 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 50 NUMBER 2 APRIL 2011
Printable (PDF) Version of this issue
Male Monkey Murder During Intergroup Interaction in Free-Ranging Hanuman Langurs, Semnopithecus entellus, Around Jodhpur (Rajasthan), by G. Sharma, C. Ram, P. Vijay, and L. S. Rajpurohit ......1
News, Information, and Announcements
Workshop Announcements......7
. . .
Lab Animal Medicine – North Carolina; Forum on European Union Housing – New Jersey
Statistics on Primate Importation into the United States in 2010, by S. McGreal......8
Call for Award Nominations......9
. . .
Nominations for the 2011 Russell and Burch Award; Send a Technician to AALAS Meeting in San Diego; Fyssen Foundation International Prize
News Briefs......10
. . .
Researchers Say Stem Cells Helped Paralyzed Monkey; New Circular Protects Experimental Animals in Israel; World Week for Animals in Labs; Scientists Recreate Heart Defect in a Lab Dish
Meeting Announcements......11
Information Available: Interesting Websites......11
Educational Opportunities......12
. . .
Online Master of Laboratory Animal Science Degree; Residency in Lab Animal and Comparative Medicine; Lab Animal Science and Pathology Courses; Medical and Experimental Mammalian Genetics; Approaches to Complex Disorders
Research and Conservation Opportunities: Brazilian Atlantic Forest......13
Resource Wanted: “Neck-Bite” Photo......14
Announcements from Publications ......15
. . .
Neotropical Primates; Journal of Evolutionary Biology Research; Bioinformatics and Sequence Analysis
Editor’s Announcement......26
Departments
Recent Books and Articles......16
* * *
Male Monkey Murder During Intergroup Interaction in Free-Ranging Hanuman Langurs, Semnopithecus entellus, Around Jodhpur (Rajasthan)
Goutam Sharma, Chena Ram, Prateek Vijay, and L. S. Rajpurohit
Animal Behaviour Unit, Department of Zoology, Jai Narain Vyas University, Jodhpur
Introduction
The langur groups usually have foraging and feeding grounds, provisioning places, water holes, resting sites, night lodgings, etc. These places are often shared by two or more neighboring groups because of overlapping home ranges (Mohnot, 1977). All-male bands have been observed to interact with several unimale bisexual groups within their extensive nomadic range. The langurs have also been observed interacting (grooming and playing) with cows, goats, and dogs. The interactions with all-male bands may increase conflict and aggression, leading to temporary changes such as inter-group infant kidnapping (Mohnot, 1980) or permanent social changes (Mohnot, 1971, Makwana, 1979).
Intensity of interactions between bisexual troops and all-male bands differs from location to location in this habitat. In some locations, such as the Mandore Garden, Daijar, Bhadershawer, and Bhimbharak, groups come in contact with one another daily because of the overlapping of considerable parts of their home ranges. In other locations, like Sursagar, Kailana, and Bijolai, contact is less frequent because of less overlapping. During artificial feeding two bisexual troops may feed together peacefully, but when a bisexual troop and an all-male band interact at such times, the interaction is more agonistic.
Intertroop interaction has been observed in many mammals including several primate species. Male replacement and infant killings in Hanuman langurs have been reported at several study sites in India and in Nepal: Dharwar (Sugiyama, 1965), Jodhpur (Mohnot, 1971; Makwana, 1979; Sommer & Mohnot, 1985; Agoramoorthy & Mohnot, 1988; Rajpurohit et al., 2003; Sharma et al., 2010), Mount Abu (Hrdy, 1974), Kanha (Newton, 1986), Tirunelveli (Ross, 1993), and Nepal (Borries, 1997). But in the present paper we describe an unsuccessful attempted takeover or male replacement. One male of an all-male band was killed by focal troop B-1’s resident male.
Resident males of bisexual troops show great aggressiveness, which is an important behavior in this species during intertroop interactions. Sometimes the result may be a great social change, periodically seen in the unimale bisexual troops: a resident may lose his residentship. Interactions between all-male bands are mostly mild and peaceful. When two groups meet, they either sit close to each other or separate without any engagement. The present paper describes the interaction between a bisexual troop and several members of an all-male band.
Interactions between bisexual troops and all-male bands are usually not peaceful: there is much excitement and aggressive interactions occur. The resident male of a bisexual group starts “whooping”, either at the sight of an all-male group or when males of the latter give “whoop” calls from a distance of 1 to 3 kilometers. Then the resident male leaps, jumps, and goes around the “preferred area” of his home range, goes from branch to branch, and thumps and shakes horizontal tree branches in excitement, interrupting with vocalizations such as “whoops”, canine grinding, barking, air-biting, and coughing. The attention of females is diverted toward these acts of their leader. At that time young male infants continuously produce screams which develop into a “chorus” when the resident male utters “whoops”; the older male infants run towards the intruding leader and may hit or bite him. Estrous females often present to the intruder males, which may lead to severe fights with the resident male, and even end in group fission. In the initial stages of intrusion, the intruder males follow the females and, if excited, threaten them and try to chase them toward their own band.
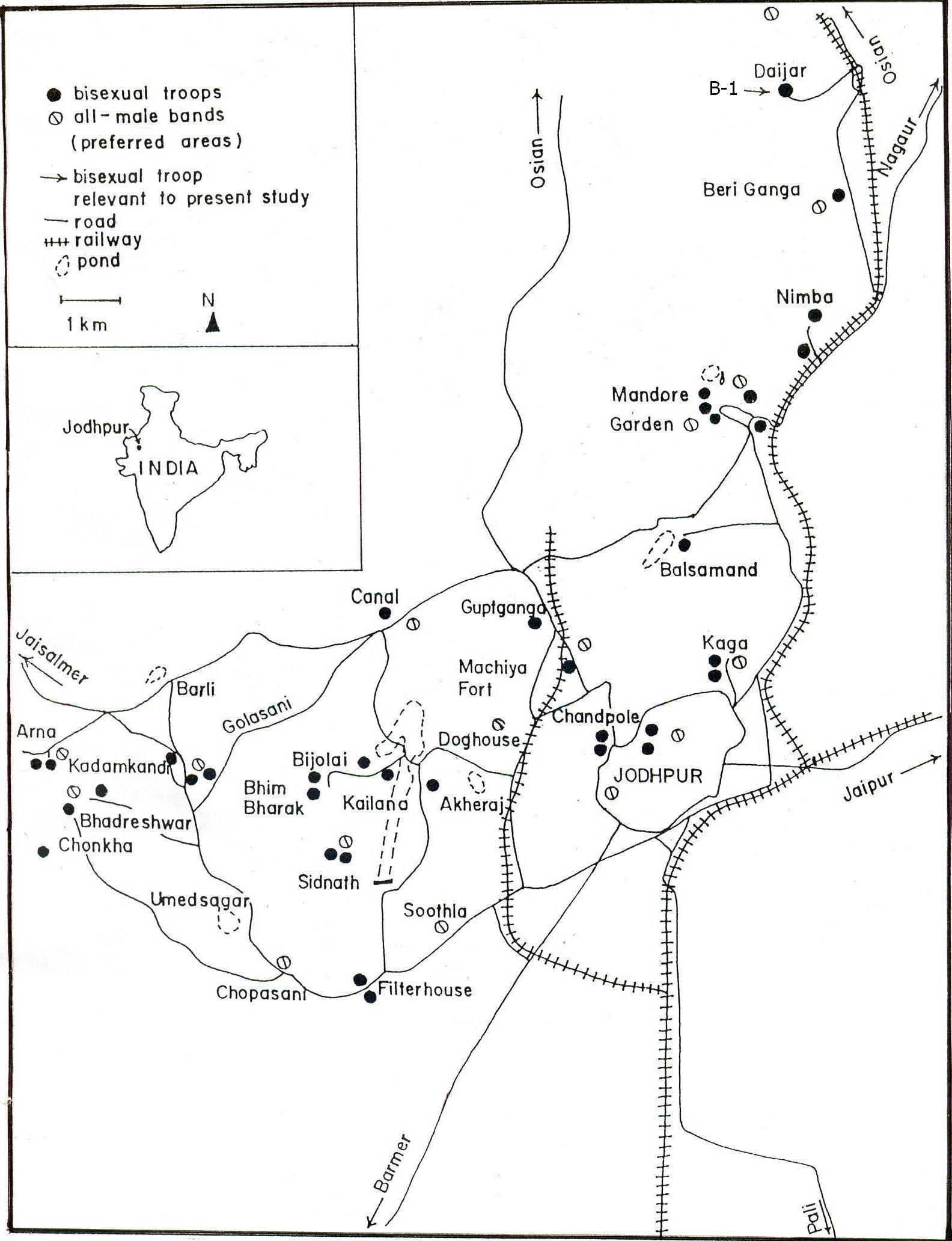
Figure 1: Location of bisexual troops and all-male bands around Jodhpur.
Bisexual troop B-1 lives on a hill at Daijar Mother Temple outside of Jodhpur city. It is about 15 km northwest of the city. The resident male of troop B-1 had been observed with the troop since 2009. In March, 2010, he was seen to be more aggressive. The troop size was 148, including the resident male, 75 adult females, 16 sub-adult females, and juveniles and infants. On April 8 four males of an all-male band attacked and tried to take over. They failed, and the alpha male of the all-male band was killed in the interaction.

Figure 2: Several members of bisexual troop B-1. Resident male is on the right.
The all-male band (AMB-1) had 25 males (18 adults and 7 sub-adults and juveniles), and was frequently seen near the bisexual troop. The present data, based on ad libitum samples (Altmann, 1974), were collected during 2010
. The responses of the focal group B-1, near Daijar, during encounters with other groups or lone males, were classified as ‘‘tolerant’’ (remains in proximity without displays), ‘‘avoiding’’ (moves quietly away from), or ‘‘aggressive’’ (vocal displays, chest-beating, and/or physical fighting).
On January 10, 2010, troop B-1, comprised of 148 langurs with one resident male, was looking normal. Common interactions, such as mild chasing, teeth grinding, and barking, were observed between troop B-1 and AMB-1. It is a daily routine, since their home ranges overlap. At feeding time, when they are provisioned by local people, it is easy to observe interactions between B-1 females and males of AMB-1. In the next several weeks, such mild interactions were observed and then, in March, the resident male became aggressive towards the AMB-1 males. The following account is based on field notes and discussion among the observers:
February 17, 7:30 a.m.: Normal, common interactions were observed between troop B-1 and AMB-1.
February 23, 6:50 a.m.: Troop B-1 seen on hill in resting position and appeared normal.
March 5, 6:00 a.m.: Troop B-1 appeared normal; no males other than resident male were seen around.
March 24, 7:00 a.m.: AMB-1 was seen interacting with troop B-1, but no fights were recorded.
March 25, 6:05 a.m.: Some AMB-1 males were sitting near the females of troop B-1 at feeding time. The females were trying to chase two males away from the feeding spot. At 6:35 a.m., the resident male looked restless and aggressive. Then he called “whoop” and ran towards the males of AMB-1 and chased them. At 7:00 a.m., the resident male was still restless, was on top of the hill and was teeth grinding and air-biting. In the next 5-7 minutes, the outside males went away. At 7:30 a.m., some males of AMB-1 came back to the home range of troop B-1; the resident started teeth grinding and air-biting, and tried to chase the intruders again. The AMB-1 males now moved a little farther away (about 200 m), but one of those males was still sitting close to the troop’s females. The resident showed aggressiveness towards that male and took a position for fighting. After a few minutes the resident male chased the other male away.
March 31, 6:30 a.m.: Troop B-1 was in the same position on the hill and some troop members were sitting on the ground waiting for feeding. In five minutes AMB-1 members appeared and started interacting with this troop. Then the resident male whooped and came down, starting teeth grinding. One male of AMB-1 was looking aggressive and was also continuously teeth grinding towards the resident male. Then the alpha male of AMB-1 jumped on the resident male and cut his leg. The resident male bit the neck of the alpha. Both males started bleeding and fighting. After 10 minutes, the resident male chased the alpha, who was injured and looking tired.
April 5: After this incident AMB-1, including the injured male, interacted peacefully with B-1. There was no objection from the resident male.
April 6: The alpha male of AMB-1 was observed with a severe fresh injury on his face, which indicated that there might have been a fight between the B-1’s resident male and the AMB-1 members. After this, AMB-1 kept their distance from B-1’s resident male.

Figure 3: Male killed in an encounter with troop B-1 (Daijar).
April 8, 8:00 a.m.: It was observed that the AMB-1 alpha male was dead near the temple, with a deep cut on his neck. The B-1 resident male was very aggressive towards AMB-1 members who were sitting near the dead monkey. He was continually teeth grinding towards them. It was presumed that the resident male had killed the alpha male of AMB-1. At 8:15 a.m. the resident male barked 5 or 6 times, whooped and jumped on a rock – and came near the male band. Then he sat down near them. AMB-1 were angry and teeth grinding. The resident was looking very aggressive. He was barking again and again and chasing members of AMB-1. Males of AMB-1 were also behaving aggressively towards the resident male, who tried to chase them, but they moved only a few steps. After 8 or 9 minutes, the resident male successfully chased them away.
During the study period, 13 encounters – 3 “tolerant”, 3 “avoiding”, and 7 “aggressive” – between the focal group and AMB-1 or lone males were recorded. Two encounters were with lone males, which occurred when lone males silently tracked B-1, occasionally for as long as two consecutive days. The location of encounters differed, depending upon whether they were with other groups or lone males. The context of encounters was not always clear (food? male/male competition?), although many occurred at or near the feeding spot.
The focal troop B-1’s response to lone males usually consisted of aggressive vocal displays or avoidance. Troop B-1 tolerated other groups many times, with or without physical contact.
Although the home range of the focal group overlapped with three other groups and several lone males, most of its home range was unused by other groups. High home-range overlap and limited site fidelity are characteristic of many primates in which female transfer occurs (Isbell & Van Vuren, 1996). Intergroup encounters may facilitate the transfer of migrants into nearby groups (Harcourt, 1978; Watts, 1990).
Generally Hanuman langurs’ intergroup encounters, which are typically aggressive, are related to the acquisition of females, rather than the defense of home ranges or food resources. The aggressive response of the resident male to outer males was also consistent with male–male competition. Sometimes, however, these encounters were related to access to food.
Although vocal displays were an important component of some intergroup encounters, contact aggression was notably less frequent than has been reported for mountain gorillas (Harcourt, 1978; Sicotte, 1993).
A “whoop” vocalization by adult males is believed to act as an effective spacing mechanism. In Dharwar (southern India), Sugiyama (1964, 1967), Sugiyama et al. (1965), and Yoshiba (1968) observed that group encounters occur daily and “whoop” vocalizations are used as a display against other bisexual groups or against all-male groups at a distance. When two groups come near each other, there is usually a fight between leader males, but such fights are not severe and are more like displays. All-male groups may attack bisexual groups for temporary association with estrous females, and this may sometimes result in permanent social changes.
On spotting a group of the opposite type, the dominant male usually gives “whoop” calls which are immediately reciprocated by the dominant male of the other group. From our observations, we believe that these calls serve as warning to the animals’ own groups, rather then as a spacing mechanism. If an oncoming all-male group decides to mount an attack, the resident male and others of the bisexual group keep up excited barking, coughing, and teeth-grinding.
Intergroup interactions in nonhuman primates range from peaceful mingling to violent clashes with fatal outcomes (reviewed by Cheney, 1987; and Fashing, 2001). Among primate species characterized by female philopatry, females are often aggressive participants in inter-group encounters (e.g., macaques [Macaca sp.], blue monkeys [Cercopithecus mitis], vervets [Chlorocebus aethiops], reviewed in Cheney, 1987), and several studies have suggested that defense of resources explains variation in female behavior (e.g. Fashing, 2001; and Wich et al., 2002; reviewed in Cheney, 1987).
Male baboons (Papio spp.) frequently chase female members of their own group away from rival groups (“herding,” Stoltz & Saayman, 1970; Buskirk et al., 1974; Henzi et al., 1998; see also Packer, 1979a) and are most likely to chase females when they are in estrus (Cheney & Seyfarth, 1977; Cowlishaw, 1995).
In primate species which form all-male groups, defense by the leader of the bisexual group against the violent appropriation and abduction of females by the all-male group may result in prolonged and severe fighting with skirmishes and raids lasting for days (e.g., chimpanzees [Pan troglodytes], Bygott, 1979; Hanuman langurs, Mohnot, 1971).
When home ranges overlap extensively, the aggressive defense of a particular resource may be more costly than the simple avoidance of other groups. In such cases, intergroup competition is often mediated by the relative dominance of the groups involved, as we saw with bisexual troop B-1.
In Sri Lanka (Ceylonese gray langur [Presbytis entellus], Ripley, 1967), aggressive group encounters occur regularly within the Ceylonese gray langur home range but do not usually result in severe fights. Such encounters take place inside the home range. Ripley, therefore, regards these intergroup fights as territorial, although the langurs seem to be defending group integrity in the sense of group space rather than topographical territory in the strict and commonly understood sense.
The “female defense” hypothesis predicts that male aggression during inter-group encounters should increase when sexually receptive females are present and that chasing of males by males should increase the distance between groups. Because high-ranking males have the greatest access to sexually receptive females (e.g. Packer, 1979b; Smith, 1986; Altmann et al., 1988, 1996; Bulger, 1993; Weingrill et al., 2000; Alberts et al., 2003), high-ranking males might be expected to be the most active in defending estrous females from extra-group males (Nunn, 2000; Nunn & Lewis, 2001).
Indeed, this is what we observed in the escalated encounter of bisexual troop B-1 with the all-male band, AMB-1.
Alberts, S. C., Watts, H. E., & Altmann, J. (2003). Queuing and queue-jumping: Long-term patterns of reproductive skew in male savannah baboons, Papio cynocephalus. Animal Behaviour, 65, 821-840.
Altmann, J. (1974). Observational study of behaviour: Sampling methods. Behaviour, 49, 227-267.
Altmann, J., Alberts, S. C., Haines, S. A., Dubach, J., Muruthi, P., Coote, T., Gefen, E., Cheesman, D. J., Mututua, R. S., Saiyalel, S. N., Wayne, R. K., Lacy, R. C., & Bruford, M. W. (1996). Behavior predicts genetic structure in a wild primate group. Proceedings of the National Academy of Sciences, U.S.A., 93, 5797-5801.
Altmann, J., Hausfater, G., & Altmann, S. A. (1988). Determinants of reproductive success in savannah baboons, Papio cynocephalus. In: T. H. Clutton-Brock (Ed.), Reproductive success: Studies of individual variation in contrasting breeding systems (pp. 403-418). Chicago, IL: University of Chicago Press.
Borries, C. (1997). Infanticide in seasonally breeding multimale groups of Hanuman langurs (Presbytis entellus) in Ramnagar (South Nepal). Behavioral Ecology and Sociobiology, 41, 139-150.
Bulger, J. (1993). Dominance rank and access to estrous females in male savannah baboons. Behaviour, 124, 89-122.
Buskirk, W. H., Buskirk, R. E., & Hamilton III, W. J. (1974). Troop-mobilizing behavior of adult male chacma baboons. Folia Primatologica, 22, 9-18.
Bygott, J. D. Agonistic behaviour, dominance and social structure in wild chimpanzees of the Gombe National Park (1979). In D. A. Hamburg & D. R. McCown (Eds.), The great apes (pp. 405-427). Menlo Park, CA: Benjamin/Cummings.
Cheney, D. L. (1987). Interactions and relationship between groups. In B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, & T. T. Struhsaker (Eds.), Primate societies (pp. 267-281). University of Chicago Press.
Cheney, D. L., & Seyfarth, R. M. (1977). Behavior of adult and immature male baboons during inter-group encounters. Nature, 269, 404-406.
Cowlishaw, G. (1995). Behavioural patterns in baboon group encounters: The role of resource competition and male reproductive tactics. Behaviour, 132, 75-86.
Fashing, P. J. (2001). Male and female strategies during intergroup encounters in guerezas (Colobus guereza): Evidence for resource defense mediated through males and a comparison with other primates. Behavioral Ecology and Sociobiology, 50, 219-230.
Harcourt, A. H. (1978). Strategies of emigration and transfer by primates, with particular reference to gorillas. Zeitschrift für Tierpsychologie, 48, 401-420.
Henzi, S. P., Lycett, J. E., & Weingrill, T. (1998). Mate guarding and risk assessment by male mountain baboons during inter-troop encounters. Animal Behaviour, 55, 1421-1428.
Hrdy, S. B. (1974). Male–male competition and infanticide among the langurs (Presbytis entellus) of Abu, Rajasthan. Folia Primatologica, 22, 19-58.
Isbell, L. A., & Van Vuren, D. (1996). Differential costs of locational and social dispersal and their consequences for female group-living primates. Behaviour, 133, 1-36.
Makwana, S. C. (1979). Infanticide and social change in two groups of the Hanuman langur, Presbytis entellus at Jodhpur. Primates, 20, 293-300.
Mohnot, S. M. (1971). Some aspects of social changes and infant-killing in the Hanuman langur, Presbytis entellus (Primates: Cercopithecidae) in Western India. Mammalia, 35, 175-198.
Mohnot, S. M. (1974). Ecology and behaviour of the common Indian langur, Presbytis entellus. PhD thesis, University of Jodhpur.
Mohnot, S. M. (1977). Observations and behaviour of the langur, Presbytis entellus in India. In M. L. Roonwal (Ed.), The natural resources of Rajasthan (pp. 499-503). Univ. of Jodhpur.
Mohnot, S. M. (1980). Intergroup infant kidnapping in Hanuman langur. Folia Primatologica, 34, 259-277.
Mohnot, S. M., Agoramoorthy, G., Rajpurohit, L. S., & Srivastava, A. (1987). Ecobehavioural studies of Hanuman langur, Presbytis entellus. Technical Report (1983-86) (pp. 1-89). MAB Project, Department of Environment, Govt. of India, New Delhi.
Newton, P. N. (1986). Infanticide in an undisturbed forest population of Hanuman langurs (Presbytis entellus). Animal Behaviour, 34, 785-789.
Nunn, C. L. (2000). Collective benefits, free-riders, and male extra-group conflicts. In P. M. Kappeler (Ed.), Primate males: Causes and consequences of variation in group composition (pp. 192-204). Cambridge: Cambridge University Press.
Nunn, C. L., & Lewis, R. J. (2001). Cooperation and collective action in animal behaviour. In R. Noë, J. A. R. A. M. van Hooff, & P. Hammerstein (Eds.), Economics in nature: Social dilemmas, mate choice and biological markets (pp. 42-66). Cambridge: Cambridge University Press.
Packer, C. (1979a). Group composition, population density and some sociobiological observation of Hanuman langurs (Presbytis entellus). Animal Behaviour, 27, 1-36.
Packer, C. (1979b). Male dominance and reproductive activity in Papio anubis. Animal Behaviour, 27, 37-45.
Rajpurohit, L. S. (1987). Male social organization in Hanuman langur, Presbytis entellus. PhD Thesis, University of Jodhpur.
Rajpurohit, L. S., Changani, A. K., Rajpurohit, R. S., & Mohnot, S. M. (2003). Observation of a sudden resident male replacement in a unimale bisexual troop of Hanuman langurs, Semnopithecus entellus, around Jodhpur (India). Folia Primatologica, 74, 85-87.
Rajpurohit, L. S., Sharma, G., Devilal, Vijay, P., Swami, B., & Ram, C. (2010). Recent survey of population and its composition in and around Jodhpur, Rajasthan (India). Proceedings of the 97th Indian Science Congress Held at Thiruvananthapuram, Kerala, in January (p.72).
Ripley, S. (1967). Inter-troop encounters among Ceylonese gray langurs (Presbytis entellus). In S. A. Altmann (Ed.), Social communication among primates (pp. 237-253). Chicago University Press.
Roonwal, M. L., & Mohnot, S. M. (Eds.). (1977). Primates of south Asia. Cambridge, MA: Harvard University Press.
Ross, C. (1993). Take-over and infanticide in South Indian Hanuman langurs (Presbytis entellus). American Journal of Primatology, 30, 75-82.
Sharma, G., Ram, C., & Rajpurohit, L. S. (2010). A case study of infanticide after resident male replacement in Semnopithecus entellus around Jodhpur (India). Laboratory Primate Newsletter, 49 [4], 6-11.
Sicotte P. (1993). Intergroup interactions and female transfer in mountain gorillas: In?uence of group composition on male behavior. American Journal of Primatology, 30, 21-36.
Smith, K. S. (1986). Dominance and mating strategies of chacma baboons, Papio ursinus, in the Okavango Delta, Botswana. PhD thesis, Davis: University of California.
Sommer, V., & Mohnot, S. M. (1985). New observations on infanticide among Hanuman langur (Presbytis entellus) near Jodhpur (Rajasthan, India). Behaviour, Ecology, and Sociobiology, 16, 245-248.
Stoltz, L. P., & Saayman, G. S. (1970). Ecology and behaviour of baboons in the northern Transvaal. Annals of the Transvaal Museum, 26, 99-143.
Sugiyama, Y. (1964). Group composition, population density and some sociobiological observations of Hanuman langurs (Presbytis entellus). Primates, 5, 7-37.
Sugiyama, Y. (1965). On the social change of Hanuman langurs (Presbytis entellus) in their natural conditions. Primates, 6, 381-417.
Sugiyama, Y. (1967). Social organization of Hanuman langurs. In S. A. Altmann (Ed.), Social communication among primates (pp. 221-236). Chicago University Press.
Sugiyama, Y., Yoshiba, K., & Parthasarathy, M. D. (1965). Home range, mating season, male group and intertroop relations in Hanuman langurs (Presbytis entellus). Primates, 6, 73-106.
Watts D. P. (1990). Ecology of gorillas and its relationship to female transfer in mountain gorillas. International Journal of Primatology, 11, 21-45.
Weingrill, T., Lycett, J. E., & Henzi, S. P. (2000). Consortship and mating success in chacma baboons (Papio cynocephalus ursinus). Ethology, 106, 1033-1044.
Wich, S. A., Assink, P. R., Becher, F., & Sterck, E. H. M. (2000). Playbacks of loud calls to wild Thomas langurs (Primates: Presbytis thomasi): The effect of location. Behaviour, 139, 65-78.
Wolfheim, J. H. (Ed.) (1983). Primates of the world: Distributions, abundance and conservation. Seattle: University of Washington Press.
Yoshiba, K. (1968). Local and inter-troop variability in ecology and social behaviour of common Indian langurs. In P. Jay (Ed.), Primates: Studies in adaptation and variability (pp. 217-242). New York: Holt, Rinehart & Winston.
-------------------------------------------
Author’s address: Dr. Goutam Sharma, Animal Behaviour Unit, Dept of Zoology, Jai Narain Vyas University, Jodhpur–342005 (Rajasthan), India
[e-mail: gautam_234746 @yahoo.co.in].
-------------------------------------------
* * *
Registration is open for the 2011 North Carolina Workshop in Laboratory Animal Medicine to be held at North Carolina State University’s College of Veterinary Medicine (NSCU-CVM), May 19–22. The course description and secure, online registration are available at <cvm.ncsu.edu/conted/WorkshopinLaboratoryAnimalMedicine.html>.
This annual four-day workshop was previously presented through the Charles Louis Davis, DVM Foundation and the North Carolina Academy of Laboratory Animal Medicine (NCALAM). Beginning this year, the workshop will be presented by NCALAM and the NCSU-CVM.
As in past years, the workshop will have lectures on pertinent topics in laboratory animal medicine on Thursday, May 19, and Friday, May 20. The Mock ACLAM exam coordinated by the International Mock Board Exam Coalition (see <labsg.org/2008Mock.html>) will be given on Saturday, May 22. The large collection of images from the National Institute of Environmental Health Sciences will be available for review in the NCSU-CVM library from May 19–22.
The meeting has been approved by the North Carolina Veterinary Medical Board for 13 hours of Continuing Education and will not be submitted for approval in the Registry of Approved Continuing Education (RACE) program. Attendees are recommended to petition their individual State boards for specific Continuing Education approval.
Forum on European Union Housing – New Jersey
The New Jersey Association for Biomedical Research (NJABR) will host an Informational Forum on “EU Housing: Practices, Perceptions, & Policies” April 15, 2011, from 8:30 a.m. to 4 p.m., at the Bridgewater Marriott, Bridgewater, New Jersey. This forum will examine
* * *
Statistics on Primate Importation into the United States in 2010
Shirley McGreal
International Primate Protection League
All data here is unedited material from the spread-sheet received from the United States Fish and Wildlife Service: “Total Primate Imports to the United States for 2010”.
Total monkeys imported: 22,948
| China | 14,303 |
| Mauritius | 3,156 |
| Cambodia | 2,400 |
| Vietnam | 1,716 |
| Indonesia | 514 |
| Saint Kitts & Nevis | 158 |
| Philippines | 125 |
| Israel | 120 |
| Unknown | 74 (no country listed on spread-sheet |
| Canada | 57 |
| Guyana | 55 |
Table 1: Source countries exporting more than 50 monkeys
| Macaca fascicularis | 20,728 (crab-eating macaque) |
| Macaca mulatta | 1,855 (rhesus macaque) |
| Chlorocebus aethiops | 130 (vervet monkey) |
| Macaca nemestrina | 100 (pig-tailed macaque) |
| Saimiri spp. | 45 (squirrel monkey) |
| Cebus apella | 33 (tufted capuchin) |
| Chlorocebus sabaeus | 28 (green monkey) |
Table 2: Species (numbers under 10 omitted)
| Los Angeles, California | 13,093 |
| Chicago, Illinois | 4,140 |
| New York, New York | 3,503 |
| Champlain, New York | 1,220 |
| Calais, Maine | 444 |
| Miami, FL | 217 |
| San Francisco, California | 200 |
| Buffalo, New York | 126 |
Table 3: Sorted by port of entry (numbers greater than 10)
| Covance Research Products Inc | 8,430 |
| Charles River Laboratories, (Research Models and Services) | 2,985 |
| SNBL-USA | 2,727 |
| Worldwide Primates | 1,867 |
| Charles River Laboratories (PG) | 1,720 |
| Charles River Laboratories (BRF) | 1,718 |
| Primate Products | 825 |
| Harlan Labs | 650 |
| Charles River Laboratories Preclinical Services | 600 |
| Buckshire Corporation | 570 |
| Valley Biosystems | 324 |
| Shared Enterprises | 250 |
| University (unidentified on table) | 100 |
| Alpha Genesis Inc. | 70 |
| Three Springs Scientific | 60 |
Table 4: Sorted by importers of more than 50 monkeys.
-------------------------------------------
Author’s address: P.O. Box 766, Summerville, SC 29484 [843-871-2280; fax: 843-871-7988; e-mail: [email protected]; Web: www.ippl.org].
Working to Protect All Primates Since 1973.
This material was sent to the primfocus e-mail list on January 29, 2011.
-------------------------------------------
* * *
The Humane Society of the United States (HSUS) presents the Russell and Burch Award to scientists who have made outstanding contributions to the advancement of alternative methods in the areas of biomedical research, testing, or higher education. Alternative methods – also known as the Three Rs – are methods that can replace or reduce the use of animals in specific procedures, or refine procedures so that animals experience less pain or suffering.
The award is named in honor of William Russell and Rex Burch, the scientists who formulated the Three Rs approach of replacement, reduction, and refinement. The award is a means of recognizing the important role that scientists play in limiting the use and suffering of animals in laboratories. Candidates for the award are judged on the scientific merit of their contribution to the alternatives field, as well as their impact on, and professional commitment to, this field. Applicants should have a history of laboratory work that is above reproach on humane grounds.
Send nominations by April 10, 2011, to <[email protected]>. No special forms are necessary. Persons nominating themselves should submit a cover letter explaining their suitability for the award, a curriculum vitae, and three published articles (preferably in PDF format) representative of their alternatives work. Persons nominating others should submit a letter explaining the nominee’s suitability for the award, and arrange to have supporting documents forwarded.
Recipients of the Russell and Burch Award are selected by past honorees. The award will be presented at the World Congresses on Alternatives and Animal Use in the Life Sciences, which will be held August 21–25, 2011, in Montreal, Canada; see <www.wc8.ccac.ca>. The honoree will receive a $5,000 award and a plaque.
For further information, contact Russell and Burch Award, Animal Research Issues Section, HSUS, 2100 L St, NW, Washington, DC, 20037 [301-258-0242; fax: 301-258-7760; e-mail: [email protected]].
Send a Technician to AALAS Meeting in San Diego
“A laboratory animal’s best friend is a healthy, caring Animal Technician.”
Lab Products, Inc., is pleased to announce the 8th Annual Lab Products’ Animal Technician Award Program. This program is intended to reward a deserving animal care technician from each of the eight American Association for Laboratory Animal Science (AALAS) Districts and Canada with the opportunity to attend his/her first National AALAS Meeting. The award will be limited to animal care personnel with at least one year of laboratory animal care experience, who have never attended a National AALAS Meeting. Recipients of the Lab Products’ Animal Technician Award will each receive an award recognition plaque; airfare, hotel, and registration for the National AALAS Meeting; one year membership in National AALAS; and $350 to cover incidentals while attending the AALAS Meeting. For more information write to Lab Products, Inc., P.O. Box 639, Seaford, DE 19973.
Award selection criteria:
Send nominations to: Attn: Awards Selection Committee, Lab Products, Inc., P.O. Box 639, Seaford, DE 19973 [800-526-0469; fax: 302-628-4309]. Nominations must be received by June 17, 2011. Award recipients will be notified by August 5, 2011.
Fyssen Foundation International Prize
The Fyssen Foundation’s general aim is to “encourage all forms of scientific enquiry into cognitive mechanisms, including thought and reasoning, which underlie animal and human behavior, their biological and cultural bases, and phylogenetic and ontogenetic development.”
An International Prize of 60,000 Euros is awarded annually to a scientist who has conducted distinguished research in the areas supported by the Foundation. The list of all recipients since 1980 is available at <www.fondation-fysson.org>.
The topic for the 2011 International Prize is “The Epigenetics of Cognition”. Nominations must be proposed by recognized scientists, and should include: a CV of the nominee; a list of publications; and a summary (four pages maximum) of the research work upon which the nomination is based. Nominations should be sent in 14 copies to the Secrétariat de la Fondation Fyssen, 194, rue de Rivoli, 7500l Paris, France. Deadline for receipt of nominations is October 31, 2011.
* * *
Japanese researchers said in December that they had used stem cells to restore partial mobility in a small monkey that had been paralyzed from the neck down by a spinal injury. “It is the world’s first case in which a small-size primate recovered from a spinal injury using stem cells,” Professor Hideyuki Okano of Tokyo’s Keio University told Agence France-Presse (AFP).
Okano’s research team, which earlier helped a mouse recover its mobility in a similar treatment, injected so-called induced pluripotent stem (iPS) cells into a paralyzed marmoset, he said. The team planted four types of genes into human skin cells to create the iPS cells, according to Kyodo News.
The injection was given on the ninth day after the injury, considered the most effective timing, and the monkey started to move its limbs again within two to three weeks, Okano said. “After six weeks, the animal had recovered to the level where it was jumping around,” he told AFP. “It was very close to the normal level. Its gripping strength on the forefeet also recovered to up to 80 percent.”
Okano called the research project a major stride to pave the way for a similar medical technique to be used on humans. Scientists say the use of human embryonic stem cells as a treatment for cancer and other diseases holds great promise, but the process has drawn fire from religious conservatives and others who oppose it.
Embryonic stem cell research is controversial because human embryos are destroyed in order to obtain the cells capable of developing into almost every tissue of the body. – From Agence France-Presse, December 8, 2010
New Circular Protects Experimental Animals in Israel
Two years have passed since the global Helsinki Declaration amendment protecting animals used in medical experimentation was passed. It has not been turned into an updated Israeli law, so Health Ministry Director-General Dr. Ronni Gamzu decided to introduce changes through a circular.
The previous rules going back to 1975 required all human medical experimentation to be preceded by experiments on animals. In 2000 the Helsinki Declaration added that animal experimentation may be carried out only “as appropriate”, meaning that only the minimum amount of animals be included to reach conclusions regarding medications or procedures for human experimentation. In 2008 it was further updated to say that consideration must be taken to protect the animals’ welfare; meaning – among other things – that they should not suffer unnecessarily.
As nothing was done to make the 2008 amendment law in Israel, Gamzu said he would add it as an “addition” to the previous law and released the stipulations to all relevant parties. Now, in every case of animal experimentation, only the number of animals needed to test efficacy and safety can be used, and the animals’ welfare must be protected.
Gamzu continued that, in each case, if simulations can effectively be used instead of animal testing, animals should not be experimented on. – By Judy Siegel-Itzkovich, Jerusalem Post, January 10, 2011
World Week for Animals in Labs
The dates for World Week for Animals in Labs (WWAIL) 2011 have been announced by Michael Budkie of Stop Animal Exploitation Now (SAEN). The event will run from April 16 to April 24, 2011, although no specific activities have been scheduled as yet. Watch <www.saenonline.org> for listings. – from Total E-Clips No. 2241, February 3, 2011
Scientists Recreate Heart Defect in a Lab Dish
Using skin cells taken from children with a rare heart defect, researchers have created beating heart cells in the lab with the same heart defect, allowing researchers to test new drugs in human cells instead of mice. – Reuters, February 9, 2011 – <news.yahoo.com/s/nm/20110209/hl_nm/us_heart_stemcells> * * *
* * *
The 20th annual meeting of the International Behavioral Neuroscience Society will be held May 24–29, in Steamboat Springs, Colorado. See <www.ibnshomepage.org/annualmtg11.htm>.
The Fourth Annual Enrichment Extravaganza, an environmental enrichment and behavioral management symposium, will be held on June 13, 2011, from 8:30 a.m. to 4 p.m., at the Atlantic City, New Jersey, Convention Center. The event is sponsored by the New Jersey Branch of the American Association for Laboratory Animal Science (AALAS), the New Jersey Association for Biomedical Research, and the Enrichment Record, and will feature guest speakers, vendors, and breakout sessions. Guest speakers will be: Mollie Bloomsmith, Yerkes National Primate Research Center; Lisa Leon, U.S. Army Research Institute of Environmental Medicine; and Greg Whittaker, Moody Gardens Aquarium. For more information and registration, see <www.njabr.org/content/extravaganza-registration> or contact Annette McCabe [908-228-2203; e-mail: [email protected]].
And the Delaware Valley, New Jersey, and Metro New York AALAS Branches invite you to the 2011 Tribranch Symposium, to be held at the same location, June 13–15. “Things Old; Things New, and Things to Review” is the Symposium theme. Topics will include sessions on emerging and re-emerging infectious disease; research animal care and use; planning for a plant closing; clean bedding dispensing systems; ingestible enrichment; a round table on personal protective equipment; and other subjects. Your abstracts for poster, technical, or scientific sessions are welcome. For more information, see <www.tribranch.org/ataglance11.html>.
A joint meeting of the International Ethological Conference (IEC) and the Animal Behavior Society (ABS) will be held July 25–30, at Indiana University, Bloomington, Indiana. See <www.indiana.edu/~behav11>.
The 45th Congress of the International Society for Applied Ethology will be held July 31– August 4, at the Hyatt Regency, Indianapolis, Indiana. See <www.applied-ethology.org/isaemeetings.htm>.
The Research Institute of Medical Primatology of the Russian Academy of Medical Sciences (IMP RAMS) will host its 2nd International Symposium, “Fundamental and Applied Aspects of Medical Primatology”, dedicated to the 90th anniversary of the birth of the world’s oldest primatologist, Boris Lapin. This symposium will take place August 7–11, 2011, in Sochi-Adler, Russia. For more information, see <www.primatologia.ru>.
The 34th Annual Meeting of the American Society of Primatologists will be held at the Hilton Austin, in Austin, Texas, September 16–19, 2011. See <www.asp.org/meetings>.
The 62nd AALAS National Meeting will be held October 2–6, 2011, in San Diego, California. See <nationalmeeting.aalas.org>.
The 29th Annual Nonhuman Primate Models for AIDS Meeting will be held October 25–28, 2011, in Seattle, Washington, sponsored by the Washington NPRC. See <www.wanprc.org/wp-content/misc/nhp2011_flier.pdf>.
An extraordinary joint meeting of the European Society for Evolutionary Biology and four North American societies (Society for the Study of Evolution, American Society of Naturalists, Society of Systematic Biologists, and Canadian Society for Ecology and Evolution) will be held in July, 2012, in Ottawa, Canada.
The International Primatological Society’s XXIV Congress will be held August 13–17, 2012 in Veracruz, Mexico. See <www.citrouv.edu.mx/ips2012>.
* * *
* * *
The Office of Professional Studies in the Health Sciences at Drexel University College of Medicine offers a three-year, 36-credit, Online Master of Laboratory Animal Science (Online MLAS) program. The program is designed specifically for individuals who are seeking career advancement in laboratory animal facility management, but are unable to physically relocate and attend the program in Philadelphia. The courses in the online program are substantially the same as those offered in our traditional program in Philadelphia, but allow maximum flexibility for the student. For the online program, the summer practicum is waived; instead, applicants must have a minimum of two years’ experience working in the field of laboratory animal science. The online program has a part-time curriculum that enables individuals to work and advance their education concurrently. Courses maximize the opportunities for learning from other students as well as from our highly experienced faculty.
Applications will be accepted until July 9th for the 2011–2012 academic year. Please visit <www.drexelmed.edu/Home/ForProspectiveStudents.aspx>; or contact Erin Vogelsong, Academic Administrator [215-762-7968; fax: 215.762.7961], for additional information.
Residency in Lab Animal and Comparative Medicine
The National Heart Lung and Blood Institute (NHLBI) and the Division of Veterinary Resources, parts of the National Institutes of Health (NIH), and the Department of Health and Human Services (HHS) seek applications from highly motivated individuals for residency training in laboratory animal and comparative medicine. This two-year program provides individuals with a strong foundation in laboratory animal medicine and in biomedical research and prepares them for board certification by the American College of Laboratory Animal Medicine (ACLAM). The training program is recognized by ACLAM; it takes advantage of a wealth of training opportunities at NIH and in the greater Washington, DC, area. Faculty and staff are drawn from several Institutes within NIH and include many ACLAM diplomates, veterinary pathologists, and surgeons. The program is designed to provide trainees with the didactic and experiential training necessary to prepare for a career in laboratory animal medicine and to obtain ACLAM certification. The program emphasizes the clinical, pathological, managerial, and administrative aspects of laboratory animal medicine. Trainee responsibilities include providing biomedical research support to investigators and research staff, providing clinical support for a wide variety of species, serving as liaison to Institutional Animal Care and Use Committees, conducting a research project, and preparing a manuscript.
Qualifications: DVM or equivalent degree, citizen or noncitizen national of the United States, graduation from an AVMA-accredited institution, and a license to practice veterinary medicine in at least one state in the U.S. Prior clinical or research experience is very desirable. Candidates with a strong interest in pursuing an academic career in laboratory animal medicine, research, and teaching are encouraged to apply. Preferred candidates will be highly motivated and have excellent communication and interpersonal skills. Veterinarians in private practice are encouraged to apply. Starting salary is based on the NIH scale for post-doctoral students. The start date is July 1, 2011. Applicants should submit: (1) a letter of intent that describes your interest in laboratory animal medicine, previous experiences related to the field, and career goals; (2) curriculum vitae; (3) complete transcripts from college(s) and veterinary school(s); (4) GRE scores (if available); and (5) three letters of recommendation to: Drs. Robert Hoyt [e-mail: [email protected]] and Charmaine Foltz [e-mail: [email protected]].
HHS and NIH are Equal Opportunity Employers. Applications from women, minorities, and persons with disabilities are strongly encouraged. The NHLBI/NIH is a smoke-free workplace.
Lab Animal Science and Pathology Courses
The 10th Annual Current Lab Animal Science Seminar (CLASS) and 54th Annual Pathology of Lab Animals Course (POLA) will be held June 5–7 (CLASS) and June 8–11 (POLA) at the Sheraton Virginia Beach Oceanfront Hotel, 3501 Atlantic Avenue, Virginia Beach, Virginia 23451. Attendees can participate in CLASS for $350, POLA for $475, or both for $700.
The schedule for June 5 includes: Welcome and Announcements; What’s New in the “New Guide”; Post-Approval Monitoring; Lab Animal Behavior; and ACLAM Board Prep Tips. June 6 includes: Histopathology for Lab Animal Vets and Laws and Regulations. June 7 has Phenotyping 101 plus rodent material. June 8 (POLA starts) is all rodents until the evening Breakout Session: Phenotyping Slide Seminar. June 9 includes two sessions on Pathology of Non-Human Primates plus a Breakout Session: TBA. June 10 has two sessions on Clinical Pathology of Lab Animals plus a Breakout Session: Mock Lab Animal Gross Exam. June 11 has no primate material.
Rooms have been reserved at a reduced rate until May 1. Reservations received after this date will be filled on a space available basis. When contacting the hotel to make your reservation, please inform them that you are attending the C. L. Davis / Armed Forces Institute of Pathology’s Current Laboratory Animal Science Seminar and/or the Pathology of Laboratory Animals courses. Hotel phone numbers are 800-325-3535 and 757-425-9000.
More information and registration materials are available at: <www.cldavis.org/upcoming.html>.
Medical and Experimental Mammalian Genetics
The 52nd Annual Short Course on Medical and Experimental Mammalian Genetics will be held July 17–29, 2011, at the Jackson Laboratory, Bar Harbor, Maine. This two-week course includes daily lectures in the mornings and evenings, supplemented by afternoon mini-symposia, workshops, tutorials, a poster session, and demonstrations. There is special emphasis on student-faculty interaction. The faculty is diverse in terms of disciplines and the students in terms of stage of career and fields of concentration. This diversity makes for an exciting experience for all. Although the course schedule is quite full, there is time set aside to enjoy the natural beauty of Mount Desert Island.
The content of the Annual Short Course focuses on (1) an up-to-date presentation of genetics in experimental animals and humans, (2) the relationship of heredity to disease in experimental animals and humans, and (3) the importance of molecular genetics in the diagnosis and treatment of inherited disorders. The development and uses of modern techniques in bioinformatics, mathematical genetics, genome manipulation, mutagenesis, and phenotyping are taught in lectures and in afternoon workshop sessions. The Short Course includes an implicit focus on translational biology and clinical applications.
There will be a poster session at which presentations by attendees are strongly encouraged. If you are interested in presenting a poster, please prepare a one-page abstract to be compiled with others in booklet form for the course. Abstracts must fit on one side of an 8 1/2 x 11” page with 1” margins. It should have a title and author line. The attending author’s name must be underlined. Abstracts of your poster presentation should be submitted in a PDF file or Word document as an e-mail attachment. The abstract submission deadline is June 18, 2011.
See <courses.jax.org/event-listings.html> or contact Laura Ingerson [207-288-6419].
Approaches to Complex Disorders
A course on “Genomic and Proteomic Approaches to Complex Heart, Lung, Blood, and Sleep Disorders” will be held Sept 8–16, 2011, at the Jackson Laboratory, Bar Harbor, Maine. This course covers the application of statistics, molecular biology, and genetics to the analysis of complex diseases such as asthma, hypertension, and coronary heart disease. Both human and animal model data are presented and considered in depth.
Students will learn to detect the genetic basis of disease, perform linkage analysis, conduct genome scans, analyze quantitative trait loci, detect gene interactions, map complex disease genes, and clone genes. Emphasis is also placed on how to characterize gene function using the latest in biochemical and molecular genetic techniques. Contemporary approaches to gene discovery such as mutagenesis, microarray analysis, comparative genomics, and single nucleotide polymorphism discovery and analysis are also covered. Social and ethical issues surrounding the generation and use of genetic information are discussed during the course.
Intensive hands-on workshops further expose students to bioinformatics tools such as: Omim, MGD, Unigene, GeneBank, Blast, Gene-scan, SwissProt and Entrez. The formal lectures, discussion groups, demonstrations and tutorials, study sessions, recreational activities, and informal residential interaction provide a unique and varied learning opportunity.
This intensive residential course is designed for 30–40 participants who hold a doctoral degree or equivalent. The course will be held at the Jackson Laboratory’s Highseas Conference Center.
See <courses.jax.org/event-listings.html> or contact Laura Ingerson [207-288-6419].
* * *
Research and Conservation Opportunities: Brazilian Atlantic Forest
Originally, six species of primates were present at Serra Bonita:
When land was first purchased to create the reserve in 1998, only the marmosets were present in substantial numbers. Groups of capuchins, seldom seen, were formed by 4 or 5 individuals at most, and seemed very scared. Since the reserve was established, in 2001, their numbers have increased significantly. Groups numbering 16–18 individuals are frequently seen around the lodge. The guards have seen groups with up to 25 individuals at the southern end of the reserve, presumably a different group from the ones found around the lodge. The titi monkeys were never seen before; now they are heard calling frequently even around the research center and the lodge.
Some of the marmosets have become very tame. One group of marmosets is attracted by the fruits on the bird feeders. Three individuals came to the feeders for the first time about two years ago. They now number twelve individuals – they are feeding on a bunch of bananas in front of the office window – one adult is carrying a pair of tiny babies on its back.
We need studies on the population dynamics and feeding habits of these existing primate species to inform the reserve’s management plan. Of course, other aspects of the animals’ natural history can be studied as well.
Conservation Opportunities
Two of the original primate species are extinct in the reserve and could be reintroduced: the northern muriqui and the brown howler monkey. The golden-headed lion tamarin is a lowland species, seen once at the lower edges of the reserve, but never at the higher elevations. Given the topography of the reserve, it is probably not a good candidate for reintroduction, at least at this early stage.
Since 2005 three rangers have been employed and hunting has been reduced almost to nothing, especially at the higher elevations around the research center and the lodge. Considering this, the size of the area, and characteristics of the vegetation (more than 50% mature forest and the rest old growth secondary forest), the reserve provides an appropriate opportunity to reintroduce the extinct species.
Expressions of Interest
Individuals or research teams who would be interested in pursuing research and conservation activities with primates at Serra Bonita are encouraged to submit by e-mail a letter and any supporting materials describing their interests, experience, and qualifications to the address below. We encourage potential participants to visit the reserve as part of their deliberations.
The management of the reserve will facilitate studies to the extent that its resources allow, but we will expect our research partners to take a lead role in securing the necessary permits and in obtaining additional funding. We especially encourage Brazilian scientists as individuals or as participants in international teams to submit inquiries. Contact Vitor O. Becker, Scientific Director, Instituto Uiraçu, Serra Bonita Reserve, P.O. Box 01, 45880-000 Camacan, Bahia, Brazil [e-mail: [email protected]]. – posted by Max Snodderly to the Primate-Science mailing list, January 10
* * *
She would, of course, acknowledge the photographer and/or supplier of the photo, and the owner of the image would maintain copyright. Contact Clara at Community Conservation, Inc., 50542 One Quiet Lane, Gays Mills, WI 54631 [608-735-4717; Fax: 512-519-8494; e-mail: [email protected]]. See also <www.communityconservation.org/index.htm>.
* * *
“For over 15 years, Neotropical Primates has been printed and mailed free to subscribers. Now, in order to save paper and conserve forests, and to reduce the considerable costs of distribution and printing, we are asking our readers to let us know if they wish to continue to receive hard copies. You are all welcome to read and/or print copies from <www.primate-sg.org/np.htm>.”
Journal of Evolutionary Biology Research
The Journal of Evolutionary Biology Research (JEBR) is a multidisciplinary, peer-reviewed, open-access journal, published monthly by Academic Journals, <www.academicjournals.org/JEBR>. JEBR is dedicated to increasing the depth of evolutionary biology research across disciplines, with the ultimate aim of improving evolutionary research.
JEBR welcomes the submission of manuscripts that meet the general criteria of significance and scientific excellence in this subject area, and will publish:
JEBR is fully committed to the Open Access Initiative and provides free access to all articles as soon as they are published.
Bioinformatics and Sequence Analysis
Professor A. Badr, Editor of the Journal of Bioinformatics and Sequence Analysis (JBSA) writes:
“The Journal of Bioinformatics and Sequence Analysis is a multidisciplinary peer-reviewed journal, published monthly by Academic Journals <www.academicjournals.org/JBSA>. JBSA is dedicated to increasing the depth of the subject across disciplines with the ultimate aim of expanding knowledge of the subject.
“Editors and reviewers: JBSA is seeking qualified researchers to join its editorial team as editors, subeditors, or reviewers. Please send your resume to <[email protected]>.
“Call for Papers: JBSA will cover all areas of the subject. The journal welcomes the submission of manuscripts that meet the general criteria of significance and scientific excellence, and will publish original articles in basic and applied research, as well as case studies, critical reviews, surveys, opinions, commentaries, and essays. We invite you to submit your manuscript(s) to <[email protected]> for publication. Instructions for authors and other details are available at <www.academicjournals.org/JBSA/Instruction.htm>.
“JBSA is an Open Access Journal: One key request of researchers across the world is unrestricted access to research publications. Open access gives a worldwide audience larger than that of any subscription-based journal and thus increases the visibility and impact of published works. It also enhances indexing and retrieval power and eliminates the need for permissions to reproduce and distribute content. JBSA is fully committed to the Open Access Initiative and will provide free access to all articles as soon as they are published.”
* * *
Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
Books
* Human Evolutionary Biology. M. P. Muehlenbein (Ed.). West Nyack, NY: Cambridge University Press, 2010. 624 pp. [Price: paperback: $70.00; hardback: $130.00]
. . . Sections are Part I: Theory and Methods; Part II: Phenotypic and Genotypic Variation; Part III: Reproduction; Part IV: Growth and Development; and Part V: Health and Disease.
* Manipulative Monkeys: The Capuchins of Lomas Barbudal (2nd Printing). S. Perry with J. H. Manson. Cambridge, MA: Harvard University Press, 2011. [Price: paperback, $22.95; hardcover, $49]
. . . Report on the lives of four generations of capuchins.
* Small Animal Bandaging, Casting, and Splinting Techniques. S. F. Swaim, W. C. Renberg, & K. M. Shike. New York: Wiley-Blackwell, 2011. 144 pp. [Price: $49.99].
* American Journal of Primatology, 2011, 73[2], <onlinelibrary.wiley.com/doi/10.1002/ajp.v73.2/issuetoc>.
. . . Contents: Measuring infant attachment security in rhesus macaques (Macaca mulatta): Adaptation of the attachment Q-set, by J. J. Warfield, K. Kondo-Ikemura, & E. Waters; Differences between the normal vaginal bacterial community of baboons and that of humans, by A. J. Rivera, J. A. Frank, R. Stumpf, A. A. Salyers, B. A. Wilson, G. J. Olsen, & S. Leigh; Associations between the drill (Mandrillus leucophaeus) and sympatric monkeys in Korup National Park, Cameroon, by C. Astaras, S. Krause, L. Mattner, C. Rehse, & M. Waltert; A comparative study of crested gibbons (Nomascus), by A. R. Mootnick & P.-F. Fan; Morphometrics and pattern of growth in wild sifakas (Propithecus edwardsi) at Ranomafana National Park, Madagascar, by S. J. King, T. L. Morelli, S. Arrigo-Nelson, F. J. Ratelolahy, L. R. Godfrey, J. Wyatt, S. Tecot, J. Jernvall, & P. C. Wright; Patterns of gastro-intestinal parasites and commensals as an index of population and ecosystem health: The case of sympatric western chimpanzees (Pan troglodytes verus) and guinea baboons (Papio hamadryas papio) at Fongoli, Senegal, by M. E. Howells, J. Pruetz, & T. R. Gillespie; Estrous asynchrony causes low birth rates in wild female chimpanzees, by A. Matsumoto-Oda & Y. Ihara; Characterization of opsin gene alleles affecting color vision in a wild population of titi monkeys (Callicebus brunneus), by J. A. Bunce, L. A. Isbell, M. Neitz, D. Bonci, A. K. Surridge, G. H. Jacobs, & D. G. Smith; and Space use selectivity by chimpanzees and gorillas in an indoor–outdoor enclosure, by S. R. Ross, S. Calcutt, S. J. Schapiro, & J. Hau.
* American Journal of Primatology, 2011, 73 [4], <onlinelibrary.wiley.com/doi/10.1002/ajp.v73.4/issuetoc>.
. . . Contents: How feeding competition determines female chimpanzee gregariousness and ranging in the Taď National Park, Côte d’Ivoire, by J. Riedel, M. Franz, & C. Boesch; Coevolutionary relationship between striatum size and social play in nonhuman primates, by K. L. Graham; Great apes use weight as a cue to find hidden food, by C. Schrauf & J. Call; Resource partitioning in sympatric langurs and macaques in tropical rainforests of the central Western Ghats, south India, by M. Singh, K. Roy, & M. Singh; Habitat selection of black-and-white snub-nosed monkeys (Rhinopithecus bieti) in Tibet: Implications for species conservation, by Z.-F. Xiang, S. Huo, & W. Xiao; Male–immature relationships in multi-male groups of mountain gorillas (Gorilla beringei beringei), by S. Rosenbaum, J. B. Silk, & T. S. Stoinski; Sexual interference in the golden snub-nosed monkey (Rhinopithecus roxellana): A test of the sexual competition hypothesis in a polygynous species, by X.-G. Qi, B. Yang, P. A. Garber, W. Ji, K. Watanabe, & B.-G. Li; Social and developmental influences on urinary androgen levels in young male white-faced marmosets (Callithrix geoffroyi), by A. K. Birnie, A. S. Smith, C. Nali, & J. A. French; Patterns of mineral lick visitation by spider monkeys and howler monkeys in Amazonia: Are licks perceived as risky areas? by A. Link, N. Galvis, E. Fleming, & A. Di Fiore; and Third-party postconflict affiliation of aggressors in chimpanzees, by T. Romero & F. B. M. de Waal.
* Animal Behaviour, 2010, 81[1], <www.sciencedirect.com/science/journal/00033472>.
. . . Contents include: Using motivation to feed as a way to assess the importance of space for broiler chickens, by S. Buijs, L. J. Keeling, & F. A. M. Tuyttens; Infant access and handling in sooty mangabeys and vervet monkeys, by C. Fruteau, E. van de Waal, E. van Damme, & R. Noë; Environmental enrichment induces optimistic cognitive bias in rats, by N. M. Brydges, M. Leach, K. Nicol, R. Wright, & M. Bateson; and Wild bearded capuchin monkeys (Cebus libidinosus) place nuts in anvils selectively, by Q. Liu, D. Fragaszy, B. Wright, K. Wright, P. Izar, & E. Visalberghi.
* Animal Behaviour, 2010, 81[2], <www.sciencedirect.com/science/journal/00033472>.
. . . Contents include: Social-learning abilities of wild vervet monkeys in a two-step task artificial fruit experiment, by E. van de Waal & R. Bshary; Grooming reciprocity in wild male chimpanzees, by N. E. Newton-Fisher & P. C. Lee; Identifying Sykes’ monkeys’, Cercopithecus albogularis erythrarchus, axes of fear through patch use, by S. E. Emerson, J. S. Brown, & J. D. Linden; and Autonomic reactions indicating positive affect during acoustic reward learning in domestic pigs, by M. Zebunke, J. Langbein, G. Manteuffel, & B. Puppe.
* Comparative Medicine, 2010, 60[6], <tinyurl.com/2b4r3mm>.
. . . Contents include: Refinement of vascular access port placement in nonhuman primates: Complication rates and outcomes, by M. L. Graham, L. A. Mutch, E. F. Rieke, M. Dunning, E. K. Zolondek, A. W. Faig, B. J. Hering, & H.-J. Schuurman; and Blood D-(?)-3-Hydroxybutyrate concentrations after oral administration of trioctanoin, trinonanoin, or tridecanoin to newborn rhesus monkeys (Macaca mulatta), by M. A. Tetrick, F. R. Greer, & N. J. Benevenga.
* Folia Primatologica, 2010, 81[4], <www.karger.com/fpr>.
. . . Allometry and evolution in the galago skull, by M. J. Ravosa, A. N. Daniel, & D. B. Costley; Can overmarking be considered as a means of chemical mate guarding in a wild callitrichid? by Y. Lledo-Ferrer, F. Peláez, & E. W. Heymann; The relationship of accentuated lines in enamel to weaning stress in juvenile baboons (Papio hamadryas anubis), by W. Dirks, L. T. Humphrey, M. C. Dean, & T. E. Jeffries; Preliminary assessment of methods used to demonstrate nut-cracking behavior to five captive chimpanzees (Pan troglodytes), by S. R. Ross, M. S. Milstein, S. E. Calcutt, & E. V. Lonsdorf; and Altitudinal movements of Guizhou snub-nosed monkeys (Rhinopithecus brelichi) in Fanjingshan National Nature Reserve, China: Implications for conservation management of a flagship species, by K. Niu, C. L. Tan, & Y. Yang.
* Folia Primatologica, 2010, 81[5], <www.karger.com/fpr>.
. . . Social structure and group dynamics of the Cao Vit gibbon (Nomascus nasutus) in Bangliang, Jingxi, China, by P. Fan, H. Fei, Z. Xiang, W. Zhang, C. Ma, & T. Huang; A direct comparison of scan and focal sampling methods for measuring wild chimpanzee feeding behaviour, by I. C. Gilby, A. A. Pokempner, & R. W. Wrangham; A preliminary study on the function of food begging in Sichuan snub-nosed monkeys (Rhinopithecus roxellana): Challenge to begging for nutritional gain, by Z. Zhang, Y. Su, & R. C. K. Chan; Influence of the task on hand preference: Individual differences among gorillas (Gorilla gorilla gorilla), by E. Pouydebat, E. Reghem, P. Gorce, & V. Bels; Contact with human facilities appears to enhance technical skills in wild vervet monkeys (Chlorocebus aethiops), by E. van de Waal & R. Bshary; An exploratory study on the combined effects of external and internal morphology on load dissipation in primate capitates: Its potential for an understanding of the positional and locomotor repertoire of early hominins, by G. A. Macho, I. R. Spears, M. G. Leakey, D. J. McColl, Y. Jiang, R. Abel, M. Nakatsukasa, & Y. Kunimatsu; and A population estimate of blue-eyed black lemurs in Ankarafa Forest, Sahamalaza-Iles Radama National Park, Madagascar, by M. S. N. Volampeno, J. C. Masters, & C. T. Downs.
* IPPL News, December, 2010, 37[3], <www.ippl.org/news.php>. (Intl Primate Protection League, P.O. Box 766, Summerville, SC 29484 [e-mail: [email protected]]).
. . . Contents include: The doucs of Vietnam; Hoolock gibbons in peril; Help stop China’s abuse of Guinea’s chimpanzees; announcements; and requests for help.
* Journal of Evolutionary Biology, 2010, 23[12], <onlinelibrary.wiley.com/doi/10.1111/jeb. 2010.23.issue-12/issuetoc>.
. . . Contents include: Parasite-mediated evolution of the functional part of the MHC in primates, by L. Z. Garamszegi & C. L. Nunn; and Night and day: The comparative study of strepsirrhine primates reveals socioecological and phylogenetic patterns in olfactory signals, by J. Delbarco-Trillo, B. A. Burkert, T. E. Goodwin, & C. M. Drea.
* Journal of Medical Primatology, 2011, 40[1], <onlinelibrary.wiley.com/doi/10.1111/jmp. 2010.40.issue-1/issuetoc>.
. . . Contents: A possible case of hantavirus infection in a Borneo orangutan and its conservation implication, by C.-C. Chen, K. J.-C. Pei, C.-M. Yang, M.-D. Kuo, S.-T. Wong, S.-C. Kuo, & F.-G. Lin; Primary pneumonic plague in the African Green monkey as a model for treatment efficacy evaluation, by R. C. Layton, T. Brasel, A. Gigliotti, E. Barr, S. Storch, L. Myers, C. Hobbs, & F. Koster; Cynomolgus monkeys (Macaca fascicularis) may not become infected with equine herpesvirus 9, by A. Kodama, T. Yanai, M. Kubo, N. El-Habashi, S. Kasem, H. Sakai, T. Masegi, H. Fukushi, T. Kuraishi, M. Yoneda, S. Hattori, & C. Kai; Effects of fasting on hematologic and clinical chemical values in cynomolgus monkeys (Macaca fascicularis), by X.-C. Zeng, C.-M. Yang, X.-Y. Pan, Y.-S. Yao, W. Pan, C. Zhou, Z.-R. Jiang, Y. Chang, & J. Ma; Fetal and maternal factors associated with infant mortality in vervet monkeys, by K. Kavanagh, B. L. Dozier, T. J. Chavanne, L. A. Fairbanks, M. J. Jorgensen, & J. R. Kaplan; Unilateral ovarian absence in two capuchin monkeys, by S. F. S. Domingues, D. V. C. Almeida, S. R. R. A. Sealercio, J. P. Kastelic, & R. R. Santos; Flow cytometric characterization of the lymphocyte composition in a variety of mucosal tissues in healthy rhesus macaques, by T. Schultheiss, N. Stolte-Leeb, S. Sopper, & C. Stahl-Hennig; Species diversity and relative abundance of lactic acid bacteria in the milk of rhesus monkeys (Macaca mulatta), by L. Jin, K. Hinde, & L. Tao; and AIDS animal model comes of age, by T. M. Folks.
* Journal of the American Association for Laboratory Animal Science, 2011, 50[1], <tinyurl.com/4shyred>.
. . . Contents include: Harmonic analysis of noninvasively recorded arterial pressure waveforms in healthy bonnet macaques (Macaca radiata), by O. Olafiranye, L. Salciccioli, H. Kamran, M. Stewart, J. Carter, & J. M. Lazar; Peripheral ossifying fibroma and juxtacortical chondrosarcoma in cynomolgus monkeys (Macaca fascicularis), by B. Schmelting, M. Zöller, & J. Kaspareit; Mycobacterium tuberculosis infection in a closed colony of rhesus macaques (Macaca mulatta), by K. S. Payne, J. J. Novak, K. Jongsakul, R. Imerbsin, Y. Apisitsaowapa, J. A. Pavlin, & S. B. Hinds; Severe thrombocytopenia in aged rhesus macaques (Macaca mulatta) infected with simian varicella virus, by L. C. Halliday & J. D. Fortman; and Abstracts of Scientific Papers from the 2010 Association of Primate Veterinarians Workshop.
* Neotropical Primates. A Journal and Newsletter of the IUCN/SSC Primate Specialist Group, December, 2009, 16[2], <www.primate-sg.org/PDF/NP16.2.pdf>.
. . . Contents: Sympatric Alouatta seniculus and Cebus capucinus in an Andean forest fragment in Colombia: A survey of population density, by N. R. Duque & C. Gómez-Posada; Relaçőes sociais, emigraçăo e fissăo de um grupo de Alouatta guariba clamitans em um remanescente de Floresta Ombrófila Mista, by R. F. Moro-Rios, J. M. Deliberador Miranda, & F. de Camargo Passos; Nectar feeding on an exotic tree (Grevillea robusta) by Alouatta caraya and its possible role in flower pollination, by R. Pavé, S. M. Peker, M. Rańo, C. Ramírez Orjuela, G. E. Zunino, & M. M. Kowalewski; Yeasts isolated from Alouatta palliata, Ateles geoffroyi, Cebus capucinus and Saimiri oerstedii (Primates: Cebidae), by N. Gross, J. Diego Castro, O. Guerrero, M. Chinchilla, R. Sánchez and G. Gutiérrez-Espeleta; New sightings of northern muriqui (Brachyteles hypoxanthus) females in forest fragments surrounding the Estaçăo Biológica de Caratinga-RPPN Feliciano Miguel Abdala, Minas Gerais, Brasil, by F. P. Tabacow, C. B. Possamai, F. R. Melo, S. L. Mendes, & K. B. Strier; On the identification of Callicebus cupreus and Callicebus brunneus, by J. Vermeer; Confirmation of Callicebus dubius (Pitheciidae) distribution and evidence of invasion into the geographic range of Callicebus stephennashi, by F. Röhe & J. S. e Silva-Jr.; Sleep tree use by white-faced capuchins (Cebus capucinus): Implications for differences in seedling composition, by K. Valenta, J. A. Klemens, & L. M. Fedigan; Possible evidence of male dispersal in common woolly monkeys (Lagothrix lagotricha), by A. Maldonado & S. Botero; Predation on small mammals by capuchin monkeys, Cebus cay, by M. Zattoni Milano & E. L. Araújo Monteiro-Filho; Object manipulation in a captive group of capuchin monkeys (Cebus nigritus), by D. Santos de Freitas & J. C. Bicca-Marques; and Field observation of predation of a slate-colored hawk, Leucopternis schistacea, on a juvenile saddle-back tamarin, Saguinus fuscicollis, by Y. Lledo-Ferrer, A. Hidalgo, E. W. Heymann, & F. Peláez.
* Tropical Medicine & International Health, 2011, 16[3], <onlinelibrary.wiley.com/doi/10.1111/tmi. 2011.16.issue-3/issuetoc>.
. . . Contents include: Artesunate/mefloquine paediatric formulation vs. artemether/lumefantrine for the treatment of uncomplicated Plasmodium falciparum in Anonkoua kouté, Côte d’Ivoire, by O. A. Toure, M. G. Kouame, Y. J. Didier, A. A. A. Berenger, K. Djerea, G. O. Genevieve, & L. K. Penali; and Success in controlling a major outbreak of malaria because of Plasmodium falciparum in Jamaica, by K. Webster-Kerr, J. P. Figueroa, P. L. Weir, K. Lewis-Bell, E. Baker, J. Horner-Bryce, E. Lewis-Fuller, M. Bullock DuCasse, K. H. Carter, & S. Campbell-Forrester.
* Veterinary Anaesthesia and Analgesia, 2011, 38[1], <onlinelibrary.wiley.com/doi/10.1111/vaa. 2011.38.issue-1/issuetoc>.
. . . Contents include: an Editorial: Animal welfare in biomedical publishing, by R. E. Clutton, K. W. Clarke, & P. J. Pascoe; and Abstracts presented at the American College of Veterinary Anesthesiologists 35th Annual Meeting, San Antonio, Texas, 11–15 September, 2010.
* Primates in 21st Century Ecosystems: Does Primate Conservation Promote Ecosystem Conservation? American Journal of Primatology, 2011, 73[1], <onlinelibrary.wiley.com/doi/10.1002/ajp.v73.1/issuetoc>.
. . . Contents: Editorial: 30th Anniversary of the American Journal of Primatology, by P. A. Garber; Primates in 21st century ecosystems: Does primate conservation promote ecosystem conservation? by M. A. Norconk, S. Boinski, & P.-M. Forget; Primate seed dispersers as umbrella species: A case study from Kibale National Park, Uganda, with implications for Afrotropical forest conservation, by J. E. Lambert; A glance to the past: Subfossils, stable isotopes, seed dispersal, and lemur species loss in Southern Madagascar, by B. E. Crowley, L. R. Godfrey, & M. T. Irwin; Lemurs in a complex landscape: Mapping species density in subtropical dry forests of southwestern Madagascar using data at multiple levels, by A. C. Axel & B. A. Maurer; Usefulness of species range polygons for predicting local primate occurrences in southeastern Peru, by S. Palminteri, G. Powell, W. Endo, C. Kirkby, D. Yu, & C. A. Peres; Conserving social–ecological systems in Indonesia: Human–nonhuman primate interconnections in Bali and Sulawesi, by E. P. Riley & A. Fuentes; Black and gold howler monkeys (Alouatta caraya) as sentinels of ecosystem health: Patterns of zoonotic protozoa infection relative to degree of human–primate contact, by M. M. Kowalewski, J. S. Salzer, J. C. Deutsch, M. Rańo, M. S. Kuhlenschmidt, & T. R. Gillespie; Feeding competition, cooperation, and the causes of primate sociality: A commentary on Sussman et al., by R. R. Lawler; Reply to Lawler: Feeding competition, cooperation, and the causes of primate sociality, by R. W. Sussman, P. A. Garber, & J. M. Cheverud; and A new species of snub-nosed monkey, genus Rhinopithecus Milne-Edwards, 1872 (Primates, Colobinae), from northern Kachin state, northeastern Myanmar, by T. Geissmann, N. Lwin, S. S. Aung, T. N. Aung, Z. M. Aung, T. H. Hla, M. Grindley, & F. Momberg.
* The Effects of Bonds Between Human and Non-human Primates on Primatological Research and Practice. American Journal of Primatology, 2011, 73[3], <onlinelibrary.wiley.com/doi/10.1002/ajp.v73.3/issuetoc>.
. . . Contents: Editorial: Tribute to Ymke Warren; Introduction to the special section: “The effects of bonds between human and nonhuman primates on primatological research and practice”, by A. Vitale & S. Pollo; Primatology between feelings and science: A personal experience perspective, by A. Vitale; Caring for nonhuman primates in biomedical research facilities: Scientific, moral and emotional considerations, by K. Coleman; The first chimpanzee sanctuary in Japan: An attempt to care for the “surplus” of biomedical research, by N. Morimura, G. Idani, & T. Matsuzawa; Being human and doing primatology: National, socioeconomic, and ethnic influences on primatological practice, by A. Fuentes; Of bonds and boundaries: What is the modern role of anthropomorphism in primatological studies? By P. J. Asquith; Bonding, biophilia, biosynergy, and the future of primates in the wild, by A. L. Rose; Nest site ecology of the Cross River gorilla at the Kagwene Gorilla Sanctuary, Cameroon, with special reference to anthropogenic influence, by R. A. De Vere, Y. Warren, A. Nicholas, M. E. Mackenzie, & J. P. Higham; Inter-unit contests within a provisioned troop of Sichuan snub-nosed monkeys (Rhinopithecus roxellana) in the Qinling Mountains, China, by Q. Zhao & C. L. Tan; Diet and ranging behavior of the endangered Javan gibbon (Hylobates moloch) in a submontane tropical rainforest, by S. Kim, S. Lappan, & J. C. Choe; Population-level right-handedness for a coordinated bimanual task in naturalistic housed chimpanzees: Replication and extension in 114 animals from Zambia and Spain, by M. Llorente, D. Riba, L. Palou, L. Carrasco, M. Mosquera, M. Colell, & O. Feliu; and Kinda baboons (Papio kindae) and grayfoot chacma baboons (P. ursinus griseipes hybridize in the Kafue river valley, Zambia, by C. J. Jolly, A. S. Burrell, J. E. Phillips-Conroy, C. Bergey, & J. Rogers.
* Using induced pluripotent stem cells to investigate cardiac phenotypes in Timothy syndrome. Yazawa, M., Hsueh, B., Jia, X., Pasca, A. M., Bernstein, J. A., Hallmayer, J., & Dolmetsch, R. E. (R. E. D., Dept of Neurobio., Stanford Univ. Sch. of Med., Stanford, CA 94305 [e-mail: [email protected]]. Nature, 2011, 471, 230-234, <www.nature.com/nature/journal/v471/n7337/full/nature09855.html>.
. . . “Individuals with congenital or acquired prolongation of the QT interval, or long QT syndrome (LQTS), are at risk of life-threatening ventricular arrhythmia. LQTS is commonly genetic in origin but can also be caused or exacerbated by environmental factors. A missense mutation in the L-type calcium channel CaV1.2 leads to LQTS in patients with Timothy syndrome. To explore the effect of the Timothy syndrome mutation on the electrical activity and contraction of human cardiomyocytes, we reprogrammed human skin cells from Timothy syndrome patients to generate induced pluripotent stem cells, and differentiated these cells into cardiomyocytes. Electrophysiological recording and calcium (Ca2+) imaging studies of these cells revealed irregular contraction, excess Ca2+ influx, prolonged action potentials, irregular electrical activity and abnormal calcium transients in ventricular-like cells. We found that roscovitine, a compound that increases the voltage-dependent inactivation of CaV1.2, restored the electrical and Ca2+ signalling properties of cardiomyocytes from Timothy syndrome patients. This study provides new opportunities for studying the molecular and cellular mechanisms of cardiac arrhythmias in humans, and provides a robust assay for developing new drugs to treat these diseases.”
* Vocalization induced cFos expression in marmoset cortex. Miller, C., Dimauro, A., Pistorio, A., Hendry, S., & Wang, X. (Cortical Systems & Behavior Lab., Dept of Psychology, Univ. of California, San Diego, 9500 Gilmann Dr. #109, La Jolla, CA 92093 [e-mail: corymiller @ucsd.edu]). Frontiers in Integrative Neuroscience, 2011, 4[128], doi: 10.3389/fnint.2010.00128,<www.frontiersin.org/integrative_neuroscience/10.3389/fnint.2010.00128/full>.
. . . “All non-human primates communicate with conspecifics using vocalizations, a system involving both the production and perception of species-specific vocal signals. Much of the work on the neural basis of primate vocal communication in cortex has focused on the sensory processing of vocalizations, while relatively little data are available for vocal production. Earlier physiological studies in squirrel monkeys had shed doubts on the involvement of primate cortex in vocal behaviors. The aim of the present study was to identify areas of common marmoset (Callithrix jacchus) cortex that are potentially involved in vocal communication. In this study, we quantified cFos expression in three areas of marmoset cortex – frontal, temporal (auditory), and medial temporal – under various vocal conditions. Specifically, we examined cFos expression in these cortical areas during the sensory, motor (vocal production), and sensory–motor components of vocal communication. Our results showed an increase in cFos expression in ventrolateral prefrontal cortex as well as the medial and lateral belt areas of auditory cortex in the vocal perception condition. In contrast, subjects in the vocal production condition resulted in increased cFos expression only in dorsal premotor cortex. During the sensory–motor condition (antiphonal calling), subjects exhibited cFos expression in each of the above areas, as well as increased expression in perirhinal cortex. Overall, these results suggest that various cortical areas outside primary auditory cortex are involved in primate vocal communication. These findings pave the way for further physiological studies of the neural basis of primate vocal communication.”
* Morphology of brachial plexus and axillary artery in bonobo (Pan paniscus). Kikuchi, Y., Oishi, M., & Shimizu, D. (Dept of Anat. & Physiol., Fac. of Med., Saga Univ., Saga 849-8501, Japan [e-mail: [email protected]]). Anatomia, Histologia, Embryologia, 2011, 40, 68-72, <onlinelibrary.wiley.com/doi/10.1111/ j.1439-0264.2010.01040.x/full>.
. . . A left brachial plexus and axillary artery of bonobo were examined, and the interrelation between the brachial plexus and the axillary artery was discussed. This is the first report of the brachial plexus and the axillary artery of bonobo. The bonobo brachial plexus formed a very similar pattern to that of other ape species and human. On the other hand, the branches of the bonobo axillary artery had uncommon architecture in comparison with human case. The axillary artery did not penetrate the brachial plexus and passes through all the way along anterior to the brachial plexus. Only 4.9% of human forelimbs have this pattern. Moreover, the brachial artery runs through superficially anterior to branches of the brachial plexus.
* Long-lasting reduction in hippocampal neurogenesis by alcohol consumption in adolescent nonhuman primates. Taffe, M. A., Kotzebue, R. W., Crean, R. D., Crawford, E. F., Edwards, S., & Mandyam, C. D. (C. D. M., Committee on the Neurobiology of Addictive Disorders, The Scripps Research Inst., La Jolla, CA 92037 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2010, 107, 11104–11109, <www.pnas.org/content/107/24/11104.full>.
. . . Binge alcohol consumption in adolescents is increasing, and studies in animal models show that adolescence is a period of high vulnerability to brain insults. The purpose of the present study was to determine the deleterious effects of binge alcohol on hippocampal neurogenesis in adolescent nonhuman primates. Heavy binge alcohol consumption over 11 months dramatically and persistently decreased hippocampal proliferation and neurogenesis. Combinatorial analysis revealed distinct, actively dividing hippocampal neural progenitor cell types in the subgranular zone of the dentate gyrus that were in transition from stem-like radial glia-like cells (type 1) to immature transiently amplifying neuroblasts (type 2a, type 2b, and type 3), suggesting the evolutionary conservation of milestones of neuronal development in macaque monkeys. Alcohol significantly decreased the number of actively dividing type 1, 2a, and 2b cell types without significantly altering the early neuronal type 3 cells, suggesting that alcohol interferes with the division and migration of hippocampal preneuronal progenitors. Furthermore, the lasting alcohol-induced reduction in hippocampal neurogenesis paralleled an increase in neural degeneration mediated by nonapoptotic pathways. Altogether, these results demonstrate that the hippocampal neurogenic niche during adolescence is highly vulnerable to alcohol and that alcohol decreases neuronal turnover in adolescent nonhuman primate hippocampus by altering the ongoing process of neuronal development. This lasting effect, observed two months after alcohol discontinuation, may underlie the deficits in hippocampus-associated cognitive tasks that are observed in alcoholics.
* Spontaneous voice–face identity matching by rhesus monkeys for familiar conspecifics and humans. Sliwa, J., Duhamel, J.-R., Pascalis, O., & Wirth, S. (S. W., Centre de Neuroscience Cognitive, Centre National de la Recherche Scientifique, Univ. Lyon I, 69675 Bron, France [e-mail: [email protected])]. Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 1735-1740, <www.pnas.org/content/108/4/1735.full>.
. . . “Recognition of a particular individual occurs when we reactivate links between current perceptual inputs and the previously formed representation of that person. This recognition can be achieved by identifying, separately or simultaneously, distinct elements such as the face, silhouette, or voice as belonging to one individual. In humans, those different cues are linked into one complex conceptual representation of individual identity. Here we tested whether rhesus macaques (Macaca mulatta) also have a cognitive representation of identity by evaluating whether they exhibit cross-modal individual recognition. Further, we assessed individual recognition of familiar conspecifics and familiar humans. In a free preferential looking time paradigm, we found that, for both species, monkeys spontaneously matched the faces of known individuals to their voices. This finding demonstrates that rhesus macaques possess a cross-modal cognitive representation of individuals that extends from conspecifics to humans, revealing the adaptive potential of identity recognition for individuals of socioecological relevance.”
* Readily available tissue-engineered vascular grafts. Dahl, S. L. M., Kypson, A. P., Lawson, J. H., Blum, J. L., Strader, J. T., Li, Y., Manson, R. J., Tente, W. E., DiBernardo, L., Hensley, M. T., Carter, R., Williams, T. P., Prichard, H. L., Dey, M. S., Begelman, K. G., & Niklason, L. E. (Humacyte Inc., Durham, NC 27709 [e-mail: [email protected]]). Science Translational Medicine, 2011, 3, 68ra9, <stm.sciencemag.org/content/3/68/68ra9.full>.
. . . “Autologous or synthetic vascular grafts are used routinely for providing access in hemodialysis or for arterial bypass in patients with cardiovascular disease. However, some patients either lack suitable autologous tissue or cannot receive synthetic grafts. Such patients could benefit from a vascular graft produced by tissue engineering. Here, we engineer vascular grafts using human allogeneic or canine smooth muscle cells grown on a tubular polyglycolic acid scaffold. Cellular material was removed with detergents to render the grafts nonimmunogenic. Mechanical properties of the human vascular grafts were similar to native human blood vessels, and the grafts could withstand long-term storage at 4°C. Human engineered grafts were tested in a baboon model of arteriovenous access for hemodialysis. Canine grafts were tested in a dog model of peripheral and coronary artery bypass. Grafts demonstrated excellent patency and resisted dilatation, calcification, and intimal hyperplasia. Such tissue-engineered vascular grafts may provide a readily available option for patients without suitable autologous tissue or for those who are not candidates for synthetic grafts.”
* Successful respiratory immunization with dry powder live-attenuated measles virus vaccine in rhesus macaques. Lin, W.-H., Griffin, D. E., Rota, P. A., Papania, M., Cape, S. P., Bennett, D., Quinn, B., Sievers, R. E., Shermer, C., Powell, K., Adams, R. J., Godin, S., & Winston, S. (D. E. G., Dept of Molecular Microbio. & Immunol., Johns Hopkins Bloomberg Sch. of Public Health, Baltimore, MD 21205 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 180, 2987-2992, <www.pnas.org/content/108/7/2987.full>.
. . . “Measles remains an important cause of childhood mortality worldwide. Sustained high vaccination coverage is the key to preventing measles deaths. Because measles vaccine is delivered by injection, hurdles to high coverage include the need for trained medical personnel and a temperature-controlled supply chain, waste of vaccine in multidose vials, and risks associated with needle use and disposal. Respiratory vaccine delivery could lower these barriers and facilitate sustained high coverage. We developed a novel single unit dose, dry powder live-attenuated measles vaccine (MVDP) for respiratory delivery without reconstitution. We tested the immunogenicity and protective efficacy in rhesus macaques of one dose of MVDP delivered either with a mask or directly intranasal with two dry powder inhalers, PuffHaler and BD Solovent. MVDP induced robust measles virus (MeV)-specific humoral and T-cell responses, without adverse effects, which completely protected the macaques from infection with wild-type MeV more than one year later. Respiratory delivery of MVDP was safe and effective and could aid in measles control.”
* Vulnerability of the fetal primate brain to moderate reduction in maternal global nutrient availability. Antonow-Schlorke, I., Schwab, M., Cox, L. A., Li, C., Stuchlik, K., Witte, O. W., Nathanielsz, P. W., & McDonald, T. J. (T. J. M., Univ. of Texas Health Science Center, San Antonio, TX 78227-5301 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 3011-3016, <www.pnas.org/content/108/7/3011>.
. . . “Moderate maternal nutrient restriction during pregnancy occurs in both developing and developed countries. In addition to poverty, maternal dieting, teenage pregnancy, and uterine vascular problems in older mothers are causes of decreased fetal nutrition. We evaluated the impact of global 30% maternal nutrient reduction (MNR) on early fetal baboon brain maturation. MNR induced major cerebral developmental disturbances without fetal growth restriction or marked maternal weight reduction. Mechanisms evaluated included neurotrophic factor suppression, cell proliferation and cell death imbalance, impaired glial maturation and neuronal process formation, down-regulation of gene ontological pathways and related gene products, and up-regulated transcription of cerebral catabolism. Contrary to the known benefits from this degree of dietary reduction on life span, MNR in pregnancy compromises structural fetal cerebral development, potentially having an impact on brain function throughout life.”
* Euthanasia: In defense of a good, ancient word. Pavlovic, D., Spassov, A., & Lehmann, C. (Ernst-Moritz-Arndt Univ., Friedrich-Loeffler-Str. 23b, 17487 Greifswald, Germany [e-mail: [email protected]]). Clinical Research & Bioethics, 2011, 2, 105, <www.omicsonline.org/2155-9627/2155-9627-2-105.digital/2155-9627-2-105.html>.
. . . “It has been suggested that the word euthanasia should be abandoned because widespread imprecise use of the word has deprived it of its precise meaning. On the basis of the primary necessary condition for euthanasia – that it be in the interests of the one who will die – we examine the use and meaning of the expression. We demonstrate that the above suggestions rely upon flagrant misuse or inappropriate use of the word. The attention of the reader is drawn to morally problematic consequences should the expression ‘euthanasia’ be abandoned and the argument that abandoning the term would be justified is rejected. Instead of abandoning the word ‘euthanasia,’ care should be taken to use the term correctly in the future. The original meaning of the word would, thereby, be preserved and we could only benefit from the rich tradition that impregnates that ancient word.”
* Chimpanzees know that others make inferences. Schmelz, M., Call, J., & Tomasello, M. (Dept of Development & Comp. Psych., Max Planck Inst. for Evolutionary Anthropology, D-04103 Leipzig, Germany [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 180, 3077-3079, <www.pnas.org/content/108/7/3077.full>.
. . . “If chimpanzees are faced with two opaque boards on a table, in the context of searching for a single piece of food, they do not choose the board lying flat (because if food were under there it would not be lying flat) but, rather, they choose the slanted one — presumably inferring that some unperceived food underneath is causing the slant. Here we demonstrate that chimpanzees know that other chimpanzees in the same situation will make a similar inference. In a back-and-forth foraging game, when their competitor had chosen before them, chimpanzees tended to avoid the slanted board on the assumption that the competitor had already chosen it. Chimpanzees can determine the inferences that a conspecific is likely to make and then adjust their competitive strategies accordingly.”
* Responses to the Assurance game in monkeys, apes, and humans using equivalent procedures. Brosnan, S. F., Parrish, A., Beran, M. J., Flemming, T., Heimbauer, L., Talbot, C. F., Lambeth, S. P., Schapiro, S. J., & Wilson, B. J. (Neurosci. Inst., Georgia State Univ., Atlanta, GA 30302 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 3442-3447, <www.pnas.org/content/108/8/3442.full>.
. . . “There is great interest in the evolution of economic behavior. In typical studies, species are asked to play one of a series of economic games, derived from game theory, and their responses are compared. The advantage of this approach is the relative level of consistency and control that emerges from the games themselves; however, in the typical experiment, procedures and conditions differ widely, particularly between humans and other species. Thus, in the current study, we investigated how three primate species, capuchin monkeys, chimpanzees, and humans, played the Assurance (or Stag Hunt) game using procedures that were, to the best of our ability, the same across species, particularly with respect to training and pretesting. Our goal was to determine what, if any, differences existed in the ways in which these species made decisions in this game. We hypothesized differences along phylogenetic lines, which we found. However, the species were more similar than might be expected. In particular, humans who played using “nonhuman primate-friendly” rules did not behave as is typical. Thus, we find evidence for similarity in decision-making processes across the order Primates. These results indicate that such comparative studies are possible and, moreover, that in any comparison rating species’ relative abilities, extreme care must be taken in ensuring that one species does not have an advantage over the others due to methodological procedures.”
* An assessment of gum-based environmental enrichment for captive gummivorous primates. Huber, H. F., & Lewis, K. P. (Dept of Anthropology, Southern Illinois Univ., Carbondale, IL 62903 [e-mail: [email protected]]). Zoo Biology, 2011, 30, 71-78, <onlinelibrary.wiley.com/doi/10.1002/zoo.20321/full>.
. . . “In the wild, many primates consume gums exuded from trees, and many species are gum specialists. In spite of this, few data exist concerning gum feeding in captivity. Using a Web-based survey of 46 zoos in 12 countries, we evaluated the extent to which zoos feed gum to primates. We found that although callitrichids and galagos receive gum-based enrichment, cercopithecines generally do not. Environmental enrichment is important for stimulating naturalistic behavior to promote the psychological wellbeing of animals. Thus, gum-based enrichment is important for captive gummivores. Our study highlights the need to improve environmental enrichment for captive gummivores, in particular that of cercopithecines. This is most striking for the patas monkey (Erythrocebus patas), an obligate gummivore. The exchange of ecological data between field research and captive settings is crucial, and is just one way primate caretakers can contribute to the conservation and welfare of some of our closest living relatives.”
* Longitudinal patterns of viremia and oral shedding of rhesus rhadinovirus and retroperitoneal fibromatosis herpesviruses in age-structured captive breeding populations of rhesus macaques (Macaca mulatta). White, J., Yang, X., Todd, P., & Lerche, N. W. (Koelle Lab, Univ. of Washington, Seattle, WA 98102 [e-mail: [email protected]]). Comparative Medicine, 2011, 61, 60-70.
. . . “Rhesus rhadinovirus (RRV) and retroperitoneal fibromatosis herpesvirus (RFHV), 2 closely related γ2 herpesviruses, are endemic in breeding populations of rhesus macaques at our institution. We previously reported significantly different prevalence levels, suggesting the transmission dynamics of RRV and RFHV differ with regard to viral shedding and infectivity. We designed a longitudinal study to further examine the previously observed differences between RRV and RFHV prevalence and the potential influence of age, season, and housing location on the same 90 rhesus macaques previously studied. Virus- and host-genome-specific real-time PCR assays were used to determine viral loads for both RRV and RFHV in blood and saliva samples collected at 6 time points over an 18-mo period. Proportions of positive animals and viral load in blood and saliva were compared between and within viruses by age group, location, and season by using 2-part longitudinal modeling with Bayesian inferences. Our results demonstrate that age and season are significant determinants, with age as the most significant factor analyzed, of viremia and oral shedding for both RRV and RFHV, and these pathogens exhibit distinctly different patterns of viremia and oral shedding over time within a single population.”
* Functional comparison of innate immune signaling pathways in primates. Barreiro, L. B., Marioni, J. C., Blekhman, R., Stephens, M., & Gilad, Y. (Dept of Human Genetics, Univ. of Chicago, Chicago, IL 60637 [e-mail: [email protected]]). PLOS Genetics, 2010, <10.1371/journal.pgen.1001249>.
. . . “Humans respond differently than other primates to a large number of infections. Differences in susceptibility to infectious agents between humans and other primates are probably due to inter-species differences in immune response to infection. Consistent with that notion, genes involved in immunity-related processes are strongly enriched among recent targets of positive selection in primates, suggesting that immune responses evolve rapidly, yet providing only indirect evidence for possible inter-species functional differences. To directly compare immune responses among primates, we stimulated primary monocytes from humans, chimpanzees, and rhesus macaques with lipopolysaccharide (LPS) and studied the ensuing time-course regulatory responses. We find that, while the universal Toll-like receptor response is mostly conserved across primates, the regulatory response associated with viral infections is often lineage-specific, probably reflecting rapid host–virus mutual adaptation cycles. Additionally, human-specific immune responses are enriched for genes involved in apoptosis, as well as for genes associated with cancer and with susceptibility to infectious diseases or immune-related disorders. Finally, we find that chimpanzee-specific immune signaling pathways are enriched for HIV-interacting genes. Put together, our observations lend strong support to the notion that lineage-specific immune responses may help explain known inter-species differences in susceptibility to infectious diseases.”
* Microfossils in calculus demonstrate consumption of plants and cooked foods in Neanderthal diets (Shanidar III, Iraq; Spy I and II, Belgium). Henry, A. G., Brooks, A. S., & Piperno, D. R. (Dept of Anthropology, Ctr for Advanced Study of Hominid Paleobiology, Washington, DC 20052 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 486-491, <www.pnas.org/content/108/2/486.full>.
. . . “The nature and causes of the disappearance of Neanderthals and their apparent replacement by modern humans are subjects of considerable debate. Many researchers have proposed biologically or technologically mediated dietary differences between the two groups as one of the fundamental causes of Neanderthal disappearance. Some scenarios have focused on the apparent lack of plant foods in Neanderthal diets. Here we report direct evidence for Neanderthal consumption of a variety of plant foods, in the form of phytoliths and starch grains recovered from dental calculus of Neanderthal skeletons from Shanidar Cave, Iraq, and Spy Cave, Belgium. Some of the plants are typical of recent modern human diets, including date palms (Phoenix spp.), legumes, and grass seeds (Triticeae), whereas others are known to be edible but are not heavily used today. Many of the grass seed starches showed damage that is a distinctive marker of cooking. Our results indicate that in both warm eastern Mediterranean and cold northwestern European climates, and across their latitudinal range, Neanderthals made use of the diverse plant foods available in their local environment and transformed them into more easily digestible foodstuffs in part through cooking them, suggesting an overall sophistication in Neanderthal dietary regimes.”
* A shift toward birthing relatively large infants early in human evolution. DeSilva, J. M. (Dept of Anthropology, Boston Univ., Boston, MA 02215 [e-mail: jdesilva@ bu.edu]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 1022-1027, <www.pnas.org/content/108/3/1022.abstract?etoc>.
. . . It has long been argued that modern human mothers give birth to proportionately larger babies than apes do. Data presented here from human and chimpanzee infant:mother dyads confirm this assertion: humans give birth to infants approximately 6% of their body mass, compared with approximately 3% for chimpanzees, even though the female body weights of the two species are moderately convergent. Carrying a relatively large infant both pre- and postnatally has important ramifications for birthing strategies, social systems, energetics, and locomotion. However, it is not clear when the shift to birthing large infants occurred over the course of human evolution. Here, known and often conserved relationships between adult brain mass, neonatal brain mass, and neonatal body mass in anthropoids are used to estimate birthweights of extinct hominid taxa. These estimates are resampled with direct measurements of fossil postcrania from female hominids, and also compared with estimates of female body mass to assess when human-like infant:mother mass ratios (IMMRs) evolved. The results of this study suggest that 4.4-Myr-old Ardipithecus possessed IMMRs similar to those found in African apes, indicating that a low IMMR is the primitive condition in hominids. Australopithecus females, in contrast, had significantly heavier infants compared with dimensions of the femoral head (n = 7) and ankle (n = 7) than what is found in chimpanzees, and are estimated to have birthed neonates more than 5% of their body mass. Carrying such proportionately large infants may have limited arboreality in Australopithecus females and may have selected for alloparenting behavior earlier in human evolution than previously thought.
* Late Pleistocene adult mortality patterns and modern human establishment. Trinkaus, E. (Dept of Anthropology, Washington Univ, St. Louis, MO 63130 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 108, 1267-1271, <www.pnas.org/content/108/4/1267.full>.
. . . The establishment of modern humans in the Late Pleistocene, subsequent to their emergence in eastern Africa, is likely to have involved substantial population increases, during their initial dispersal across southern Asia and their subsequent expansions throughout Africa and into more northern Eurasia. An assessment of younger (20–40 y) versus older (>40 y) adult mortality distributions for late archaic humans (principally Neandertals) and two samples of early modern humans (Middle Paleolithic and earlier Upper Paleolithic) provides little difference across the samples. All three Late Pleistocene samples have a dearth of older individuals compared with Holocene ethnographic/historical samples. They also lack older adults compared with Holocene paleodemographic profiles that have been critiqued for having too few older individuals for subsistence, social, and demographic viability. Although biased, probably through a combination of preservation, age assessment, and especially Pleistocene mobility requirements, these adult mortality distributions suggest low life expectancy and demographic instability across these Late Pleistocene human groups. They indicate only subtle and paleontologically invisible changes in human paleodemographics with the establishment of modern humans; they provide no support for a life history advantage among early modern humans.
* High-temperature environments of human evolution in East Africa based on bond ordering in paleosol carbonates. Passey, B. H., Levin, N. E., Cerling, T. E., Brown, F. H., & Eiler, J. M. (Dept of Earth & Planetary Sci., Johns Hopkins Univ., 301 Olin Hall, 3400 N. Charles St, Baltimore, MD 21218 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2011, 107, 11245-11249, <www.pnas.org/content/107/25/11245.full>.
. . . “Many important hominid-bearing fossil localities in East Africa are in regions that are extremely hot and dry. Although humans are well adapted to such conditions, it has been inferred that East African environments were cooler or more wooded during the Pliocene and Pleistocene when this region was a central stage of human evolution. Here we show that the Turkana Basin, Kenya—today one of the hottest places on Earth—has been continually hot during the past 4 million years. The distribution of 13C-18O bonds in paleosol carbonates indicates that soil temperatures during periods of carbonate formation were typically above 30°C and often in excess of 35°C. Similar soil temperatures are observed today in the Turkana Basin and reflect high air temperatures combined with solar heating of the soil surface. These results are specific to periods of soil carbonate formation, and we suggest that such periods composed a large fraction of integrated time in the Turkana Basin. If correct, this interpretation has implications for human thermophysiology and implies a long-standing human association with marginal environments.”
* An extinct monkey from Haiti and the origins of the Greater Antillean primates. Cooke, S. B., Rosenberger, A. L., & Turvey, S. (Dept of Evol. Anthro., Duke Univ., Durham, NC 27708 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A, 2011, 180, 2699-2704, <www.pnas.org/content/108/7/2699. full>.
. . . A new extinct Late Quaternary platyrrhine from Haiti, Insulacebus toussaintiana, is described here from the most complete Caribbean subfossil primate dentition yet recorded, demonstrating the likely coexistence of two primate species on Hispaniola. Like other Caribbean platyrrhines, I. toussaintiana exhibits primitive features resembling early Middle Miocene Patagonian fossils, reflecting an early derivation before the Amazonian community of modern New World anthropoids was configured. This, in combination with the young age of the fossils, provides a unique opportunity to examine a different parallel radiation of platyrrhines that survived into modern times, but is only distantly related to extant mainland forms. Their ecological novelty is indicated by their unique dental proportions, and by their relatively large estimated body weights, possibly an island effect, which places the group in a size class not exploited by mainland South American monkeys. Several features tie the new species to the extinct Jamaican monkey Xenothrix mcgregori, perhaps providing additional evidence for an inter-Antillean clade.
* Gorilla and orangutan brains conform to the primate cellular scaling rules: Implications for human evolution. Herculano-Houzel, S., & Kaas, J. H. (Inst. de Cięncias Biomédicas, UFRJ, Rua Carlos Chagas Filho 337, Ilha do Fundăo, Rio de Janeiro, RJ 21941-920 Brazil [e-mail [email protected]]). Brain, Behavior and Evolution, 2011, 77, 33-44, <www.ncbi.nlm.nih.gov/pubmed/21228547>.
. . . “Gorillas and orangutans are primates at least as large as humans, but their brains amount to about one third of the size of the human brain. This discrepancy has been used as evidence that the human brain is about 3 times larger than it should be for a primate species of its body size. In contrast to the view that the human brain is special in its size, we have suggested that it is the great apes that might have evolved bodies that are unusually large, on the basis of our recent finding that the cellular composition of the human brain matches that expected for a primate brain of its size, making the human brain a linearly scaled-up primate brain in its number of cells. To investigate whether the brain of great apes also conforms to the primate cellular scaling rules identified previously, we determine the numbers of neuronal and other cells that compose the orangutan and gorilla cerebella, use these numbers to calculate the size of the brain and of the cerebral cortex expected for these species, and show that these match the sizes described in the literature. Our results suggest that the brains of great apes also scale linearly in their numbers of neurons like other primate brains, including humans. The conformity of great apes and humans to the linear cellular scaling rules that apply to other primates that diverged earlier in primate evolution indicates that prehistoric Homo species as well as other hominins must have had brains that conformed to the same scaling rules, irrespective of their body size. We then used those scaling rules and published estimated brain volumes for various hominin species to predict the numbers of neurons that composed their brains. We predict that Homo heidelbergensis and Homo neanderthalensis had brains with approximately 80 billion neurons, within the range of variation found in modern Homo sapiens. We propose that while the cellular scaling rules that apply to the primate brain have remained stable in hominin evolution (since they apply to simians, great apes and modern humans alike), the Colobinae and Pongidae lineages favored marked increases in body size rather than brain size from the common ancestor with the Homo lineage, while the Homo lineage seems to have favored a large brain instead of a large body, possibly due to the metabolic limitations to having both.”
* Genetic history of an archaic hominin group from Denisova Cave in Siberia. Reich, D., Green, R. E., Kircher, M., Krause, J., Patterson, N., Durand, E. Y., Viola, B., et al. (Dept of Genetics, Harvard Med. Sch., Boston, MA 02115 [e-mail: [email protected]]). Nature, 2010, 468, 1053-1060, <www.nature.com/nature/journal/v468/n7327/full/nature09710.html>.
. . . “Using DNA extracted from a finger bone found in Denisova Cave in southern Siberia, we have sequenced the genome of an archaic hominin to about 1.9-fold coverage. This individual is from a group that shares a common origin with Neanderthals. This population was not involved in the putative gene flow from Neanderthals into Eurasians; however, the data suggest that it contributed 4–6% of its genetic material to the genomes of present-day Melanesians. We designate this hominin population ‘Denisovans’ and suggest that it may have been widespread in Asia during the Late Pleistocene epoch. A tooth found in Denisova Cave carries a mitochondrial genome highly similar to that of the finger bone. This tooth shares no derived morphological features with Neanderthals or modern humans, further indicating that Denisovans have an evolutionary history distinct from Neanderthals and modern humans.”
* Evolution of neuronal and endothelial transcriptomes in primates. Giger, T., Khaitovich, P., Somel, M., Lorenc, A., Lizano, E., Harris, L. W., Ryan, M. M., Lan, M., Wayland, M. T., Bahn, S., & Pääbo, S. (Max-Planck-Inst. for Evol. Anthropology, Deutscher Platz, D-04103 Leipzig, Germany [e-mail: [email protected]]). Genome Biology and Evolution, 2010, 2, 284-292, <gbe.oxfordjournals.org/content/2/284.full>.
. . . “The study of gene expression evolution in vertebrates has hitherto focused on the analysis of transcriptomes in tissues of different species. However, since a tissue is made up of different cell types, and cell types differ with respect to their transcriptomes, the analysis of tissues offers a composite picture of transcriptome evolution. The isolation of individual cells from tissue sections opens up the opportunity to study gene expression evolution at the cell type level. We have stained neurons and endothelial cells in human brains by antibodies against cell type-specific marker proteins, isolated the cells using laser-capture-microdissection, and identified genes preferentially expressed in the two cell types. We analyze these two classes of genes with respect to their expression in 62 different human tissues, with respect to their expression in 44 human post mortem brains from different developmental stages, and with respect to between-species brain expression differences. We find that genes preferentially expressed in neurons differ less across tissues and developmental stages than genes preferentially expressed in endothelial cells. We also observe less expression differences within primate species for neuronal transcriptomes. In stark contrast, we see more gene expression differences between humans, chimpanzees, and rhesus macaques relative to within species differences in genes expressed preferentially in neurons than in genes expressed in endothelial cells. This suggests that neuronal and endothelial transcriptomes evolve at different rates within brain tissue.”
* Comparative and demographic analysis of orang-utan genomes. Locke, D. P., Hillier, L. W., Warren, W. C., Worley, K. C., et al. (Washington Univ. Sch. of Med., 4444 Forest Park Ave, Saint Louis, MO, 63108). Nature, 2011, 469, 529-533, <www.nature.com/nature/journal/v469/n7331/full/nature09687.html>.
. . . “‘Orang-utan’ is derived from a Malay term meaning ‘man of the forest’ and aptly describes the southeast Asian great apes native to Sumatra and Borneo. The orang-utan species, Pongo abelii (Sumatran) and Pongo pygmaeus (Bornean), are the most phylogenetically distant great apes from humans, thereby providing an informative perspective on hominid evolution. Here we present a Sumatran orang-utan draft genome assembly and short read sequence data from five Sumatran and five Bornean orang-utan genomes. Our analyses reveal that, compared to other primates, the orang-utan genome has many unique features. Structural evolution of the orang-utan genome has proceeded much more slowly than other great apes, evidenced by fewer rearrangements, less segmental duplication, a lower rate of gene family turnover and surprisingly quiescent Alu repeats, which have played a major role in restructuring other primate genomes. We also describe a primate polymorphic neocentromere, found in both Pongo species, emphasizing the gradual evolution of orang-utan genome structure. Orang-utans have extremely low energy usage for a eutherian mammal, far lower than their hominid relatives. Adding their genome to the repertoire of sequenced primates illuminates new signals of positive selection in several pathways including glycolipid metabolism. From the population perspective, both Pongo species are deeply diverse; however, Sumatran individuals possess greater diversity than their Bornean counterparts, and more species-specific variation. Our estimate of Bornean/Sumatran speciation time, 400,000 years ago, is more recent than most previous studies and underscores the complexity of the orang-utan speciation process. Despite a smaller modern census population size, the Sumatran effective population size (Ne) expanded exponentially relative to the ancestral Ne after the split, while Bornean Ne declined over the same period. Overall, the resources and analyses presented here offer new opportunities in evolutionary genomics, insights into hominid biology, and an extensive database of variation for conservation efforts.”
* Use of an implantable loop recorder in the investigation of arrhythmias in adult captive chimpanzees (Pan troglodytes). Lammey, M. L., Jackson, R., Ely, J. J., Lee, D. R., & Sleeper, M. M. (Alamogordo Primate Facility, Holloman Air Base, Alamogordo, NM, 88330 [e-mail: [email protected]]). Comparative Medicine, 2011, 61, 71-75.
. . . “Cardiovascular disease in general, and cardiac arrhythmias specifically, are common in great apes. However, the clinical significance of arrhythmias detected on short-duration electrocardiograms is often unclear. Here we describe the use of an implantable loop recorder to evaluate cardiac rhythms in 4 unanesthetized adult chimpanzees, 1 with a history of possible syncope and 3 with the diagnosis of multiform ventricular ectopy (ventricular premature complexes) and cardiomyopathy. The clinical significance of ventricular ectopy was defined further by using the implantable loop recorder. Arrhythmia was ruled out as a cause of collapse in the chimpanzee that presented with possible syncope because the implantable loop recorder demonstrated normal sinus rhythm during a so-called syncopal event. This description is the first report of the use of an implantable loop recorder to diagnose cardiac arrhythmias in an unanesthetized great ape species.”
-------------------------------------------
We would like to acknowledge Primate-Science as a source for information about new books.
-------------------------------------------
* * *
The Newsletter appeared quarterly and was intended primarily for persons doing research with nonhuman primates. We hope that it has served its purpose well. Fifty years is a long time in the life of a woman who came to this job with no background in psychology, biology, or publication.
Volume 50 will be the last volume; there will be two more issues after this one: July and October. We hope that we will still receive manuscripts (until August 1) and announcements (until September 1).
All back issues may still be purchased for $10.00 each. Obviously, we will not accept future subscriptions, since this is the last volume. Current and back issues of the Newsletter are available on the World Wide Web at <http://brown.edu/primate>. We hope to spend the last months of 2011 putting PDF copies of all the earlier back issues onto the Website.
We would also like to thank all of the people who have helped in this long project. We cannot begin to name all of you, but we will certainly mention the Assistant Editor, without whom the last 23 volumes could not have been printed; all of the Veterinarians here at Brown (and one now retired), who have read and advised; our statistician Consulting Editor; and the people of the Psychology Department here at Brown. Look at the Contents for Previous Years file, to see the names that appear again and again!
And we must never forget Allan Schrier, who had the idea, and the strength to make it a reality. Thank you, Allan!
* * *
Contents of Volumes 46-48 (2007-2009)
Contents of Volume 49-50 (2010-2011)
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, CLPS Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; fax: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
Current and back issues of the Newsletter are available on the World Wide Web at
http://www.brown.edu/primate
The Newsletter is supported by Brown University
Cover photograph of a bonobo (Pan paniscus), three year old Vic, visiting with his friend, a three year old boy in red. Taken at the Cincinnati Zoo and Botanical Gardens by Marian Brickner, June 2, 2006.
Copyright (c) 2011 by Brown University
Last updated: March 18, 2011