 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 29 NUMBER 4 OCTOBER 1990
Articles and Notes
Behavioral Responses of Female Long-Tailed Macaques (Macaca fascicularis) to Pair Formation, by S. W. Line, K. N. Morgan, H. Markowitz, J. A. Roberts, & M. Riddell ...... 1
Training Stumptailed Monkeys (Macaca arctoides) to Cooperate during In-Homecage Treatment, by V. Reinhardt & D. Cowley ...... 9
Conservation Costs, by W. R. Kingston ...... 13
News, Information, and Announcements
Editors' Notes ...... 5
. . LPN Goes Electronic, Help for Overseas Subscribers, Ketamine Shortage
Filovirus Updates ...... 7
. . From CDC, Philippine Department of Health
New APHIS Regulations ...... 8
Revised Mefloquine Dosage ...... 8
Grants Available ...... 10
. . Alan T. Waterman Award, Small Grants Program, Aging Research,
Research Training Groups, Institutional Dental Grants, Diabetes
Research
Course Announcement: Ethical Issues ...... 11
Meeting Report: ASP in Davis, CA ...... 12
Research Opportunity: Specimens for Study ...... 12
News Briefs...... 16
. . Silver Spring Monkeys Euthanized, Australian Animal Laws,
IPS Newsletter, Four Monkeys Proposed as Endangered,
Florida Key Monkeys Upset the Neighbors, Chimpanzee Classification
Information Requested and Available...... 17
. . IAMAT, WRPRC Publication, Primate Exporter, CDC Hotline
Meeting Announcements...... 18
. . Humane Care and Use of Laboratory Animals, Primates in Evolution,
XXIV World Veterinary Congress, Effect of Regulations on Animal
Welfare
Cartoon, by R. Connor ...... 6
Departments
Address Changes...... 14
Recent Books and Articles...... 19
* * *
Scott W. Line, Kathleen N. Morgan, Hal Markowitz, Jeffrey A. Roberts, and Mike Riddell
California Regional Primate Research Center and San Francisco State University
While most primate species are social in nature, it has been standard practice in many laboratories to house adult primates in individual cages. Single housing has several advantages from a husbandry standpoint, including reduced frequency of trauma and disease transmission, improved access to individual animals for experimental procedures, and increased safety for animal handlers. Considering the animals' perspective, however, individual housing eliminates the opportunity for many normal behavior patterns.
The 1985 amendment to the Animal Welfare Act has led to renewed interest in group-housing as a means of maintaining primates in research facilities. While it is not known what form the final regulations that will be used to enforce the law will take, it is likely that they will emphasize increased use of social housing. The last public proposal from the Department of Agriculture recommended social housing whenever possible (USDA, 1989).
Pair-housing is one form of social housing that may be practical for replacing individual housing of primates in many situations. Monkeys in pairs can perform a large number of social behaviors not available to those that are singly caged, and expanding the behavioral repetoire is one way of improving primate welfare (Line, 1987; Novak & Suomi, 1988). Pair-housing may also be accomplished relatively economically by modifying existing single cages. We recently added side openings to several stainless steel squeeze-back cages at a cost of approximately $150/cage. The openings are 50 cm wide by 23 cm high, approximately 20% of the side wall. When the cages are placed next to each other, two monkeys housed in them can move freely back and forth. If necessary, a solid partition can be locked in place to separate the monkeys.
Despite the conventional wisdom that unfamiliar adult macaques are more likely to fight than to coexist peacefully, Reinhardt and co-workers have shown that pair-housing can be successful for adult rhesus monkeys (Macaca mulatta) that are unfamiliar with each other and have lived in single cages for prolonged periods. They have paired juvenile rhesus with adults of both sexes (Reinhardt et al., 1987), adult females (Reinhardt et al., 1988), and adult males (Reinhardt, 1989). Reinhardt (1990b) also reported that unrelated stump-tailed macaques (M. arctoides) can be successfully pair-housed.
In this experiment we attempted to replicate these findings in another species of primate commonly used in laboratory research, the long-tailed macaque (M. fascicularis). Beyond determining whether or not compatible pairs can be formed with unfamiliar individuals, we also wanted to document general changes in behavior associated with the move from single to pair-housing. Providing monkeys with the opportunity for social interaction is only one step toward improving their welfare. We also wanted to obtain quantitative data on whether the monkeys expressed more social behaviors.
Unlike Reinhardt (1990a), we feel that monitoring the occurrence of abnormal behavior is useful in assessing primate welfare. While some abnormal behaviors have been observed among free-ranging and group-housed primates, they are much less frequent among socially housed primates than among those that are singly-housed. Still, there is no general consensus on the meaning and importance of abnormal behavior patterns. Abnormal behaviors could be viewed as positive coping responses that enable an animal to adapt to an unnatural environment. On the other hand, Dantzer (1986) argued that environments that require such coping responses are inadequate. It is our view that reductions in the frequency or duration of abnormal behaviors are evidence that welfare has improved. Consequently, we also measured abnormal behavior before and after the move to pair-housing.
Methods
The subjects were 12 adult female long-tailed macaques (mean weight = 3.4 kg, mean estimated age = 8.8 years). Eleven were wild-caught as adults and had lived in single indoor cages in the colony for a mean of 3.5 years (range = 2.0 to 7.0 years). One was born in the colony in a multi-female, multi-male group, and had been singly housed indoors for 6.4 years.
At the beginning of the experiment all subjects were housed singly in three different colony rooms containing approximately 50 long-tailed macaques. They were kept in stainless steel squeeze-back cages 0.68 m deep, 0.79 m high, and 1.83 m wide (manufactured by Lab Products/Harford, Maywood, NJ). Each cage was divided into three identically-sized sections (0.68 x 0.79 x 0.61 m) by removable stainless steel partitions, and was mounted in a double tier on a wheeled rack. All subjects were housed in upper-row cages, were fed a commercial diet twice daily, and had continuous access to automatic water dispensers. Room lights were controlled automatically on a 12:12 light:dark cycle. All animal care procedures were conducted according to standard laboratory protocols. The Primate Center is AAALAC-accredited.
Frequency and duration of behavior were recorded during 10-minute observation sessions using an automatic wand and bar-code system with a laptop computer. Frequencies and durations of each behavior were summarized individually, then combined to establish a general activity budget with the following behavioral categories: agonistic behavior (hitting, grabbing, biting, presenting, grimacing, threatening, or shaking the cage directed at other monkeys or the observer), cage manipulation (oral or manual exploration of any part of the cage), abnormal postures (self-holding, saluting, and other bizarre postures), stereotypic actions (drinking urine, self-sucking, head tossing, and rocking), stereotypic locomotion (pacing, circling, swinging, somersaulting, and other repetitive actions with no apparent purpose), self-abuse (self-biting, self-hitting, and plucking fur), foraging (holding, biting, sniffing, or manipulating pieces of food, and drinking water), grooming, standing, non-stereotypic locomotion, and resting (passive contact, sleeping, crouching, lying, and sitting while otherwise inactive). Abnormal postures, stereotypic actions, stereotypic locomotion, and self-abuse were added together for a composite abnormal behavior duration. In addition, we recorded the frequency of vocalizations, the time each subject spent in the front half of the cage and, after pairing, the time pair-mates spent in proximity to one another (i.e. within the same half of the cage). We also calculated the duration of observer-directed behavior from the amount of agonistic behavior performed while looking at the observer.
Ten baseline observations were performed on each subject while they were housed singly in their original cages. Ten additional observations were made on each monkey during the first two weeks after they were paired according to the protocols described below. A total of 36 hours of data was collected by seven observers. Inter-rater reliability exceeded 90%. Observations were conducted between 0800 and 1700 hours, primarily between 1000 and 1300. The time of day of observation was matched between the baseline and paired conditions.
The first eight subjects were housed in Room 1, and were assigned to pairs after their baseline data were analyzed. We made the assignments primarily on the rate of aggressive behavior observed during this period. Two monkeys showed much higher rates than any of the others, and these two were assigned as pair-mates to two that showed no aggressive behavior. The other two pairs were created by designating monkeys of similar weight and age as pair-mates.
We then attempted to assess compatibility between these subjects prior to pair formation. We placed the potential pair-mates into open-mesh stainless steel wire transfer boxes measuring 30 cm wide by 36 cm high by 54 cm long. The boxes were then placed next to each other on the floor of the anteroom of Room 1 for 15 minutes while an observer recorded the behavior of each macaque. Each subject was returned to its original cage after the test.
These subjects were subsequently paired by moving each monkey into an adjacent section of a new cage and removing the partition. All eight were moved to a new location in Room 1 in an attempt to reduce aggressive behavior caused by territoriality.
To test whether or not pair formation could be successfully accomplished with a minimum of labor, we did not perform the pair compatibility test for the other four subjects. These monkeys were initially housed in Rooms 2 and 3, and were assigned to pairs randomly. They were subsequently placed in adjacent cage sections in Room 1, and we paired them by simply removing the partition.
Data were analyzed by either a one-tailed or two-tailed paired t-test, depending on whether or not we expected a change in a particular direction for each behavior following pair formation. A p-value less than 0.05 was considered significant.
Results
Pair formation was successful for six of the first eight monkeys. In the unsuccessful case one subject grabbed and bit the other repeatedly during the entire ten minutes they were together. The aggressor was the animal that showed the highest rate of aggressive behavior in the baseline period. Her partner crouched in a corner of the cage, vocalizing and grimacing, but this did not appear to deter the attacks. No serious injuries occurred, but the frequency of attacks did not appear to be declining, and so the subjects were separated. No aggressive behavior was seen during the initial interactions of the other three pairs. Additional checks were made several times a day during the next week to assure continued compatibility.
Over the seven days following pair formation, four of the paired subjects sustained minor wounds, none of which required veterinary treatment. No fighting was directly observed during this time, and no additional wounds were detected after the first week.
The other four monkeys (which did not experience the pair compatibility test and were not moved prior to pair formation) were also successfully paired. No injuries or fighting were noted in any of these subjects during the first two weeks. Seven weeks after pair formation one monkey among these four sustained mild trauma. While no fighting was directly observed, multiple small lacerations were discovered on her head and neck. She was separated from her pair-mate and anesthetized to allow closer inspection and cleaning of the wounds. The two were kept apart overnight, and re-paired the next morning by removing the partition. They immediately moved together and groomed each other; no further trauma has been detected.
The five successful pairs have remained together for five to six months, and are still compatible. All subjects in these pairs were observed to groom each other and show other affiliative behaviors within the first ten minutes after pair formation. In two cases the monkeys moved together and grasped each other in a ventral-ventral embrace immediately after the partition was removed.
Figure 1: Behavioral changes of 10 adult female long-tailed macaques following pair formation. Each bar represents the mean (+SEM) % time engaged in a behavior while singly caged (open bars) and while paired (shaded bars). P-values represent significance levels for paired t-tests.
A summary of the mean activity budget of the ten compatible pair-mates before and after pairing is presented in Figure 1. The total time spent grooming increased significantly after pairing (one-tailed t-test, p = 0.0097). Most of the grooming observed in the pair condition was allogrooming. The time spent autogrooming decreased significantly from 24% to 10% after pairing (one-tailed t-test, p = 0.0053). The amount of time devoted to rest and abnormal behavior also decreased significantly after pairing (one-tailed t-tests, p = 0.0353 and p = 0.0028, respectively). Foraging also decreased significantly after pairing (two-tailed t-test, p = 0.0249). No other activity budget categories changed significantly after pair formation.
Among the other behaviors recorded, time spent in observer-directed activity declined significantly from 2.9% to 0.7% (one-tailed t-test, p = 0.0333), vocalization frequency declined from 45.5/hour to 31.3/hour (two-tailed t-test, p = 0.0241), and time spent in the front half of the cage decreased significantly from 75.5% to 40.5% (two-tailed t-test, p = 0.0216). Pair-mates spent 63.1% of the observation time in proximity to one another.
Behavior Singly caged Pair-housed P-value pace 20,85 (14.4) 9.31 (6.96) 0.2419 abnormal posture 11.65 (8.32) 1.56 (1.01) 0.1281 stereotypical action 39.29 (16.92) 4.0 (2.25) 0.0221 self-abuse 4.48 (4.06) 0.0 (0) 0.1493 total 76.24 (21.71) 14.79 (9.12) 0.0028
Table 1: Influence of housing condition on abnormal behaviors among 10 adult female long-tailed macaques. Each figure is the mean (+SEM) duration in seconds per 10 minute observation period. P-values represent the significance levels for a comparison of the two conditions with a 1-tailed t-test.
The mean duration per 10 minutes of each of the four categories of abnormal behavior in singly-caged and pair-housing conditions appear in Table 1. All four categories decreased in duration after the move to pair-housing.
Discussion
The major findings of this experiment were: 1) pair-housing of unfamiliar adult female long-tailed macaques was successful for five of six pairs, and 2) affiliative behaviors occurred frequently, while abnormal behaviors decreased significantly among pair-housed subjects. Since the pairs have only been housed together for six months or less, and the follow-up observations were performed in the first two weeks after pair formation, these conclusions should be considered preliminary. While there is a continued risk of fighting, we have seen no evidence of serious incompatibility between pair-mates thus far. We are continuing to collect data to see if the behavioral changes observed are maintained beyond the initial period of pair formation.
It is possible that the motivational systems involved in the expression of the different categories of abnormal behavior are varied, and that these behaviors should be considered separately, rather than as a composite response to the change in the social environment. While there was a decrease in the mean duration of all four categories of abnormal behavior, only the decrease in stereotypic action was statistically significant. There were decreases in the duration of each abnormal behavior for the majority of subjects, however. A larger sample size may have resulted in more of the changes reaching statistical significance. For example, self-abusive behaviors were recorded for five of the ten subjects when singly housed, but were completely absent after pair formation.
We found that the minimal-labor protocol, in which monkeys were randomly assigned to one another and paired by simply removing the partition between cage sections, was just as successful as the more detailed procedure used for the first four pair attempts. We have subsequently created two additional compatible pairs using the minimal-labor approach. Reinhardt (1990b) also reported success using a similar pairing technique with stump-tailed macaques.
We did not predict a change in the amount of time foraging after pair formation. Examination of the components of foraging revealed that the decline was entirely due to a decrease in the amount of time spent drinking. All ten subjects showed a decline in drinking. One potential explanation is that the amount of time drinking in the singly caged condition was unusually high. Although none of these subjects showed clinical signs of polydipsia, psychogenic polydipsia has been documented in singly caged rhesus macaques in our colony (Bicknese et al., 1989). It is possible that stress associated with single caging elicited excessive water consumption, and that this pattern was changed by the move to pair-housing.
All but one of the subjects were wild-caught, so we cannot be completely certain whether or not all pairs were unrelated, and unfamiliar with each other. We do know that they were not housed together while in our colony (a period ranging from two to seven years). One pair was comprised of a wild-caught and a colony-born animal. Two other pairs included wild-caught monkeys obtained from different suppliers 29 and 49 months apart. In the final two pairs both individuals were obtained from the same supplier, but at times 8 months and 15 months apart from one another. Given this background, we feel it is reasonable to assume they did not know each other prior to this experiment.
An alternative explanation for the changes in behavior following pair formation is that the subjects became habituated to the presence of observers. This might explain the decreases in abnormal behaviors, resting, observer-directed activity, and frequency of vocalizations. If this were the case, however, we would expect to see a gradual decline in each of these behaviors over time, a pattern that did not occur. There were no decreasing trends in mean level of each behavior over the course of the experiment, and repeated-measures analyses of variance on each behavior within the baseline period were nonsignificant. It is more likely that time interacting with the cage-mate replaced time devoted to these activities.
Pair-housing represents a compromise between keeping primates in large groups and keeping them in individual cages. It increases some risks to the animals and personnel, including both risk of trauma and disease transmission. It may also inconvenience some experimental protocols. Overall, however, it provides substantial benefits to the animals. In a comparison of pair-mates, perches, and pieces of wood, Reinhardt (1990c) concluded that pair-mates provided the most effective long-term stimulation for adult rhesus macaques. While the absolute cost of increased use of pair-housing will not be small, it is likely to be one of the least expensive and most effective alternatives for improving the welfare of nonhuman primates in research facilities.
References
Bicknese, E. J., Eisele, P. H., & George, J. W. (1989). Psychogenic polydipsia in individually housed adult rhesus macaques (Macaca mulatta). Laboratory Animal Science, 39, 476-477.
Dantzer, R. (1986). Behavioral, physiological and functional aspects of stereotyped behavior: A review and a re-interpretation. Journal of Animal Science, 62, 1776-1786.
Line, S. W. (1987). Environmental enrichment for laboratory primates. Journal of the American Veterinary Medical Association, 190, 854-859.
Novak, M. A., & Suomi, S. J. (1988). Psychological well-being of primates in captivity. American Psychologist, 43, 765-773.
Reinhardt, V. (1989). Behavioral responses of unrelated adult male rhesus monkeys familiarized and paired for the purpose of environmental enrichment. American Journal of Primatology, 17, 243-248.
Reinhardt, V. (1990a). Evaluating the effectiveness of environmental enrichment. Laboratory Primate Newsletter, 29 [1], 15.
Reinhardt, V. (1990b). Environmental enrichment program for caged stump-tailed macaques (Macaca arctoides). Laboratory Primate Newsletter, 29 [2], 10-11.
Reinhardt, V. (1990c). Time budget of caged rhesus monkeys exposed to a companion, a PVC perch, and a piece of wood for an extended time. American Journal of Primatology, 20, 51-56.
Reinhardt, V., Houser, W. D., Eisele, S. G., & Champoux, M. (1987). Social enrichment of the environment with infants for singly caged adult rhesus monkeys. Zoo Biology, 6, 365-371.
Reinhardt, V., Houser, W. D., Eisele, S. G., Cowley, D., & Vertein, R. (1988). Behavioral responses of unrelated rhesus monkey females paired for the purpose of environmental enrichment. American Journal of Primatology, 14, 135-140.
U. S. Department of Agriculture (1989). Animal welfare proposed rules. Federal Register, 54 [49], 10822-10954.
-------------------------------------------------------------------
First author's address: California Regional Primate
Research Center, Univ. of California, Davis, CA 95617-8542.
This work was supported by NIH grant RR00169-28 to the CRPRC.
The authors thank Carmel Stanko, Julie Ferris, Astrid Paletzki, and
Kevin Hall for assistance in data collection.
* * *
LPN Goes Electronic
Starting with the January, 1991, issue, the Laboratory Primate Newsletter will be available as an Electronic List, to be called LPN-L. All of the articles, including tables but no graphs or pictures, will be posted immediately after the issue is sent to press, approximately the 15th of the month preceeding the cover month. Each article and department will be posted as a separate file, so that subscribers may save and/or print out as much or as little as seems important.
This List may be accessed from BITNET, UUNET, CompuServe, and other networks. A person may join by sending the following message:
subscribe LPN-L your-own-name
This message should be the first line in an electronic letter sent to the following address:
from BITNETlistserv@brownvm
from other networks
[email protected]
If you do not receive a reply from the brownvm listserver, then ask your system administrator for help.
Subscribers who choose to receive the electronic version will continue to receive the printed issues unless they request to be dropped from the mailing list. We encourage those who do not need the printed version to let us know. Persons and organizations who do not currently receive the LPN may join this Electronic List.
This service should be especially helpful to readers overseas who now have a long wait for sea mail delivery. We realize that many readers will not be able to use this new feature, but believe that the next few years will bring about more nearly universal access to electronic communication, along with improved technology that will enable us to send pictures and graphs.
LPN Goes Electronic, Part 2
In 1990, eight of the articles we published were sent to us by electronic mail, and proofs returned and discussed the same way. We would like to encourage contributors to send their notices and manuscripts by electronic mail to [email protected] (or else to primate@brownvm) if it is possible. This will eliminate typing errors and mailing delays. Figures and tables, along with a single copy of the manuscript, should be sent by mail.
Help for Overseas Subscribers
Since 1974 we have asked our foreign subscribers to pay a small fee to cover the extra cost of overseas mailing (domestic mail is sent fourth class, which is very inexpensive). That fee has increased from the original $2/year to the current $5/year. In 1991 it will cost $6 to mail four issues to a foreign address by sea mail. The airmail rate will be $12. Subscribers who pay for future years before January 1, 1991, may pay at the $5 ($10 by air) rate.
Most foreign subscribers must buy bank checks or money orders for dollars, which increases their expense disproportionately. We have changed our bookkeeping system so that we can send bills for overseas postage to third parties. A few of our American subscribers have taken on the responsibility of paying for the mailing charges of foreign academics and researchers. We would like to encourage more American individuals and corporations to underwrite their colleagues in this way. You may offer to pay for a specific individual, or be assigned one. This will be especially helpful to subscribers in countries where foreign exchange is difficult to obtain.
Ketamine Shortage
All veterinary and other medical supply houses are back ordering ketamine, as it is not currently being made because of a manufacturing problem. New supplies are not expected until late November, 1990, at the earliest. You are advised to use your supplies judiciously.
* * *
From CDC
[This letter, dated August 10, 1990, was sent from William L. Roper, Director of the Centers for Disease Control, to "Interested Parties."]
This is a further update on the current status of issues related to the identification of filovirus in imported nonhuman primates.
Since our last update on July 6, we have granted special permits to import cynomolgus, African green, or rhesus monkeys to four importers. One shipment has arrived in the United States; three more shipments are anticipated during August.
As discussed in the special permit procedures that were published in the Federal Register of April 20, 1990 (55 FR 15210), one broader permit has now been granted for unlimited shipments for a period of 180 days. Additional extended permits will be considered as other importers demonstrate their ability to comply with all of the provisions of their approved import permits.
Facilities of all registered importers have been inspected with the following results: 147 registered importers as of March 1, 1990; 80 withdrawals from list of registered importers; 31 registrations retained after inspection; 36 registrations revoked.
Ten importers whose registrations were revoked have been reinstated, resulting in a total of 41 registered importers as of today.
CDC currently recommends the indirect fluorescent antibody (IFA) method for assaying filovirus antibody in support of nonhuman primate quarantine protocols. We are in the process of reviewing alternative laboratories for their capabilities to provide filovirus testing in support of quarantine release decisions.
Also, CDC recommends that filovirus antibody testing be limited to the simultaneous assay of paired sera obtained at the beginning and end of the quarantine period, as described in the Interim Protocol distributed on May 16, 1990. The purpose of this testing is to determine, by virtue of a four-fold or ,cb greater rise in antibody titer, whether active filovirus infection is present.
This recommended limited use of filovirus serology is reinforced by recent experience in conducting filovirus serosurveys of nonhuman primates in the United States. IFA assay testing of large numbers of nonhuman primate sera over the past several months has yielded many low-titered IFA reactions that are inconsistent with knowledge of documented filovirus infections. Many of these low-titered IFA results have not been consistently reproducible or confirmable by other assays. At present, low-positive IFA titers (16-32) in single, unpaired sera must be considered uninterpretable. Therefore, the testing of single serum specimens is not recommended for determination of current or prior infection with filovirus.
I trust this information is useful to you.
Philippine Department of Health
[Dr. Manuel M. Dayrit, Program Manager, Field Epidemiology Training Program, Department of Health (DOH), Manila, issued the following memo concerning a meeting with exporters of non-human primates on August 15, 1990.]
Instead of paired testing of live monkeys upon start and end of a 30-day quarantine period, DOH recommends that only monkeys which die during a 30-day quarantine period be tested. If one dead monkey is found Ebola antigen positive, it is recommended that (a) the batch within which mortality occurred be sacrificed; (b) restrictions on export from the facility be imposed for a second 30-day quarantine period when testing of dead monkeys will be undertaken; (c) lifting of export restrictions will be made when no more dead monkeys test positive for Ebola antigen.
* * *
Revised APHIS Regulations were published August 15, 1990 (55 Fed. Reg. No 158, pp. 33448-33531), and comments were solicited, with a deadline of October 1, 1990. The new regulations are a vast improvement over earlier proposed regulations issued last year. They are minimally detrimental to the conduct of good biomedical and behavioral research and management.
Some important aspects of the proposed regulations are as follows:
Positive Aspects
Potential Problems
* * *
CDC has revised the dosing recommendations for mefloquine (Lariam&S'R.) use. The new regimen consists of a single dose of mefloquine to be taken weekly, starting 1 week before travel. Prophylaxis should be continued weekly during travel in malarious areas and for 4 weeks after a person leaves such areas.
This notice updates the information in CDC's "Recommendations for the prevention of malaria among travelers" (MMWR, 1990, 39, No. RR-3) and "Health information for international travel" (DHHS publication no. (CDC)90-8280). Detailed recommendations for the prevention of malaria may be obtained 24 hours a day by calling the CDC Malaria Hotline at 404-332-4555. -- From Morbidity and Mortality Weekly Reports, 1990, 39, 630.
* * *
Viktor Reinhardt and Doug Cowley
Wisconsin Regional Primate Research Center
The handling of experimental macaques is often associated with distress for the animals, who are removed from their familiar homecages, and strain for the animal caretakers, who have to lift and carry heavy loads. Transferring a 10-kg animal in a 6-kg transport box from its cage to another room and back may constitute a considerable strain on a person's lower back, particularly when the animal is housed in an upper-row cage.
It has recently been shown that adult rhesus monkeys can easily and quickly be trained to cooperate during in-homecage venipunture, eliminating the need to move them to a restraint apparatus (Vertein & Reinhardt, 1989). The success of the training procedure was so encouraging that it was adapted to another research protocol requiring daily application of various drugs to the foreheads of stumptailed monkeys (Macaca arctoides).
There were 20 adult stumptails -- 17 males and 3 females -- living in 10 compatible pairs (Reinhardt, 1990), 5 pairs in lower-row double cages and 5 pairs in upper-row double cages. All cages were equipped with sturdy, replaceable plastic plates that fit into the cage door openings (Figure 1). Each plate had a face-shaped hole (14 cm wide, 17 cm high) fitting an adult stumptail's head and two smaller circular holes (10 cm diameter) fitting its forearms. The arrangement of the holes was such that an animal could reach out for food offered and eat it while presenting its forehead for treatment (Figure 1) . The three holes enabled an animal to secure and eat the food offered, but not to grab or bite the caretaker.
Several sham treatments were performed before the morning feeding on several days to ensure the animals' eagerness to retrieve the food as planned. Pairs were temporarily separated by means of a cage divider so that one partner could be treated without the other interfering.
Figure 1: Laboratory primates can readily be trained to cooperate during various experimental procedures in their homecages.
Only 1 to 14 training sessions, each lasting for 1-5 minutes, were required to habituate the animals to this new in-homecage treatment procedure. The small time investment paid off quickly, and the ease of the daily treatments not only avoided an unnecessary health risk for the animal caretaker but also saved the time required to get the animals out of their cages and to transfer them back and forth.
Animal care staff should be encouraged to train their charges to cooperate during routine handling procedures in the animals' familiar homecages. This improves experimental design as well as offering environmental enrichment for the experimental animals, whose intelligence is challenged. It also allows animal caretakers to expand their professional skills in creative, productive ways and hence makes their work more satisfying.
References
Reinhardt, V. (1990). Environmental enrichment program for caged stump-tailed macaques (Macaca arctoides). Laboratory Primate Newsletter, 29 [2], 10-11.
Vertein, R. & Reinhardt, V. (1989). Training female rhesus monkeys to cooperate during in-homecage venipuncture. Laboratory Primate Newsletter, 28 [2], 1-3. -------------------------------------------------------------------
Authors' address: Wisconsin Regional Primate Research
Center, 1223 Capitol Court, Madison, WI 53715.
We wish to thank Mr. Dick Pape for preparing the plastic plates and
Mr. John Wolf for editing this manuscript. The training project was
supported by NIH grant RR-00167 to the WRPRC.
* * *
Alan T. Waterman Award
This annual award recognizes an outstanding young scientist in the forefront of science. In addition to a medal and other recognition, the recipient receives grants of up to $500,000 for a period of up to three years for scientific research or advanced study in the biological, medical, or other sciences at the institution of the recipient's choice. Candidates must be U.S. citizens or permanent residents and must be 35 years of age or younger, OR not more than 5 years beyond receipt of the Ph.D. degree by December 31 of the year in which nominated.
For candidates to be considered for the 1991 Award, nominations must be received by the Foundation before December 31, 1990. For more information contact: Mrs. Susan E. Fannoney, Executive Secretary, The Alan T. Waterman Award Committee, NSF, 1800 G Street, NW, Washington, DC 20550 [202-357-7512].
Small Grants Program
The Animal Resources Program of the Division of Research Resources, NIH, continues to offer annually up to 4 one-year, non-renewable awards, to provide support for pilot projects, testing of new techniques, or feasibility studies of innovative research in the area of laboratory animal medicine, which would provide a basis for more extended research. Appropriate areas of study include but are not limited to the etiology, detection, or prevention of laboratory animal disease; methods to decrease pain and distress associated with the use of animals in research; and optimal housing requirements. The award will be for one year and will provide a maximum of $25,000 (direct costs) for technical assistance, supplies, small equipment, and travel, but not for professional salary. The award may not be used to supplement projects currently supported by any other funds, or to provide interim support for projects under review by the Public Health Service.
Any domestic non-profit or for-profit institution may apply. The Principal Investigator (PI) must be actively working in laboratory animal medicine at the time of the award. However, investigators receiving funds from grants for Laboratory Animal Diagnostic and Investigational Laboratories or Regional Primate Research Centers from the Division of Research Resources, NIH, may not act as PI's in this program.
The application receipt date is February 1 of each year. For information, contact the Director, Laboratory Animal Sciences Program, Animal Resources Branch, Division of Research Resources, NIH, 5333 Westbard Ave., Rm. 853, Bethesda, MD 20892 [301-496-5175].
Aging Research
The American Federation for Aging Research, Inc. (AFAR) is accepting grant applications for 1991. As in the past, grants up to $25,000 will be awarded, tenable July 1, 1991, to June 30, 1992, to support one-year clinical or basic research proposals on all biomedical aspects of aging. AFAR is especially interested in supporting investigators who are entering the field of aging. AFAR does not encourage former grantees or established investigators who are already working in the field of geriatrics to apply for support. Applications and further information may be obtained from AFAR, 725 Park Avenue, NYC, NY 10021 [212-570-2090]. Submission deadline is January 15, 1991.
Research Training Groups
The National Science Foundation's Biological and Behavioral Sciences Directorate is inviting applications for grants to encourage multidisciplinary training and to support the acquisition or development of state-of-the-art shared instruments and special purpose materials. U.S. Ph.D.-granting institutions that have basic research and training programs in the biological, behavioral, and social sciences are eligible for awards in basic science areas, including cellular and molecular biology, neurosciences, psychology and anthropology, but not research with disease-related goals, including work on the etiology, diagnosis or treatment of physical or mental disease.
Preproposals are due November 3, 1990, invited formal proposals March 1, 1991, for awards averaging $250,000 for 5 years. For more information, contact: Gerald Selzer, National Science Foundation Directorate for Biological, Behavioral and Social Sciences, 1800 G Street, NW, Washington, D.C. 20550 [202-357-7652].
Institutional Dental Grants
The National Institute of Dental Research (NIDR) awards National Research Service Award (NRSA) institutional grants to eligible institutions to develop or enhance research training opportunities for qualified individuals of the institution's selection who seek to prepare for careers in biomedical and behavioral oral health research. The purpose of the NRSA program is to help ensure that highly trained scientific manpower will be available in adequate numbers and in the appropriate research areas and fields to maintain the nation's biomedical and behavioral oral health research agenda.
Applications will be accepted for research training at the predoctoral level and specialized basic and/or clinical science training at the postdoctoral level. Research training grants are a useful mechanism for the postdoctoral training of dentists whose training usually provided only limited research experience. Priority will be given to programs that offer dentists the opportunity to obtain a Ph.D. degree. However, in certain cases, proposals offering training as part of a master's degree program will be acceptable. Preapplication consultation with NIDR is highly desirable.
For additional information contact: Thomas M. Valega, Ph.D., Special Assistant for Manpower Development and Training, NIDR, NIH, Westwood Building, Room 510, Bethesda, MD 20892 [301-496-6324].
Diabetes Research
Two grants are available to help fund diabetes and endocrinology research centers. A diabetes center must be an identifiable unit within a single university medical center or a consortium of cooperating institutions. An existing program in biomedical research on diabetes and related metabolic and endocrine disorders is required.
The deadline for applications is November 12, 1990. For more information contact: Dr. Sanford Garfield, Diabetes Centers Program Director, Division of Diabetes, Endocrinology and Metabolic Diseases, Westwood Bldg., Rm. 626, NIDDKD, Bethesda, MD 20892 [301-496-7418].
* * *
The Kennedy Institute of Ethics, Georgetown University, announces a course entitled "Ethical Issues of Animal Experimentation", to be held on campus, Washington, DC, March 24-28, 1991. The purpose of this academic course is to address ethical issues, convey information, and provide a forum for discussion. The course is aimed at members of Institutional Animal Care and Use Committees, biomedical scientists, philosophers, bioethicists, administrative officials, members of the public, and other interested persons.
The topics to be addressed include the moral standing of animals, rights and obligations, the application of moral standing to individual species, the roles of animal pain and suffering, trading harms and benefits, trading species such as primate vs. rat, and government responsibilities, among others. Various viewpoints will be presented. The faculty includes Tom L. Beauchamp, Ph.D., David DeGrazia, Ph.D., Rebecca Dresser, J.D., Ruth R. Faden, M.P.H., Ph.D., R. G. Frey, Ph.D., Franklin Loew, D.V.M., Ph.D., Charles McCarthy, Ph.D., F. Barbara Orlans, Ph.D., Edmund Pellegrino, M.D., Robert M. Veatch, Ph.D., Leroy Walters, Ph.D., Thomas L. Wolfle, D.V.M., Ph.D., and others.
For details, contact Ms. Michelene Sheehy, Course Administrator, Kennedy Inst. of Ethics, Georgetown Univ., Washington, DC 20057 [202-687-6766].
* * *
About 400 primatologists met July 11-14, 1990, at the University of California Davis campus for the 13th annual meeting of the American Society of Primatologists. The keynote address was given by Professor Marian C. Diamond of UC Berkeley, an anatomist who works with rat subjects. Much to the surprise of many of us, her address on the effects of environment and experience on the nervous system was extremely relevant to the efforts we are making to improve the environments of our primate subjects. Other featured speakers were Charles Snowdon (U. Wisconsin), who described his non-invasive investigations of suppression of ovulation in cotton-top tamarins; Gene Sackett (U. Washington), expressing the theory that there is a fundamental cross-purpose between veterinarians and caretakers, who strive to breed the healthiest possible animals, and researchers, who need animal models of human diseases and abnormalities; and Donald Lindburg (San Diego Zoo), who spoke of ex-situ propagation of endangered wildlife as a last resort in the face of vanishing habitat.
Symposia were held on the Socio-ecology of juvenile primates; Gorilla cognition and communication and its development; Rank and reproductive success; Genetic research on nonhuman primates: Issues in biology and management; Mechanisms of vocal communication in primates; and Psychological perspectives in primatology (a symposium in tribute to Gary Mitchell). A workshop was held on Invasive and noninvasive applications of ultrasound in macaques, and another, in conjunction with the Association of Primate Veterinarians (who met in Sacramento 14-16 July), on USDA standards for outdoor primate facilities. Several films and videos were shown one evening, and there were 50 posters presented in two sessions. There were also 10 sessions of contributed papers on various subjects. Abstracts of the presentations have been published in the American Journal of Primatology, 1990, 20 [3].
Four students were selected as winners of the Student Presentation Awards. C. Drea (Emory U.) and M. Libet (UC Davis) presented papers entitled, respectively, "Color discrimination and color reversal acquisition in group-living rhesus monkeys," and "Differential tolerance across rank groups during social feeding in a captive group of rhesus monkeys." L. Digby (UC Davis) and T. Justice (Wake Forest U.) presented posters on "An experimental test of dispersal choices in Callithrix jacchus," and "Changes in the interfemale social behavior of Macaca fascicularis over the menstrual cycle."
Officers were elected for the next 2 years: President, Joyce Sirianni; President-elect, Rich Rawlins; Past President, Bill Mason; Executive Secretary, Nancy Caine; and Treasurer, Jeff French.
The 14th annual meeting will be held jointly with the Mexican Primate Society on June 24-28, 1991, in Vera Cruz, Mexico.
* * *
The Caribbean Primate Research Center Museum maintains a collection of primate skeletons and alcohol-preserved tissue specimens. The skeletal collection consists of approximately 2000 complete or nearly complete primate skeletons, predominately Macaca mulatta and Erythrocebus patas. The alcohol-preserved tissue specimens were obtained from necropsy of these primates. Support data such as matrilines (of Cayo Santiago M. mulatta), age, sex, and medical histories are available for most of the specimens. All specimens are available for study on site by qualified scientists. Interested investigators should write a letter of inquiry for further information regarding the collection to: Dr. Matt Kessler, Director, Caribbean Primate Research Center, University of Puerto Rico, Medical Sciences Campus, P.O. Box 1053, Sabana Seca, PR 00749.
* * *
W. R. Kingston
A symposium, "Beyond Captive Breeding: Reintroducing Endangered Species to the Wild," was held by the Zoological Society of London on November 24-25, 1989. The proceedings of this symposium have not yet been published, but I have heard that, according to a paper presented by Kleiman and Beck (in press), costs to date of the Golden Lion Tamarin Conservation Project have amounted to some $23,000 per captive bred animal released into the reserve. There is no doubt that the sustained and dedicated efforts of Dr. Kleiman and others have achieved excellent results, especially in captive breeding. Since breeding stock, almost entirely from already captive sources, had to be transported almost worldwide to make up genetically suitable matings, it is not at all surprising that costs have been very high. However, at least two other things are equally obvious.
First, if these are the costs of breeding and successfully reintroducing one very small, relatively prolific, and free-breeding primate, then there is a poor prospect for doing many other Callitrichids, let alone much larger primates such as Brachyteles, Chiropotes, and Uakaris. Second, fund raising from the general public by conservation organizations is bound to be depressed by these figures, considering the efforts that go into raising a few hundred dollars by flag days and coffee mornings, which is how much of the donations are raised in the U.K. The high costs of a black footed ferret conservation project have already raised considerable comment here.
I have presented and published papers (e.g., Kingston, 1972) based on my experiences as chief animal technician for Fison Pharmaceuticals, managing the PAHO/NIH primate breeding and conservation center in Iquitos, Peru from 1976 to 1979, and as a consultant for the Centro Nacional de Primatas in Belem, Brazil, from 1980 to 1985. These activities included the breeding of callitrichids on a large scale, but working within the limits of a very restricted budget. This practical experience demonstrated 1) that very suitable breeding facilities could be constructed in South America at relatively low cost; 2) that breeding stock could be trapped quickly and inexpensively using locally recruited staff; and 3) that very suitable low cost diets could be developed from locally available materials. These conclusions are set out in more detail elsewhere (Kingston, 1983, 1986). My personal experience includes 12 species of Callitrichidae as well as a number of Cebids, but does not include Leontopithecus. However, in conversation with a number of people working with this genus it appears that they are certainly no more difficult than some Saguinus species. I should point out that I had a very humiliating experience with Saguinus mystax in Peru, in which 100 pairs failed to produce any young whatsoever, although several other species identically housed and managed bred freely!
There are a number of callitrichids, including of course the other two Leontopithecus species, on the endangered list. These include Callithrix aurita, C. flaviceps, C. argentata leucippe, and C. humeralifer chrysoleuca. There is absolutely no reason to suppose that these would prove any more difficult than C. jacchus, C. argentata argentata, etc., all of which breed freely and easily. Endangered Saguinus include S. bicolor, S. imperator, and S. leucopus and, although this genus is a little more difficult, these should prove no more difficult than S. oedipus and fuscicollis. Even my personal bête noire, S. mystax, has bred reasonably in other hands.
All the above could be bred in adequate numbers with reasonable genetic diversity to ensure the survival of the genome if the project is tackled in an efficient, straightforward manner. It should be done in the country of origin of the species and, if at all possible, in the locality in which it is hoped to reintroduce captive bred stock. At least twenty pairs should be trapped and housed in simple, reasonable sized cages with provision for the all-important facility of being able to catch the animals without risk of injury, particularly to carried offspring.
The actual work should be done by local people with the help of one experienced advisor, through whom the funding agency should keep control of funds provided. When a reasonable number of captive bred young are available, suitable unrelated pairs should be distributed to appropriate institutions abroad, which should certainly pay the costs of transport if not an actual price -- after all, primates of all kinds make excellent exhibits! This should ensure a position similar to that achieved with the Golden Lion Tamarins: an adequate pool of breeding stock to allow for expansion if suitable secure habitat becomes available. The reduced costs would make it possible to deal with many more species, even the larger primates which are much more expensive to cage and feed, and which reproduce more slowly. Undeniably more and more species are going to become endangered; funding is always going to be a problem; and no one wants to lose a primate species. If the ideal of secure natural habitat for all species in not immediately attainable it is surely better to have a viable nucleus living in captivity than to lose the species altogether.
I believe that any primate can be successfully kept and bred in captivity. The supposedly difficult cebids -- Alouatta, Chiropotes, Pithecia, and Cacajao -- are breeding in a number of centers and zoos, as are the Colobus and Presbytis of the Old World. The lemurs of Madagascar could certainly be bred under the conditions I describe and survival of at least a viable nucleus of all species assured.
With regard to reintroduction into the wild of captive bred animals, I would suggest the following: young animals, kept with their parents until they have had experience helping raise siblings, should be released into a good-sized wire enclosure sited within an "island" of natural habitat formed by clear felling a suitably wide strip all around it. After a reasonable interval the enclosure, which is provisioned of course, should be opened to allow the animals free range within the isolated area. Provisioning should be continued for a while, during which the very rapid growth of vegetation in the cleared area will soon allow the animals to disperse into the reserve itself.
References
Kingston, W. R. (1972). The breeding of endangered species of marmosets and tamarins. In D. Bridgwater (Ed.), Saving the Lion Marmoset (pp. 86-91).
Kingston, W. R. (1975). The cost of developing and managing a marmoset colony. In D. G. Kleiman (Ed.), The Biology and Conservation of the Callitrichidae. (pp. 311-315). Washington: Smithsonian Institution.
Kingston, W. R., & Muniz, J. A. P. C. (1983). Preliminary report on the establishment in Brazil of a breeding colony of marmosets (Callithrix humeralifer humalifer). Laboratory Primate Newsletter, 22 [4], 1-2.
Kingston, W. R. (1986). Captive breeding of endangered species. Primate Eye, 30, 27-30.
Kleiman, D., & Beck, B. (In press). Reintroduction of New World primates, with emphasis on golden lion tamarins.
-------------------------------------------------------------------
Author's address: The Old Smithy, Bishops Frome, Worcester WR6 5BA, England.
* * *
M. B. Ballinger, Animal Health & Services, D-403, AP-13, Abbott Labs, Abbott Park, IL 60064.
Bernadette Bresard, La Grande Planche, 18410 Argent sur Sauldre, France.
Arnold Chamove, Dept. of Psychology, Massey Univ., Palmerson, North New Zealand.
Maribeth Champoux, NIH Animal Center, P.O. Box 289, Poolesville, MD 20837.
Kevin L. Clark, 1486 N. High St., Apt. C, Columbus, OH 43201.
A. Susan Clarke, Harlow Primate Laboratory, 22 N. Charter Street, Madison, WI 53715.
CLEA Japan, Inc., No. 2 Inari Bldg., 20-14, Aobadai-2, Meguko-ku, Tokyo, Japan.
Col. William C. Cole, Deputy Commander, Letterman Army Inst. of Research, Presidio of San Francisco, CA 94129-6800.
Robert W. Cooper, Monkey Jungle, 14805 S.W. 216th St., Miami, FL 33170.
Gerald W. Cortright, 1350 W. Bethune Ave., Apt. 2002, Detroit, MI 48202.
Harold Davis, USAS/SAMUSP, Brooks Air Force Base, San Antonio, TX 78235.
Judith A. Davis, 12423 Loft Lane, Silver Spring, MD 20904.
Robert C. Dysko, Unit for Lab. Animal Medicine, 018 ARF, Box 0614, Univ. of Michigan School of Medicine, Ann Arbor, MI 48109.
Dorothy Fragaszy, Psychology Dept., Univ. of Georgia, Athens, GA 30602.
Laura A. Gillespie, 352 N. Summit Ave., Gaithersburg, MD 20877.
Joe Held, 1300 Crystal Dr. #505, Arlington, VA 22202-3234.
D. Helwig, 56 Glenview Dr., San Francisco, CA 94131.
Billy W. Howard, 2814D Buttonwood Court, Aberdeen Proving Ground, MD 21005.
Janis Joslin, Animal Health Dept., Woodland Park Zoo, 5500 Phinney Avenue North, Seattle, WA 98103.
Barbara A. Kintner, Laboratory Animal Care Unit, Michigan Dept. of Public Health, P.O. Box 30035, Lansing, MI 48909.
Martin F. Kreite, 12434 Walnut Cove Circle, Germantown, MD 20874.
Randall C. Kyes, Dept. of Comparative Medicine, Bowman Gray School of Medicine, 300 S. Hawthorne Rd., Winston-Salem, NC 27103.
Fred Kyle, Columbia Univ. Inst. Comparative Medicine, Black Bldg, Room 1810, 630 West 168th St., New York, NY 10032.
R. M. Letscher, 319 Cave Lane, San Antonio, TX 78209.
Peter Leung and K. Luke, 1015 Tower A, Hunghom Commercial Centre, 39 Matauwai Road, Hunghom, Kowloon, Hong Kong.
Thomas Mayer, UNSMRU3, P.O. Box 103, FPO NY 09527-1600.
Daniel R. Michalowski, Director, Seneca Park Zoo, 2222 St. Paul St., Rochester, NY 14621-1097.
Kathleen N. Morgan, Dept. of Psychology, Cal. State Univ., 4400 Univ. Parkway, San Bernardino, CA 92407.
Michael J. Parnell, Rocky Mountain Labs., 903 S. Fourth St., Hamilton, MT 59840.
Theopolis Peace, 114 Rollingwood Dr., San Rafael, CA 94901.
David K. Peters, Univ. of Missouri, Laboratory Animal Center, 1015 East 50th Street, Kansas City, MO 64110.
Shannon Pinkston, Psychology Dept., Univ. of Georgia, Athens, GA 30602.
James A. Porter, P.O. Box 970768, Miami, FL 33197.
Michael Rand, Univ. Animal Care, College of Medicine, Health Sciences Center, Univ. of Arizona, Tucson, AZ 85724.
William Reid, Animal Research Facility, P.O. Box 0614, Univ. of Michigan Medical School, Ann Arbor, MI 48109.
Reed W. Rings, ARF, UNMC, 600 S. 42nd Street, Omaha, NE 68198-6385.
Joe Safron, Sinclair Comparative Medicine Research Farm, R.R. 15, 5650 South Sinclair Road, Columbia, MO 65304-9197.
Maj. Creighton J. Trahan, USUHS, Dept. Lab. Animal Medicine, 4301 Jones Bridge Rd., Bethesda, MD 20814-4799.
Joseph E. Wagner, Dept. of Veterinary Pathology, College of Veterinary Medicine, Univ. of Missouri, Columbia, MO 65211.
J. H. Wong, Assoc. Dir. of Assessments, Canadian Council on Animal Care, 1000-151 Slater St., Ottawa, PO K1P 5H3, Canada.
* * *
Silver Spring Monkeys Euthanized
Three of the so-called Silver Spring monkeys were euthanized for humane reasons on July 6 at Louisiana's Delta Regional Primate Research Center. In ordering the euthanasia, the NIH was following the advice of attending veterinarians and carrying out its responsibilities under the Animal Welfare Act and the Public Health Service Act. The condition of these monkeys had been deteriorating for several years. They had been candidates for euthanasia since 1988, but litigation brought by animal rights groups prevented this action until Federal Court in Washington, DC refused to grant another restraining order sought by the Physicians Committee for Responsible Medicine. An appeal of this decision was denied on July 3.
One other monkey had been euthanized in January (see Laboratory Primate Newsletter, 1990, 29, [2], 12). The remaining four Silver Spring monkeys will stay at Delta in the legal custody of NIH. -- From the NABR Update, 1990, 11[24].
Australian Animal Laws
The National Health and Medical Research Council, Australia's main funding organization for medical research, announced last week a code revision tightening rules for laboratory animal use. According to the revised code, researchers must convince animal ethics committees that alternative techniques are not suitable, justify the species and number of animals to be used, and judge procedures as to whether vertebrate animals experience pain the same way humans do. "If a human needs an anesthetic for a procedure, then it should be assumed the animal will need one, too," said Warwick Anderson, chairman of the group that revised the code. He added that the code provided "the most comprehensive guidelines dealing with this issue in the world." -- From the NABR Update, 1990, 11[26].
IPS Newsletter
Dorothy Fragaszy, Psychology Dept., Univ. of Georgia, Athens, GA 30602, is the new editor of the International Primatological Society's IPS Newsletter.
Four Monkeys Proposed as Endangered
Ten animal species -- four monkeys and six birds -- were proposed by the U.S. Fish and Wildlife Service on January 16, 1990, for listing as Endangered species. If the proposals are approved, these animals will receive the protection available to foreign taxa under the Endangered Species Act (ESA). They are already on Appendix I of the Convention on International Trade in Endangered Species of Wild Fauna and Flora, which restricts commerce in rare animals and plants. Listing them under the ESA would reinforce the prohibition on U.S. import or export of these species without a permit. The Act also authorizes the Service to assist foreign wildlife management agencies in the conservation of listed species by providing training, personnel, and (if available) limited funding.
The monkey species are all Rhinopithecus. R. roxellana, the Sichuan or golden snub-nosed monkey, is found on the southeastern slopes of the Tibetan Plateau in the Chinese provinces of Sichuan, Shaanxi, Hubei, Gansu, and Yunnan; the Yunan or black snub-nosed monkey, R. bieti, occurs in the Yunling Range of Tibet and Yunnan; R. brelichi, the Guizhou or gray snub-nosed monkey, is native to the Fanjin Range in the Chinese province of Guizhou; and the Tonkin snub-nosed monkey, R. avunculus, is found in northern Viet Nam. -- From the Endangered Species Technical Bulletin, 1990, 15[2], 4.
Florida Key Monkeys Upset the Neighbors
Residents of the Florida Keys are fed up with Bausch and Lomb's monkey business. The vision-care giant put thousands of monkeys on two small, deserted islands more than a decade ago to breed, offering a steady supply of laboratory animals to biomedical researchers. But people who live near the islands where the rhesus monkeys have been allowed to run free object to what they deem needless environmental destruction.
The rhesus monkeys have been good business since they were brought over from India 18 years ago. Handlers regularly dispense monkey chow and capture more than a thousand each year, generating more than $1.5 million in annual sales.
The breeding colonies became more important to U.S. researchers after India, Pakistan, and Bangladesh banned monkey exports in the late 1970s, said Dr. Joseph Held, vice president for primate research at Charles River Laboratories, the division of Bausch and Lomb that runs the islands. And because they are free of Herpes B virus and other infections from the outside world, they are much more valuable than other primates destined for the nation's laboratories.
More than 1900 breeding animals and several thousand of their offspring roam on 300 acres of partially submerged wetlands on Key Lois and Raccoon Key. Even Charles River Labs agrees that the monkeys have made a mess of things. The company has agreed to pay for replacing thousands of red mangroves, which the monkeys stripped of leaves, and has begun refoliating the islands. The company also plans to gradually phase down the colony operations, and eventually transfer the monkeys elsewhere. Meanwhile, although the water around Key Lois has been fouled, leaving a band of yellowish-brown algae, the company routinely tests the water quality, and states that there is no health hazard. -- [From an A.P. report by Michael Warren, August 21, 1990.]
Chimpanzee Classification
Details on the reclassification of the chimpanzee and pygmy chimpanzee have been set forth in the March 12, 1990, Federal Register. Under the new ruling, all populations of Pan paniscus are listed as Endangered. In the case of P. troglodytes, however, captive populations will remain classified as Threatened, and individuals of that species held legally in the United States will continue to be covered by a special regulation that allows their use for certain activities (such as medical research) that are otherwise prohibited.
* * *
IAMAT
The International Association for Medical Assistance to Travellers (IAMAT), a division of the Foundation for the Support of International Medical Training, Inc., offers its members several interesting publications. This year members received a directory of English-speaking physicians in 138 countries, all of whom have agreed to a set payment schedule for IAMAT members, and who assure qualified medical assistance 24 hours a day. Also included were pamphlets on malaria, and a world immunization chart. IAMAT is now offering for sale, at their own cost of $75 plus postage, a 5-pound, easy-to-assemble, bed net. Membership in IAMAT is free, but donations are welcomed. For information, contact IAMAT, 417 Center Street, Lewiston, NY 14092 [716-754-4883].
WRPRC Publication
The Wisconsin Regional Primate Research Center is publishing a quarterly newsletter, called Centerline, of research and personnel notes, along with news items. The address of the Center is 1223 Capitol Court, Madison, WI 53715.
Primate Exporter
In keeping with our long-term policy of mentioning, but not recommending, businesses which send us announcements, we pass on the news that the Katabi Export & Trading Co., P.O. Box 1049, Dar es Salaam, Tanzania, has introduced themselves as exporters of vervet monkeys and baboons to the United States. Mr. A. B. Mtuga, the Director, would like to contact importers. Their Telex number is 41207 TPTC TZ Attention Mtuga; FAX number is 34794.
CDC Hotline
CDC's Traveler's Health Section, Division of Quarantine, Center for Prevention Services, now has a 24-hour-a-day automated telephone system that provides information for international travelers on vaccine requirements and recommendations by geographic area. Menu options include information on malaria, food and water precautions, travelers' diarrhea, immunizations for children less than 2 years of age, pregnant travelers, and disease outbreaks. To access this information, call 404-332-4559.
* * *
Humane Care and Use of Laboratory Animals
The National Institutes of Health (NIH), Office for Protection from Research Risks (OPRR), will sponsor four workshops on implementing the Public Health Service Policy on Humane Care and Use of Laboratory Animals. Each of the four workshops scheduled will focus on a specific topic. The workshops are open to institutional administrators, members of Institutional Animal Care and Use Committees, laboratory animal veterinarians, investigators and other institutional staff who have responsibility for high-quality management of sound institutional animal care and use programs. For overall information on the series, contact Mrs. Roberta Sonneborn, Executive Assistant for Animal Welfare Education, OPRR, NIH, Building 31, Room 5B59, 9000 Rockville Pike, Bethesda, MD 20892 [301-496-7163]. The workshops are as follows:
"Problems of Small vs. Large Institutions", December 3-4, 1990, University of Rhode Island, 36 Upper College Road, Kingston, RI 02881. Contact URI Conference Center [401-792-2170] or Mr. Kevin McAndrews [401-792-2833].
"Surgery and Post-surgical Care", April 4-5, 1991, Medical University of South Carolina, ATTN: Carol Reed, Registration Coordinator, Dept. of Comparative Medicine, 704 BSB, Charleston, SC 29425-2216. Contact Dr. M. Michael Swindle [803-792-3625].
"Recurrent Controversies in Protocol Review", May 2-3, 1991, Washington University School of Medicine, Continuing Medical Education, Box 8063, 660 South Euclid, St. Louis, MO 63110. Contact Ms. Loretta Giacoletto [314-362-6891 or 800-325-9862].
"Resolving the Ethical Dilemmas in Animal Protocol Review", September 12-13, 1991, University of Washington, Dept. of Comparative Medicine, Box SB-42, Seattle, WA 98195. Contact Ms. Gail Woltz [206-543-9678].
Primates in Evolution
John Napier's students, colleagues, and friends are taking the opportunity of a memorial meeting in his honor to come together in London, December 17-18, for workshop and symposium sessions to review and assess the field of primatology today. A major purpose of the meeting will be to discuss the continuing role of primatology in the 21st century in the areas which concerned John, such as behavior, comparative anatomy, and evolution. The meetings will also be the occasion of the Osman Hill Memorial Lecture, sponsored by the Primate Society of Great Britain, to be delivered by Professor R. D. Martin, and the L. S. B. Leakey Memorial Lecture, sponsored by the L. S. B. Leakey Trust, to be delivered by Professor Alan Walker. For further information and registration contact Dr. L. C. Aiello, Department of Anthropology, University College London, Gower Street, London WC1E 6BT, UK.
XXIV World Veterinary Congress
"The Veterinary Profession and North-South Cooperation in Benefit of Mankind" is the title of the XXIV World Veterinary Congress, to be held August 18-23, 1991, in Rio de Janeiro, Brazil. Topics will include animal welfare, advances in knowledge on animal production, public health, and the environment. For further information, contact the President of the Central Organizing Committee, Joselio de Andrade Moura, SCS-Ed. Ceará -- 14.0 Andar, Brasilia DF, Brazil.
Effect of Regulations on Animal Welfare
A conference on the Effect of Current Regulations on Laboratory Animal Welfare will be co-sponsored by the Scientists Center for Animal Welfare (SCAW) and the Johns Hopkins University School of Medicine, October 29, 1990, at the Tremont Plaza Hotel, Baltimore, MD. The purpose is to explore current issues of concern and responsibility for members of Animal Care and Use Committees (ACUCs), with particular emphasis on current and changing regulations. Workshops will be on AAALAC accreditation, Employee training programs, ACUC community members, Educational materials for IACUCs, and How to develop an IACUC at a small institution. For information, contact Lee Krulisch, Executive Director, SCAW, 4805 St. Elmo Ave., Bethesda, MD 20814 [301-654-6390] by October 22.
* * *
Books
*Understanding chimpanzees. P. G. Heltne & L. A. Marquardt
(Eds.). Cambridge: Harvard University Press, 1989. 407 pp.
[Price: $40.00]
. . Papers presented at the
symposium "Understanding Chimpanzees," which was organized
by the Chicago Academy of Sciences and held in November 1986.
Contents:
Current Fieldwork: Pan Troglodytes. Gombe.
Gombe: Highlights and current research, by J. Goodall.
Chimpanzee use of medicinal leaves, by R. W. Wrangham & J. Goodall.
Methods for isolating Chimpanzee vocal communication, by C. Boehm.
The research at Gombe: its influence on human knowledge, by R. S. Fouts.
Mahale Mountains.
Research at Mahale, by T. Nishida.
Social interactions between resident and immigrant female
chimpanzees, by T. Nishida. Sexual behavior of immigrant and resident
female chimpanzees at Mahale, by T. Hasegawa. Sex differences in
the behavioral development of chimpanzees at Mahale, by M.
Hiraiwa-Hasegawa.
Additional sites.
Feeding ecology of chimpanzees in the Kibale Forest, Uganda, by G.
Isabirye-Basuta. Recent research on chimpanzees in West Africa, by
W. C. McGrew. Population dynamics of chimpanzees at Bossou,
Guinea, by Y. Sugiyama. The use of stone tools by wild-living
chimpanzees, by A. Kortlandt. ChimpanZoo, by J. Goodall.
Current fieldwork:
Pan paniscus. Introduction: The fourth ape, by F. B. M. de Waal.
Behavioral contrasts between bonobo and chimpanzee, by F. B. M.
de Waal. The sexual behavior of pygmy chimpanzees, by T. Kano.
Developmental retardation and behavioral characteristics of
pygmy chimpanzees, by S. Kuroda. Social organization of pygmy
chimpanzees, by F. J. White.
The Chimpanzee Mind.
Are animals intelligent? Wolfgang Köhler's approach, by
E. W. Menzel, Jr. Cross-fostered chimpanzees: I. Testing
vocabulary, by R. A. Gardner and B. T. Gardner. Cross-fostered
chimpanzees: II. Modulation of meaning, by B. T. Gardner and
R. A. Gardner.
Signing interactions between mother and infant chimpanzees, by
D. H. Fouts. Spontaneous pattern construction in a chimpanzee,
by T. Matsuzawa. Symbol acquisition and use by
Pan troglodytes, Pan paniscus, Homo sapiens, by S. Savage-Rumbaugh,
M. A. Romski, W. D. Hopkins, & R. A. Sevcik. Current and future research
on chimpanzee intellect, by D. M. Rumbaugh.
Chimpanzee Conservation.
Population status of wild chimpanzees
(Pan troglodytes) and threats to survival, by G. Teleki.
Demography of chimpanzees in captivity, by K. Latinen.
Area status report: Tanzania, by J. Goodall. Conservation status of
Pan paniscus,
by R. K. Malenky, N. Thompson-Handler, & R. L. Susman.
Evolution and Epilogue.
Hominoid sociobiology and hominid social evolution, by M. P.
Ghiglieri. Epilogue: Understanding chimpanzees and bonobos,
understanding ourselves, by P. G. Heltne.
*Perspectives in Primate Biology, Volumes 2 and 3. P. K. Seth &
S. Seth (Eds.). New Delhi: Today & Tomorrow's Printers & Publishers,
1989. Vol. 2: 208 pp. [$69]; Vol. 3: 260 pp. [$69].
. . Volume 2, Contents: Evolutionary radiations in humans and great
apes: Some quantitative evidence, by C. Oxnard. Classification,
distribution and ecology of Indian macaques, by J. Fooden. Quantitative
and experimental approaches to primate locomotion: A review of recent
advances, by F. K. Jouffroy. The morphology, variability and function of
the teeth of extant primates, by D. R. Swindler. Primate relationships:
social dispositions and physiological responses, by S. P. Mendoza & W.
A. Mason. Blood group serology of apes and monkeys, by W. W. Socha.
Chromosomal evolution in Malagasy prosimians, by Y. Rumpler. Functional
differences between serum protein coded by codominant alleles at the
transferrin and albumin loci in
Macaca mulatta, by D. G. Smith, F. W. Lorey, B. K. Rolfs, & C. E.
Ahlfors.
. . Volume 3, Contents: A review of variables affecting primate
socialization, by F. E. Poirier. Food-sharing in primates: A critical
review, by A. T. C. Feistner & W. C. McGrew. Sexual behaviour of the
great apes, by R. D. Nadler & J. F. Dahl. Primate social signals:
Structure and function, by M. Maürus. Ecology and behaviour of the
Malagasy primates, by I. Tattersall & R. W. Sussman. Bringing
experimentation to the field of primate vocalisation, by R. W. Byrne.
The structure and usage of long calls in marmosets and tamarins, by C.
T. Snowdon. Cross-specific review of Asian colobine social organization
and certain behaviours, by F. E. Poirier & M. C. Kanner. Behaviour of
the genus
Papio: Ethogram, taxonomy, methods, and comparative measures, by A.
M. Coelho, Jr. & C. A. Bramblett. Social behaviour of the lesser apes,
by D. J. Chivers. Social structure and dynamics of the chimpanzee: A
review, by T. Nishida. Field studies of wild orang-utans: Current state
of knowledge, by J. R. MacKinnon. A global overview of primate
conservation, by R. A. Mittermeier. Behavioural phylogeny of rhesus
monkeys in India, by P. K. Seth, S. Seth, P. K. Chopra, & G. J. Reddy.
The successful captive breeding of endangered primates, by C. T.
Snowdon.
*Medical Management of the Orangutan. S. K. Wells, E. L. Sargent, M. E. Andrews, & D. E. Anderson. New Orleans: Audubon Park Zoo, 1990. 215 pp. [Price: $20. Order from Susan Wells, Audubon Park Zoo, 6500 Magazine St., New Orleans, LA 70118]
Bibliographies
*A Jubilee Bibliography, 1963-1988. Compiled by C. M. Perkins. Jersey: Wildlife Preservation Trust. (Systemic and 8 other indexes) [Jersey Zoological Park, Jersey, Channel Islands, UK]
Bulletins
*ICLAS Bulletin, Number 66, Spring, 1990. [2.50 Pounds from UFAW, 8
Hamilton Close, North Mimms, Potters Bar, Herts. EN6 3QD, UK]
. . This issue includes articles on "Laboratory animal management
around the world," by D. Jol, and "The status of laboratory animal
management in Kenya," by M. A. Suleman.
Directories
*Directory of the Consortium of Aquariums, Universities and
Zoos. 1990-91.
[Price: $15. Order from D. Hardy, Dept. of Psychology,
California State Univ., Northridge, CA 91330]
. . Names, addresses, and specific interests and projects of 624
individuals from 300 institutions.
Newsletters and Reports
*Research Resources Reporter, 1990, 14 [3]. (Published by the
Division of Research Resources, NIH)
. . This issue includes a report from the Yerkes Regional Primate
Research Center, in which M. E. Wilson describes how rhesus monkeys
raised under controlled indoor conditions mature more rapidly, with
accelerated sexual and bone maturation, than those raised outdoors.
*Research Resources Reporter, 1990, 14 [4]. (Same publisher as
above)
. . This issue includes an article by J. P. Swyers entitled "Animal
models offer hope for AIDS vaccines," summarizing work at the Delta and
New England Regional Primate Research Centers.
*Research Resources Reporter, 1990, 14 [6]. (Published by the
National Center for Research Resources, NIH)
. . This issue includes an article by J. P. Swyers entitled
"Successful breeding program benefits chimpanzee conservation and
research," describing the National Chimpanzee Breeding and Research
Program.
*Gorilla Conservation News. No. 4, 1990 [K. J. Stewart, Dept.
of Anthropology, Univ. of California, Davis, CA 95616].
. . The newsletter of the Gorilla Advisory Committee of the Primate
Specialist Group, International Union for the Conservation of Nature and
Natural Resources.
*RI PC Annual Report 1989. [151, Lange Kleiweg, P.O. Box 5815,
2280 HV Rijswijk, The Netherlands]
. . The annual report of the Radiobiological Institute and the TNO
Primate Center, which in the future will be united as the Institute of
Applied Radiobiology and Immunology TNO. Of the many short notes
describing the accomplishments of the organizations, 18 refer to primate
models and subjects, with special emphasis on virology, retrovirology,
and autoimmune diseases.
*The 60th Year: 1989-1990, Reflecting on the Past and Building
on the Future. Yerkes Regional Primate Research Center (Emory
Univ., Atlanta, GA 30322).
. . The emphasis of this report is on the benefits to humans of
primate medical and behavioral research.
Monographs
*Environmental Enrichment. The Response of Captive Orang Utans to Pictures. P. McEwen. Hertfordshire: UFAW, 1989. 16pp. [UFAW, 8 Hamilton Close, South Mimms, Potters Bar, Herts EN6 3QD, UK]
Special Journal Issues
*New World Monkeys. Journal of Human Evolution, 1989, 18.
. . This issue, dedicated to Dr. Adelmar Coimbra-Filho, will be
followed by an issue on the platyrrhine fossil record. Contents:
Neotropical primate conservation, by R. A. Mittermeier, W. G. Kinzey, &
R. B. Mast. Vocal communication in New World monkeys, by C. T. Snowdon.
The nocturnal primate niche in the New World, by P. C. Wright. The
positional behavior and substrate use of squirrel monkeys: ecological
implications, by S. Boinski. Sympatric Brazilian callitrichids: The
black tufted-ear marmoset,
Callithrix kuhli, and the golden-headed lion tamarin,
Leontopithecus chrysomelas, by A. B. Rylands. Comparative feeding
ecology of the uakari and bearded saki,
Cacajao and Chiropotes, by J. M. Ayres. Adaptive radiation of the
ateline primates, by A. L. Rosenberger & K. B. Strier.
*Nonhuman primate models for AIDS II.
Journal of Medical Primatology, 1990, 19 [3/4].
. . A selection of papers from the 6th annual meeting on nonhuman
primate models for AIDS, held at the Southwest Foundation for Biomedical
Research at San Antonio, TX, 1988. Contents: Nonhuman primate models for
acquired immunodeficiency syndrome, by M. K. Axthelm & S. M. Shiigi. The
simian immunodeficiency virus (SIV) rev gene regulates env expression,
by S.-M. Cheng, M. Blume, S.-G. Lee, P. P. Hung, V. M. Hirsch, & P. R.
Johnson. Cellular immune response of SIV-infected rhesus macaques, by T.
P. McGraw, B. R. Vowels, M. E. Gershwin, & M. B. Gardner.
Mesenchymoproliferative disorders (MPD) in simian AIDS associated with
SRV-2 infection, by C. C. Tsai, C.-C. Tsai, S. T. Roodman, & M.-D. Woon.
Maternal transmission of type D simian retrovirus (SRV-2) in pigtailed
macaques, by C.-C. Tsai, K. E. Follis, K. Snyder, S. Windsor, M. E.
Thouless, L. Kuller, & W. R. Morton. Mutational analysis of simian
immunodeficiency virus from African green monkeys and human
immunodeficiency virus type 2, by R. Shibata, A. Adachi, H. Sakai, A.
Ishimoto, T. Miura, & M. Hayami. Identification of SIV/SMM viral
proteins that induce T cell response in experimentally infected rhesus
macaques and naturally infected sooty mangabeys by the cellular western
blot assay, by J. D. Powell, F. Villinger, T. Yehuda-Cohen, M.
Vuchetich, H. M. McClure, K. W. Sell, & A. Ahmed-Ansari.
*Adaptation and adaptability of Capuchin monkeys.
Folia Primatologica, 1990, 54 [3-4].
. . Most of these papers are from a symposium at the IPS meeting in
Brasilia, 1988. Contents: Variability and adaptability in the genus
Cebus, by D. Fragaszy, E. Visalberghi, & J. G. Robinson. Early
behavioral development in Capuchins
(Cebus), by D. Fragaszy. Conceptual learning in Capuchin monkeys,
by L. E. Adams-Curtis. Use of objects as hammers to open nuts by
Capuchin monkeys
(Cebus apella) , by J. R. Anderson. Tool use in
Cebus, by E. Visalberghi. Social processes affecting the appearance
of innovative behaviors in Capuchin monkeys, by D. Fragaszy & E.
Visalberghi. Significance of kin relations and individual preferences in
the social behaviour of
Cebus apella, by C. Welker, H. Höhmann, & C. Schäfer-Witt.
Infanticide in wedge-capped Capuchin monkeys,
Cebus olivaceus, by X. Valderrama, S. Srikosamatara, & J. G.
Robinson. Dietary differences between neighboring
Cebus capucinus groups: Local traditions, food availability or
responses to food profitability? by C. A. Chapman & L. M. Fedigan.
Dietary variability in
Cebus apella in extreme habitats: Evidence for adaptability, by A.
D. Brown & G. E. Zunino. Vertebrate predation in
Cebus capucinus: Meat eating in a neotropical monkey, by L. M.
Fedigan. Cytogenetic variability within and across populations of
Cebus apella in Argentina, by M. D. Mudry. A comparative study of
neonatal skeletal development in
Cebus and other primates, by E. S. Watts. Scientific films on
Capuchin monkeys, by K. Izawa & E. Visalberghi.
*Special issue: Ecological and behavioral correlates of polyspecific
primate troops.
American Journal of Primatology, 1990, 21 [2].
. . Papers from a symposium at the IPS meeting in Brasilia, 1988.
Contents: Introductory remarks: Ecological and behavioral correlates of
polyspecific primate troops, by M. A. Norconk. Mixed flocks and
polyspecific associations: Costs and benefits of mixed groups to birds
and monkeys, by J. Terborgh. Mixed-species association of east African
guenons: General patterns or specific examples, by M. Cords.
Interspecific relations in a mixed-species troop of moustached tamarins,
Saguinus mystax, and saddle-back tamarins, Saguinus fuscicollis
(Platyrrhini: Callitrichidae), at the Rio Blanco, Peruvian Amazonia, by
E. W. Heymann. Association between olive colobus
(Procolobus verus), Diana guenons (Cercopithecus diana),
and other forest monkeys in Sierra Leone, by J. F. Oates & G. H.
Whitesides. Effects of mixed-species association on resource use by
Saimiri sciureus and Cebus apella, by R. D. Podolsky. Mechanisms
promoting stability in mixed
Saguinus mystax and S. fuscicollis troops, by M. A. Norconk.
*Primate socioecology, communication, and hominid evolution, Part I.
International Journal of Primatology, 1990, 11 [1].
. . Papers from a symposium presented at the 86th annual meeting of
the American Anthropological Association, Chicago, IL. Contents:
Introduction: Current studies on primate socioecology and evolution, by
I. DeVore. New World primates, new frontiers: Insights from the woolly
spider monkey, or muriqui
(Brachyteles arachnoides), by K. B. Strier. Ecology of gorillas and
its relation to female transfer in mountain gorillas, by D. P. Watts.
Fission-fusion social organization in
Ateles and Pan, by M. M. Symington. A comparative perspective on
the evolution of tamarin and marmoset social systems, by A. W. Goldizen.
*Annual Resource Guide 1990. Special Edition of Continuing
Listings. [Primate Supply Information Clearinghouse, PIC,
RPRC, SJ-50, Univ. of Washington, Seattle, WA 98195]
. . Primates, tissues, products, equipment, laboratories, and services,
needed or available on a long-term basis, are listed.
Videotapes
*Training Corral-living Rhesus Monkeys for Fecal and Blood
Sample Collection. M. R. Clarke, K. M. Phillippi, J. A. Falkenstein,
E. A. Moran, & S. J. Suomi. 1990. [Price: $20 from Delta R.P.R.C.,
Tulane Univ., Three Rivers Rd., Covington, LA 70433]
. . A training film, showing in detail the steps leading to successful
collection.
*Captive Chimpanzee Care and Enrichment: A Day in the Life. M.
D. Bodamer, R. S. Fouts, D. H. Fouts, & D. Shipley (Dept. of Psychology,
Central Washington Univ., Ellensburg, WA 98926).
. . Demonstrations of the environment and activities provided at CWU.
*Conduct of a Field Course in Primate Social Ecology and Student
Assistance in the Collection of Quantitative Data. D. R. Rasmussen,
L. Guss, J. A. Dubois, & L. Marsh.
.br;*"Proyecto Titi:" An Effective Conservation Education Program in
Colombia. A. Savage & H. Giraldo.
. . These 40 minute videos may be borrowed for classroom use from
the Library of the Wisconsin Regional Primate Research Center.
Animal Models
*L-dopa pharmacokinetics in plasma and cisternal and lumbar
cerebrospinal fluid of monkeys. Hammerstad, J. P., Woodward, W. R.,
Gliessman, P., Boucher, B., & Nutt, J. G. (Dept. of Neurology, L226,
Oregon Health Sciences Univ., 3181 S.W. Sam Jackson Park Rd., Portland,
OR 97201).
Annals of Neurology, 1990, 27, 495-499.
. . The pharmacokinetics of L-dopa in cisternal CSF are similar to
those in plasma and are likely to reflect those in the brain, while
those in the lumbar CSF are different.
*Symptomatic and asymptomatic
1-methyl-4-
phenyl-1,2,3,6-tetrahydropyridine-treated primates:
Biochemical changes in striatal regions. Elsworth, J. D., Deutch, A. Y.,
Redmond, D. E., Jr., Taylor, J. R., Sladek, J. R., Jr., & Roth, R. H.
(Dept. of Pharmacology, Yale Univ. School of Medicine, 333 Cedar St.,
New Haven, CT 06510).
Neuroscience, 1989, 33, 323-331.
. . Vervet monkeys treated with MPTP fell into symptomatic and
asymptomatic groups. Relatively small, but distinct, differences in
dopamine and homovanillic acid depletions between the groups in some
regions indicates the potential of reversing parkinsonian motor
abnormalities with intrastriatal grafts or implants that produce
relatively low amounts of dopamine.
*Cyclosporin-induced hypertension in marmosets: A new model of
hypertension sensitive to angiotensin-converting enzyme inhibition.
Clozel, J.-P., & Fischli, W. (Pharmaceutical Research Dept., F.
Hoffmann-La Roche, CH-4002 Basel, Switzerland).
Journal of Cardiovascular Pharmacology, 1989, 14, 77-81.
. . Hypertension can consistently be induced in marmosets by chronic
treatment with cyclosporin.
*Oral contraceptives and coronary artery atherosclerosis of
cynomolgus monkeys. Clarkson, T. B., Shively, C. A., Morgan, T. M.,
Koritnik, D. R., Adams, M. R., & Kaplan, J. R. (Arteriosclerosis
Research Center, Bowman Gray School of Medicine, Wake Forest Univ.,
Winston-Salem, NC 27103).
Obstetrics & Gynecology, 1990, 75, 217-222.
. . Three groups of adult female cynomolgus monkeys were fed a
moderately atherogenic diet: a control group, a group given ethinyl
estradiol and norgestrel, and a group given ethinyl estradiol and
ethynodiol diacetate. Both treatments lowered the plasma concentrations
of HDL cholesterol, but the extent of coronary artery atherosclerosis
was also lessened, especially among females at high risk based on their
plasma lipid profiles.
*Transtentorial brain herniation in the monkey: Analysis of brain
stem auditory and somatosensory evoked potentials. Stone, J. L., Ghaly,
R. F., Subramanian, K. S., Roccaforte, P., & Kane, J. (Div. of
Neurosurgery, Cook County Hospital, 1835 W. Harrison St., Chicago, IL
60612).
Neurosurgery, 1990, 26, 26-31.
. . Transtentorial brain herniation (TBH) was developed experimentally
over a 4-hour period. Intracranial pressure rose to twice baseline
value, and there was a 25% increase in Wave V amplitude, 1 hour before
TBH. Analysis of Wave V of the brain stem auditory evoked potentials was
as sensitive as ICP in warning of TBH.
*Bilirubin excretion and bile flow in fed and fasted Brazilian
squirrel monkeys
(Saimiri sciureus). Cornelius, C. E., Myers, B. A., Bruss, M. L., &
George, J. W. (Dept. of Physiological Sciences, School of Veterinary
Medicine, University of California, Davis, CA 95616).
Veterinary Research Communications, 1989, 13, 395-401.
. . Bolivian squirrel monkeys (BoSMs), unlike Brazilian (BrSMs),
exhibit an exaggerated and marked unconjugated hyperbilirubinaemia upon
fasting, reduced plasma clearance of bilirubin and lower hepatic
uridinediphosphoglucuronate glucuronosyl transferase activity. BoSMs
have been used as experimental animal models for human Gilbert's
syndrome type I. It appears that BrSMs represent the best animals
studied to date to use as controls, since they do not exhibit marked
fasting hyperbilirubinaemia and thus resemble normal humans.
Behavior
*Standing and climbing a pole by members of a captive group of
Japanese monkeys. Machida, S. (Dept. of Comparative & Developmental
Psychology, Fac. of Human Sciences, Osaka Univ., Yamadaoka, Suita,
Osaka, 565 Japan).
Primates, 1990, 31, 291-298.
. . A juvenile female in a captive group spontaneously began standing
poles against a concrete wall and climbing up them. Within 4 years, 3
juvenile females out of 39 monkeys had acquired the behavior.
*Fragile sons or harassed daughters? Sex differences in mortality
among juvenile primates. Van Schaik, C. P. & de Visser, J. A. G. M.
(Dept. of Biological Anthropology & Anatomy, Duke Univ., 3705 Erwin Rd.,
Durham, NC 27705).
Folia Primatologica, 1990, 55, 10-23.
. . Among primates, females tend to be less viable as juveniles than
males. Three hypotheses were tested with over 40 data sets drawn from
the literature. Local resource competition provides the best explanation
for the observed patterns. The contrast between the findings for
primates and those for other taxa is attributed to the low growth rate
of immatures and the widespread occurrence of conditions conducive to
local resource competition in primates.
*Peer interaction in infant chimpanzees. Tomasello, M., Gust, D. A.,
Evans, A. (Dept. of Psychology, Emory Univ., Atlanta, GA 30322).
Folia Primatologica, 1990, 55, 33-40.
. . Six infant
Pan troglodytes, aged 18 to 50 months, were observed in a
seminatural context. Results indicate that a number of factors may
influence the peer affiliations of infant chimpanzees, including the age
of the infant and the mother's social relationships.
*Species differences in response to conspecific intruders in
Callithrix jacchus and Saguinus oedipus. Harrison, M. L. &
Tardif, S. D. (Dept. of Sociology and Anthropology, Kent State Univ.,
Kent, OH 44242).
International Journal of Primatology, 1989, 10, 343-362.
. . Responses elicited in newly mated individuals by conspecific
intruders were compared in 4 pairs of each species. The species differed
both within and between their mating and social systems. These
differences may result from differences in habitat quality, and in the
size of the territory that must be defended from conspecifics.
*Alloparental behaviour in Barbary macaques
(Macaca sylvanus). Small, M. F. (Dept. of Anthropology, Cornell
Univ., Ithaca, NY 14853).
Animal Behaviour, 1990, 39, 297-306.
. . Macaques vary in the amount of infant-non-mother interactions, and
over 300 hours of focal-animal samples of mothers and non-mother females
in a semi-free ranging group show that Barbary macaques are at the
extreme interactive end of this continuum.
*Intergenerational transmission of maternal rejection rates among
free-ranging rhesus monkeys. Berman, C. M. (Dept. of Anthropology, SUNY,
Buffalo, NY 14261).
Animal Behaviour, 1990, 39, 329-337.
. . Rejection rates for individual mothers were consistent from infant
to infant and similar to those of their own mothers. Association with
the mother after infancy may play an important role in the transmission
of rejection rates from mothers to daughters.
*Bachelor groups of gorillas in captivity: The situation in the
wild. Harcourt, A. H. (Large Animal Research Group, Dept. of Zoology,
Cambridge Univ., Cambridge CB3 0DT, UK).
Dodo, 1988, 25, 54-61.
. . A discussion of the benefits and drawbacks of keeping adult male
gorillas together in captivity, based on a review of observations in the
wild.
*Group fission in Nilgiri langurs
(Presbytis johnii). Hohmann, G. (Indian Inst. of Science,
Bangalore, India).
International Journal of Primatology, 1989, 10, 441-454.
. . Four cases of group fission were observed which differed markedly
from group changes reported in other
Presbytis species. Although the presence of all male bands and
single males indicate that male replacement by invasive takeovers may
occur, it is suggested that for
P. johnii, noninvasive fissions are probably another common
mechanism for the foundation and maintenance of bisexual one-male
groups.
*Habitual fission-fusion and social organization of the Hakone troop
T of Japanese macaques in Kanagawa Prefecture, Japan. Fukuda, F.
(2-27-12, Shohnandai, Fujisawa, Kanagawa, 252 Japan).
International Journal of Primatology, 1989, 10, 419-439.
. . A thorough socioecological study and analysis of the seasonal
pattern of fission-fusion in one troop, which formed small subgroups
from spring to autumn. Seasonal variation of pattern and composition of
each subgroup could be attributed to alternation of attractive food
plant species, as well as age, rank, and sexual activity of both sexes.
*Proboscis monkey
(Nasalis larvatus) social organization: Group structure. Yeager, C.
P. (Psychology Dept., Univ. of Tennessee, Knoxville, TN 37996-0900).
American Journal of Primatology, 1990, 20, 95-106.
. . A 12-month study in Indonesia, using scan sampling and event
sampling while following groups. Proboscis monkeys were found to follow
the typical Asian colobine pattern of one-male social groups, with
extra-group males forming all-male groups.
*Formal biting in stumptailed macaques
(Macaca arctoides). Demaria, C. & Thierry, B. (B. Thierry, Lab. de
Psychophysiologie, 7 rue de l'Université, Univ. Louis Pasteur, 67000
Strasbourg, France).
American Journal of Primatology, 1990, 20, 133-140.
. . Formal biting, slowly gripping with teeth a part of the body of an
apparently willing partner, was studied in a captive group of 1 adult
male and 10 females. This behavior seems to represent a ritualized
interaction expressing formal dominance between partners.
*Discrimination of chirp vocalizations in the cotton-top tamarin.
Bauers, K., & Snowdon, C. T. (WRPRC, 1223 Capitol Court, Madison, WI
53715).
American Journal of Primatology, 1990, 21, 53-60.
. . Natural exemplars of two types of vocalizations, sharing many
acoustic features but used in different circumstances, were presented to
8 groups of
Saguinus o. oedipus in a playback paradigm. Results suggest that
tamarins can discriminate subtle acoustic cues, and indicate that the
playback paradigm can be an efficient means of testing discrimination
between different call types.
Care
*Cotton-top tamarins
(Saguinus (o.) oedipus) in a semi-naturalistic captive colony.
Price, E. C. & McGrew, W. C. (Dept. of Psychology, Univ. of Stirling,
Stirling, FK9 4LA, Scotland).
American Journal of Primatology, 1990, 20, 1-12.
. . Data from 6.5 years of records imply that the breeding success of
captive cotton-top tamarins may be improved by maintaining them in
groups whose size and age-sex composition resemble those of wild groups.
Mean litter size decreased, but percentage infant survival increased.
*Increased cage size does not alter heart rate or behavior in female
rhesus monkeys. Line, S. W., Morgan, K. N., Markowitz, H., & Strong, S.
(CPRC, Univ. of California, Davis, CA 95616).
American Journal of Primatology, 1990, 20, 107-113.
. . Responses of 10 animals to three different cage sizes showed only
some differences in vocalizations. Apparently modest increases in cage
size are less likely to enrich the environment of singly caged
laboratory primates than are changes in social opportunities or increases
in environmental complexity.
*Effects of management methods, social organization, and physical
space on primate behavior and health. Erwin, J. & Sackett, G. P. (G. P.
Sackett, RPRC SJ-50, Univ. of Washington, Seattle, WA 98195).
American Journal of Primatology, 1990, 20, 23-30.
. . The incidence of clinical treatment of a large colony of pigtailed
macaques was studied retrospectively to assess the effectiveness of
different housing and management protocols. A protocol dedicating a
two-room suite to each group, with only one room used at a time,
maximized isolation and reduced the available space compared to others,
but trauma, enteritis, respiratory disorders, and inflammation all
decreased markedly. Behavioral studies identified beneficial social
behavior patterns associated with spatial reduction as the basis of the
decrease in aggression, trauma, and disease.
*Time budget of caged rhesus monkeys exposed to a companion, a PVC
perch, and a piece of wood for an extended time. Reinhardt, V. (WRPRC,
1223 Capitol Court, Madison, WI 53715).
American Journal of Primatology, 1990, 20, 51-56.
. . Records of 60 animals over at least 1.5 years show that all three
stimuli remain effective, but a compatible companion is of greater
stimulatory value, especially for adults, than are inanimate objects.
*Nonhuman primate bites. Janda, D. H., Ringler, D. H., Hilliard, J.
K., Hankin, R. C., & Hankin, F. M. (University Hospitals [TC2912], Univ.
of Michigan, 1500 E. Medical Center Drive, Ann Arbor, MI 48109).
Journal of Orthopaedic Research, 1990, 8, 146-150.
. . Emergency rooms and physicians should be aware of the potential
complications of nonhuman primate bites. Lab personnel working with
primates should be informed as to the potential for life-threatening
viral sequelae from, in particular, macaque bites, and should notify
physicians/
emergency room personnel when bite incidents involve macaque
monkeys.
Conservation
*Population dynamics of hoolock gibbons
(Hylobates hoolock) in Assam, India. Choudhury, A. (Near Gate No. 1
of Nehru Stadium, Islampur Road, Guwahati 781 007, Assam, India).
American Journal of Primatology, 1990, 20, 37-41.
. . Data on several groups studied between 1986 and 1988 show fewer
young than reported in earlier studies.
Development
*Growth curves of body weight and their relationship to sexual
maturity in laboratory-bred male African green monkeys
(Cercopithecus aethiops). Hiyaoka, A., Yoshida, T., Cho, F., &
Goto, N. (Corp. for Production and Research of Laboratory Primates,
Hachimandai, Tsukuba-shi, Ibaraki 305, Japan).
Experimental Animals, 1990, 39, 345-352. (Japanese, with English
abstract)
. . Body weight was measured from birth to 6 years for 5 animals, and
5 growth models were applied to the data. Five tables and 4 graphs
help describe the results.
*Behavioral development and socialization of infants in a
free-ranging group of howling monkeys
(Alouatta palliata). Clarke, M. R. (Delta R.P.R.C., Tulane Univ.,
Three Rivers Road, Covington, LA 70433).
Folia Primatologica, 1990, 54, 1-15.
. . A 22-month study of 6 female and 5 male infants from birth
to death, emigration, or end of study, showed that emigration patterns
resulted in groups of adults of low relatedness, and that females, which
were more sociable as adults, were also more sociable as infants and
exhibited less weaning stress than males. Males were forced out
of the group sooner, remained solitary longer, primarily interacted
with adult females as adults, were forced to be independent sooner,
reacted negatively to interactions, and ceased interacting with adult
males by 3 months of age.
Disease
*An epizootic of whooping cough among chimpanzees in a zoo.
Gustavsson, O. E. A., Röken, B. O., & Serrander, R. (Dept. of
Epidemiology, National Bacteriology Lab., 105 21 Stockholm, Sweden).
Folia Primatologica, 1990, 55, 45-50.
. . Fifteen of 19 chimpanzees became ill within several days in June,
suffering from a dry, racking cough combined with a general
deterioration in condition. Six had slight nasal catarrh. At the end of
the second week, one 9-month-old unexpectedly died of purulent
bronchopneumonia. A second outbreak of coughing began in August, which
finally affected 14 chimpanzees and a gorilla (which was separated by a
fence). The coughing gradually decreased and eventually ceased in
September. Symptomatic and antibiotic treatments did not seem to be
effective.
*A cross sectional survey for B virus antibody in a colony of group
housed rhesus macaques. Weigler, B. J., Roberts, J. A., Hird, D. W.,
Lerche, N. W., & Hilliard, J. K. (Dept. of Epidemiology & Preventive
Medicine, School of Veterinary Medicine, Univ. of California, Davis, CA
95616).
Laboratory Animal Science, 1990, 40, 257-261.
. . Of 146 monkeys housed in 3 different outdoor breeding corrals, 97%
of animals older than 2.5 years were seropositive, compared to only 22%
of younger animals. Neither gender nor social dominance ranking were
predictive of B virus antibody status. Venereal transmission of B virus
is hypothesized.
*Genetic organization of a chimpanzee lentivirus related to HIV-1.
Huet, T., Cheynier, R., Meyerhans, A., Roelants, G., & Wain-Hobson, S.
(Lab. de Biologie et Immunologie Moléculaires des Rétrovirus, Inst.
Pasteur, 28 Rue de Dr Roux, 75724 Paris cedex 15, France).
Nature, 1990, 345, 356-358.
. . Sera from two Gabon chimpanzees cross reacted with all the HIV-1
proteins including the envelope glycoproteins. The overall genetic
organization was the same as that of HIV-1, but the sequence was more
divergent than any HIV-1 sequence reported so far. The 2 chimpanzees
were both infants and in relatively good health. None of the 'feral'
SIVs, the African green monkey and mandrill viruses, is associated with
disease, perhaps because diseased animals would not survive in the
jungle. Simian AIDS has only been described among captive monkeys.
*Reactivation of HSV-1 in primates by transcorneal iontophoresis of
adrenergic agents. Rootman, D. S., Haruta, Y., & Hill, J. M. (J. M.
Hill, LSU Eye Center, 2020 Gravier St., Suite B., New Orleans, LA
70112-2234).
Investigative Ophthalmology & Visual Science, 1990, 31, 597-600.
. . Results suggest that the rabbit models developed to date are
analogous to induced HSV-1 reactivation in a nonhuman primate, and that
the conclusions drawn from rabbits may be applicable to human ocular
herpes disease.
*Expression of infectious viral particles by primary chimpanzee
hepatocytes isolated during the acute phase of non-A, non-B hepatitis.
Jacob, J. R., Burk, K. H., Eichberg, J. W., Dreesman, G. R., & Lanford,
R. E. (R. E. Lanford, Dept. of Virology & Immunology, Southwest
Foundation for Biomedical Research, P.O. Box 28147, San Antonio, TX
78284).
Journal of Infectious Diseases, 1990, 161, 1121-1127.
. . The non-A, non-B hepatitis virus (NANBH) is implicated in over 90%
of posttransfusion-associated hepatitis cases. About 40-50% of
NANBH-infected individuals will develop chronic liver disease. The first
experimental transmission of NANBH to chimpanzees, the only reliable
animal model for this disease, was demonstrated over 10 years ago.
Recently, a hormonally defined, serum-free medium with the capacity to
maintain primary baboon hepatocytes for more than 3 months was developed.
Liver
wedge biopsies were obtained from chimpanzees during the acute phase of
experimental NANBH infections. Primary chimpanzee hepatocytes were
maintained for over 4 weeks in vitro. Immunocytochemical staining for a
NANBH-associated antigen revealed expression of this cytoplasmic marker
during the culture period. There were histopathologic alterations
indicative of viral hepatitis, and the appearance of liver cell
cytoplasmic tubules diagnostic for NANBH. Concentrated tissue culture
medium examined by electron microscopy contained virus-like particles
with an average diameter of 39-46 nm, which exhibited an envelope and
inner 37-nm core structure.
*Typhoid immunization. Morbidity and Mortality Weekly Report
Recommendations and Reports, 1990, 39, No. RR-10. 1-5.
. . Vaccination is indicated for travelers to areas that have a
recognized risk of exposure to
Salmonella typhi, particularly those going to developing countries
(especially Latin America, Asia, and Africa) for prolonged periods.
Such travelers need to be cautioned that typhoid vaccination is not a
substitute for careful selection of food and drink, since typhoid
vaccines are not 100% effective.
*Preliminary report: Isolation of Ebola virus from monkeys imported
to USA. Jahrling, P. B., Geisbert, T. W., Dalgard, D. W., Johnson, E.
D., Ksiazek, T. G., Hall, W. C., & Peters, C. J. (Disease Assessment
Div., US Army Med. Research Inst. of Infectious Disease, Fort Detrick,
MD 21701).
Lancet, 1990, 335, 502-505.
. . In 5 sick animals from the Virginia epizootic in October, 1989,
tissues that yielded Ebola (EBO)-like viruses also contained EBO viral
antigens by both ELISA and immunohistochemistry, and filovirus particles
by electronmicroscopy. Further studies to define the virulence
potentials of these new filovirus isolates and their relation to the EBO
virus prototype strains are of high priority.
*Clinical and parasitologic aspects of cryptosporidiosis in nonhuman
primates. Miller, R. A., Bronsdon, M. A., Kuller, L., & Morton, W. R.
(Div. of Infectious Disease, Seattle Veteran's Admininstration Medical
Center, Seattle, WA 98108).
Laboratory Animal Science, 1990, 40, 42-46.
. . Report of 81 cases of acute cryptosporidiosis among 157 infant
primates at the Washington R.P.R.C. The risk of infection was related to
the length of time the animal was housed in the nursery and to social
interaction with other monkeys.
*Experimental cryptosporidiosis in a primate model. Miller, R. A.,
Bronsdon, M. A., & Morton, W. R. (M. Bronsdon, RPRC SJ-50, Univ. of
Washington, Seattle, WA 98195).
Journal of Infectious Diseases, 1990, 161, 312-315.
. . Inoculation with either 2 x 10&S'5 or 10
Cryptosporidium oocysts resulted in clinical enteritis and the fecal
passage of large numbers of oocysts. The size of the inoculum had no
apparent effect on the severity or duration of disease. The small
inoculum size coupled with the passage of large numbers of oocysts
contributes to the highly contagious nature of cryptosporidiosis among
captive primates and may be relevant to the epidemiology and control of
the disease in humans.
*Giardia isolates from primates and rodents display the same
molecular polymorphism as human isolates. De Jonckheere, J. F.,
Majewska, A. C., & Kasprzak, W. (Dept. of Microbiology, Inst. of Hygiene
& Epidemiology, J. Wytsmanstraat 14, B-1050 Brussels, Belgium).
Molecular and Biochemical Parasitology, 1990, 39, 23-28.
. . Two profiles of restriction fragment length polymorphism were
found in
Giardia isolated from animals, which are identical to two profiles
found previously in strains of human origin. Isoenzyme and total protein
patterns obtained with agarose isoelectric focusing divided the strains
in the same two groups. Pulsed field gradient gel electrophoresis showed
non-identical chromosomal band patterns.
*Characterization of a simian hepatitis A virus (HAV): Antigenic and
genetic comparison with human HAV. Brown, E. A., Jansen, R. W., & Lemon,
S. M. (Div. of Infectious Diseases, Dept. of Medicine, Univ. of North
Carolina, Chapel Hill, NC 27599-7030).
Journal of Virology, 1989, 4932-4937.
. . The close similarity between a strain (PA21) of HAV recovered from
a naturally infected captive owl monkey and a reference human virus was
confirmed with an extended panel of 18 neutralizing murine monoclonal
antibodies, but other data indicate that PA21 represents a unique
genotype of HAV and suggest the existence of an ecologically isolated
niche for HAV among feral owl monkeys.
*In vivo replication and reversion to wild type of a
neutralization-resistant antigenic variant of hepatitis A virus. Lemon,
S. M., Binn, L. N., Marchwicki, R., Murphy, P. C., Ping, L.-H., Jansen,
R. W., Asher, L. V. S., Stapleton, J. T., Taylor, D. G., & LeDuc, J. W.
(Address same as above).
Journal of Infectious Diseases, 1990, 161, 7-13.
. . Results from inoculating 6 owl monkeys with an antigenic variant
of hepatitis A virus further define virologic events in hepatitis A and
show that in vivo replication of an antigenic variant was restricted
compared with that of wild-type virus.
*Serological survey for two simian retroviruses in macaques and
African green monkeys. Krugner-Higby, L., Kucera, L., Lerche, N., Sever,
J., Fucillo, W., Allan, J., & Benveniste, R. (Dept. of Comparative
Medicine, Wake Forest Univ. Medical Center, Winston-Salem, NC 27103).
Laboratory Animal Science, 1990, 40, 24-28.
. . Colonies at the Bowman Gray School of Medicine
were tested for SIV and SRV-1 antibodies by IFA and western blot.
36% of cynomolgus, 3% of rhesus, and 12% of stumptails were positive for
SRV-1 antibodies by IFA. 36% of African green monkeys were IFA positive
for SIV antibodies. The IFA was highly predictive for SIV in the African
green monkeys, but far less so for SRV-1 in the macaques.
*Immunogenicity of liposomal malaria sporozoite antigen in monkeys:
Adjuvant effects of aluminium hydroxide and non-pyrogenic liposomal
lipid A. Richards, R. L., Swartz, G. M., Jr., Schultz, C., Hayre, M. D.,
Ward, G. S., Ballou, W. R., Chulay, J. D., Hockmeyer, W. T., Berman, S.
L., & Alving, C. R. (Dept. of Membrane Biochemistry, Walter Reed Army
Inst. of Research, Washington, DC 20307-5100).
Vaccine, 1989, 7, 506-512.
. . A recombinant protein (R32tet&s'32.) that contains sequences from
the repeat region of the CS protein of
P. falciparum in combination with aluminium hydroxide (alum) has
undergone phase 1 testing in humans. Although slow titers of antibodies
were obtained in humans, the feasibility of the approach was
demonstrated in that one of two individuals given the highest dose of
antigen was protected against sporozoite challenge. In each case
liposomal lipid A was 25-fold to 200-fold less pyrogenic than free lipid
A. Excellent antibody levels were also observed in monkeys immunized
with a combination of R32tet&s'32 encapsulated in alum-adsorbed
liposomes containing non-pyrogenic doses of monophosphoryl lipid A and
alum. It was concluded that a non-pyrogenic formulation of alum-adsorbed
liposomes, in which the liposomes contained both lipid A and an
encapsulated synthetic sporozoite antigen, shows considerable promise
for inducing high titers of antibodies to sporozoites.
*Characterization of simian immunodeficiency virus-specific
T-cell-mediated cytotoxic response of infected rhesus macaques. Vowels,
B. R., Gershwin, M. E., Gardner, M. B., Ahmed-Ansari, A., & McGraw, T.
P. (T. P. McGraw, Dept. of Medical Pathology, Univ. of California, Davis,
CA 95616).
AIDS, 1989, 3, 785-792.
. . This study demonstrates the presence of SIV&s'MAC.-specific
cytotoxic T lymphocytes (CTL) in the peripheral blood of healthy rhesus
macaques that have been infected with SIV&s'MAC.. An enigma of AIDS is
the evidence that there are levels of what would normally be protective
humoral and cellular immune activity in individuals with AIDS. Possible
explanations for the progressive clinical decline in the presence of
'protective' immune responses include the destruction of normal CD4+
lymphocytes by immunologic mechanisms following adsorption of virus or
viral antigens to their surface. In a host with defective T-cell
maturation, this would lead to the elimination of the cells central to
responding to challenges by opportunistic and other infectious agents.
*Simian immunodeficiency virus-specific T-cell-mediated
proliferative response of infected rhesus macaques. McGraw, T. P.,
Vowels, B. R., Gardner, M. B., Ahmed-Ansari, A., & Gershwin, M. E.
(Address same as above).
AIDS, 1990, 4, 191-198.
. . This study demonstrates that significant levels of T-cell
proliferative responses are present during the early course of SIV
infection in rhesus macaques, and that loss of in vitro proliferation
corresponds with clinical decline.
*Simian immunodeficiency viruses from African green monkeys display
unusual genetic diversity. Johnson, P. R., Fomsgaard, A., Allan, J.,
Gravell, M., London, W. T., Olmsted, R. A., & Hirsch, V. M. (Retroviral
Pathogenesis Sec., Div. of Molecular Virology & Immunology, Dept., of
Microbiology, Georgetown Univ., 12441 Parklawn Dr., Rockville, MD
20852).
Journal of Virology, 1990, 64, 1086-1092.
. . At least 6 species of African primates are naturally infected with
SIV, and do not manifest the AIDS-like illness observed in macaques
experimentally infected with SIV. The variation among SIV&s'agm.
isolates is greater than the difference between a simian and a human
lentivirus, suggesting that SIV&s'agm. has been in the African green
monkey population for a long time and may be the oldest primate
lentivirus group in existence.
*Peripheral neuropathy associated with osteosarcoma in a Japanese
monkey. Yasuda, H., Taniguchi, Y., & Shigeta, Y. (3rd Dept. of Medicine,
Shiga Univ. of Medical Science, Ohtsu-shi, Shiga 520-21, Japan).
Experimental Animals, 1990, 39, 285-289.
. . This is the first case found of a monkey dying of neuropathy
associated with malignancy, which is not infrequent in humans.
*Studies on Hepatocystis sp. in rhesus monkeys from Yunnan,
China. Takenaka, T., Hashimoto, K., Gotou, H., Matsumoto, S., &
Nishikawa, T. (Drug Safety Research Center, Otsuka Pharm. Co., 463-10
Kagasuno, Kawauchi-cho, Tokushima-shi, Tokushima 771-01, Japan).
Experimental Animals, 1990, 39, 273-279. (Japanese with English
abstract and figure captions)
. . In 6 of 10 cases, examination of blood smears and livers showed
the presence of
Hepatocystis sp. 7 microphotographs and a photograph are included.
*Sero-epidemiological studies of malaria in Indian tribes and
monkeys of the Amazon basin of Brazil. de Arruda, M., Nardin, E. H.,
Nussenzweig, R. S., & Cochrane, A. H. (Dept. of Entomology, Inst.
Oswaldo Cruz, Rio de Janeiro, Brazil).
American Journal of Tropical Medicine and Hygiene, 1989, 4,
379-385.
. . Low incidence of malaria, but high levels of antibodies to both
blood stages and sporozoites of
Plasmodium spp., was found in 4 Indian tribes.
P. brasilianum were found in the blood of indigenous monkeys often
kept as pets, suggesting that the monkeys serve as reservoir hosts.
Anopheles darlingi mosquitoes, infected with P. spp. were found
in the study area.
Ecology
*Ecological constraints on group size in three species of
neotropical primates. Chapman, C. A. (Dept. of Anthropology, Harvard
Univ., Cambridge, MA 02138).
Folia Primatologica, 1990, 55, 1-9.
. . The foraging strategies and association patterns of
Ateles geoffroyi, Alouatta palliata, and Cebus capucinus were
studied over a 5-year period to test the hypothesis that the size,
density, and distribution of food resources influence the size of animal
groups. Only the Capuchins did not respond to changes in ecological
variables by forming subgroups or changing the cohesion of their group.
*Plant feeding of yellow baboons
(Papio cynocephalus) in Mikumi National Park, Tanzania, and the
relationship between seasonal feeding and immature survival. Rhine, R.
J., Norton, G. W., Wynn, G. M., & Wynn, R. D. (Psychology Dept., Univ.
of California, Riverside, CA 92521).
International Journal of Primatology, 1989, 10, 319-342.
. . Focal-animal feeding data from 64 adults and 46 infants during a
3-year period were used to evaluate hypotheses predicting selection for
a birth peak and to study the baboon's eclectic/selective feeding
adaptation.
*Comparative density, demography, and ranging behavior of Barbary
macaques
(Macaca sylvanus) in marginal and prime conifer habitats. Mehlman,
P. T. (Laboratory Animal Breeders & Services, Yemassee, SC 29945).
International Journal of Primatology, 1989, 10, 269-292.
. . Six groups of 162 animals inhabiting (marginal) fir forests in the
Rif mountains are compared to a population in the (prime) Moyen Atlas.
Small groups, occupying larger ranges, in the marginal habitat have
better rates of growth than small groups in prime habitat.
*Gorillas
(Gorilla gorilla gorilla) in the Likouala swamp forests of north
central Congo: Preliminary data on populations and ecology. Fay, J. M.,
Agnagna, M., Moore, J., & Oko, R. (Dept. of Anthropology, Washington
Univ., St. Louis, MO 63110).
International Journal of Primatology, 1989, 10, 477-486.
. . Two recent, independent surveys extend the known range of
G. g. gorilla very close to the Oubangui River, where they are
found at high densities in the Likouala swamp.
Genetics
*Biochemical genetic markers of squirrel monkeys and their use for
pedigree validation. VandeBerg, J. L., Aivaliotis, M. J., Williams, L.
E., & Abee, C. R. (C. R. Abee, Dept. of Genetics, Southwest Foundation
for Biomedical Research, P.O. Box 28147, San Antonio, TX 78228-0147).
Biochemical Genetics, 1990, 28, 41-56.
. . Family data for biochemical genetic markers of squirrel monkeys
were derived from 73 pedigreed progeny and both parents of each. It was
concluded that the pedigree records of 7 progeny were incorrect.
Although 5 of the pedigree errors were proved to be mistaken paternity,
the others apparently were the consequence of infant swapping between
dams shortly after birth.
*Genetic variability in natural populations of the Brazilian night
monkey
(Aotus infulatus). Schneider, M. P. C., Sampaio, M. I. C.,
Schneider, H., & Salzano, F. M. (Dept. de Genética, Centro de
Ciências Biológicas, Univ. Federal do Pará, Campus Univ. do
Guamá, 66059 Belém, PA, Brazil).
International Journal of Primatology, 1989, 10, 363-374.
. . Examination for 12 blood protein loci in 71 animals from both
banks of the Tocantins River in Amazonia indicate remarkable differences
between the subpopulations of the two banks. These results and the
difficulties in delimiting species in this genus may indicate a
relatively recent spur of evolutionary differentiation.
*Three new blood groups of rhesus monkeys. Frederick, D. L.,
Samples, N. K., & Stone, W. H. (W. H. Stone, Dept. of Biology, Trinity
Univ., 715 Stadium Dr., San Antonio, TX 78212).
American Journal of Primatology, 1990, 20, 127-131.
. . Three new blood group systems, called "T," "U," and "V," have
been indentified in the rhesus monkey, making 16 different blood group
systems and well over one hundred million possible phenotypes in this
species.
Instruments & Techniques
*Effects of chair restraint on plasma enzyme values in the rhesus
monkey (Macaca mulatta). Tatsumi, T., Komatsu, H., & Adachi, J.
(Drug Development Labs, Chugai Pharmaceutical Co., 3-41-8
Takada, Toshima-ku, Tokyo 171, Japan).
Experimental Animals, 1990, 39, 353-359. (Japanese with English
abstract)
. . Six monkeys were restrained for 8 hours. Plasma enzyme values
increased within 3 hours and remained elevated for up to 2 days after
release. Four graphs and a table illustrate the data.
*Effects of repeated chair restraint on physiological values in the
rhesus monkey (Macaca mulatta). Tatsumi, T., Koto, M., Komatsu,
H., & Adachi, J. (Address same as above).
Experimental Animals, 1990, 39, 353-359. (Japanese with English
abstract)
. . Although heart rate and blood pressure increased significantly
with the outset of chair restraint, they gradually decreased with
repetition of restraint as well as with lapse of time after release.
Finally no elevation was detected after repetitions lasting 8
hours/day for 5 days. Plasma enzymes showed similar trends. There are
8 tables of data.
*Systems for collection of urine in the captive common marmoset,
Callithrix jacchus. Lunn, S. F. (MRC Reproductive Biology Unit,
Centre for Reproductive Biology, 37 Chalmers St., Edinburgh EH3 9EW,
UK).
Laboratory Animals, 1989, 23, 353-356.
. . Two systems are described, and results reported from 1210 24-h
samples from 84 adult animals. No significant volume differences were
observed between pregnant and non-pregnant females, nor did bilateral
gonadectomy cause any difference in either sex.
Nutrition
*Long-term parenteral nutrition in unrestrained nonhuman primates:
An experimental model. Friday, K. E. & Lipkin, E. W.
American Journal of Clinical Nutrition, 1990, 51, 470-476.
. . Twelve Macaca fascicularis received total parenteral
nutrition through a freely mobile jacket and tether system for 6-8
weeks. Major complications included catheter sepsis, hyperglycemia,
diarrhea, and premature death in 6 animals.
Physiology
*Hypoglycemia of squirrel monkey neonates: Implications for infant
survival. Brady, A. G., Williams, L. E., & Abee, C. R. (Dept. of
Comparative Medicine, College of Medicine, Univ. of South Alabama,
Mobile, AL 36688).
Laboratory Animal Science, 1990, 40, 262-265.
. . Serum glucose determinations were done at 1, 3, 7, 10, 14 days and
1 month of age using a standard laboratory test. Glycosylated hemoglobin
measurements were made at 1 day, 1 week, 2 weeks, 1 month, 2 months, 1
year, and in adults. Blood glucose concentration is significantly lower
in squirrel monkey neonates than in older infants, juveniles, and
adults. The marginal hypoglycemic state of these infants places them at
high risk for clinical hypoglycemia as a sequel to maternal rejection,
trauma, and associated problems.
*Mitogenic responses of peripheral blood mononuclear cells of vervet
monkeys
(Cercopithecus aethiops): Apparent role of adherent cells. Olobo,
J. O. & Reid, G. D. F. (Inst. of Primate Research, P.O. Box 24481,
Karen, Nairobi, Kenya).
American Journal of Primatology, 1990, 20, 31-36.
. . This paper describes baseline immune responses of normal
colony-housed vervet monkeys to several stimuli.
*Gonadal hormones influence the emergence of cortical function in
nonhuman primates. Clark, A. S. & Goldman-Rakic, P. S. (P. S.
Goldman-Rakic, Section of Neuroanatomy, Yale Univ. School of Medicine,
New Haven, CT 06510).
Behavioral Neuroscience, 1989, 103, 1287-1295.
. . A study of the impact of gonadal steroids on the ontogeny and
maturation of other brain areas and the expression of nonreproductive
behaviors. Normal male and female rhesus monkeys, monkeys given orbital
prefrontal cortex (ORB) lesions at 50 days of age, and female monkeys
given androgen at different ages were tested on an object discrimination
reversal task at 75 days of age. Gender influenced the performance of
monkeys during normal development and after ORB lesions. Females treated
with androgen performed similarly to normal males. The findings suggest
that gonadal hormones may play an inductive role in the differentiation
of higher cortical function in nonhuman primates.
*Neural and hormonal control of blood pressure in conscious monkeys.
Cornish, K. G., Barazanji, M. W., & Iaffaldano, R. (Dept. of Physiology,
Univ. of Nebraska College of Medicine, Omaha, NB 68105-1065).
American Journal of Physiology, 1990, 258, H107-H112.
. . Results imply that the sympathetic nervous system is the primary
controlling mechanism for blood pressure in the conscious primate, with
arginine vasopressin making a minor contribution. The release of renin
would appear to be primarily under the control of the sympathetic
nervous system.
*Species differences in regulation of alpha-adrenergic receptor
function. Shen, Y.-T., Vatner, D. E., Gagnon, H. E., & Vatner, S. F. (S.
F. Vatner, NERPRC, 1 Pine Hill Dr., Southborough, MA 01772).
American Journal of Physiology, 1989, 257, R1110-R1116.
. . Results suggest that major species differences in physiological
responsiveness to alpha -adrenergic agonists and autonomic receptor density
exist. These major differences must be recognized in extrapolation of
results among species from cardiovascular physiological and biochemical
studies.
*Brain 5-HT and inhibition of aggressive behavior in animals: 5-HIAA
and receptor subtypes. Miczek, K. A., Mos, J., & Olivier, B. (Dept. of
Psychology, Tufts Univ., Medford, MA 02155).
Psychopharmacology Bulletin, 1989, 25, 399-403.
. . Review of studies in various nonhuman primate species. Future
development of appropriate 5-HT2 agonists and 5-HT1ABC and D
antagonists will allow the roles of these receptors in aggressive
behavior to be characterized more definitively.
Reproduction
*1989 International Studbook Golden-Headed Lion Tamarin (Leontopithecus chrysomelas). Number Two. J. J. C. Mallinson, Studbook holder. Jersey: Wildlife Preservation Trust [Jersey Zoological Park, Jersey, Channel Islands, UK]. 41 pp.
*Male reproductive success in harem troops of Hanuman langurs
(Presbytis entellus) . Sommer, V. & Rajpurohit, L. S. (Inst. für
Anthropologie, Univ. Göttingen, Bürgerstrasse 50, 34 Göttingen,
FRG).
International Journal of Primatology, 1989, 10, 293-317.
. . Analysis of cross-sectional and longitudinal studies from
1967-1988.
*Sires influence fetal death in pigtailed macaques
(Macaca nemestrina). Sackett, G. P. (RPRC SJ-50, Univ. of
Washington, Seattle, WA 98195).
American Journal of Primatology, 1990, 20, 13-22.
. . Colony records were used to identify dams and sires whose
reproductive histories indicated high or low risk for producing fetal
deaths. In subsequent matings, high-risk sires had a 4.86 times higher
relative risk of producing fetal death than low-risk sires, while in
high-risk dams the risk was only 2.64 times higher. Low-risk dams
experienced a 3-fold increase in fetal deaths when mated with a
high-risk sire, whereas high-risk dams had a nearly 50% reduction in
fetal deaths when mated with a low-risk sire. These results show that
male factors beyond fertility need to be considered when examining the
causes of reproductive failure.
*5-Hydroxyeicosatetraenoic acid, leukotriene C4., and
prostaglandin F2alpha in amniotic fluid before and during term and
preterm labor. Walsh, S. W. (Dept. of Ob/Gyn, Univ. of Texas Medical
School, 6341 Fannin St., Suite 3.204, Houston, TX 77030).
American Journal of Obstetrics and Gynecology, 1989, 161,
1352-1360.
. . Amniotic fluid concentrations of 5-HETE, LTC4, and PGF2alpha in
5 pregnant, chronically catheterized rhesus monkeys indicate:
5-HETE and LTC4, but not PGF2alpha, are associated with uterine
contractility after preterm intrauterine surgery; 5-HETE and LTC4 are
present in higher
concentrations than PGR2alpha; all three increase with the
onset of labor; Labor can occur with suppressed PGF2alpha levels, but
increasing 5-HETE and LTC4 levels. These data suggest that
5-HETE and LTC4 are important components of the parturitional
process, and challenge the dogma that PGs are the universal mediators of
labor.
*Diagnosis of abortion and fetal death by the ultrasonographical
device in cynomolgus monkeys under indoor individually-caged conditions.
Kohno, M., Suzuki, M. T., Ono, T., Ogawa, H., & Cho, F. (Corp. for
Production and Research of Laboratory Primates, National Inst. of
Health, 1-Hachimandai, Tsukuba-shi, Ibaraki 305, Japan).
Experimental Animals, 1990, 39, 291-294. (Japanese, with English
abstract)
. . The combined rate of abortion and fetal death was 7.0% in wild-born
and 7.8% in colony-born animals. There were no significant differences in
the rate with respect to age and time in colony.
*Social contraception in naked mole-rats and marmoset monkeys.
Abbott, D. H., Barrett, J., Faulkes, C. G., & George, L. M. (MRC/AFRC
Comparative Physiology Research Group, Inst. of Zoology, Zoological
Society of London, Regent's Park, London NW1 4RY, UK).
Journal of the Zoological Society of London, 1989, 219, 703-710.
. . Review of recent advances in understanding of social suppression
of reproduction in 2 species which display this to an extreme degree.
*Progesterone antagonist lilopristone: A potent abortifacient in the
common marmoset. Puri, C. P., Patil, R. K., Kholkute, S. D., Elger, W.
A. G., & Swamy, X. R. (Inst. for Research in Reproduction (ICMR), Parel,
Bombay-400012, India).
American Journal of Obstetrics and Gynecology, 1989, 161,
248-253.
. . This study suggests that ZK 98.734 has potential for fertility
regulation, and that clinical trials for postcoital contraception,
induction of menstruation, and early abortifacient effects are
warranted.
*Postpartum ovulation and conception in Goeldi's monkey
Callimico goeldii. Ziegler, T. E., Snowdon, C. T., & Warneke, M.
(Dept. of Psychology, Univ. of Wisconsin, Madison, WI 53706).
Folia Primatologica, 1989, 52, 206-210.
. . The occurrence of postpartum ovulations in all 6 postpartum periods
of 4 animals demonstrates that
Callimico does not have lactational suppression of ovulation.
*Reproduction of the owl monkey
(Aotus nancymai) (Primates: Cebidae) in captivity. Gozalo, A., &
Montoya, E. (Proyecto Peruano de Primatología-IVITA, P.O. Box 621,
Iquitos, Peru).
American Journal of Primatology, 1990, 21, 61-68.
. . Reproductive data from a breeding colony of 79 pairs, which
produced 180 offspring in the past 10 years.
Taxonomy
*Titis, new world monkeys of the genus
Callicebus (Cebidae, Platyrrhini): A preliminary taxonomic review.
Hershkovitz, P. (Dept. of Zoology, Field Museum of Natural History,
Chicago, IL 60605-2496).
Fieldiana: Zoology, New Series, No. 55.
. . Descriptions in terms of external, cranial, postcranial, dental,
and cerebral characters, with comparisons with other platyrrhines,
including all other nonprehensile-tailed cebids.
*Genetic differentiation between baboon subspecies: Relevance for
biomedical research. Williams-Blangero, S., Vandeberg, J. L., Blangero,
J., Konigsberg, L., & Dyke, B. (Dept. of Genetics, Southwest Foundation
for Biomedical Research, P.O. Box 28147, San Antonio, TX 78228).
American Journal of Primatology, 1990, 20, 67-81.
. . Failure to identify the complete taxonomic classification of
research subjects presents a source of error for scientists attempting
to evaluate results or to repeat experiments. Analyses of the genetic
distances among five baboon subspecies are presented. Statistical tests
on the distances showed that significant genetic differences exist. Two
of the subspecies also differ significantly for biomedically relevant
lipoprotein cholesterol levels, as can be predicted from the genetic
distances.
*Subspecific morphological variation in the saddle-back tamarin
(Saguinus fuscicollis). Cheverud, J. M., & Moore, A. J. (Dept. of
Anatomy & Neurobiology, Washington Univ. School of Medicine, Box 8108,
660 S. Euclid Ave., St. Louis, MO 63110).
American Journal of Primatology, 1990, 21, 1-15.
. . Analysis of 11 facial measurements from 104 specimens indicate an
absence of geographically related morphological variation across the
species range while phylogenetic relationships are reflected in
morphological similarities among groups and individuals. This finding is
consistent with the formation of subspecific populations by relatively
small founding populations budding off from a larger parent population
and remaining relatively isolated.
* * * -------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. (Phone: 401-863-2511)
Judith_Schrier@brown.edu
ACKNOWLEDGMENTS
The Newsletter is supported by U. S. Public Health
Service Grant RR-00419 from the Animal Resources Program,
Division of Research Resources, N.I.H.
Cover drawing of a mountain gorilla (Gorilla gorilla berengei) by Dr. Robert M. George, Department of Anatomy, University of South Carolina
Copyright @1990 by Brown University
Editor: Judith E. Schrier, M. Sc.
Associate Editor: James S. Harper, D.V.M.
Consulting Editor: Morris L. Povar, D.V.M.
Copy Editor: Elva Mathiesen, B. A.
Founding Editor: Allan M. Schrier, Ph.D.
{kind=link}
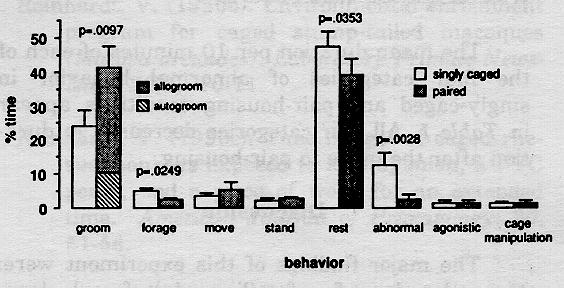{kind=link}
{kind=link}