest the full phenotype of Alzheimer’s disease (AD). To assess the biophysical properties of Aβ that might govern its pathogenic potential in humans and nonhuman primates, we incubated the benzothiazole imaging agent Pittsburgh Compound B (PIB) with cortical tissue homogenates from normal aged humans, humans with AD, and from aged squirrel monkeys, rhesus monkeys, and chimpanzees with cerebral Aβ-amyloidosis. Relative to humans with AD, high-affinity PIB binding is markedly reduced in cortical extracts from aged nonhuman primates containing levels of insoluble Aβ similar to those in AD. The high-affinity binding of PIB may be selective for a pathologic, human-specific conformation of multimeric Aβ, and thus could be a useful experimental tool for clarifying the unique predisposition of humans to Alzheimer’s disease.”
* Transgenic rhesus monkeys produced by gene transfer into early-cleavage-stage embryos using a simian immunodeficiency virus-based vector. Niu, Y., Yu, Y., Bernat, A., Yang, S., He, X., Guo, X., Chen, D., Chen, Y., Ji, S., Si, W., Lv, Y., Tan, T., Wei, Q., Wang, H., Shi, L., Guan, J., Zhu, X., Afanassieff, M., Savatier, P., Zhang, K., Zhou, Q., & Ji, W. (W. J., Kunming Primate Res. Ctr, Chinese Acad. of Sciences, Kunming 650223, P. R. China [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2010, 107, 17663-17667, <www.pnas.org/content/107/41/17663.full>.
. . . “The development of transgenic technologies in monkeys is important for creating valuable animal models of human physiology so that the etiology of diseases can be studied and potential therapies for their amelioration may be developed. However, the efficiency of producing transgenic primate animals is presently very low, and there are few reports of success. We have developed an improved methodology for the production of transgenic rhesus monkeys, making use of a simian immunodeficiency virus (SIV)-based vector that encodes EGFP and a protocol for infection of early-cleavage-stage embryos. We show that infection does not alter embryo development. Moreover, the timing of infection, either before or during embryonic genome activation, has no observable effect on the level and stability of transgene expression. Of 70 embryos injected with concentrated virus at the one- to two-cell stage or the four- to eight-cell stage and showing fluorescence, 30 were transferred to surrogate mothers. One transgenic fetus was obtained from a fraternal triple pregnancy. Four infant monkeys were produced from four singleton pregnancies, of which two expressed EGFP throughout the whole body. These results demonstrate the usefulness of SIV-based lentiviral vectors for the generation of transgenic monkeys and improve the efficiency of transgenic technology in nonhuman primates.”
* Extensive spontaneous plasticity of corticospinal projections after primate spinal cord injury. Rosenzweig, E. S., Courtine, G., Jindrich, D. L., Brock, J. H., Ferguson, A. R., Strand, S. C., Nout, Y. S., Roy, R. R., Miller, D. M., Beattie, M. S., Havton, L. A., Bresnahan, J. C., Edgerton, V. R., & Tuszynskimtuszynski, M. H. (M. H. T., Dept of Neurosciences, U.C. San Diego, La Jolla, CA 92093 [e-mail: [email protected]]. Nature Neuroscience, 2010, 13, 1505-1510, <www.nature.com/neuro/journal/vaop/ncurrent/full/nn.2691.html>.
. . . “Although axonal regeneration after CNS injury is limited, partial injury is frequently accompanied by extensive functional recovery. To investigate mechanisms underlying spontaneous recovery after incomplete spinal cord injury, we administered C7 spinal cord hemisections to adult rhesus monkeys and analyzed behavioral, electrophysiological and anatomical adaptations. We found marked spontaneous plasticity of corticospinal projections, with reconstitution of fully 60% of pre-lesion axon density arising from sprouting of spinal cord midline-crossing axons. This extensive anatomical recovery was associated with improvement in coordinated muscle recruitment, hand function and locomotion. These findings identify what may be the most extensive natural recovery of mammalian axonal projections after nervous system injury observed to date, highlighting an important role for primate models in translational disease research.”
* Pool deemed positive for primates. AWI Quarterly, 2010, 59[3], 21, <www.awionline.org/ht/d/Contents/icids/24076/pid/24077>.
. . .Providing monkeys with “swimming pools” during the hot summer months is probably one of the most attractive environmental enrichments for them. Macaques and baboons are good swimmers and divers, but simply playing with water can fascinate primates for long periods of time.
* Refinement of the use of food and fluid control as motivational tools for macaques used in behavioural neuroscience research: Report of a working group of the NC3Rs. Prescott, M. J., Brown, V. J,, Flecknell, P. A., Gaffan, D., Garrod, K., Lemon, R. N., Parker, A. J., Ryder, K., Schultz, W., Scott, L., Watson, J., & Whitfield, L. (NC3Rs, 20 Park Crescent, London W1B 1AL, U.K. [e-mail: [email protected]]). Journal of Neuroscience Methods, 2010, 193, 167-188, <dx.doi.org/10.1016/j.jneumeth.2010.09.003>.
. . .This report provides practical guidance on refinement of the use of food and fluid control as motivational tools for macaques used in behavioral neuroscience research. The guidance is based on consideration of the scientific literature and, where data are lacking, expert opinion and professional experience, including that of the members of a Working Group convened by the United Kingdom National Centre for the Replacement, Refinement and Reduction of Animals in Research (NC3Rs). The report should be useful to researchers, veterinarians and animal care staff responsible for the welfare of macaques used in food and fluid control protocols, as well as those involved with designing, performing and analyzing studies that use these protocols. It should also assist regulatory authorities and members of local ethical review processes or institutional animal care and use committees concerned with evaluating such protocols. The report provides a framework for refinement that can be tailored to meet local requirements. It also identifies data gaps and areas for future research and sets out the Working Group’s recommendations on contemporary best practice.
* Temporal discounting and inter-temporal choice in rhesus monkeys. Hwang, J., Kim, S., & Lee, D. (D. L., Dept of Neurobiol., Yale Univ. Sch. of Med., 333 Cedar St, New Haven, CT 06510 [e-mail: daeyeol.lee@ yale.edu]). Frontiers in Behavioral Neuroscience, 2009, 3[9], <www.frontiersin.org/neuroscience/behavioralneuroscience/paper/10.3389/neuro.08/009.2009>.
. . . “Humans and animals are more likely to take an action leading to an immediate reward than actions with delayed rewards of similar magnitudes. Although such devaluation of delayed rewards has been almost universally described by hyperbolic discount functions, the rate of this temporal discounting varies substantially among different animal species. This might be in part due to the differences in how the information about reward is presented to decision makers. In previous animal studies, reward delays or magnitudes were gradually adjusted across trials, so the animals learned the properties of future rewards from the rewards they waited for and consumed previously. In contrast, verbal cues have been used commonly in human studies. In the present study, rhesus monkeys were trained in a novel inter-temporal choice task in which the magnitude and delay of reward were indicated symbolically using visual cues and varied randomly across trials. We found that monkeys could extract the information about reward delays from visual symbols regardless of the number of symbols used to indicate the delay. The rate of temporal discounting observed in the present study was comparable to the previous estimates in other mammals, and the animal’s choice behavior was largely consistent with hyperbolic discounting. Our results also suggest that the rate of temporal discounting might be influenced by contextual factors, such as the novelty of the task. The flexibility furnished by this new inter-temporal choice task might be useful for future neurobiological investigations on inter-temporal choice in nonhuman primates.”
* Case Report: Bilateral phacoemulsification in an orangutan (Pongo pygmaeus). Montiani-Ferreira, F., Lima, L., Bacellar, M., D’Otaviano Vilani, R. G., Fedullo, J. D., & Lange, R. R. (Universidade Federal do Paraná (UFPR), Rua dos Funcionários, 1540, 80035-050, Curitiba, PR, Brazil [e-mail: [email protected]]). Veterinary Ophthalmology, 2010, 13[Suppl. s1], 91-99, <onlinelibrary.wiley.com/doi/10.1111/j.1463-5224.2009.00733.x/full>.
. . .A 14-year-old, female, captive-born orangutan (Pongo pygmaeus) developed bilateral cataracts. Ultrasonography, electroretinography, and cataract correction using phacoemulsification were performed bilaterally. This case report aims to describe the ophthalmic procedures performed in this animal critically endangered of extinction. The surgery successfully restored vision and normal activity to the patient.
* Being Jane Goodall. D. Quammen. National Geographic, October, 2010, <ngm.nationalgeographic.com/2010/10/jane-goodall/quammen-text>.
. . .An overview and appreciation of Jane Goodall’s fifty years as a primatologist. It includes a photo gallery, <ngm.nationalgeographic.com/2010/10/jane-goodall/gombe-photography>, all photos from National Geographic articles by and about her, and articles <ngm.nationalgeographic.com/gombe-hub>.
* Island biogeography reveals the deep history of SIV. Worobey, M., Telfer, P., Souquičre, S., Hunter, M., Coleman, C. A., Metzger, M. J., Reed, P., Makuwa, M., Hearn, G., Honarvar, S., Roques, P., Apetrei, C., Kazanji, M., & Marx, P. A. (P. A. M., Tulane NPRC, Covington, LA 70433 [e-mail: [email protected]]). Science, 2010, 329, 1487, <www.sciencemag.org/cgi/content/full/sci;329/5998/1487>.
. . . “Simian immunodeficiency virus (SIV) lineages have been identified that are endemic to Bioko Island. The time the island formed offers a geological time scale calibration point for dating the most recent common ancestor of SIV. The Bioko viruses cover the whole range of SIV genetic diversity, and each Bioko SIV clade is most closely related to viruses circulating in hosts of the same genus on the African mainland rather than to SIVs of other Bioko species. Our phylogeographic approach establishes that SIV is ancient and at least 32,000 years old. Our conservative calibration point and analyses of gene sequence saturation and dating bias suggest it may be much older.”
* Origin of the human malaria parasite Plasmodium falciparum in gorillas. Liu, W., Li, Y., Learn, G. H., Rudicell, R. S., Robertson, J. D., Keele, B. F., Ndjango, J.-B. N., Sanz, C. M., Morgan, D. B., Locatelli, S., Gonder, M. K., Kranzusch, P. J., Walsh, P. D., Delaporte, E., Mpoudi-Ngole, E., Georgiev, A. V., Muller, M. N., Shaw, G. M., Peeters, M., Sharp, P. M., Rayner, J. C., & Hahn, B. H. (B. H. H., Dept of Med., Univ. of Alabama, Birmingham, AL 35294 [e-mail: [email protected]]). Nature, 2010, 467, 420-425, <www.nature.com/nature/journal/v467/n7314/full/nature09442.html>.
. . . “Plasmodium falciparum is the most prevalent and lethal of the malaria parasites infecting humans, yet the origin and evolutionary history of this important pathogen remain controversial. Here we develop a single-genome amplification strategy to identify and characterize Plasmodium spp. DNA sequences in fecal samples from wild-living apes. Among nearly 3,000 specimens collected from field sites throughout central Africa, we found Plasmodium infection in chimpanzees (Pan troglodytes) and western gorillas (Gorilla gorilla), but not in eastern gorillas (Gorilla beringei) or bonobos (Pan paniscus). Ape plasmodial infections were highly prevalent, widely distributed, and almost always made up of mixed parasite species. Analysis of more than 1,100 mitochondrial, apicoplast, and nuclear gene sequences from chimpanzees and gorillas revealed that 99% grouped within one of six host-specific lineages representing distinct Plasmodium species within the subgenus Laverania. One of these from western gorillas comprised parasites that were nearly identical to P. falciparum. In phylogenetic analyses of full-length mitochondrial sequences, human P. falciparum formed a monophyletic lineage within the gorilla parasite radiation. These findings indicate that P. falciparum is of gorilla origin and not of chimpanzee, bonobo, or ancient human origin.”
* Identification of a receptor for an extinct virus. Soll, S. J., Neil, S. J. D., & Bieniasz, P. D. (P. D. B., Aaron Diamond AIDS Research Ctr, Rockefeller Univ., New York, NY 10016 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2010, 107, 19496-19501, <www.pnas.org/content/107/45/19496.long>.
. . . “The resurrection of endogenous retroviruses from inactive molecular fossils has allowed the investigation of interactions between extinct pathogens and their hosts that occurred millions of years ago. Two such paleoviruses, chimpanzee endogenous retrovirus-1 and -2 (CERV1 and CERV2), are relatives of modern MLVs and are found in the genomes of a variety of Old World primates, but are absent from the human genome. No extant CERV1 and -2 proviruses are known to encode functional proteins. To investigate the host range restriction of these viruses, we attempted to reconstruct functional envelopes by generating consensus genes and proteins. CERV1 and -2 enveloped MLV particles infected cell lines from a range of mammalian species. Using CERV2 Env-pseudotyped MLV reporters, we identified copper transport protein 1 (CTR1) as a receptor that was presumably used by CERV2 during its ancient exogenous replication in primates. Expression of human CTR1 was sufficient to confer CERV2 permissiveness on otherwise resistant hamster cells, and CTR1 knockdown or CuCl2 treatment specifically inhibited CERV2 infection of human cells. Mutations in highly conserved CTR1 residues that have rendered hamster cells resistant to CERV2 include a unique deletion in a copper-binding motif. These CERV2 receptor-inactivating mutations in hamster CTR1 are accompanied by apparently compensating changes, including an increased number of extracellular copper-coordinating residues, and this may represent an evolutionary barrier to the acquisition of CERV2 resistance in primates.”
* Advanced antisense therapies for postexposure protection against lethal filovirus infections. Warren, T. K., Warfield, K. L., Wells, J., Swenson, D. L., Donner, K. S., Van Tongeren, S. A., Garza, N. L., Dong, L., Mourich, D. V., Crumley, S., Nichols, D. K., Iversen, P. L., & Bavari, S. (S. B., USAMRIID, Fort Detrick, MD 21702-9255 [e-mail: [email protected]]). Nature Medicine, 2010, 16, 991-994, <www.nature.com/nm/journal/v16/n9/full/nm.2202.html>.
. . . “Currently, no vaccines or therapeutics are licensed to counter Ebola or Marburg viruses, highly pathogenic filoviruses that are causative agents of viral hemorrhagic fever. Here we show that administration of positively charged phosphorodiamidate morpholino oligomers (PMOplus), delivered by various dosing strategies initiated 30–60 min after infection, protects >60% of rhesus monkeys against lethal Zaire Ebola virus (ZEBOV) and 100% of cynomolgus monkeys against Lake Victoria Marburg virus (MARV) infection. PMOplus may be useful for treating these and other highly pathogenic viruses in humans.”
* Cellular scaling rules for the brains of an extended number of primate species. Gabi, M., Collins, C. E., Wong, P., Torres, L. B., Kaas, J. H., & Herculano-Houzel, S. (S. H.-H., Inst. de Cięncias Bioméd., UFRJ, Av. Carlos Chagas Filho, 373, Ilha do Fundăo, Rio de Janeiro, RJ 21941-902, Brazil [e-mail: [email protected]]). Brain, Behavior and Evolution, 2010, 76, 32-44.
. . . “What are the rules relating the size of the brain and its structures to the number of cells that compose them and their average sizes? We have shown previously that the cerebral cortex, cerebellum and the remaining brain structures increase in size as a linear function of their numbers of neurons and non-neuronal cells across six species of primates. Here we describe that the cellular composition of the same brain structures of five other primate species, as well as humans, conform to the scaling rules identified previously, and that the updated power functions for the extended sample are similar to those determined earlier. Accounting for phylogenetic relatedness in the combined dataset does not affect the scaling slopes that apply to the cerebral cortex and cerebellum, but alters the slope for the remaining brain structures to a value that is similar to that observed in rodents, which raises the possibility that the neuronal scaling rules for these structures are shared among rodents and primates. The conformity of the new set of primate species to the previous rules strongly suggests that the cellular scaling rules we have identified apply to primates in general, including humans, and not only to particular subgroups of primate species. In contrast, the allometric rules relating body and brain size are highly sensitive to the particular species sampled, suggesting that brain size is neither determined by body size nor together with it, but is rather only loosely correlated with body size.”
* Cellular scaling rules for primate spinal cords. Burish, M. J., Peebles, J. K., Baldwin, M. K., Tavares, L., Kaas, J. H., & Herculano-Houzel, S. (address same as above). Brain, Behavior and Evolution, 2010, 76, 45-59.
. . . “The spinal cord can be considered a major sensorimotor interface between the body and the brain. How does the spinal cord scale with body and brain mass, and how are its numbers of neurons related to the number of neurons in the brain across species of different body and brain sizes? Here we determine the cellular composition of the spinal cord in eight primate species and find that its number of neurons varies as a linear function of cord length, and accompanies body mass raised to an exponent close to 1/3. This relationship suggests that the extension, mass and number of neurons that compose the spinal cord are related to body length, rather than to body mass or surface. Moreover, we show that although brain mass increases linearly with cord mass, the number of neurons in the brain increases with the number of neurons in the spinal cord raised to the power of 1.7. This faster addition of neurons to the brain than to the spinal cord is consistent with current views on how larger brains add complexity to the processing of environmental and somatic information.”
* Rhesus monkeys (Macaca mulatta) do recognize themselves in the mirror: Implications for the evolution of self-recognition. Rajala, A. Z., Reininger, K. R., Lancaster, K. M., & Populin, L. C. (Dept of Psych., Univ. of Wisconsin, Madison, WI 53706 [e-mail: [email protected]]). PLoS ONE, 2010, 5[9], e12865, <www.plosone.org/article/info:doi/10.1371/journal.pone.0012865>.
. . . “Self-recognition in front of a mirror is used as an indicator of self-awareness. Along with humans, some chimpanzees and orangutans have been shown to be self-aware using the mark test. Monkeys are conspicuously absent from this list because they fail the mark test and show persistent signs of social responses to mirrors despite prolonged exposure, which has been interpreted as evidence of a cognitive divide between hominoids and other species. In stark contrast with those reports, the rhesus monkeys in this study, who had been prepared for electrophysiological recordings with a head implant, showed consistent self-directed behaviors in front of the mirror and showed social responses that subsided quickly during the first experimental session. The self-directed behaviors, which were performed in front of the mirror and did not take place in its absence, included extensive observation of the implant and genital areas that cannot be observed directly without a mirror. We hypothesize that the head implant, a most salient mark, prompted the monkeys to overcome gaze aversion inhibition or lack of interest in order to look and examine themselves in front of the mirror. The results of this study demonstrate that rhesus monkeys do recognize themselves in the mirror and, therefore, have some form of self-awareness. Accordingly, instead of a cognitive divide, they support the notion of an evolutionary continuity of mental functions.”
* New Oligocene primate from Saudi Arabia and the divergence of apes and Old World monkeys. Zalmout, I. S., Sanders, W. J., MacLatchy, L. M., Gunnell, G. F., Al-Mufarreh, Y. A., Ali, M. A., Nasser, A.-A. H., Al-Masari, A. M., Al-Sobhi, S. A., Nadhra, A. O., Matari, A. H., Wilson, J. A., & Gingerich, P. D. (Museum of Paleontology, Univ. of Michigan, 1109 Geddes Ave, Ann Arbor, MI 48109 [e-mail: [email protected]]). Nature, 2010, 466, 360-364, <www.nature.com/nature/journal/v466/n7304/full/nature09094.html>. “It is widely understood that Hominoidea (apes and humans) and Cercopithecoidea (Old World monkeys) have a common ancestry as Catarrhini deeply rooted in Afro-Arabia. The oldest stem Catarrhini in the fossil record are Propliopithecoidea, known from the late Eocene to early Oligocene epochs (roughly 35–30 Myr ago) of Egypt, Oman and possibly Angola. Genome-based estimates for divergence of hominoids and cercopithecoids range into the early Oligocene; however, the mid-to-late Oligocene interval from 30 to 23 Myr ago has yielded little fossil evidence documenting the morphology of the last common ancestor of hominoids and cercopithecoids, the timing of their divergence, or the relationship of early stem and crown catarrhines. Here we describe the partial cranium of a new medium-sized (about 15–20 kg) fossil catarrhine, Saadanius hijazensis, dated to 29–28 Myr ago. Comparative anatomy and cladistic analysis shows that Saadanius is an advanced stem catarrhine close to the base of the hominoid–cercopithecoid clade. Saadanius is important for assessing competing hypotheses about the ancestral morphotype for crown catarrhines, early catarrhine phylogeny and the age of hominoid–cercopithecoid divergence. Saadanius has a tubular ectotympanic but lacks synapomorphies of either group of crown Catarrhini, and we infer that the hominoid–cercopithecoid split happened later, between 29–28 and 24 Myr ago.”
* Human-specific evolution and adaptation led to major qualitative differences in the variable receptors of human and chimpanzee natural killer cells. Abi-Rached, L., Moesta, A. K., Rajalingam, R., Guethlein, L. A., & Parham, P. (P. P., Dept of Structural Biol., Stanford Univ. Sch. of Med., Stanford, CA 94305-5101 [e-mail: [email protected]]). PLoS Genetics, 2010, 6[11], <www.plosgenetics.org/article/info:doi/10.1371/journal.pgen.1001192>.
. . . “Natural killer (NK) cells serve essential functions in immunity and reproduction. Diversifying these functions within individuals and populations are rapidly-evolving interactions between highly polymorphic major histocompatibility complex (MHC) class I ligands and variable NK cell receptors. Specific to simian primates is the family of Killer cell Immunoglobulin-like Receptors (KIR), which recognize MHC class I and associate with a range of human diseases. Because KIR have considerable species-specificity and are lacking from common animal models, we performed extensive comparison of the systems of KIR and MHC class I interaction in humans and chimpanzees. Although of similar complexity, they differ in genomic organization, gene content, and diversification mechanisms, mainly because of human-specific specialization in the KIR that recognizes the C1 and C2 epitopes of MHC-B and -C. Humans uniquely focused KIR recognition on MHC-C, while losing C1-bearing MHC-B. Reversing this trend, C1-bearing HLA-B46 was recently driven to unprecedented high frequency in Southeast Asia. Chimpanzees have a variety of ancient, avid, and predominantly inhibitory receptors, whereas human receptors are fewer, recently evolved, and combine avid inhibitory receptors with attenuated activating receptors. These differences accompany human-specific evolution of the A and B haplotypes that are under balancing selection and differentially function in defense and reproduction. Our study shows how the qualitative differences that distinguish the human and chimpanzee systems of KIR and MHC class I predominantly derive from adaptations on the human line in response to selective pressures placed on human NK cells by the competing needs of defense and reproduction.”
* Endocranial shape changes during growth in chimpanzees and humans: A morphometric analysis of unique and shared aspects. Neubauer, S., Gunz, P., & Hublin, J.-J. (P.G., Dept of Human Evol., Max Planck Inst. for Evolutionary Anthropology, Deutscher Platz 6, D-04103, Leipzig, Germany [e-mail: [email protected]]). Journal of Human Evolution, 2010, 59, 555-566, <linkinghub.elsevier.com/retrieve/pii/S0047248410001442>.
. . . “Compared to our closest living and extinct relatives, humans have a large, specialized, and complex brain embedded in a uniquely shaped braincase. Here, we quantitatively compare endocranial shape changes during ontogeny in humans and chimpanzees. Identifying shared and unique aspects in developmental patterns of these two species can help us to understand brain evolution in the hominin lineage. Using CT scans of 58 humans and 60 chimpanzees varying in age from birth to adulthood, we generated virtual endocasts to measure and analyze 29 three-dimensional endocranial landmarks and several hundred semilandmarks on curves and the endocranial surface; these data were then analyzed using geometric morphometric methods. The ontogenetic shape trajectories are nonlinear for both species, which indicates several developmental phases. Endocranial shape is already distinct at birth and there is no overlap between the two species throughout ontogeny. While some aspects of the pattern of endocranial shape change are shared between humans and chimpanzees, the shape trajectories differ substantially directly after birth until the eruption of the deciduous dentition: in humans but not in chimpanzees, the parietal and cerebellar regions expand relatively (contributing to neurocranial globularity) and the cranial base flexes within the first postnatal year when brain growth rates are high. We show that the shape changes associated with this early ‘globularization phase’ are unique to humans and do not occur in chimpanzees before or after birth.”
* Brain development after birth differs between Neanderthals and modern humans. Gunz, P., Neubauer, S., Maureille, B., & Hublin, J.-J. (Address same as above). Current Biology, 2010, 20, R921-R922, <download.cell.com/current-biology/pdf/PIIS0960982210012820.pdf>.
. . . “Neanderthals had brain sizes comparable to modern humans, but their brain cases were elongated and not globular as in Homo sapiens. It has, therefore, been suggested that modern humans and Neanderthals reached large brain sizes along different evolutionary pathways. Here, we assess when during development these adult differences emerge. This is critical for understanding whether differences in the pattern of brain development might underlie potential cognitive differences between these two closely related groups. Previous comparisons of Neanderthal and modern human cranial development have shown that many morphological characteristics separating these two groups are already established at the time of birth, and that the subsequent developmental patterns of the face are similar, though not identical. Here, we show that a globularization phase seen in the neurocranial development of modern humans after birth is absent from Neanderthals.”
* Radiometric dating of the type-site for Homo heidelbergensis at Mauer, Germany. Wagner, G. A., Krbetschek, M., Degering, D., Bahain, J.-J., Shao, O., Falguéres, C., Voinchet, P., Dolo, J.-M., Garcia, T., & Rightmire, G. P. (Geographisches Inst., Univ. Heidelberg, D-69120 Heidel-berg, Germany [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2010, 107, 19726-19730, <www.pnas.org/content/107/46/19726.full>.
. . . “The Mauer mandible, holotype of Homo heidelbergensis, was found in 1907 in fluvial sands deposited by the Neckar River 10 km southeast of Heidelberg, Germany. The fossil is an important key to understanding early human occupation of Europe north of the Alps. Given the associated mammal fauna and the geological context, the find layer has been placed in the early Middle Pleistocene, but confirmatory chronometric evidence has hitherto been missing. Here we show that two independent techniques, the combined electron spin resonance/U-series method used with mammal teeth and infrared radiofluorescence applied to sand grains, date the type-site of Homo heidelbergensis at Mauer to 609 ± 40 ka. This result demonstrates that the mandible is the oldest hominin fossil reported to date from central and northern Europe and raises questions concerning the phyletic relationship of Homo heidelbergensis to more ancient populations documented from southern Europe and in Africa. We address the paleoanthropological significance of the Mauer jaw in light of this dating evidence.”
* Evolutionary relationships of wild hominids recapitulated by gut microbial communities. Ochman, H., Worobey, M., Kuo, C.-H., Ndjango, J.-B. N., Peeters, M., Hahn, B. H., & Hugenholtz, P. (Dept of Ecology & Evolutionary Biol., Yale Univ., West Haven, CT 06520 [e-mail: [email protected]]). PLoS Biology, 2010, 8[11], <www.plosbiology.org/article/info:doi/10.1371/journal.pbio.1000546>.
. . . “Multiple factors over the lifetime of an individual, including diet, geography, and physiologic state, will influence the microbial communities within the primate gut. To determine the source of variation in the composition of the microbiota within and among species, we investigated the distal gut microbial communities harbored by great apes, as present in fecal samples recovered within their native ranges. We found that the branching order of host-species phylogenies based on the composition of these microbial communities is completely congruent with the known relationships of the hosts. Although the gut is initially and continuously seeded by bacteria that are acquired from external sources, we establish that over evolutionary timescales, the composition of the gut microbiota among great ape species is phylogenetically conserved and has diverged in a manner consistent with vertical inheritance.”
* First skull of Antillothrix bernensis, an extinct relict monkey from the Dominican Republic. Rosenberger, A. L., Cooke, S. B., Rimoli, R., Ni, X., & Cardoso, L. (Dept of Anthro. & Archaeol., Brooklyn Coll., CUNY, New York, NY [e-mail: [email protected]]). Proceedings of the Royal Society, B, 2011, 278, 67-74, <rspb.royalsocietypublishing.org/content/278/1702/67>.
. . .The nearly pristine remains of Antillothrix bernensis, a capuchin-sized (Cebus) extinct platyrrhine from the Dominican Republic, have been found submerged in an underwater cave. This represents the first specimen of an extinct Caribbean primate with diagnostic craniodental and skeletal parts in association, only the second example of a skull from the region, and one of the most complete specimens of a fossil platyrrhine cranium yet discovered. Cranially, it closely resembles living cebines but is more conservative. Dentally, it is less bunodont and more primitive than Cebus, with crowns resembling Saimiri (squirrel monkeys) and one of the oldest definitive cebines, the late Early Miocene Killikaike blakei from Argentina. The tricuspid second molar also resembles the enigmatic marmosets and tamarins, whose origins continue to present a major gap in knowledge of primate evolution. While the femur is oddly short and stout, the ulna, though more robust, compares well with Cebus. As a member of the cebid clade, Antillothrix demonstrates that insular Caribbean monkeys are not monophyletically related and may not be the product of a single colonizing event. Antillothrix bernensis is an intriguing mosaic whose primitive characters are consistent with an early origin, possibly antedating the assembly of the modern primate fauna in greater Amazonia during the La Venta horizon. While most Greater Antillean primate specimens are quite young geologically, this vanished radiation, known from Cuba (Paralouatta) and Jamaica (Xenothrix) as well as Hispaniola, appears to be composed of long-lived lineages like several other mainland clades.
* Chronology of the Grotte du Renne (France) and implications for the context of ornaments and human remains within the Châtelperronian. Higham, T., Jacobi, R., Julien, M., David, F., Basell, L., Wood, R., Davies, W., & Ramsey, C. B. (Res. Lab. for Archaeology & History of Art, Univ. of Oxford, Oxford OX1 3QY, U.K. [e-mail: thomas [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2010, 107, 20234-20239, <www.pnas.org/content/107/47/20234.full>.
. . .There is extensive debate concerning the cognitive and behavioral adaptation of Neanderthals, especially in the period when the earliest anatomically modern humans dispersed into Western Europe, around 35,000–40,000 B.P. The site of the Grotte du Renne (at Arcy-sur-Cure) is of great importance because it provides the most persuasive evidence for behavioral complexity among Neanderthals. A range of ornaments and tools usually associated with modern human industries, such as the Aurignacian, were excavated from three of the Châtelperronian levels at the site, along with Neanderthal fossil remains (mainly teeth). This extremely rare occurrence has been taken to suggest that Neanderthals were the creators of these items. Whether Neanderthals independently achieved this level of behavioral complexity and whether this was culturally transmitted or mimicked via incoming modern humans has been contentious. At the heart of this discussion lies an assumption regarding the integrity of the excavated remains. One means of testing this is by radiocarbon dating; however, until recently, our ability to generate both accurate and precise results for this period has been compromised. A series of 31 accelerator mass spectrometry ultrafiltered dates on bones, antlers, artifacts, and teeth from six key archaeological levels shows an unexpected degree of variation. This suggests that some mixing of material may have occurred, which implies a more complex depositional history at the site and makes it difficult to be confident about the association of artifacts with human remains in the Châtelperronian levels.
* Survey of live laboratory animals reared in Japan (2009). Yagami, K., Mashimo, T., Sekiguchi, F., Sugiyama, F., Yamamura, K., & Serikawa, T. (LARC, Univ. of Tsukuba, 1-1-1 Tennodai, Tsukuba, Ibaraki 305-8575, Japan). Experimental Animals, 2010, 59, 531-535, <www.jstage.jst.go.jp/article/expanim/59/4/59_531/_article>.
. . .A survey on the number of live laboratory animals reared in research institutions, including universities, biomedical institutes, testing laboratories, pharmaceutical companies, and animal breeders, was conducted on June 1, 2009. One thousand seventy-four replies were recovered from 1,593 institutions, and 471 of them had used live laboratory animals from June 1, 2008 to May 31, 2009. A total of 11,337,334 live laboratory animals were being maintained on June 1, 2009 in Japan. 11,416 (.1%) were nonhuman primates.
* Hypnotic effects and pharmacokinetics of a single bolus dose of propofol in Japanese macaques (Macaca fsucata fsucata[sic]). Miyabe-Nishiwaki, T., Masui, K., Kaneko, A., Nishiwaki, K., Shimbo, E., & Kanazawa, H. (K. M., Dept of Anaesthesiology, Natl Defense Med. College, Namiki 3-2, Tokorozawa, Saitama, 359-8513 Japan. [e-mail: [email protected]]). Veterinary Anaesthesia and Analgesia, 2010, 37, 501-510, <onlinelibrary.wiley.com/doi/10.1111/j.1467-2995.2010.00564.x/full>.
. . . “Four male macaques (5–6 years old, 8.0–11.2 kg), were restrained and 8 mg/kg of propofol was administered intravenously at 6 mg/kg/minute. Behavioral changes without stimuli (first experiment), then responses to external stimuli (second experiment) were assessed every 2 minutes for 20 minutes. Venous blood samples were collected before and at 1, 5, 15, 30, 60, 120 and 210 minutes after drug administration, and plasma concentrations of propofol were measured (third experiment). Pharmacokinetic modelling was performed using NONMEM VI. The macaques were recumbent without voluntary movement for a mean 14.0 ± 2.7 SD (range 10.5–16.2) or 10.0 ± 3.4 (7.2–14.5) minutes and recovered to behave as pre-administration by 25.1 ± 3.6 (22.1–30.1) or 22.2 ± 1.5 (21.1–24.3) minutes after the end of propofol administration without or with stimuli, respectively. Respiratory and heart rates were stable throughout the experiments (28–68 breaths/minute and 72–144 beats/minute, respectively). Our final pharmacokinetic model included three compartments and well described the plasma concentration of propofol. The population pharmacokinetic parameters were: V1 = 10.4 L, V2 = 8.38 L, V3 = 72.7 L, CL1 = 0.442 L/minute, CL2 = 1.14 L/minute, CL3 = 0.313 L/minute, (the volumes of distribution and the clearances for the central, rapid and slow peripheral compartments, respectively). We conclude that intravenous administration of propofol (8 mg/kg) at 6 mg/kg/minute to Japanese macaques had a hypnotic effect lasting more than 7 minutes. A three-compartment model described propofol plasma concentrations over more than 3 hours. The developed pharmacokinetic parameters may enable simulations of administration protocols to maintain adequate plasma concentration of propofol.”
-------------------------------------------
We would like to acknowledge Primate-Science as a source for information about new books.
-------------------------------------------
Contents of Volumes 46-48 (2007-2009)
Contents of Volume 49-50 (2010-2011)
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, CLPS Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; fax: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
Current and back issues of the Newsletter are available on the World Wide Web at
http://www.brown.edu/primate
The Newsletter is supported by Brown University
Cover photograph of a bonobo (Pan paniscus), three year old Vic, visiting with his friend, a three year old boy in red. Taken at the Cincinnati Zoo and Botanical Gardens by Marian Brickner, June 2, 2006.
Copyright (c) 2010 by Brown University
Last updated: December 17, 2010