 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 30 NUMBER 2 APRIL 1991
Articles and Notes
A Canine Companion for a Nursery-Reared Infant Chimpanzee, by M. A. Thompson, M. A. Bloomsmith, & L. L. Taylor...... 1
Comparing the Effectiveness of PVC Swings versus PVC Perches as Environmental Enrichment Objects for Caged Female Rhesus Macaques (Macaca mulatta), by J. Kopecky & V. Reinhardt...... 5
Social Housing Ameliorates Behavioral Pathology in Cebus apella, by K. Bayne, S. Dexter, & S. Suomi...... 7
Klebsiella pneumoniae Infection in a New World Nonhuman Primate Center, by A. Gozalo & E. Montoya...... 13
Effects of Peer Deprivation on Behavior of Captive Patas Monkeys (Cercopithecus [Erythrocebus] patas), by S. Willard & J. Loy...... 20
News, Information, and Announcements
Letters: Controversial Project...... 4
Grants Available...... 6
. .
AmFAR Scholar Awards, Wenner-Gren Foundation for Anthropological
Research, Basic Research on Osteoporosis, Models for SIDS,
Communication Disorders Grants, Lessening Animals' Pain
and Distress, Model for Vaccination in Pregnancy, Alzheimer's
Disease, Cancer Research Institute, H. F. Guggenheim Foundation
A Role Model in the Field of Primate Psychology: George M. Haslerud (1906-1990), by G. Mitchell...... 12
Travellers' Health Notes...... 15
. .
Traveler's Diarrhea, Cholera in Peru, Bubonic Plague Alert,
Malaria Prophylaxis, Dengue Epidemic 1990
Meeting Announcements...... 18
. .
IPS in '92, Cal State Fullerton, Royal Society Discussion
Meeting, IV World Veterinary Congress, AALAS and ICLAS,
Travel to ASP, NABR Conference
Contraceptive Implants in Callitrichids: A Call for Information, by E. C. Price & S. Evans...... 19
Course Announcement: Pathology of Lab Animals...... 23
News Briefs: USDA Standards Finalized...... 24
Research and Education Opportunities...... 25
. .
Wildlife Preservation Trust, Earthwatch, AALAS Certification
Courses, New World Monkeys for Research
Information Requested and Available...... 35
. .
Bonobo Network, PIC 1991 Fee Schedule and Catalog, Reading and
Reference Lists, Specimen Resources Directory, Orangutan
Project, NSF Outreach Service, WRPRC Library Announcements,
Prenatal Drug Exposure, Environmental Enrichment Ideas, Animal
Research Information Board
Cartoon ...... 25
Departments
Address Changes...... 8
Positions Available: Primate Foundation of Arizona...... 24
Recent Books and Articles...... 26
* * *
M. A. Thompson, M. A. Bloomsmith, and L. L. Taylor
University of Calgary, University of Texas, University of Miami
Introduction
Mother rearing is the best means to raise infant chimpanzees, but because of infant or maternal ill health or maternal incompetence, infants must sometimes be raised by humans in a nursery. Such rearing has immediate and long-term consequences. Nursery rearing may cause infants to exhibit abnormal or stereotyped behaviors (e.g. rocking, self-orality, self-clutching) (Capitanio, 1986). These behaviors may be normal infantile response patterns directed, in the absence of the mother, toward the infant's own body (Berkson, 1968) and serve no apparent function (Davenport & Menzel, 1963). In the long term, nursery-reared chimpanzees, as adults, may experience problems with sociality, reproduction, maternal competence, and persistent behavioral pathologies (Berkson & Mason, 1964; Capitanio, 1986; Davenport & Menzel, 1963; Davenport & Rogers, 1970; Dienske & Griffin, 1978; Rogers & Davenport, 1969, 1970; Turner, Davenport, & Rogers, 1969).
The physical contact and constant stimulation provided by the mother during the infant's first year of life (Nicolson, 1977) cannot be met by a human caretaker unless the human is present 24 hours a day -- a condition that most nurseries cannot provide. One solution has been to rear primate infants with dogs as alternative companions (Capitanio, 1984; Mason & Capitanio, 1988; Mason & Kenney, 1974). Young rhesus monkeys of various ages developed specific attachments when housed individually with dogs (Mason & Kenny, 1974), and dog-reared animals were more active in social situations than those raised without canine companions (Capitanio, 1984). To date, this type of companionship has not been quantitatively evaluated for chimpanzees, although dogs are currently being used at a few chimpanzee facilities, including the one at the M. D. Anderson Cancer Center (see also Fritz & Fritz, 1985; Struthers, Rodriguez, Cooper, & Rowell, 1990). The purpose of this pilot study was to document the interaction between a dog and an infant chimpanzee to determine if rearing with a canine companion would (1) reduce stress-related behaviors, (2) reduce stereotyped behaviors such as rocking, and (3) increase locomotion and play in the chimpanzee.
Methods
Subjects: A male infant chimpanzee (Pan troglodytes) born March 23, 1990, was taken from his mother at 5.5 weeks of age because of potentially life-threatening maternal abuse. The mother's social setting was changed in hopes of improving maternal behavior. The infant was later reintroduced to her, but the mother continued to be abusive, so at 6 weeks of age the infant was permanently removed and nursery reared. The infant was 13 weeks old at the beginning of this study. The dog was a female collie-cross which has been used as a companion for nursery-reared infants at the facility for five years.
Housing: The infant was kept in an aluminum mesh playpen measuring 3 x 3 feet. He had fleece pads (cloth blankets) and assorted toys available at all times. The playpen allowed him to stand bipedally and to climb up the side when he was able. After the first week of the study, the playpen was put into a room that allowed the infant to see other nursery-reared chimpanzees.
Procedure: The study consisted of control and experimental conditions. In the control condition the dog was not present in the nursery room. In the experimental condition the dog was present and the chimpanzee was allowed to have contact with her. During all test periods the playpen was turned on its side so full contact between the infant and the dog was possible.
Data Collection: Data were collected for six weeks between June 27 and July 31, 1990. Tests were done four to seven times a week, with each test consisting of a 20-minute control and a 20-minute experimental observation, balanced for order. One-zero sampling at 10-second intersample intervals was used with the chimpanzee as the focal animal. A total of 26 tests (17.3 hours) were done. Data were collected using 33 dependent measures that were grouped into the following categories for analysis: play, stress-related facial expressions, other stress-related behaviors, grasp, quadrupedal locomotion, other locomotion, orality, resting/stationary, interact with dog, stereotyped rocking, watching another chimpanzee or the dog, and miscellaneous behaviors.
Analysis: Control and experimental data were summarized by computing the percentages of the total number of data points in each behavioral category. When behaviors occurred simultaneously, each was recorded. Therefore the sum of all percentages in each category can be greater than 100%. Data were analyzed using the Student's t-test, with an alpha level of .05.
Results
Three behavioral categories indicative of stress were significantly reduced when the dog was present. The categories were rocking (t = 2.6; df = 25; p = .01), stress-related facial expressions (t = 2.4; df = 25; p = .03), and other stress-related behaviors (t = 2.1; df = 25; p = .01) (See Table 1). The incidence of grasping (t = 2.5; df = 25; p= .02) and other locomotion (bipedal and climbing) (t = 2.5; df = 25; p= .02) were significantly lower when the dog was present. The category of rest/stationary showed a significant increase (t = 3.6; df = 25; p = .001) when the dog was present. Most of this increase seemed to be due to the infant lying stationary and watching the dog.
+---------------------------------------------------+
| Control Experimental|
|Behavior Condition Condition |
+---------------------------------------------------+
|Rock 4.0% 1.9%* |
|Stress-related facial |
| expressions 1.0 .3* |
|Stress-related behaviors 3.0 0.6* |
|Grasp 17.7 11.5* |
|Other locomotion 11.0 4.6* |
|Rest/Stationary 28.1 43.4* |
|Quadrupedal locomotion 1.5 1.2 |
|Play 11.0 9.6 |
|Orality 7.3 5.4 |
|Watch chimpanzee or dog 6.4 4.9 |
|Interact with dog NA 12.0 |
+---------------------------------------------------+
*Significant difference at an alpha level of .05.
Table 1. Percentages of collected data points that the infant chimpanzee spent in various activities in control and experimental phases of the study. (Simultaneous behaviors account for the total exceeding 100%).
Behaviors not significantly affected by the presence of the dog were play, quadrupedal locomotion, and orality. Dog-directed behaviors were not eliminated or decreased after six weeks of exposure to the dog. On average, the chimpanzee interacted with the dog during 12% of the sampled points. Thus, the dog remained an attractive companion across time.
Discussion
The dog's presence had a largely positive effect on the infant's behavior; the hypotheses of decreasing stress-related behavior and abnormal behavior were supported. These decreases appeared to be the result of additional social stimulation supplied by the dog companion. This is consistent with the idea that stereotyped behavior in nursery- reared chimpanzees may be related to the absence of social and physical stimuli, particularly kinesthetic stimulation, ordinarily provided by the mother (Davenport & Rogers, 1970), and that they may serve the function of self-stimulation. Such behaviors are most prevalent in highly restrictive environments that provide few sources of social and physical stimulation and do not allow the animal to modify its environment (Baumeister & Forehand, 1973; Berkson & Mason, 1964; Davenport & Rogers, 1970; Walsh, Bramblett, & Alford, 1982).
As in this study, rhesus macaques reared with dogs show some abnormal behavior, although less than similar monkeys raised without canine partners (Capitanio, 1984). Such decreases in stress behaviors may develop because the dog is an animate being that moves and responds to the infant. An attachment may form between the two animals that is comforting for the infant in stressful situations. Rhesus have formed strong and specific attachments to their canine companions (Mason & Kenney, 1974). Although the infant in this study appeared to show some interest in the dog throughout testing, contact with the dog was more frequent near the end of the study. This may be indicative of the bond formation process as well as the infant's development.
Dog-reared rhesus monkeys were more active in social situations than those reared with inanimate surrogates (Capitanio, 1984). However, in the current study the infant chimpanzee showed no difference in play behavior and decreased some locomotor behaviors when the dog was present. Thus, the third hypothesis of the dog increasing locomotor activity and play behavior was not supported. These behaviors were not eliminated by the dog's presence, but were simply reduced. It appeared that the infant spent time watching the dog while lying stationary rather than moving around himself. This infant was quite young; vigorous social play and grooming had not yet developed. Based on subjective impressions of other infants in our nursery living with dogs, we expect playing and grooming to increase as the infant matures.
Fritz and Fritz (1985) have recommended allowing a dog to assist in the rearing of infants and note that the dog may act as a playmate, groomer, groomee, and disciplinarian. They have suggested that infants raised with human caretakers, dogs, and peers exhibit more exploring and play and less rocking than those reared just with humans and peers. Our current study has shown quantitatively that rocking was decreased in this subject when the dog was present, thus offering statistical support for the Fritzs' observation. Dogs at our facility seem to act as disciplinarians, sometimes barking, snarling, or snapping at a chimpanzee if play gets too rough. This discipline may be important because nursery-reared chimpanzees show rough behavior toward peers and may have difficulties in interpreting social communication signs (Nankivell, Fritz, Nash, & Fritz, 1988). Dogs may help to reduce this potential behavioral problem. No injuries have resulted from the dogs' disciplinary behavior, which may prove beneficial when resocializing nursery-reared infants with other chimpanzees. At our facility we begin the process of supervised introduction of the infant to the dog when the infant is a few weeks old. We have safely housed dogs with single infants as well as with small groups of infants, until infants reach about 3 years of age. One potential negative outcome of housing dogs with chimpanzees is disease transmission between them. At the U.T. facility we have observed no such problems after housing two dogs with a total of 12 nursery-reared chimpanzees over a five-year period. The dogs are on a regular vaccination program and are monitored closely for internal and external parasites.
Although quantitative data describing the role of dogs in raising young chimpanzees are few, this pilot study shows many potential benefits for the infants. Reductions in rocking and other stress-related behaviors in the dog's presence is a strong argument for providing nursery-reared chimpanzees with dog companions. We highly recommend nursery-rearing chimpanzees with dogs as social companions, particularly when no other primates of appropriate ages are available as social partners. We plan future studies involving more subjects raised with dogs over a longer term to document the effects of this rearing strategy as the chimpanzees reach maturity.
References
Baumeister, A., & Forehand, R. (1973). Stereotyped acts. International Review of Research in Mental Retardation, 6, 55-96.
Berkson, G. (1968). Development of abnormal stereotyped behaviors. Developmental Psychobiology, 1, 118-132.
Berkson, G. & Mason, W. (1964). Stereotyped behaviors of chimpanzees; Relation to general arousal and alternative activities. Perceptual and Motor Skills, 19, 635-653.
Capitanio, John P. (1984). Early experience and social processes in rhesus macaques (Macaca mulatta): 1. Dyadic social interaction. Journal of Comparative Psychology, 98, 35-44.
Capitanio, J. P. (1986). Behavioral pathology. In G. Mitchell & J. Erwin (Eds.), Comparative primate biology: Behavior, conservation, and ecology (Vol. 2, Part A, pp. 411-454). New York: Alan R. Liss.
Davenport, R. K. & Menzel, E. W. Jr. (1963). Stereotyped behavior of the infant chimpanzee. Archives of General Psychiatry, 8, 115-120.
Davenport, R. & Rogers, C. M. (1970). Differential rearing of the chimpanzee. The Chimpanzee, 3, 337-360.
Dienske, H. & Griffin, R. (1978). Abnormal behavior patterns developing in chimpanzee infants during nursery care -- a note. Journal of Child Psychology and Psychiatry, 19, 387-391.
Fritz, J. & Fritz, P. (1985). The hand-rearing unit: Management decisions that may affect chimpanzee development. In C. E. Graham & J. A. Bowen (Eds.), Clinical management of infant great apes (pp. 1-34). New York: Alan R. Liss.
Mason, W. A. & Capitanio, J. P. (1988). Formation and expression of filial attachment in rhesus monkeys raised with living and inanimate mother substitutes. Developmental Psychobiology, 21, 401-430.
Mason, W. A. & Kenney, M. D. (1974). Redirection of filial attachments in rhesus monkeys: Dogs as mother surrogates. Science, 183, 1209-1211. Nankivell, B., Fritz, J., Nash, L., & Fritz, P. (1988). Competent maternal behavior by a hand-reared, resocialized, primiparous chimpanzee: A case history. Laboratory Primate Newsletter, 27 [4], 5-8.
Nicolson, N. (1977). A comparison of early behavioral development in wild and captive chimpanzees. In S. Chevalier-Skolnikoff & F. E. Poirier (Eds.), Primate bio-social development (pp. 529-560). New York: Garland.
Rogers, C. M. & Davenport, R. K. (1969). Effects of restricted rearing on sexual behavior of chimpanzees. Developmental Psychology, 1, 200-204.
Rogers, C. M. & Davenport, R. K. (1970). Chimpanzee maternal behavior. The Chimpanzee, 3, 361-368.
Struthers, E. J., Rodriguez, P., Cooper, P., & Rowell, J. (1990). Xenospecific enrichment at the Primate Research Institute. Laboratory Primate Newsletter, 29[2], 14-15.
Turner, C. H., Davenport, R. K., & Rogers, C. M. (1969). The effect of early deprivation on the social behavior of adolescent chimpanzees. American Journal of Psychiatry, 125, 85-90.
Walsh, S., Bramblett, C. A., & Alford, P. L. (1982). A vocabulary of abnormal behaviors in restrictively reared chimpanzees. American Journal of Primatology, 3, 315-319.
-------------------------------------------------------------------
Second author's address: University of Texas M. D. Anderson Cancer
Center, Science Park, Dept. of Veterinary Resources, Route 2, Box
151-B1, Bastrop, TX 78602.
The authors would like to thank Dr. P. L. Alford and the chimpanzee
caretaking staff for their help with this project. Thanks also to Carolyn Crain for manuscript preparation. This work was financially supported by the National Institutes of Health, National Center for
Research Resources grants R24-RR03578 and U42-RR03589.
* * *
Jennifer Kopecky and Viktor Reinhardt
Wisconsin Regional Primate Research Center
Introduction
Pending federal rules require that "the physical environment in the primary enclosures" of nonhuman primates "must be enriched by providing means of expressing noninjurious species-typical activities." As examples of environmental enrichment, the rules list "perches, swings, mirrors." It has been shown that perches as well as mirrors distract selected nonhuman primate species, including rhesus macaques, by stimulating such activities for a substantial portion of time (Anderson, 1983; Bramblett, 1989; Reinhardt, 1990; Schmidt et al., 1989; Suarez & Gallup, 1985; Wolff, 1989).
The rules also list swings as examples of environmental enrichment. Intuition may suggest that caged monkeys would spend most of their time doing acrobatics if only they were provided with a swing. Supportive data, however, are lacking and it is unknown if a swing is indeed a useful object to enrich the environment of caged nonhuman primates. The present study addresses this issue.
Methods
Fourteen adult (older than 5 years) laboratory-reared female rhesus macaques, Macaca mulatta, were observed for 60 minutes between 11:00-13:00. The monkeys were housed in pairs in stainless-steel cages 85 cm deep, 170 cm wide, and 85 cm high. One half of each cage was provided with a perch, the other with a swing. The animals had been exposed to both objects for more than one month. Swings and perches were of the same material (polyvinyl chloride pipes), and had the same diameter (4.8 cm). Perches were 120 cm long; they were installed diagonally at a mean height of 50 cm (see description in Reinhardt, 1990). The swings were 53 cm long; they were suspended in the center of the cage with two chains from the cage ceiling, also at a height of 50 cm.
Observations were made by the first author, who was familiar to the study subjects. All physical contact with the swing and perch was recorded with a stopwatch, and the percentage of time each animal spent with either of the two objects was computed.
Results
Of the 14 rhesus monkeys tested, 11 used the perches and 10 used the swings during the one hour observation. The animals spent an average of 10.8(+/- 15.6)% of the time with the perch, but only 1.4(+/- 2.1)% of the time with the swing. The difference was statistically significant (p<0.05; Mann Whitney test). The swing elicited the following species-typical behaviors: climbing, swinging, balancing, perching.
Discussion
The adult female rhesus monkeys in this study showed a clear preference for perches, using them almost 8 times as much as the swings. Swings elicited noninjurious, species-typical behavior patterns, but the amount of time they distracted the animals was negligible.
The animals' preference for perches was probably related to the fact that perches, unlike swings, are fixed structures permitting continuous relaxed postures rather than short-term balancing. Moreover, perches, unlike swings, permit the animals to sit right in front of the cage with optimal visual control of the environment outside of the cage. While sitting on the perch, the monkeys may, therefore, experience a stronger feeling of security than while balancing on the swing.
The study leads to the conclusion that swings offer a different reinforcement than perches and, since they are used much less often, should not be the only enriching device in a cage. Future investigations should evaluate possible species differences.
References
Anderson, J. R. (1983). Mirror-image stimulation and short separations in stumptail monkeys. Animal Learning and Behavior, 11, 139-143.
Bramblett, C. A. (1989). Enrichment options for guenons in the laboratory. American Journal of Primatology, Supplement, 1, 59-63.
Reinhardt, V. (1990). Comparing the effectiveness of PVC perches versus wooden perches as environmental enrichment objects for singly caged rhesus monkeys. Laboratory Primate Newsletter, 29[1], 13-14.
Schmidt, E. M., Dold, G. M., & McIntosh, J. S. (1989). A perch for primate squeeze cages. Laboratory Animal Science, 39, 166-167.
Suarez, S. D. & Gallup, G. G,. (1986). Social responding to mirrors in rhesus macaques (Macaca mulatta): Effects of changing mirror location. American Journal of Primatology, 11, 239-244.
U.S. Department of Agriculture (1990). Animal Welfare; Standards; Proposed Rule. Federal Register, 55, 33521-33531.
Wolff, A. V. (1989). Polyvinyl chloride piping as perch material for squirrel monkeys. Laboratory Primate Newsletter, 28[1], 7.
-------------------------------------------------------------------
Authors' address: Wisconsin Regional Primate Research Center, 1223 Capitol Court, Madison, WI 53715. This project was supported by NIH grant RR-00167 to the WRPRC. Caged rhesus monkeys were provided with both a swing and a perch. The time individuals spent using the swing was compared with that spent using the perch to determine which object provides more effective enrichment.
* * *
Kathryn Bayne, Sandy Dexter, and Stephen Suomi
National Institutes of Health
Introduction
As the USDA formulates regulations for Part 3 of the 1985 amendment to the Animal Welfare Act, a clear trend toward social housing of group-living species of nonhuman primates is evident in published draft proposals (Federal Register, 1989, 1990). It is documented for some species of primates that experience with conspecifics is requisite for normal reproductive and maternal behaviors (Fritz, 1989; Harlow, 1962; Harlow & Harlow, 1971; Mason, 1968; Moor-Jankowski & Mahoney, 1989). Sustained contact with conspecifics can ameliorate many of the aberrant behaviors caused by rearing or holding conditions with insufficient social stimulation (Fritz, 1989; Reinhardt et al., 1988; Suomi & Harlow, 1972).
One primate species commonly used in the laboratory is the capuchin monkey (Cebus apella). Under free-ranging conditions it lives in groups of all age/sex combinations of 10 to 20 individuals (Robinson & Janson, 1987; Welker et al., 1990). In the laboratory capuchins are housed in a number of ways, including individually and in small groups. Incomplete data are available regarding the impact of individual housing on the behavior of capuchins, or on the potential for social recovery (i.e., the return to a normal behavioral profile after exposure to appropriate environmental stimuli) by this species. We conducted a pilot study on the behavior of individually housed capuchins and the changes in their behavior when they were housed socially.
Subjects
Four adult male (6 yrs of age), two adult female (6 yrs of age), and one juvenile (1 yr old) capuchin were used in this study. The animals came from diverse backgrounds, but were adapted to the laboratory during a quarantine period. A behavioral profile was obtained for each animal (see Table 1).
+---------------------------------------------------+
|A153 (Pierce) Male, Age = 10 years. Timid, |
| loner, associates with A154, |
| dominant as of May 1990. |
| |
|A154 (Thor) Male, Age = 10 years. Very |
| aggressive, dominant. |
| |
|A155 (Eddie) Male, Age = 10 years. Associ- |
| ates with A156, subordinate. |
| |
|A156 (Tippy) Male, Age = 10 years. Subor- |
| dinate. |
| |
|D524 (Basil) Female, Age = 10 years. Asso- |
| ciates with D525, huddles, shy, |
| dominant as of May 1990. |
| |
|D525 (Vega) Female, Age = 10 years. More |
| aggressive, interacts frequently |
| with the males, dominant. | |
| |
|D526 (Nike) Female, Age = 1 year. Timid, |
| huddles, circles. |
+---------------------------------------------------+Table 1: Capuchin Demographics
Methods
An instantaneous sampling technique (Martin & Bateson, 1986) was used to evaluate the subjects in each of the two housing conditions. Sessions lasted 5 minutes, with observations made every 10 seconds. Each animal was evaluated once a day, some time between 0900 and 1400 hours, for a total of 34 days in each housing condition. A check sheet of 9 qualitative or quantitative abnormal behaviors was used to record the subjects' activities during the observation session. Passive behavior and environmental exploration were also recorded (see Table 2) .
+--------------+-------------------------------+
|Category |Behavior |
+--------------+-------------------------------+
| | |
|Stereotypic |Auto-Erotic |
| |Rock |
| |Huddle |
| |Stereotypic (e.g., salute, |
| palate-rubbing, regurgitation)|
+--------------+-------------------------------+
|Repetitive |Locomotion (fixed, rigid, |
|Locomotion | and redundant patterns) |
+--------------+-------------------------------+
|Self-Directed |Self-Bite |
| |Self-Mouth |
| |Self-Abuse (e.g., slapping, |
| | head banging, hair plucking) |
+--------------+-------------------------------+
|Self-Groom |Excessive Self-Groom |
+--------------+-------------------------------+
|Passive |Sitting or standing motionless |
+--------------+-------------------------------+
|Environmental |Environmental Exploration |
|Exploration | (e.g. manipulation of cage) |
+--------------+-------------------------------+Table 2: Behaviors Recorded on Check Sheet and Categories for Analysis.
All animals were initially housed in Group 3 stainless steel cages (in accordance with the Guide for the Care and Use of Laboratory Animals, 1985). The animals had lived in these cages for about 2 months before the study began. The animals were on a 12:12 light:dark cycle, and were fed twice daily, with water ad libitum. The animals had visual, auditory, and olfactory contact, with limited physical contact between the cages. No specific enrichment devices were included in the cages. However, many of the animals perched on the squeeze apparatus.
After the behaviors of the animals were recorded in the single cage condition, the animals were formed into a compatible group and housed in an indoor/outdoor run (total space 20' x 20') equipped with multiple swings and perches. The photoperiod varied slightly with the summer season, as did the diurnal ambient temperatures.
Statistical Analysis
The 11 behaviors were collapsed into six categories for statistical analysis (Passive, Repetitive Locomotion, Self-Grooming, Self-Directed, Stereotypic, and Environmental Exploration). The Wilcoxon Signed Ranks Test was used to compare the percent of observations for each behavior category for the two housing conditions.
Results
All of the behaviors on the check sheet were observed in the subject group. Table 3 presents the percent of observations recorded for each of the six behavioral categories by animal. Two of the behaviors occurred at significantly different rates between the individual housing condition and the group housing arrangement. Specifically, more stereotypic behavior was recorded in the baseline condition than the experimental condition (p < .04) and more passive behavior was recorded when the monkeys were housed socially than when they were housed individually (p < .02). None of the other behaviors showed a significant change in occurrence between the two housing conditions (see Figure 1).
+-------+------+-----+----+-----+-------+-------+
|Animal |Stereo|Loco |Self|Self-|Passive|Environ|
|Number | | | |Groom| |Explore|
+-------+------+-----+----+-----+-------+-------+
|A153 | | | | | | |
| BASE |20.00 |21.32|2.00|7.32 |28.66 |20.66 |
| EXPT |.29 |17.25|--- |6.37 |76.28 |11.29 |
| | | | | | | |
|A154 | | | | | | |
| BASE |1.34 |41.32|2.00|11.32|33.34 |10.0 |
| EXPT |1.28 |18.47|.20 |18.33|61.72 |9.77 |
| | | | | | | |
|A155 | | | | | | |
| BASE |3.34 |37.34|--- |5.34 |22.68 |31.32 |
| EXPT |.29 |52.55|.29 |2.74 |44.12 |46.66 |
| | | | | | | |
|A156 | | | | | | |
| BASE |3.98 |19.34|5.34|6.66 |31.32 |33.32 |
| EXPT |--- |25.98|--- |5.78 |68.24 |12.62 |
| | | | | | | |
|D524 | | | | | | |
| BASE |--- |22.86|1.43|7.60 |54.29 |13.8 |
| EXPT |.29 |24.02|.29 |4.41 |70.88 |11.22 |
| | | | | | | |
|D525 | | | | | | |
| BASE |20.00 |21.40|.94 |--- |34.30 |23.3 |
| EXPT |.10 |31.67|--- |10.10|58.14 |10.33 |
| | | | | | | |
|D526 | | | | | | |
| BASE |41.40 |17.60|.47 |7.16 |28.60 |4.76 |
| EXPT |11.57 |27.16|4.02|6.18 |51.08 |24.55 |
| | | | | | | |
|x BASE |12.87 |25.88|1.74|6.48 |33.31 |19.59 |
|x EXPT |1.97 |28.16|0.69|7.70 |61.49 |17.92 |
+-------+------+-----+----+-----+-------+-------+Table 3: Percent of Time-Sampling Observations
Figure 1: The overall mean percent of occurrence for each recorded category of behavior when all animals are considered together.
Discussion
Several behaviors which were considered abnormal in either the quality or quantity of their occurrence were recorded in the subjects. The most prevalent behaviors recorded followed a pattern that approximates that observed for other nonhuman primates (Bayne et al., 1991; Line et al., 1990): passive, repetitive locomotion, environmental exploration, stereotypic, self-grooming, and self-directed in the individual housing condition; passive, repetitive locomotion, environmental exploration, self-grooming, stereotypic, and self-directed in the social housing condition.
The social housing condition effectively reduced stereotypic behaviors; however, it also was associated with more passive behaviors being exhibited by the subjects. The concurrent shifts in these components of the behavioral repertoire suggests that the animals were in a calmer state when housed socially.
Although neither locomotion nor environmental exploration changed significantly between the two housing conditions, it must be remembered that these behaviors were recorded at approximately the same rate even though the floor area available to the animals was increased almost 100 times (from 4.3 sq-ft in the single cage to 400 sq-ft in the indoor/outdoor run). Thus, it may be proposed that on a square foot basis both locomotion and exploratory behaviors were reduced in the experimental housing condition.
The reduction of the occurrence of a behavior did not appear to be related to the level of motor activity involved with the behavior. For example, two low-level activity behaviors, self-directed and grooming, did not change significantly between the two housing conditions, while passive behavior did. Conversely, repetitive locomotion, a high activity behavior, and stereotypic, frequently a stationary behavior, were both affected by social housing and increased space.
Finally, based on these data, it is reasonable to suggest that individual housing has some adverse behavioral consequences for capuchin monkeys. The effect was primarily fixed, repetitive locomotion and stereotypic behaviors. It does appear, however, that even during this short pilot study some degree of social recovery was achieved as reflected by the significant reduction of stereotypic behaviors.
References
Bayne, K., Mainzer, H., Dexter, S., Campbell, G., Yamada, F., & Suomi S. (1991). The reduction of abnormal behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology, 23, 23-35.
Fritz, J. (1989). Resocialization of captive chimpanzees: An amelioration procedure. American Journal of Primatology, Suppl. 1, 79-86.
U.S. Dept. of Health and Human Services (1985). Guide for the Care and Use of Laboratory Animals. Bethesda: PHS. NIH Publication 86-23, Revised.
Harlow, H. R. (1962). The heterosexual affectional system in monkeys. American Psychologist, 17, 1-9.
Harlow, H. F., & Harlow, M. K. (1971). Psychopathology in monkeys. In H. D. Kimmel (Ed.), Experimental Psychopathology: Recent research and theory (pp. 203-229). New York: Academic Press.
Line, S. W., Morgan, K. N., Roberts, J. A., & Markowitz, H. (1990). Preliminary comments on resocialization of aged rhesus macaques. Laboratory Primate Newsletter, 29[1], 8-12.
Martin, P. & Bateson, P. (1986). Measuring behavior: An introductory guide. Cambridge: Cambridge University Press.
Mason, W. A. (1968). Early social deprivation in the nonhuman primates: Implications for human behavior. In D. Glass (Ed.), Biology and behavior: Environmental influences (pp. 70-137). New York: Rockefeller University Press.
Moor-Jankowski, J., & Mahoney, C. J. (1989). Chimpanzees in captivity: Humane handling and breeding within the confines imposed by medical research and testing. Journal of Medical Primatology, 18, 1-26.
Reinhardt, V., Houser, D., Eisele, S., Cowley, D., & Vertein, R. (1988). Behavioral responses of unrelated rhesus monkey females paired for the purpose of environmental enrichment. American Journal of Primatology, 14, 135-140.
Robinson, J. G. & Janson, C. H. (1987). Capuchins, squirrel monkeys and atelines: Socioecological convergence with old world primates. In B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, & T.T. Struhsaker (Eds.), Primate societies (pp. 69-82). Chicago: University of Chicago Press.
Suomi, S. J. & Harlow, H. F. (1972). social rehabilitation of isolate-reared monkeys. Developmental Psychology, 6, 487-496.
U. S. Department of Agriculture (1989). Animal Welfare; Proposed Rules. Federal Register, 54, 10822-10954.
U. S. Department of Agriculture (1990). Animal Welfare; Standards; Proposed Rule. Federal Register, 55, 33448-33531.
Welker, C., Becker, P., Hohmann, H., & Schafer-Witt, C. (1990). Social relations in groups of the black-capped capuchin (Cebus apella) in captivity: Interactions of group-born infants during their second half-year of life. Folia Primatologica, 54, 16-33.
-------------------------------------------------------------------
First author's address: Office of Animal Care and Use, National
Institutes of Health, Bldg. 14D, Room 313, Bethesda, MD 20892.
The authors would like to express their gratitude to Ms. Amy Onufrock for her assistance with this study and Ms. Kristin Abbot for providing the demographic data.
* * *
Alfonso Gozalo and Enrique Montoya
Proyecto Peruano de Primatologia
Introduction
Klebsiella pneumoniae infections have occurred in Old and New World monkeys (Fiennes, 1967, Houser et al., 1970; Hunt et al., 1968; Snyder et al., 1970; Schmidt & Butler, 1971) and are reported to cause pneumonia (Fiennes, 1967), meningitis, peritonitis, cystitis (Houser et al., 1970), and lesions indicative of septicemia (Snyder et al., 1970; Schmidt & Butler, 1971; McClure et al., 1986). In humans the disease spectrum includes severe pneumonia, lobular or lobar, urinary tract infection, and miscellaneous septic lesions, but no enterotoxicity (Von Lichtenberg, 1989).
In this report we describe the clinical course, symptoms, postmortem gross and microscopic lesions, and control of an outbreak of K. pneumoniae infection in a New World nonhuman primate colony.
Subjects and Methods
The animal colony at the Center for Reproduction and Conservation of Nonhuman Primates (CRCP) in Iquitos, Peru contains an average of about 700 monkeys (420 Aotus sp., 70 Saguinus sp., and 210 Saimiri sp.), housed in separate buildings according to species. Six species (Aotus nancymae, A. vociferans, Saguinus labiatus, S. mystax, Saimiri boliviensis peruviensis, and S. sciureus macrodon) are maintained for breeding purposes, and small numbers of other genera are maintained for conservation purposes (Ateles, Cacajao, Callimico, Cebus, Lagothrix). The monkeys are kept in open-system breeding units, consisting of concrete and wooden-frame wire-covered pens inside a block construction, screened building. Pens are approximately 1m x 1m x 2m. The monkeys are fed a standard in-house prepared, grain-based baked diet, plus ripe bananas and water ad-lib.
Between May and October 1990, a number of monkeys in this colony died suddenly. Necropsies were performed on all dead monkeys except those with advanced autolysis. Tissue samples were taken for microbiological and histological analysis. Microbiological analyses were performed according to standard procedures (WHO, 1987). Antibiotic sensitivity testing was performed on all isolates. Tissue samples were fixed in 10% neutral buffered formaline, embedded in paraplast, sectioned at 5m, and stained routinely with hematoxilin and eosin.
Results
Seventeen monkeys died as a consequence of purulent or septicemic lesions (Table 1). Klebsiella pneumoniae was isolated from 14 of the 17 cases; three cases were not cultured due to advanced autolysis. Due to the acute course of the disease, antemortem diagnosis was often not made. In a few cases diarrhea or prostration and anorexia was observed prior to death. The disease affected mostly Saguinus and Aotus monkeys, and one case was observed in a juvenile male Saimiri (Table 1). The highest mortality, ten cases, occurred in September. At necropsy, purulent peritonitis was the most common finding, followed by pyothorax, purulent lobular pneumonia, and hepatic abscess (Table 1). In the Saguinus monkeys the most common finding was purulent peritonitis with no evidence of intestinal involvement. Mesenteric lymph nodes were swollen and abscessed. In Aotus, acute inflammation and suppuration of the Peyer's patches and lymphoid tissue of the cecum was the first lesion noted, followed by abscess formation and peritoneal cavity infection. One case, of pyothorax and peritonitis, was observed in a juvenile male Saimiri. Antibiotic sensitivity testing of K. pneumoniae isolates revealed that it was sensitive to Gentamicin, Tetracycline, and Flumequine. We decided to use water soluble Tetracycline (55 mg/k body weight for five consecutive days) in the drinking water to avoid handling and stressing the monkeys. Only one more monkey died after therapy was instituted.
+-------------------------------------------------------------+ |Date Species Sex Age Lesions Microorganism | | isolated | +-------------------------------------------------------------+ |May 12 S. labiatus F A Peritonitis not done | |Jun 6 A. nancymae M J Peritonitis K. peumoniae | |Aug 11 S. mystax M A Peritonitis K. pneumoniae | |Aug 13 S. mystax F I Pneumonia K. pneumoniae | |Aug 28 S. mystax M J Peritonitis K. pneumoniae | |Aug 30 S. mystax F A Peritonitis K. pneumoniae | |Sep 3 S. mystax F A Peritonitis K. pneumoniae | |Sep 7 A. vociferans F A Peritonitis not done | |Sep 8 A. vociferans M A Peritonitis K. pneumoniae | |Sep 9 S. labiatus F A Peritonitis K. pneumoniae | |Sep 10 S. mystax F A Peritonitis K. pneumoniae | |Sep 11 A. nancymae M J Peritonitis K. pneumoniae | |Sep 11 A. nancymae F A Peritonitis not done | |Sep 13 S. mystax M A Hepatic abcess K. pneumoniae | |Sep 14 S. mystax F A Pyothorax & not done | | Peritonitis | |Sep 18 Saimiri s. m. M J Pyothorax & K. pneumoniae | | Peritonitis | |Oct 1 A. nancymae M J Peritonitis K. pneumoniae | +-------------------------------------------------------------+
Table 1: Monkeys which died from Klebsiella infection at the CRCP in 1990.
Discussion
Acute septicemia and bronchopneumonia from Klebsiella infection causes high mortality in owl monkeys (Snyder et al., 1970). Air sacs have been proposed as a potential primary site for Klebsiella infection (Giles et al., 1974). However, inoculation of the microorganism into the air sacs of three healthy owl monkeys failed to produce lesions (Giles et al., 1974). According to Fiennes (1967), Klebsiella is commonly isolated from the mouths of many healthy monkeys. It is generally considered to be an opportunistic pathogen that causes disease when an animal is stressed or exhibits an altered defense mechanism (Giles et al., 1974; Snyder et al., 1970; Weil, 1966). According to Snyder et al. (1970), the main pathologic finding in 23 owl monkeys dying from Klebsiella infection was septicemia (73%). In owl monkeys K. pneumoniae produces inflammation and suppuration of the Peyer's patches and lymphoid tissue in cecum, and peritonitis with septicemia, with or without intestinal ulceration and abscesses of the mesenteric lymph nodes. Clinically the monkeys exhibit fever up to 106~F, and have distended, painful abdomens upon palpation. Seven isolates from K. pneumoniae were identified at the Centers for Disease Control in Atlanta as belonging to the K65 serotype, and three samples obtained from the colons of apparently healthy monkeys were found to belong to the K4, K65, and K70 serotypes (Escajadillo, 1984). Apparently wild Aotus have no contact with K. pneumoniae, being very susceptible to a pathogenic strain (Escajadillo, 1984; Giles et al., 1974). A killed-virus vaccine from one strain of K. pneumoniae isolated from Aotus was administered with poor results (Escajadillo, personal communication).
We believe that Saguinus and Aotus monkeys are more susceptible to pathogenic strains of K. pneumoniae than other New World monkeys. In our opinion the most frequent route of infection is fecal-oral and, less frequently, inhalation. We have not observed air sacculitis in the owl monkeys from our colony. Pathologically there are differences between Saguinus and Aotus infections that could be due to genetic or anatomic bases. Most of the monkeys infected were in good condition with no signs of illness or weight loss. Occasionally we have observed cases of purulent peritonitis caused by K. pneumoniae in Aotus monkeys from our colony (four cases in 1989). We have also isolated the microorganism from diarrheic as well as normal monkey feces and food scraps. It is unclear if the monkeys were or were not exposed to this bacterium in the wild; it is more probable that they were exposed to a highly pathogenic strain of this microorganism in captivity (see the serotyping done in Panamanian owl monkeys by Escajadillo, 1984). The control and prevention of this infection is feasible by providing the monkeys via drinking water with an adequate antibiotic determined by a sensitivity test performed on isolates from this pathogenic bacterium.
References
Escajadillo, A. (1984). Mantenimiento en el Laboratorio y algunos usos experimentales del mono nocturno Aotus. Revista Medica de Panama, 9, 274-287.
Fiennes, R. (1967). Zoonoses of primates. Ithaca: Cornell University Press.
Giles, R. C. Jr., Hildebrandt, P. K., & Tate, C. (1974). Klebsiella air sacculitis in the owl monkey (Aotus trivirgatus). Laboratory Animal Science, 24, 610-616.
Houser, W. D., Norback, D. H., & Ragland, W. L. (1970). Atypical Klebsiella infections in infant monkeys. Mainly Monkeys, 1, 20.
Hunt, D. E., Pitillo, R. F., Deneau, G. A., Schabel, F. M. Jr., & Mellett, L. B. (1968). Control of an acute Klebsiella pneumoniae infection in a rhesus monkey colony. Laboratory Animal Care, 18, 182-185.
McClure, H. M., Brodie, A. R., Anderson, D. C., & Swenson, R. B. (1986). Bacterial infections of nonhuman primates. In K. Benirschke (Ed.), Primates: The road to self-sustaining populations (pp. 531-556). New York: Springer Verlag.
Schmidt, R. E. & Butler, T. M. (1971). Klebsiella-Enterobacter infections in chimpanzees. Laboratory Animal Science, 21, 946-949.
Snyder, S. B., Lund, J. E., Bone, J., Soave, O. A., & Hirsch, D. C. (1970). A study of Klebsiella infections in owl monkeys. Journal of the American Veterinary Medical Association, 157, 1935-1939.
Von Lichtenberg, F. (1989). Infectious disease: Viral, chlamydial, rickettsial, and bacterial diseases. In W. B. Saunders' Staff (Eds.), Robbin's pathologic basis of disease, 4th edition (pp. 307-384). Philadelphia: W. B. Sanders Co.
Weil, A. J. (1966). Nosocomial infection with Klebsiella, type 25. New England Journal of Medicine, 275, 17-22.
World Health Organization (1987). Second part of bench-level procedure manual on basic bacteriology. WHO/LAB/87.1, 63.
-------------------------------------------------------------------
First author's address: Proyecto Peruano de Primatologia-IVITA, Apartado 621, Iquitos, Peru. This work was done as part of the activities of the Peruvian Primatological Project Manuel Moro Sommo supported by the Peruvian government and the Pan American Health Organization. We also want to thank Arnulfo Romaina and Jarvel Lopez for technical assistance.
* * *
Scott Willard and James Loy
University of Rhode Island
Introduction
Studies of rhesus macaques (Macaca mulatta) have shown that infants reared for long periods with only their mother as a companion often show behavioral abnormalities later in life. It is generally believed that lack of experience with other youngsters (peer deprivation) is related to the behavioral effects. Peer deprivation for the first eight to twelve months of life often results in deficiencies in affiliative behaviors, and hyperaggressiveness (Alexander, 1966; Spencer-Booth, 1969, as cited in Hinde, 1971).
This paper reports the effects of a year and a half of peer deprivation on three female patas monkeys (Cercopithecus [Erythrocebus] patas). As detailed below, the main deprivation effects in these African monkeys were maternal inadequacy and (in one case) some hyperaggressiveness.
Methods
Animals and Housing: The subjects of this study were members of the captive patas group maintained by the University of Rhode Island (URI). The URI monkeys are usually housed in large indoor gang-cages but, because of laboratory renovations, all animals were moved off campus in late 1980 and single-caged in standard baboon/macaque laboratory cages at a boarding facility. Three females were pregnant when boarded, and each gave birth to a daughter in April, 1981 (Table 1). Each female infant lived alone with its mother (although all infants could see, hear, and smell other patas monkeys, including other mother-infant pairs) for 12-19 months before the animals were returned to URI and again gang-caged. Two infants (EL and RH) experienced 18 months of mother-only rearing, while the third (VP) was mother-only reared for 12 months, until her mother died. VP was then caged alone for almost seven more months (Table 1).
+------+-------------+---------------------+------------+
| | |Length of Mother-only| |
| | | Rearing | Age When |
|Female|Date of Birth| (Peer-Deprivation) | Gang-caged |
| | | | |
+------+-------------+---------------------+------------+
| EL |27 April 1981| 18.2 months | 1.51 years |
| | | | |
| RH |23 April 1981| 18.4 months | 1.52 years |
| | | | |
| VP |15 April 1981| 18.8 months* | 1.55 years |
+------+-------------+---------------------+------------+
*VP's mother died on 11 April 1982, and thus VP was caged
alone for 6.75 months before her return to URI.
Table 1: Histories of peer-deprived patas females.
In November, 1982, the entire colony was returned to campus. The three peer-deprived females (now about 1.5 years old) were immediately introduced into a 35 m� gang-cage and housed as a trio for 71 days. In late January, 1983, three normally-reared, adult females were added to the gang-cage, and seven months later, a normally-reared, adult male was introduced into the group.
Observations: Between 18 November 1982 and 27 January 1983, the three peer-deprived females were focal sampled (Altmann, 1974) for behavioral data by JL and three undergraduate assistants. Each animal was the subject of 12-13 fifteen minute samples. Another brief period of (ad libitum) behavioral sampling occurred in September 1983, immediately after the introduction of the adult male. During both sampling periods, data were collected on agonism (fights, supplantations), sexual behavior (solicits, mounts, copulations), and affiliative behaviors (social grooming, sitting in proximity, sitting-touching) (see Loy, 1981, for definitions of behaviors). Data on the parturient and maternal behaviors of the peer-deprived females were collected ad libitum following their various pregnancies from 1984 to 1986.
Results
Peer-deprived trio (11/18/82 - 1/27/83): During their 71 days together as a trio, the peer-deprived females frequently engaged in affiliative behaviors. Sitting in proximity to one another was the most common affiliative pattern, with sitting-touching next in frequency. Although social grooming was relatively rare, all three animals participated in a few episodes. Fights and supplantations occurred much less often than affiliative interactions. Dominance relations within the trio remained ambiguous throughout the observation period.
Peer-deprived Trio plus Normal Adults (8/29/83 - 9/29/83): During the second sampling period (seven months after the introduction of the adult females, and immediately after the introduction of the adult male), the peer-deprived females (now almost 2-1/2 years old) behaved very similarly to the normally-reared adults. Although ad libitum sampling precluded rigorous interindividual comparisons, the peer-deprived females appeared to engage in almost as many affiliative interactions (mostly with each other) as the normal animals, and further, the peer-deprived monkeys seemed neither hyper- nor hypoaggressive. The dominance hierarchy of the gang-caged animals was linear (Loy & Harnois, 1988), and the three peer-deprived females occupied upper and middle ranks of the female hierarchy (second, third, and fourth out of six).
There was considerable sexual activity within the group following the adult male's introduction. Normal females gave the species-typical solicitation display (Loy, 1981) to the male several times per hour. Peer-deprived females EL and VP also immediately began soliciting the male, but did so much less frequently than normal females. The third peer-deprived female, RH, was not observed to give a clear solicitation display until October 1983.
Reproductive Histories of Peer-deprived Females: The reproductive histories of EL, VP, and RH are summarized in Table 2. EL reproduced first; she conceived in October 1983, and gave birth in April 1984. VP's first conception occurred in June 1984 and RH's in January 1985.
+------+-------------+-----------+---------------+
| |Approx. Dates| Dates of |Fate of Infants|
|Female|of Conception|Parturition| and Remarks |
+------+-------------+-----------+---------------+
| EL | 19 Oct 1983 | 5 Apr 1984|Removed DOB and|
| | | |hand-raised |
| | | | |
| EL | 20 Sep 1984 | 8 Mar 1985|Removed DOB and|
| | | |hand-raised |
| | | | |
| EL | 23 Aug 1985 | 8 Feb 1986|Mother-reared |
| | | |for 3 days, |
| | | |removed and |
| | | |hand-raised |
| | | | |
| RH | 7 Jan 1985 |25 Jun 1985| Died DOB |
| | | | |
| RH | 2 Oct 1985 |20 Mar 1986| Died DOB |
| | | | |
| VP | 21 Jun 1984 | 7 Dec 1984| Died DOB |
| | | | |
| VP | 6 Jul 1985 |22 Dec 1985|Removed DOB for|
| | | |hand-raising, |
| | | |died next day |
| | | | |
| VP | 2 Jun 1986 |18 Nov 1986|Mother-raised, |
| | | |first in single|
| | | |cage, then in |
| | | |group |
+------+-------------+-----------+---------------+
Table 2: Reproductive histories of peer-deprived patas females. Conception dates calculated by backdating 170 days from DOB.
Between April 1984 and November 1986, the three peer-deprived females produced eight full-term infants, only four of whom survived. The females handled their new infants awkwardly, occasionally dropping them roughly to the floor. Furthermore, in contrast to normal patas mothers who are extremely attentive, the peer-deprived females often left their babies unattended and lying for long periods on the cold concrete floor of the gang-cage. Two babies (RH-86, VP-84) were found dead on the mornings of their births. Two others (RH-85, VP-85) were alive when they had to be removed on the day of birth because of maternal neglect or rough handling; both died within 24 hours. Neither RH nor VP showed any noticable improvement in maternal behavior between their first and second infants.
The four cases of infant survival were all due to human intervention. VP was isolated in a small cage a few days before her third baby (VP-86) was due. She gave birth in isolation, and successfully reared her third infant first in the small cage and then in the group. EL's first two babies were removed from the group on the day of birth after having been left on the floor so long they were in danger of hypothermia. Each was hand-raised for three months before being reintroduced into the group. EL showed significant improvement in maternal behavior with her third baby. This infant was carefully cleaned and groomed after its birth, then huddled and carried ventro-ventrally. After three days of quite normal maternal care and nursing, however, EL abandoned her third baby on the floor and it, too, had to be removed and hand-reared (see Hemmalin & Loy, 1989, for additional details on EL's parturient and postpartum behaviors).
EL's infants were hand-reared for three months each and then reintroduced into the group. For unknown reasons, EL was extremely aggressive toward her returning offspring. After the introduction of both her first and second babies, EL attacked them so severely that she had to be removed from the group for several weeks to insure the infants' safety.
Discussion
Alexander (1966) observed rhesus monkeys that were mother-only reared (and thus peer-deprived) for the first four to eight months of life and found that they showed subtle behavioral effects. When tested shortly after deprivation, the young monkeys were somewhat more aggressive and less affiliative than normal animals (but see Griffin, 1966, for contrasting results), and by adulthood they were definitely "socially deficient" (Mitchell, 1970, p.210). Similarly, Spencer-Booth (1969, as cited in Hinde, 1971) found that rhesus infants that were mother-only reared for one year proved to be less social than group-reared infants.
In several ways, the three patas females observed at URI seemed to show fewer behavioral effects of peer-deprivation than reported for rhesus monkeys, although differences in study designs make this a very tentative conclusion. When observed as young adults, our deprived patas monkeys engaged in both affiliative and agonistic interactions nearly as often as normally-reared monkeys. Only one peer-deprived patas female (EL) showed any evidence of unusual aggressiveness and then only in one particular circumstance, when her hand-reared infants were reintroduced to the gang-caged group. This behavior reminds one of Alexander's (1966) observation of eight month peer-deprived rhesus monkeys who were extremely aggressive toward strange infants added to their group.
All three peer-deprived patas females began to show normal sexual behavior soon after exposure to an adult male, and all conceived and carried infants to term. Following parturition, however, all three showed strong maternal inadequacy, expressed mainly as a lack of attentiveness, and awkward or rough handling of their babies. Only one female, EL, showed spontaneous improvement in her maternal behavior. EL rejected her first two infants on the day of birth, but managed to care for her third infant for three days before rejecting it. VP successfully reared her third baby, but only after human intervention, i.e., isolation for parturition.
The females' poor performances as mothers were most likely related to peer-deprivation, although the exact cause-and-effect relationship is unclear. With the exception of EL's first delivery, the patas females had considerable opportunities within the gang-cage to observe adequate mothering and/or to handle other females' infants prior to the birth of their own babies (Table 3). In addition, as infants at the boarding facility, they were able to see each other being adequately mothered in nearby single cages. Such opportunities to learn adequate maternal behaviors by watching and interacting with normal mother-infant pairs had little positive effect on the peer-deprived females, however, and it seems likely that all of their infants would have died except for human intervention. In contrast, studies of rhesus monkeys have shown that when peer-deprived females are allowed, starting as four-month-olds, to watch successful mothering, they usually care appropriately for their own first (and subsequent) infants (Dienske, van Vreeswijk, & Koning, 1980). This comparison suggests that prolonged peer-deprivation due to mother-only rearing may have stronger effects on maternal behavior among patas monkeys than among rhesus macaques. Why such an interspecific difference should exist is unknown.
+-------------+--------------------+--------------------+
| | Exposure to Other |Degree of Access to |
| |Mother/Infant Pairs | Other's Infants |
+------+------+---------+----------+---------+----------+
|Mother|Infant|No. Pairs|Total Days| Good | Limited |
| EL | 1984 | 1 | 8 | x | |
| EL | 1985 | 1 | 208 | x | |
| EL | 1986 | 2 | 384 | x | |
| RH | 1985 | 1 | 365 | | x |
| RH | 1986 | 3 | 564 | | x |
| VP | 1984 | 1 | 253 | | x |
| VP | 1985 | 2 | 472 | | x |
| VP | 1986 | 4 | 746 | | x |
+------+-------+--------+----------+---------+----------+
Table 3: Preparturition exposure of peer-deprived patas females to "normal" mother-infant pairs in the URI gangcage. Females RH and VP had limited access to other females' infants because of their middle positions in the female dominance hierarchy.
References
Alexander, B. V. K. (1966). The effects of early peer-deprivation on juvenile behavior of rhesus monkeys. Unpublished dissertation, University of Wisconsin, Madison.
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227-263.
Dienske, H., van Vreeswijk, W., & Koning, H. (1980). Adequate mothering by partially isolated rhesus monkeys after observation of maternal care. Journal of Abnormal Psychology, 89, 489-492.
Griffin, G. A. (1966). The effects of multiple mothering on the infant-mother and infant-infant affectional systems. Unpublished dissertation, University of Wisconsin, Madison.
Hemmalin, H. & Loy, J. (1989). Observations of parturition among captive patas monkeys (Erythrocebus patas). American Journal of Primatology, 19, 183-188.
Hinde, R. A. (1971). Development of social behavior. In A. M. Schrier & F. Stollnitz (Eds.), Behavior of nonhuman primates (Vol. 3, pp. 1-68). New York: Academic Press.
Loy, J. (1981). The reproductive and heterosexual behaviours of adult patas monkeys in captivity. Animal Behaviour, 29, 714-726.
Loy, J. & Harnois, M. (1988). An assessment of dominance and kinship among patas monkeys. Primates, 29, 331-342.
Mitchell, G. (1970). Abnormal behavior in primates. In L. A. Rosenblum (Ed.), Primate behavior: Developments in field and laboratory research (Vol. 1, pp. 195-249). New York: Academic Press.
Spencer-Booth, Y. (1969). The effects of rearing rhesus monkey infants in isolation with their mothers on their subsequent behaviour in a group situation. Mammalia, 33, 80-86.
-------------------------------------------------------------------
Second author's address: Department of Sociology and Anthropology, University of Rhode Island, Kingston, RI 02881.
* * *
Several years ago I served as a reviewer of Mary Willard's research proposal (see this Newsletter, 1991, 30[1], 18-19) for the Veterans Administration. The biggest problem that I and other reviewers had was that it was unscientific but competing for scientific funds. After a second round of reviews, resulting primarily from pressure to see the project funded for its political value, I recommended that if the VA was determined to fund it, they should do so under the auspices of established patient therapy procedures rather than research. At that point, I lost track of the project.
Clearly, the project did not use established scientific methods; had limited substantive data to support its justification; raised numerous emotional conflicts about human health and sanitation; and raised difficult questions about animal welfare, particularly because of the aversive shock discipline procedure and total tooth extraction. Although data were limited, there also appeared to be a high failure rate among placed monkeys.
While the project makes great press, I personally concluded that beyond its scientific merit or lack thereof, I could not justify it in my own mind. -- Conrad B. Richter, Professor and Director, Div. of Laboratory Animal Resources, Duke University Medical Center, P.O. Box 3180, Durham, NC 27710.
* * *
AmFAR Scholar Awards
The American Foundation for AIDS Research (AmFAR) is dedicated to meeting an urgent need for biomedical research on AIDS and the conditions related to it. AmFAR Scholars are usually scientists who have completed two or three years of postdoctoral training, but do not have a tenured position. Scholar awards, of approximately $34,000 per year, are made for a period of three years, and are not renewable. For further information, contact AmFAR, Grants Dept., 5900 Wilshire Blvd., Second Floor, East Satellite, Los Angeles, CA 90036-5032 [213-857-5900].
Wenner-Gren Foundation for Anthropological Research
This foundation supports research in all branches of anthropology, including cultural/social anthropology, ethnology, biological/physical anthropology, archaeology, and anthropological linguistics, and in closely related disciplines concerned with human origins, development, and variation. Their deadline for applications is May 1, 1991. For more information contact: The Wenner-Gren Foundation for Anthropological Research, Inc., 220 Fifth Avenue, 16th floor, New York, NY 10001-7780 [212-683-5000].
Basic Research on Osteoporosis
Applications are invited for grants on the following basic research areas: (1) the effect of local and systemic growth regulators and osteoinductive factors on bone metabolism relevant to osteoporosis; (2) the development of animal and cell culture models for osteoporosis; (3) the role of mechanical/gravitational stress in maintaining bone mass and preventing bone loss; and (4) studies of the mechanism of action on bone of factors with therapeutic potential. The deadline for applications is May 15, 1991. For more information contact: Dr. Joan A. McGowan, Bone Biology and Bone Diseases Program Director, NIAMSD, 5333 Westbard Avenue, Westwood Bldg., Room 403, Bethesda, MD 20892 [301-496-7495].
Models for Sudden Infant Death Syndrome
The National Institute of Child Health and Human Development invites applications for studies using fetal and/or young developing animals to elucidate environmental factors and developmental mechanisms during pregnancy and early postnatal life that predispose the young animal to a SIDS-like event (i.e., sudden death during a sleep period), or the inability to recover from hypoxic or other life-threatening stresses. Developmental studies that span the fetal and early postnatal period are encouraged.
For further information, contact Marian Willinger, Ph.D., Health Scientist Administrator, Pregnancy and Perinatology Branch Center for Research for Mothers and Children, NICHHD, Executive Plaza North, Room 643, Bethesda, MD 20892 [301-496-5575]. Application deadline is May 15, 1991.
Communication Disorders Grants
The National Institute on Deafness and Other Communication Disorders announces a Small Grant Program, to provide support for pilot research to determine the feasibility of a subsequent research project. The research must be focused on areas within the mission of NIDCD, that is, hearing, balance/vestibular, smell, taste, voice, speech, or language. This Program is designed solely to support basic and clinical scientists with limited research experience. Current or previous recipients of NIH research awards (R01 or R29) are ineligible for this Small Grant Program. These funds may not be used to support thesis or dissertation research. Applicants may request up to $25,000 (direct costs) per year. The grant may not exceed two years and is not renewable. Following completion of the Small Grant support, investigators are encouraged to seek support for research through an Individual Research Project Grant (R01) or a First Independent Research Support and Transition Award (R29).
Submission dates for 1991 are May 6 and September 16. Applications and further information are available from the Scientific Review Branch, NIDCD, NIH, Executive Plaza South, Rm. 750, 6120 Executive Plaza Blvd., Rockville, MD 20852 [301-496-5061].
Lessening Animals' Pain and Distress
The National Institutes of Health (NIH) and the Alcohol, Drug Abuse, and Mental Health Administration (ADAMHA) invite grant applications for investigations into research methods that do not use vertebrate animals, use fewer vertebrate animals, or produce less pain and distress in vertebrate animals used for research.
Many strategies are currently in place to reduce the pain and distress of laboratory animals; however, new methods and technologies are encouraged.
Grant applications are requested for projects that will increase the extent and depth of knowledge needed to develop methods of biomedical research that: *reduce the number of vertebrate animals used in research *produce less pain and distress in vertebrate animals than methods currently used. Support will be provided for up to five years (renewable for subsequent periods) subject to the availability of funds and progress achieved. Research grant applications may be submitted by both nonprofit and profit-making organizations and institutions, State or local governments and their agencies, and eligible agencies of the Federal Government.
Before submitting an application, contact Dr. Louise E. Ramm, Director, Biological Models and Materials Research Program, National Center for Research Resources, NIH, Westwood Bdlg., Room 8A07, Bethesda, MD 20892 [301-402-0630].
Model for Vaccination in Pregnancy
The National Institute of Allergy and Infectious Diseases (NIAID), has a requirement to develop and characterize a non-human primate animal model system to study the effects of vaccination of pregnant females on their offspring. The long-term goal of the research is to gather evidence for the safety and efficacy of maternal immunization during pregnancy for the protection of the newborn by placentally transferred antibody against frequent infant pathogens. The purpose of this research effort would be to provide baseline information, using a primate model, on several candidate vaccines for the maternal immunization approach.
This NIAID-sponsored project will take approximately five (5) years to complete. One cost-reimbursement level of effort contract is anticipated and that one award will be made. This is an announcement for an anticipated Request for Proposals (RFP). RFP-NIH-NIAID- DMID-91-34 shall be issued on or about February 15, 1991, with a closing date tentatively set for April 9, 1991. Request for the RFP shall be directed in writing to: Phillip Hastings, Contract Management Branch, NIAID, NIH, Control Data Building, Room 326P, 6003 Executive Blvd., Bethesda, MD 20892. To receive a copy of the RFP, please supply this office with two self-addressed mailing labels.
Alzheimer's Disease
Research is invited on the etiology, pathology, diagnosis, and potential therapies for Alzheimer's disease and related cerebral degenerative disorders. Topics of interest include: (1) genetics; (2) mechanisms of cell death; (3) nerve growth factors; (4) animal modeling; and (5) neuroimaging and aspects of differential diagnosis. The deadline for applications is May 6, 1991. For more information contact: Eugene Oliver, Health Scientist Administrator, Division of Demyelinating, Atrophic and Dementing Disorders, NINDS, NIH, Federal Bldg., Rm. 806, Bethesda, MD 20892 [301-496-1431].
Cancer Research Institute
The Institute funds research aimed at furthering the development of immunological approaches to the diagnosis, treatment, and prevention of cancer. The Institute offers its Investigator Award to qualified scientists at the assistant professor level who are working in the field of cancer immunology or general immunology. The deadline for applications is May 1, 1991. For more information contact: Ms. Ursula Hahn, Grant Administrator, Cancer Research Institute, 133 East 58th Street, New York, NY 10022-1236.
Harry Frank Guggenheim Foundation
Support is offered for research to advance the understanding of human social problems related to dominance, violence, and aggression through short-term grants. These grants support scientific research and scholarly study concerning man's relations to man, including projects which are designed to reveal basic physiological mechanisms; elucidate fundamental psychological processes; analyze critical social interrelations; and formulate and test unifying theories of those mechanisms, processes and interrelations. Grants are made either to an institution on behalf of an individual or to an individual directly. Proposals may be submitted for one-, two- or three- year projects, although initial awards are for one year. Awards range from $15,000 to $35,000 a year for periods of one or two years. Support may include the costs of salaries, employee benefits, research assistants, computer time, supplies and equipment, field work, secretarial and technical help and other necessary expenses. Funds are not awarded for institutional overhead, travel to professional meetings, self education, elaborate fixed equipment or dissertation support. Deadlines are February 1 and August 1 each year. Contact the Harry Frank Guggenheim Foundation, 527 Madison Ave., New York, NY 10022-4301 [212-644-4907].
* * *
Amy E. Morris, Tulsa 200, 5701 E. 36th St. N., Tulsa, OK 74115.
F. Barbara Orlans, Kennedy Inst. of Ethics, Georgetown Univ., Washington DC 20057.
Shelly L. Williams, Language Research Center, Georgia State Univ., Atlanta, GA 30303-3083.
Ellen Mary Wilson, F.D.A., Office of Beltsville Technical Operations, Module I Facility, 8301 Muirkirk Rd., Laurel, MD 20708.
* * *
George M. Haslerud, teacher, researcher, scholar, and retired psychology professor, died April 10, 1990, in Dover, NH. A primate psychologist, he was recognized internationally for his research in learning and motivation. Professor Haslerud taught the psychology of learning, physiological psychology, and many other classes for 27 years at the University of New Hampshire, Durham, until his retirement in 1972.
In the middle and late 1930's, Haslerud studied general early behavioral development of chimpanzees, as well as the effects of movement on their avoidance reactions. He wrote the introduction to a recent reprinting of Yerkes' book, "The Mental Life of Monkeys and Apes." Yerkes was in regular contact with Haslerud and Yerkes often cited him in his own writings. Haselrud also did research on the brain areas of importance to delayed response in monkeys.
The most wonderful things about George Haslerud were the breadth of his interests and his commitment to international intellectual cooperation. While at UNH, he was awarded two Fulbright professorships. One of these, in 1953, took him to Kyoto University in Japan. In a letter to me in 1981, he stated that his visit to Japan was still paying dividends in international friendship and intellectual stimulation. On his second Fulbright, he visited the Universidad Catolica in Lima, Peru. In 1981, nine years retired, he traveled to the Middle East via one of the World Affairs Council's diplomatic-cultural tours. He and his group received briefings from the heads of administration and parliaments in various countries and saw ancient cultural sights under optimal conditions. In Egypt, they had a 40-minute conference with then-President Anwar el Sadat. In a 1981 letter to me, Haslerud called Sadat "the bravest man in the world. "
Professor Haslerud was an exceptional, compassionate teacher, able to draw from his students abilities which they did not know they had. As one of his undergraduates, I became a co-author on four publications before even considering graduate school. I have tried to encourage early research involvement in my own students ever since. His commitment to his students lasted throughout their careers. Long after his retirement he met with, and wrote to, many of my students (Maple, Erwin, Schapiro) about primatology. It was a privilege to have been one of his "academic sons. "
George Haslerud's research was reported in respected professional journals of all kinds, and his book "Transfer, Memory, Creativity " was published in 1972 by the University of Minnesota Press (UM was his alma mater).
He was an erudite, quiet, encouraging, and kind man with interests in behavior including, but not restricted to, nonhuman primates. Still, he is important in the history of behavioral primatology and he was the most important individual to me in my own developing years as a psychologist. I was drawn to primates by his many lectures and anecdotes of the Yerkes days, as were many others. I miss his letters already. He is survived by his wife, Dr. Ethelyn Haslerud; three daughters, Beth, Karen, and Anna; and three grandsons. -- G. Mitchell, Psychology Dept., Univ. of California, Davis, CA 95616.
* * *
Traveler's Diarrhea
High risk areas for traveler's diarrhea (incidence 20 to 50%) are the developing countries of Latin America, Africa, the Middle East, and Asia. There is intermediate risk in southern Europe, the USSR, and some Caribbean islands, and low risk in Canada, northern Europe, Australia, New Zealand, and the United States. The most common agents are 1) Bacteria: E. coli (most common, enterotoxigenic, enteroinvasive, enteroadherent); Salmonella, Shigella, Campylobacter jejuni, Aeromonas hydrophila, Yersinia enterocolitica, Plesiomonas Shigelloides, Vibrio cholerae (non-0l), V. fluvialis, V. parahaemolyticus. 2) Viruses: Norwalk agent; Rotavirus (?). 3) Parasites: Giardia lamblia, Entamoeba histolytica, Cryptosporidium.
Prevention is chiefly avoiding raw vegetables, raw meat, raw seafood, tapwater, ice, unpasturized milk and dairy products, and unpeeled fruit. Preventative agents with documented efficacy of 50-85% are Doxycycline (100 mg/day), Sulfa-trimethoprim (1 double strength tablet/day), and Bismuth subsalicylate (Pepto-Bismol; 2 oz or 2 300 mg tablets four times/day).
Treatment begins with oral intake to maintain fluid and electrolyte balance: potable fruit juice, caffeine-free soft drinks, and salted crackers. For severe symptoms, the WHO oral replacement solution is recommended: 3/4 tsp. salt (NaCl), 1 tsp. baking soda (NaHCO�), 1 c. orange juice or 2 bananas (for KCl), and 4 Tbsp. sugar per quart or liter of boiled water. For mild diarrhea (less than 3 stools/day, without blood, pus, or fever), the following antimotility drugs are recommended: Diphenoxylate (Lomotil, 2 2.5mg tablets 3 or 4 times/day); Loperamide (Imodium, 4mg, then 2mg after each loose stool to maximum of 16mg/day); Bismuth subsalicylate (Pepto-Bismol; 30 ml or 2 tablets every 30 minutes, up to 8 doses). For moderate or severe diarrhea, antimicrobial agents are recommended: Sulfa-trimethoprim (1 double strength tablet twice a day for 5 days); Trimethoprim (200 mg twice a day for 5 days); Doxycycline (100 mg twice a day for 5 days). For persistent diarrhea with serious fluid loss, fever, or stools showing blood or mucus, seek medical attention ! -- Adapted from NIH Consensus Development Panel, 1985. See Journal of the American Medical Society, 1985, 253, 2700.
Cholera in Peru
From January 24 through February 9, 1991, 1859 persons with symptoms of cholera (profuse watery diarrhea and vomiting) and 66 deaths were reported to the Ministry of Health (MOH) in Lima, Peru. Vibrio cholerae O1, Inaba, biotype El Tor, was isolated from patients' stools from Chancay and Chimbote. Additional cases have been reported from the cities of Chimbote, Piura, Trujillo, and Chiclayo. The MOH has recommended 1) the exclusive use of boiled water for drinking, 2) careful cleaning of fruit and vegetables, and 3) avoidance of raw or inadequately cooked fish or seafood.
This is the first time this century that epidemic cholera has been identified in South America. A pandemic of cholera began in 1961 in southeast Asia, and has affected many areas of Asia, the Middle East, Europe, Oceania, and Africa. The El Tor pandemic strain grows in many foods and can persist in aquatic environments. In previous epidemics, documented vehicles of transmission have included contaminated water, raw or undercooked seafood, moist-grain gruels, and leftover rice.
The risk to foreign travelers of acquiring cholera in endemic areas is low. Cholera vaccination confers only brief and incomplete protection and is not recommended. The usual precautions to prevent traveler's diarrhea should be observed carefully; particularly, raw seafood and potentially contaminated water should be avoided. A traveler who develops severe watery diarrhea, or diarrhea and vomiting, during or following travel to an area with known cholera should seek medical attention immediately. -- From Morbidity and Mortality Weekly Report, 1991, 40, 108-110.
Bubonic Plague Alert
In June, 1990, a mammologist from the United States who was traveling in the area of La Paz, Bolivia, had onset of an acute illness consisting of severe headache, chills, fever, sweating, loss of appetite, pain and swelling in her right axilla, and muscle pains in the lower back and hip. A physician diagnosed a respiratory infection and treated her with intramuscular ampicillin. The pain and swelling in the axilla gradually increased, and she had difficulty moving her right arm and shoulder. The next day, she traveled to her home in the District of Columbia; the pain in her right axilla had increased, and she had developed a dry cough. On examination by a physician, her temperature was 38.5 C (101.3 F), and she had an enlarged 2.4-cm fluctuant lymph node in the right axilla with surrounding boggy edema. She was immediately hospitalized with a presumptive diagnosis of bubonic plague.
A lymph node aspiration was performed, and Gram and Wayson stains of the aspirate revealed rare bipolar staining gram-negative organisms suggestive of Yersinia pestis. On June 22, a culture of the aspirate was positive for Y. pestis organisms; the finding was subsequently confirmed by the Plague Reference Diagnostic Laboratory in CDC's Division of Vector-Borne Infectious Diseases. The patient was treated with 1 g streptomycin intramuscularly twice daily. On the third day of treatment, the patient's fever defervesced, and the enlarged axillary node began to gradually regress. The patient was discharged on June 26 and completed a 10-day course of streptomycin as an outpatient. Following treatment, she has remained well.
The patient had camped in the vicinity of rural towns for nearly 2 months, collecting small mammals, including rice rats (Oryzomys sp.). Because of weight restrictions, she used nembutal injections rather than chloroform for euthanizing animals (nembutal kills the animals but not fleas on the animal; chloroform kills both), and while skinning rats she crushed some fleas with her fingers. She had received a primary plague immunization series in 1957 but had not received booster immunizations since 1971.
Human plague is endemic and occasionally epidemic in South American, Africa, and Asia. In 1989, 770 cases were reported from 11 countries (WHO, 1990), including 374 cases (49%) from Vietnam and 180 (23%) from Madagascar. Brazil reported 26 cases. Plague has been reported from Bolivia in 7 of the past 10 years (WHO, 1990), and the La Paz area in Bolivia is a recognized endemic focus (WHO, 1983).
Because plague vaccine boosters are necessary to maintain protective immunity, persons at continuing risk should receive booster doses at 1- to 2-year intervals (CDC, 1990). The primary series consists of three immunizations given on days 0 and 30 and 3-6 months after dose 2. The final dose of the primary series should be followed by a booster dose at 6 months and at 1 year. -- From Morbidity and Mortality Weekly Reports, 1991, 39, 895, 901.
Change of Dose for Malaria Prophylaxis
CDC recommends mefloquine (Lariam ®) alone as the drug of choice for malaria prevention for travelers to areas with drug-resistant Plasmodium falciparum malaria. Based on accumulating experience with this drug, the prophylactic dosing regimen has been revised to a single dose of mefloquine to be taken every week. The first dose should be taken 1 week before travel, and it should be continued weekly during the entire period of travel in malarious areas and for 4 weeks after departure from such areas. The previous dosing regimen (in which one dose was taken each week for 4 weeks, followed by one dose every other week) compromises the effectiveness of mefloquine.
All studies confirm that mefloquine is well tolerated when used for prophylaxis. No serious adverse reactions to mefloquine prophylaxis have been observed among Peace Corps volunteers or among 18,462 persons enrolled in prophylactic drug trials and surveys of travelers who were taking mefloquine weekly. However, serious adverse reactions have been reported, especially when mefloquine was used for treatment of patients with malaria. Because mefloquine has been used in the United States for only 18 months, monitoring of adverse reactions remains important. Physicians are encouraged to report serious adverse reactions in persons using mefloquine to CDC's Malaria Branch, Division of Parasitic Diseases, Center for Infectious Diseases [404-488-4046].
Consistent with previous guidelines, mefloquine is not recommended for use by travelers with known hypersensitivity to mefloquine; children less than 15 kg (30 lbs); pregnant women; travelers using beta blockers; travelers involved in tasks requiring fine coordination and spatial discrimination, such as airplane pilots; and travelers with histories of epilepsy or psychiatric disorder. Travelers to areas of risk where chloroquine-resistant P. falciparum is endemic and for whom mefloquine is contraindicated may elect to use daily doxycycline alone or chloroquine alone.
If chloroquine is used, the traveler needs to be aware of the need to seek medical attention for febrile episodes and to carry a treatment dose of pyrimethamine-sulfadoxine (Fansidar ®) to be used if medical care is not available within 24 hours.
The CDC publication Health Information for International Travel (CDC-90-8280) provides detailed information for each country on the risk for malaria to travelers and on the presence of drug-resistant P. falciparum.
Health information for travelers is available 24 hours a day from the CDC automated telephone system [404-332-4555]. Periodic shortages of mefloquine have occurred in the United States. Travelers who cannot obtain mefloquine before departure may be able to purchase it in Europe while in transit to countries with endemic malaria. Prescriptions written in the United States are accepted at airport pharmacies in Frankfurt and Paris (both Charles deGaulle and Orly airports). The pharmacy at Heathrow Airport in London requires prescriptions written in Great Britain. At the airport pharmacy in Brussels, a prescription from the airport physician is required. Mefloquine is not available at the airports in Amsterdam and Rome and at Gatwick (London).
In some countries, a fixed combination of mefloquine and pyrimethamine-sulfadoxine is marketed under the name Fansimef®. Fansimef® should not be confused with mefloquine, and it is not recommended for prophylaxis of malaria. -- From Morbidity and Mortality Weekly Reports, 1991, 40, 72-73.
Dengue Epidemic, 1990
From March to July, 1990, an epidemic of dengue caused by dengue types 1 and 4 occurred in Iquitos, Peru, and the surrounding area. A smaller outbreak was reported in Tarapoto in the neighboring department. Predominant manifestations were fever, headache, and malaise. Five of 20 pools of mosquitoes collected near Iquitos yielded dengue virus. Aedes aegypti, the epidemic vector of dengue, was declared eradicated from Peru in 1958, but has been gradually reinfesting the country. From Morbidity and Mortality Weekly Reports, 1991, 40, 145-147.
* * *
IPS in '92
The International Primatological Society announces that its 14th Congress will take place in Strasbourg, France, August 16 to 21, 1992, at the Congress Palast. The host institution is the Societe Francophone de Primatologie. The official language of the congress will be English with an eventual translation to and from French. Symposia on evolution, ecology, social behavior, physiology, nutrition, genetics, and communication have been suggested. The Scientific Committee would be grateful for further suggestions, and for early indications of intentions to participate in the congress and to contribute to the program.
For further information, contact N. Herrenschmidt, Univ. Louis Pasteur, Centre de Primatologie, 7, rue de l'Universite, F-67000 Strasbourg, France [33-8856-12-68; FAX: 33-8856-02-30]. Tentative locations for future meetings are Madagascar in 1994 and Indonesia in 1996.
Cal State Fullerton
The Anthropology Department and the Anthropology Students Association of California State University, Fullerton, have scheduled a Primate Symposium for April 20, 1991. The topic will be the Great Apes, with Sandy Harcourt, Kelly Stewart, James Moore, Richard Tenaza, and Gary Shapiro speaking. For more information, contact Norm Rosen, Anthropology Department, Cal State Fullerton, Fullerton, CA 92631 [213-473-9641].
Royal Society Discussion Meeting
Drs. E. Widdowson and A. Whiten have organized a discussion of Foraging Strategies and Natural Diet of Monkeys, Apes and Humans for May 30, 1991, in London. For further information, write The Royal Society, 6, Carlton Terrace, London SW1Y 5AG, England.
IV World Veterinary Congress
The IV World Veterinary Congress will be held in Rio de Janeiro, August 18 to 23, 1991. For information contact the Congress Secretariat, Congrex Do Brasil, Rua do Ouvidor 60 Grupo 414, 20040 Rio de Janeiro RJ Brasil [021-224-6080; FAX: 021-231-1492]. The World Association of Wildlife Veterinarians will also hold their meeting at this Congress. Contact J. E. Cooper FRCVS, Royal College of Surgeons of England, 35-43 Lincoln's Inn Fields, London WC2A 3PN, England.
AALAS and ICLAS
The American Association for Laboratory Animal Science and the International Council for Laboratory Animal Science will hold a joint meeting in Buffalo, NY, October 20 to 24, 1991. For information, contact AALAS, 70 Timber Creek Drive, Suite 5, Cordova, TN 38018.
Travel to ASP
Videk Enterprises, run by Dick and Viciki Dukelow, are arranging special roundtrip airfares to the American Society of Primatologists meeting in Veracruz, Mexico, this June 24-28. Fares are available from any Continental Airlines "Gateway" city in the US. For booking and further information, call Joann (of Getaway Travel 'n' Tours) at 800-748-0435, or contact Videk Enterprises, Box 233, Holt, MI 48842 [517-694-8454].
Breitenbach Travel, 3594 N. Oakland Ave., Milwaukee, WI 53211 [800-366-3828] is also mentioned as being on the alert to take care of ASP attendees. Dr. A. Estrada, Local Arrangements Chair for the meeting, is available to answer questions about travel and accomodations, as well as about a post-Congress field trip to Lake Catemaco and Los Tuxtlas [FAX: 011 52 294-30115]
NABR Conference
The National Association for Biomedical Research will hold a Conference on Government Relations, May 19-21, 1991, at the Capitol Hilton in Washington, DC. Representative Vin Weber (R-MN), founder of the Congressional Animal Welfare Caucus, and other Members of Congress and their staff have been invited to participate in discussions. Conference participants will also travel to Capitol Hill to ask for support for the Caucus and the federal facility protection bills, which Representative Stenholm (D-TX) and Senator Heflin (D-AL) are expected to introduce in the near future. For more information, contact NABR, 818 Connecticut Ave., N.W., Suite 303, Washington, DC 20006 [202-857-0540; FAX: 202-659-1902].
* * *
Eluned C. Price and Sian Evans
Jersey Wildlife Preservation Trust and the University of Miami
The use of contraceptive implants to control breeding is now an important component in the captive management of two of the most attractive and most endangered callitrichid primates, the golden lion tamarin (Leontopithecus rosalia) and the cotton-top tamarin (Saguinus oedipus).
The captive population of golden lion tamarins, some 500-600 animals, is tightly managed, and implants containing the hormone melengestrol acetate have now been placed in a number of females in zoos and research colonies (Ballou & Kleiman, 1989).
The captive population of cotton-top tamarins, in contrast, is much larger (probably around 1500 individuals; Tardif & Colley, 1988), and is not now subject to such systematic management. This species can be bred very successfully in captivity (e.g., Price & McGrew, 1990), and as a result the space available for cotton-tops is reaching its limit. Several United Kingdom institutions holding cotton-tops have decided to follow the example of the golden lion tamarin program and limit breeding by implanting females. However, we have become aware of several problems that have arisen in the use of implants in cotton-top tamarins in the UK. These include the removal of sutures by other family members, necessitating prolonged separation of females from their families to allow the wound to heal (E. C. Price, personal observation); migration and eventual expulsion of an implant through the original incision (E. C. Price, personal observation); and abortions in females implanted during pregnancy (J. B. Carroll, personal communication).
Improved techniques and the wider dissemination of information may well reduce the occurence of such difficulties, and at this point it seems useful to attempt to compile a data base, so that potential users of implants are aware of possible problems, and of solutions developed by others. We therefore request details of experiences from anyone who has used implants in cotton-top tamarins, or in other species. We will collate and publish the results, with full acknowledgement of all contributions. We are particularly interested in details of techniques used for putting implants in place (including anesthesia, positioning of the implant, and the most appropriate size for implants in a given species); any difficulties that occurred during or after implantation (such as insertion of the implant, migration or expulsion of implants, removal of sutures, or problems with healing); abortions, deaths, or other health problems; and any disruptions of family relationships.
Please send the information to either of the following addresses: *Dr. Sian Evans, Dept. of Biology, Univ. of Miami, P.O. Box 249118, Coral Gables, Miami, FL 33124, USA. *Dr. Eluned Price, Jersey Wildlife Preservation Trust, Les Augres Manor, Trinity, Jersey, JE3 5BF, Channel Islands, GB.
References
Ballou, J. & Kleiman, D. G. (1989). Species survival plan report: Golden lion tamarin. AAZPA Newsletter, 30[2], 6-7.
Price, E. C. & McGrew, W. C. (1990). Cotton-top tamarins (Saguinus (o.) oedipus) in a semi-naturalistic captive colony. American Journal of Primatology, 20, 1-12.
Tardif, S. D. & Colley, R. (1988). International Cotton-top Tamarin Studbook. Oak Ridge: Oak Ridge Associated Universities.
* * *
The Armed Forces Institute of Pathology (AFIP) and the American Registry of Pathology are co-sponsoring a continuing education course entitled "Pathology of Laboratory Animals," August 12-16, 1991, at the Holiday Inn, 8120 Wisconsin Ave., Bethesda, MD 20814.
The course is designed primarily for professionals who are responsible for the recognition and interpretation of lesions in laboratory animals. It is intended particularly to help them interpret spontaneous disease which may affect experimental results or alter the supply of suitable lab animals. Pathology will receive major emphasis, but other features of diseases such as etiology, pathogenesis, diagnosis, and control will also be given attention. A wide range of diseases from iatrogenic through infectious will be discussed in a variety of lab animal species.
Application forms and programs for the course may be obtained by contacting the American Registry of Pathology, AFIP, Washington, DC 20306-6000 [202-576-4549]. Tuition fee is $250, but Department of Defense, Veterans Administration, and Public Health Service applicants will only be assessed an administrative fee of $90.
* * *
Primate Foundation of Arizona
The Primate Foundation has three job openings. One is for a staff veterinarian as listed in the last issue of this Newsletter. A second is for a registrar to maintain and develop computer record systems to account for inventory and health status of a chimpanzee breeding colony, and various other office duties. Some computer experience is desirable, and the applicant must type a minimum of 40 WPM and have good communication skills.
The third position is for a research associate, requiring a Ph.D. (thesis pending considered) in psychology, zoology, or anthropology, and experience in behavioral research. This person will work directly under the Director of the colony, and be responsible for assistance with study design, analysis, and write-up, as well as training and daily supervision of the Staff Observers and Interns, maintenance of daily records, and collection and computer entry of behavioral data. The job requires a solid background in statistics, good organization, managerial and communication skills, and the ability to work well with students and other investigators.
All personnel must have a negative T.B. skin test and a negative Hepatitis B surface antigen test prior to employment. We are an Equal Opportunity Employer and offer excellent benefits. All salaries are negotiable. Send a letter of interest (with requested salary), resume, and three letters of reference to: Jo Fritz, Director, Primate Foundation of Arizona, P.O. Box 86, Tempe, AZ 85280.
* * *
USDA Standards Finalized
The last subparts of animal welfare regulations and standards necessary as a result of 1985 amendments to the federal Animal Welfare Act were published in final form on February 15 by the Animal and Plant Health Inspection Service (APHIS) of the USDA. New standards for dogs, cats, and nonhuman primates (9 CFR Part 3, Subparts A and D) appeared in the February 15, 1991, Federal Register (56 Fed Reg 6426-6505). APHIS administrator James W. Glosser told the AP wire service the agency had produced "a practical and enforceable blend of performance and engineering standards." He said, "We have made room for variations in the way individual animals behave and for differences in the way they are used and housed."
This final version differs very little in substance from the reproposal which appeared on August 15, 1990. While the new standards are effective as of March 18, 1991, research facilities will be given time to put their plans in place. Written documents describing programs for environmental enrichment for nonhuman primates must be developed and implemented by August 14, 1991. Such written plans and procedures must then be available to APHIS inspectors. In order to minimize their economic impact, major capital improvements, such as changes in cage size, may be phased in over three years. Compliance with six specific provisions which may require significant investment by some institutions can be delayed until February 15, 1994. See the "effective date" discussion on page 6431 of the Federal Register for complete details.
Page 6369 of the same issue of the Federal Register contains a joint announcement by the USDA's National Agricultural Library (NAL) and the NIH. The NAL, in cooperation with the National Library of Medicine, is preparing and will continue to update bibliographies pertaining to the psychological well-being of nonhuman primates. Such bibliographies, the NIH "Nonhuman Primate Intramural Management Plan," and model institutional plans, once they are developed, will be available to requestors from the NAL. A series of regional meetings (San Francisco, April 10; St. Louis, May 1; Washington, DC, June 6) will be cosponsored by APHIS and the NIH Office for Protection from Research Risks Division of Animal Welfare, to assist institutions in interpreting the new standards and solicit comments of interested parties. -- From the February 15, 1991 issue of NABR Alert, XIII[2].
* * *
Wildlife Preservation Trust
The Wildlife Preservation Trust, an international non-profit organization, develops and maintains captive breeding programs worldwide, as well as conducting research, educational programs, and professional training. Their Training Program is a 10 or 16 week internship designed for postgraduates and zoo personnel to give intensive training in propagation techniques with a variety of species. For application and further information, contact Training Program, Wildlife Preservation Trust International, 34th St. & Girard Ave., Philadelphia, PA 19104 [215-222-3636]. The Summer Course is designed for advanced undergraduates and graduate students. For details, write to John McNicholas, Assistant Training Officer, Summer School Coordinator, Jersey Wildlife Preservation Trust, Les Augres Manor, Trinity, Jersey, Channel Islands, Britain.
Earthwatch
Earthwatch provides volunteers and funds to scholars doing fieldwork. Among the projects and scholars listed in their 1991 catalog are studies of rain forest fauna in Mexico (A. Estrada), macaques in Bali (B. Wheatley), bush fauna in Zimbabwe (S. Alibhai), and ecology of Lake Naivasha (D. Harper & K. Mavuti). Volunteers, who pay their own transportation expenses plus a share of the costs of the expedition, learn to participate in fieldwork. Interested scientists should submit a two-page outline of their research subject and design, dates, budget, and plans for the use of volunteers, to the Center for Field Research 680 Mount Auburn St., Box 403, Watertown, MA 02272 [617-926-8200]. Volunteers should apply to the same address.
AALAS Certification Courses
The New York Metropolitan Branch of the American Association for Laboratory Animal Science offers self-testing exams and course materials to instructors designing course plans or to technicians preparing for AALAS technologist certification. Registration can be at any time. Contact Dr. D. M. Stark, Box 23A, 500 E. 63rd St., New York, NY 10021.
New World Monkeys for Research
The Interagency Research Animal Committee, chartered by the U.S. Public Health Service (PHS), supports and coordinates a program with the Pan American Health Organization that makes possible the importation of New World monkeys into the United States for use in biomedical research projects. Transfers are limited to a few species, neither threatened nor endangered. For information on qualifications for participation, species, purchase procedures, and other matters, contact the Interagency Research Animal Committee, Ms. Suzanne Moore, NIH, Bldg 14A, Rm 100, 9000 Rockville Pike, Bethesda, MD 20892 [301-496-5424].
* * *
Bonobo Network
An informal newsletter for the exchange of information on the topics of pygmy chimpanzee/bonobo research, conservation, and captive management has been started. It is an outgrowth of a round-table discussion on the bonobo, which was held at the IPS meeting in Nagoya last July. To contribute to or receive this newsletter, contact Evelyn Ono Vineberg, 10603 Sunset Ridge Dr., San Diego, CA 92131 [619-549-8906; FAX: 619-566-1230].
PIC 1991 Fee Schedule and Catalog
The Primate Information Center has put out its 1991 Fee Schedule and catalog of Topical Bibliographies in print. For a copy, contact PIC, RPRC, SJ-50, Univ. of Washington, Seattle, WA 98195 [206-543-4376; FAX: 206-685-0305].
Reading and Reference Lists
Cathy A. Johnson-Delaney, of the Primate Information Center, has prepared several reference lists, which should be useful to many readers. The lists are: Primate References for Young Readers 1919 (Ele- mentary school); Primate References for Students 1991 (Middle and High School); General Interest Primate Reading List 1991; Pet Own- er's List of Primate Societies, Sanctuaries, and Foundations; and Reference List for Practitioners, which includes a resource list and an article on health care and husbandry presented by Dr. Johnson-Delaney at the Eastern States Veterinary Conference in January, 1990. For copies of any of these lists, contact Dr. Johnson-Delaney at the PIC (address given above).
Specimen Resources Directory
The Registry of Comparative Pathology of the Armed Forces Institute of Pathology has published the second edition of the directory, Resources of Biomedical and Zoological Specimens. The first edition was published in 1981. The new, 47 page directory contains 99 listings of collections all over the world, and is available free from the Registry of Comparative Pathology, Armed Forces Inst. of Pathology, Washington, DC 20306 [202-576-2452].
Orangutan Project
Dr. Joe Kemnitz's project on the relationship between obesity and diabetes in the orangutan continues. Any institution that houses a diabetic orangutan and that has not already participated should contact Dr. Anne Baker at the Brookfield (Chicago) Zoo [708-485-0263], as should any other instutitions that would like to provide baseline information from nondiabetic animals. A description of the project and its protocol are available from either Dr. Baker or Lori Perkins at Zoo Atlanta [404-624-5600]. -- From the AAZPA Communique, September 1990.
NSF Outreach Service
NSF's Outreach Service encourages NSF staff members on official business to give orientations about NSF funding opportunities at all institutions, especially those that are not among the leading recipients of NSF funds. These include predominantly minority or women's colleges and universities and primarily undergraduate institutions.
The Outreach Service will respond to written requests for NSF speakers by attempting to arrange, where possible, for travelling staff to visit the requesting institution. To avoid duplication, requests should be made through a research or similar office of the requesting institution. Because of limited availability of staff time and travel funds, NSF is not able to respond to all requests. Priority will be given to requests for presentations and visits that would serve several institutions at the same time.
Interested persons should contact Mr. Patrick Olmert, NSF Outreach Coordinator, States Liaison Section, NSF Office of Legislative and Public Affairs, Washington, DC 20550 [202-357-7196].
WRPRC Library Announcements
The Wisconsin RPRC has announced a schedule of fees for the loan of audiovisual material to persons outside the University at Madison, to offset shipping and packing costs. Videotapes, films, slide sets, or audiotapes cost $5 per item, plus $5 per order. Slides cost 50 cents each, plus $3 per order. Non-U.S. borrowers' fees will vary depending on postage or shipping charge. Contributors of substantive audiovisual materials will receive free borrowing privileges for one year.
The Library has also produced the first two of a series of audiovisual bibliographies, one on chimpanzees (1929-1990; Publ. No. 002-90) and one on gorillas (1938-1990; Publ. No. 003-90). They contain annotated citations of videotapes, films, records, etc, including items not held by the WRPRC Library, and are available for purchase at $10 each. The second edition of their comprehensive catalog of audiovisual holdings (Publ. No. 001-90) is also now available for $10 ($21.50 outside the U.S.). Send check payable to: Primate Center Library, WRPRC, 1220 Capitol Court, Madison, WI 53715-1299 [608-263-3512; FAX: 608-263-4031].
Prenatal Drug Exposure
Mary F. Dent, Dept. of Family Medicine, 4N51 Brody, ECU School of Medicine, Greenville, NC 27858-4354, would like to know if anyone is studying prenatal exposure to cocaine or sympathomimetics in non-human primates. She is currently preparing her own study using standard rats and believes some of the puzzles in this area could be cleared up with a good primate study. She would like to communicate with anyone addressing this area. She can also be reached by e-mail at [email protected]
Environmental Enrichment Ideas
Dorothy Fragaszy, editor of the IPS Newsletter, has started a column of suggestions for practical environmental enrichment in that newsletter. Send your ideas to her at the Psychology Dept., Univ. of Georgia, Athens, GA 30602.
Animal Research Information Board
The Animal Research Information Board, available only through BITNET or INTERNET, will have information and items of interest on the use of animals in research. It will include the latest information on:
* * *
Books
*The Chimpanzees of the Mahale Mountains: Sexual and Life History
Strategies. T. Nishida (Ed.). Tokyo: Univ. of Tokyo Press, 1990. 328
pp. [Price: $69.50. Distributed by Columbia Univ. Press.]
. .
J. Itani and his associates report on 25 years of demographic and
behavioral research on certain chimpanzee bands in Tanzania. The book
includes photographs and many tables of data. Contents: A quarter century of research in the Mahale Mountains: An overview, by T. Nishida.
Safari surveys of the vegetation and the chimpanzee groups in the
northern half of the Mahale Mountains, by J. Itani. Demography and
reproductive profiles, by T. Nishida, H. Takasaki, & Y. Takahata. Sex
differences in ranging patterns, by T. Hasegawa. Sperm competition
and mating behavior, by T. Hasegawa & M. Hiraiwa-Hasegawa. Adult
males' social relations with adult females, by Y. Takahata. Social
relationships among adult males, by Y. Takahata. Alpha males' interactions and social skills, by K. Kawanaka. Social context of pantgrunting in young chimpanzees, by H. Hayaki. Age differences in anteating by adult and adolescent males, by K. Kawanaka. A preliminary
report on age differences in plant-feeding behaviors among adult and
adolescent males, by S. Uehara. Some socio-behavioral manifestations
of old age, by M. A. Huffman. Maternal investment before weaning, by
M. Hiraiwa-Hasegawa. Role of food-sharing between mother and infant
in the ontogeny of feeding behavior, by M. Hiraiwa-Hasegawa. A note
on the ontogeny of feeding, by M. Hiraiwa-Hasegawa. Deceptive behavior in young chimpanzees: An essay, by T. Nishida.
*Rearing Condition and the Acquisition of Phobic Behaviour in Monkeys. E. L. Roder. Nijmegen: Doctoral dissertation, 1990. 109 pp.
*Biomedical Research in Primates. M. Jonker, Ed. Rijswijk: TNO Primate Center, 1988. 96 pp. [Price: $25]
. .
Proceedings of the TNO Symposium on
Biomedical Research in Primates,
Amsterdam, March 10-11, 1988. Contents: Opening speech, by J.
van London. TNO Primate Center: Past, present, and future, by C.
Pries. Breeding and maintenance of non-human primates, by C. Goosen.
Primates and biotherapy, by H. Schellekens. Hepatitis delta infection
in chimpanzees, by A. Kos & H. Schellekens. The primate role of the
MHC in transplantation, disease and health, by R. R. P. de Vries.
Experimental allergic encephalomyelitis in non-human primates: An
excellent model of multiple sclerosis, by E. C. Alvord, Jr., L. M.
Rose, S. Hruby, T. L. Richards, R. Petersen, C.-M. Shaw, E. A. Clark,
L. H. Ericsson, W. A. Stewart, D. W. Paty, & M. W. Kies. The use of a
primate model in the preclinical testing of monoclonal antibodies, by
P. J. Friend, G. Hale, & H. Waldmann. The use of non-human primates
for the evaluation of efficacy and safety of therapeutic monoclonal
antibodies, by M. Jonker. Ethological studies on social deprivation in
monkeys, apes and humans, by H. Dienske. Studies of disturbed behaviour in macaques, by C. Goosen. Ethics in research using non-human
primates, by Tj. De Cock Buning. The living environment of non-human
primates in captivity, by J. Goodall.
Bibliographies
* Tuberculosis in Nonhuman Primates 1982-1990: A Bibliography. 2nd edition. C. A. Johnson-Delaney. Seattle: Primate Information Center, 1990. 8 pp. (79 citations) [Price: $6.50. Stock #90-011. Send order to Primate Information Center, RPRC, SJ-50, Univ. of Washington, Seattle, WA 98195]
*Simian and Human Retroviruses in Nonhuman Primates: Infection Disease and Animal Model Studies: A Bibliography. Annual update. C. A. Johnson-Delaney. Seattle: Primate Information Center, 1990. 35 pp. (336 citations) [Price: $10. Stock #90-012. Ordering information as above]
*Primate Field Studies in Brazil: A Bibliography. A. T. Bernardes, A. B. Rylands, C. M. C. Valle, R. B. Machado, A. F. Coimbra-Filho, & L. R. B. Fischer. 1988. 114 pp. [Belo Horizonte, Minas Gerais, Brazil: Soc. Brasileira de Primatologia]
Directories
*Resources of Biomedical and Zoological Specimens. Second edition.
Washington, DC: RCP, 1990. 47 pp. [Free from the Registry of Comparative Pathology, Armed Forces Inst. of Pathology, Washington, DC 20306
[202-576-2452].
. .
Contains 99 listings of collections all over the world.
New Journals
* Live Animal Trade & Transport Magazine [Published quarterly;
$20/year. P.O. Box 441110, Fort Washington, MD 20749-0653].
. .
Starting its third year, this publication has departments on regulations, animal welfare, health, services, and equipment.
Magazines, Newsletters, and Reports
*Primate News, 1990, Volume 24. [Published by the Oregon RPRC, 505
N.W. 185th Ave., Beaverton, OR 97006.]
. .
A special report issue, "Toward Better Health,"
on the role of primates in biomedical research at the Regional Primate Research Centers.
*Primate Report, No. 28, June, 1990. [Published in cooperation with
the German Primate Center (DPZ). Price $8]
. .
The 1989 Annual Scientific Report of the DPZ includes the following
articles: Inclusion body hepatitis in infant tamarins (Saguinus oedipus, S. fuscicollis, S. labiatus), of the German Primate Center
(DPZ), by M. Brack, G. Baskin, & H. Klensang. Urinary neopterin and
biopterin in various primate species, by M. Goldberg & E. Fuchs. Does
the duration of living with the breeding pair influence the first
reproductive success in Saguinus fuscicollis daughters? by I. Kuderling.
*Inside Yerkes, Winter, 1991. [Yerkes RPRC, Emory Univ., Atlanta, GA
30322]
. .
Reports on current research at the Center, and a review of its 60
year history.
Special Journal Issues
*Captive propagation and reintroduction: A strategy for preserving
endangered species? Endangered Species UPDATE, 1990, 8[1].
. .
Contains general articles and case histories of 17 species, including one primate. Of particular interest to primatologists are: The
role of genetics in captive breeding and reintroduction for species
conservation, by A. R. Templeton; and The conservation program for the
golden lion tamarin, Leontopithecus rosalia, by D. G. Kleiman, B. B.
Beck, A. J. Baker, J. D. Ballou, L. A. Dietz, & J. M. Dietz.
*Primatology in China. Primate Report, No. 26, March, 1990. [Published in cooperation with the German Primate Center (DPZ). Price $8]
. .
Fifteen articles on fossil and living primates in China, on primatological work at Chinese institutes, on the main research fields, and an invitation for intensive international cooperation.
*Tactical deception in primates: The 1990 database. Primate Report,
No. 27, May, 1990. [Publishing information and price same as above.]
. .
R. W. Byrne and A. Whiten present 253 reports by primatologists of
incidents which suggest deception.
Anatomy
*Morphologic development of the adrenal cortex in squirrel monkeys
(Saimiri sciureus). Miller, C. J., Prahalada, S., Coe, C., &
Benirschke, K. (CPRC, Univ. of California, Davis, CA 95616). Journal
of Medical Primatology, 1990, 19, 651-661.
. .
Adrenals of fetal monkeys had a broad central zone of large eosinophilic cells (fetal zone) surrounded by a subcapsular band of small
dark cells (definitive zone). Adrenals of stillborn and neonatal monkeys had a reduced fetal zone and expanded definitive zone. By day 18
postpartum, no remnant of the fetal zone remained.
Animal Models
*Serial transmission of a putative causative virus of enterically
transmitted non-A, non-B hepatitis to Macaca fascicularis and
Macaca mulatta. Uchida, T., Win, K. M., Suzuki, K., Komatsu, K.,
Iida, F., Shikata, T., Rikihisa, T., Mizuno, K., Soe, S., Myint, H.,
Tin, K. M., & Nakane, K. (Dept. of Pathology, Nihon Univ. School of
Medicine, Oyaguchi-Kamimachi, Itabashi-ku, Tokyo 173, Japan). Japanese Journal of Experimental Medicine, 1990, 60, 13-21.
. .
Fecal extract from patients with NANB (called type E) hepatitis
caused acute hepatitis in primates, in first, second, and third passage.
*Enterically-transmitted non-A, non-B hepatitis. Bradley, D. W. (Hepatitis Branch, CDC, Atlanta, GA 30333). British Medical Bulletin, 1990, 46, 442-461.
. .
Description of experimental studies with cynomolgus, African green,
and rhesus monkeys, tamarins, and chimpanzees.
*Effects of captopril on atherosclerosis in cynomolgus monkeys.
Aberg, G. & Ferrer, P. (Dept. of Pharmacology, Squibb Inst. for Medical Research, P.O. Box 4000, Princeton, NJ 08543-4000). Journal of
Cardiovascular Pharmacology, 1990, 15 (Suppl 5), S65-S72.
. .
Monkeys given captopril showed a significantly reduced progression
of arterial lesions; the effects were most evident in the coronary
arteries, which were practically free from atherosclerosis in treated
animals.
*Amyloid in the brains of aged squirrel monkeys. Walker, L. C., Masters, C., Beyreuther, K., & Price, D. L. (Neuropathology Lab., Johns
Hopkins Univ. School of Medicine, 600 N. Wolfe St., 509 Pathology
Bldg., Baltimore, MD 21205-2181). Acta Neuropathologica, 1990,
30, 381-387.
. .
The squirrel monkey may be a useful model for the study of the
pathogenesis of amyloid in the aged brain. Unlike rhesus monkeys,
squirrel monkeys show a preponderance of amyloid deposition in the
walls of blood vessels rather than in the cores of senile plaques.
This species difference could be exploited to determine the mechanisms
by which amyloid forms within the vascular wall and the relationship,
if any, between vascular amyloidosis and senile plaques in the aged
brain.
Behavior
*Do old female monkeys have a specific social role? Pavelka, M. S. M.
(Dept. of Anthropology, Univ. of Calgary, 2500 University Dr. NW, Calgary, Alberta T2N 1N4 Canada). Primates, 1990, 31, 363-373.
. .
The expectation that old monkeys will occupy a distinct social role
because of their advanced age is discussed in terms of biological vs.
social aging, human vs. nonhuman aging, the evolution of behavioral
change in old age, and the application of the social role concept to
old female monkeys. Data from 450 hours of observation of 40 Japanese
monkeys gives little evidence that the behavior of the aged females
differs significantly from that of other adult females.
*Proboscis monkey (Nasalis larvatus) social organization: Intergroup patterns of association. Yeager, C. P. (Psychology Dept., Univ. of Tennassee, Knoxville, TN 37996). American Journal of Primatology, 1991, 23, 73-86.
. .
Data on 10 one-male groups and two all-male groups show a secondary
level of organization, "bands,"
distinct from the groups. Adult
females direct the majority of their affiliative behavior towards
their offspring and other adult females, males do not engage in active
herding, and one-male groups spend time apart from other groups.
*Effects of the presence of a second male on pair-tests of captive
cynomolgus monkeys (Macaca fascicularis): Role of dominance. Zumpe,
D. & Michael, R. P. (R. P. M., Dept. of Psychiatry, Emory Univ.
School of Medicine, GMHI, 1256 Briarcliff Rd., Atlanta, GA 30306).
American Journal of Primatology, 1990, 22, 145-158.
. .
Stimulation of a test male by a second male either immediately
before or during a pair-test had highly significant but different
effects. Ejaculatory activity increased in tests that followed stimulation, mainly in males dominant to the stimulus male, while sexual
and social activity decreased in tests during stimulations, but mainly
in males subordinate to the stimulus male. This suggests that dominance confers a mating advantage on males even when there is no oppor-unity for mating interference or active mate competition.
*Consistency in maternal behavior within families of free-ranging rhesus monkeys: An extension of the concept of maternal style. Berman, C. M. (Dept. of Anthropology, SUNY, Buffalo, NY 14261). American Journal of Primatology, 1990, 22, 159-169.
. .
Comparing several measures of mother-infant interaction between
pairs with the same mothers, and pairs with different mothers, as well
as over time, suggest that the concept of "maternal style" can be used
to describe variation between both individual mother-infant pairs and
between individual mothers.
*Evidence for a "monitoring call" in red-bellied tamarins. Caine, N.
G. & Stevens, C. (Dept. of Psychology, Bucknell Univ., Lewisburg, PA
17837). American Journal of Primatology, 1990, 22, 251-262.
. .
Three hypotheses about the "slide call" of red-bellied tamarins
were tested; all were supported, implying that social monitoring
appears to be at least one important function of this call, especially
in circumstances where coordinated and/or cohesive behavior is
required of the group.
*Variability in spider monkeys' vocalizations may provide basis for
individual recognition. Chapman, C. A. & Weary, D. M. (D. M. W., Dept.
of Biology, McGill Univ., Montreal, P.Q. H3A 1B1 Canada). American
Journal of Primatology, 1990, 22, 279-284.
. .
Analysis of long-range vocalizations given by spider monkeys
revealed consistent acoustic differences among the calls of individuals. Acoustic recognition of callers would facilitate the choice of
which subgroups to join, allowing individuals to manipulate the size
and composition of their subgroups.
*Sex differences in mortality among langurs (Presbytis entellus) of
Jodhpur, Rajasthan. Rajpurohit, L. S. & Sommer, V. (V. S., Inst. für
Anthropologie, Burgerstr. 50, D-3400, Gottingen, Germany). Folia
Primatologica, 1991, 56, 17-27.
. .
Long-term demographic monitoring of 144 known individuals up to age
30 months confirmed a higher juvenile female mortality. Once males
emigrate from their natal troop, their mortality begins to exceed that
of females. Male infants are more subject to infanticide.
*Object manipulation in wild red colobus monkeys living in the Abuko
Nature Reserve, The Gambia. Starin, E. D. (Dept. of Anthropology, Graduate Center, CUNY, 33 W. 42nd St., New York, NY 10021). Primates, 1990, 31, 385-391.
. .
Variation in the form and frequency of manipulative activities
shows clear sex differences which may relate to sex differences in social behavior.
*A comparative study of tolerance and social organization in captive
lemurs. Blanckenhorn, W. U. (Dept. of Biological Sciences, SUNY,
Albany, NY 12222). Folia Primatologica, 1990, 55, 133-141.
. .
A troop of Varecia variegata showed higher levels of agonistic
behavior and lower levels of grooming behavior than similar troops of
Lemur fulvus and L. catta. This supports the few indications that
wild V. variegata occur in smaller, perhaps family or extended family groups, as well as the prediction that species forming larger multimale associations, like L. catta and L. fulvus, have evolved higher degrees of social tolerance.
*Species preference by infant macaques with controlled social experi-
ence. Fujita, K. (Dept. of Psychology, Primate Research Inst., Kyoto Univ., Inuyama, Aichi, 484, Japan). International Journal of Primatology, 1990, 11, 553-573.
. .
Japanese monkeys and rhesus monkeys, reared with various experience
with their own and the other species, were offered levers to press to
observe pictures of the two species. Results suggest that infants of
both species have a native tendency to prefer to see physical characteristics of rhesus monkeys over Japanese monkeys.
*Delayed figure reconstruction by a chimpanzee (Pan troglodytes) and
humans (Homo sapiens). Fujita, K. & Matsuzawa, T. (Address same as
above). Journal of Comparative Psychology, 1990, 104, 345-351.
. .
A chimpanzee showed excellent visual reproductive memory, compared
to humans, but there appeared to be differences in some aspects of
information processing.
*Do rhesus mothers suggest friends to their offspring? de Waal, F. B.
M. (WRPRC, 1223 Capitol Court, Madison, WI 53715-1299). Primates,
1990, 31, 597-600.
. .
23 rhesus mothers were observed 240 times holding their own infants
and the infant of another female. In about 90% of cases, the second
infant was from a dominant matriline. It is suggested that females try
to promote future association between their own offspring and high-ranking youngsters.
*Spontaneous tool use and sensorimotor intelligence in Cebus compared with other monkeys and apes. Chevalier-Skolnikoff, S. (CPRC, Univ. of California, Davis, CA 95616). Behavioral and Brain Sciences, 1989, 12, 561-627.
. .
62 hours of observing captive groups, plus a literature survey,
suggest that strong tool-using propensity among primates is based on
advanced sensorimotor ability rather than fortuitous discovery. 23
pages of comments by 31 peers are followed by 11 pages of response by
the author.
*Natal male baboon rank rises and successful challenges to resident
alpha males. Hamilton, W. J. III & Bulger, J. B. (Div. of Environmental Studies, Univ. of California, Davis, CA 95616). Behavioral Ecology and Sociobiology, 1990, 26, 357-362.
. .
Male baboons gain status slowly within their natal troop, but
quickly after they emigrate. Observations of over 120 animals for 4
years showed the departing rank of natal emigrants to be 5.4, while
the first measured rank, shortly after transfer into a new troop, was
1.5.
Care
*Auditory enrichment for Lar gibbons Hylobates lar at London Zoo.
Shepherdson, D., Bemment, N., Carman, M., & Reynolds, S. (Zoological
Soc. of London, Regent's Park, London NW1 4RY, England). International Zoo Yearbook, 1989, 28, 256-260.
. .
A pair of Lar gibbons responded to playing of Lar gibbon vocalizations with increased activity and vocalization, and this response was sustained over a period of several months.
*A mealworm dispenser for the slender-tailed meerkat Suricata suricatta at London Zoo. Shepherdson, D., Brownback, T., & James, A. (Address same as above). International Zoo Yearbook, 1989, 28, 268-271.
. .
A 50 cm length of 5 cm diameter plexiglas tubing, with the open
ends covered with removeable caps, and ten 6 mm holes drilled along
one side, was filled with a mixture of 25-30 mealworms and sawdust and
fixed to the roof of the animals' enclosure. As the worms moved about,
they would eventually fall out, taking about 8 hours to "dispense" themselves.
*The application and evaluation of behavioural enrichment in zoos.
Shepherdson, D. J. (Address same as above). Primate Report, 1988,
22, 35-41.
. .
Descriptions are included of artificial "foraging" devices for
chimpanzees and marmosets.
*The reduction of abnormal behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. Bayne, K., Mainzer, H., Dexter, S., Campbell, G., Yamada, F., & Suomi, S. (Bldg. 14D, Rm. 313, NIH, 9000 Rockville Pike, Bethesda, MD 20892). American Journal of Primatology, 1991, 23, 23-35.
. .
A piece of plexiglass covered with artificial fleece, with food
particles rubbed into it, was attached to the outside of each animal's
home cage. All animals foraged from the board to the point that a significant reduction in the level of abnormal behavior was noted.
Conservation
*The endangered species act: A guide to its protections and implementation. [Price: $13.50, including shipping, from the Stanford Environmental Law Society, Stanford Law School, Stanford, CA 94305-8610.]
Development
*Teeth emergence in wild olive baboons in Kenya and formulation of a
dental schedule for aging wild baboon populations. Kahumbu, P. & Eley,
R. M. (R. M. E., Inst. of Primate Research, National Museums of Kenya,
Box 24481, Karen, Nairobi, Kenya). American Journal of Primatology,
1991, 23, 1-9.
. .
Based on analysis of 95 wild olive baboons plus data from other
researchers on wild and captive yellow and olive baboons, full permanent dentition in wild baboons is predicted to occur over 1 year later
than in captive animals.
*Behavioral responsiveness to strangers in young chimpanzees (Pan troglodytes). Miller, L. C., Bard, K. A., Juno, C. J., & Nadler, R. D. (Yerkes RPRC, Emory Univ., Atlanta, GA 30322). Folia Primatologica, 1990, 55, 142-155.
. .
Young chimpanzees were tested by exposure to a medium-sized, non-assertive stranger, and a large, assertive stranger, both in the presence of their caretaker and in a familiar setting. Their behavior is
described as wary, rather than fearful. Previous studies reporting
severe distress among strangers may be attributable to a strange setting and/or absence of the caretaker.
*Estimating the age of infant chacma baboons (Papio cynecephalus
ursinus). Whitehead, S. I., Henzi, S. P., & Piper, S. E. (Dept. of Zoology, Univ. of Natal, Pietermaritzburg, South Africa). Folia Primatologica, 1990, 55, 185-188.
. .
Muzzle and ear color provide reliable means of estimating the age
of chacma infants under the age of 7 months.
Disease
*Diabetes mellitus in nonhuman primates: Recent research advances and
current husbandry practices. Howard, C. F. Jr. & Yasuda, M. (Div. of
Primate Medicine, ORPRC, Beaverton, OR 97006). Journal of Medical
Primatology, 1990, 19, 609-625.
. .
A review article updating recent research, which also furnishes
some information about therapeutic management and husbandry of monkeys
with predispositions toward diabetes or with overt diabetes.
*A new record of Bertiella studeri (Blanchard, 1891) Stiles and Hassal, 1902 from the crab-eating macaque Macaca fascicularis in Peninsular Malaysia. Krishnasamy, M., Jeffery, J., & Sirimanne, R. A. (Div. Medical Ecology, Inst. for Medical Research, Pahang Rd., 50588 Kuala
Lumpur, Malaysia). Tropical Biomedicine, 1988, 5, 89-90.
. .
Report of the tapeworm Bertiella studeri, which is of zoonotic
importance because human infections have been reported in many countries.
Ecology
*Polyspecific association of two tamarin species, Saguinus labiatus
and Saguinus fuscicollis, in Bolivia. Buchanan-Smith, H. (Dept. of
Psychology, Univ. of St. Andrews, St. Andrews, Fife, KY16 9JU, Scotland). American Journal of Primatology, 1990, 22, 205-214.
. .
The two species formed stable mixed associations and each mixed
species troop shared a common home range which they jointly defended
against neighboring mixed species troops. Both species appeared to
occupy a similar ecological niche, but differences were highlighted in
their methods of insect foraging, preferred height in the forest, and
mode of travel.
*Strategies of habitat use by mountain gorillas. Watts, D. P. (Dept.
of Biological Anthropology and Anatomy, Duke Univ., Durham, NC 27706).
Folia Primatologica, 1991, 56, 1-16.
. .
Variation in habitat quality influenced both long-term area occupation densities and shorter-term movement parameters. Home range use
patterns increased foraging efficiency. Interactions with other gorillas and poaching risk had effects superimposed on food distribution
patterns. There was a positive influence of group size on day journey
length, but it was only apparent at very small and relatively large group sizes.
*Grouping patterns of a mountain population of Macaca sylvanus in
Algeria -- A fission-fusion system? Menard, N., Hecham, R., Vallet,
D., Chikhi, H., & Gautier-Hion, A. (CNRS, UA 373, Station Biologique
de Paimpont F-35380, Plelan-le-Grand, France). Folia Primatologica,
1990, 55, 166-175.
. .
About 182 monkeys were studied from 1983 to 1988. The study population was characterized by a higher adult/immature ratio than forestliving troops, probably as a result of a higher rate of infant mortality. Possible causes of grouping patterns and population
characteristics are discussed in relation to environmental and social constraints.
Evolution
*The use of stone tools by wild-living chimpanzees. Kortlandt, A.
(88, Woodstock Rd., Oxford OX2 7ND, UK). The Newsletter (P.F.A.),
1991, 2[3], 1-3.
. .
The use of stone tools by chimpanzees and by earliest hominids show
no indication of homology, functional equivalence, similarity of motor
patterns, or identity of motivation.
General
*Proceedings of the XIIIth Congress of the International Primatological Society. International Journal of Primatology, 1990, 11, 619-628.
. .
Minutes of the meeting, held July 18-21, 1990, in Nagoya, and July
23-24, 1990, in Kyoto, Japan.
Genetics
*Use of isozymes in genetic management of nonhuman primate colonies.
Dyke, B., Williams-Blangero, S., Dyer, T. D., & VandeBerg, J. L.
(Dept. of Genetics, Southwest Foundation for Biomedical Research, San
Antonio, TX 78284). In Z. Ogita et al. (Eds.), Isozymes: Structure,
Function, and Use in Biology and Medicine (pp. 563-574). New York:
Wiley-Liss, 1990.
. .
Simulations based on 47 breeding-age males that were potential
sires of 115 infants lead to the following conclusions: Paternity
exclusion, using randomly assigned genotypes at 18 biochemical loci
based on known distributions of biochemical genetic markers, suggests
that up to 33% of the infants might be expected to have known sires.
About 8 breeding-age males might have sired each infant. If only half
the adult males were present, the proportion of pedigreed progeny
would increase to about 44%, and the number of potential sires per
birth would be reduced to 5. This would be marginal improvement, given
the loss of genetic variability that would result. The estimates
depend strongly on the demographic structure of the population and the
assumption of random mating. Different values are likely to hold for
other caged or wild populations.
Instruments & Techniques
*A bar code scoring system for behavioral research. Forney, K. A., Leete, A. J., & Lindburg, D. G. (D. G. L., Zoological Soc. of San Diego, P.O. Box 551, San Diego, CA 92112). American Journal of Primatology, 1991, 23, 127-135.
. .
A description of an application of bar code technology, and a discussion of advantages and restrictions of its use.
Nutrition
*Diet in one group of woolly spider monkeys, or muriquis (Brachyteles
arachnoides). Strier, K. B. (Dept. of Anthropology, Univ. of Wisconsin, 1180 Observatory Dr., Madison, WI 53706). American Journal of Primatology, 1991, 23, 113-126.
. .
During a 14 month study muriquis devoted an average of 51% of their
feeding time to leaves, 32% to fruits, and 11% to flowers. They
devoted a greater proportion of their feeding time to patchy fruit and
flower resources than expected from comparisons with smaller, sympatric howler monkeys. Leaves appeared to be eaten primarily to provide
necessary protein and/or bulk.
*Problems associated with long-term feeding of purified diets in rhesus monkeys. Golub, M. S., Gershwin, M. E., Lonnerdal, B., Keen, C.
L., Hurley, L. S., & Hendrickx, A. G. (CPRC, Univ. of California,
Davis, CA 95616). Primates, 1990, 31, 579-588.
. .
In an attempt to study the effect of dietary zinc, female rhesus
were fed either a marginally zinc-deprived diet or a comparable zinc-replete purified diet. Poor pregnancy outcome was noted in both
groups, especially after more than 2 years on the purified diet. These
data demonstrate the difficulty of constructing adequate purified
diets for long-term studies in primates, even when based upon available and recommended nutritional information.
Physiology
*Sociophysiology of squirrel monkeys. Mendoza, S. P., Lyons, D. M., & Saltzman, W. (Address same as above). American Journal of Primatology, 1991, 23, 37-54.
. .
A major consequence of social stimulation is altered regulation of pituitary-adrenal, pituitary-gonadal, and autonomic function. adult relationships are associated with distinctly different sociophysiological profiles than have been found for the mother-infant relationship, and do not conform to the stress/buffering model within which they are often interpreted.
*The complex relationship between behavioral attributes, social status, and whole blood serotonin in male Macaca fascicularis. Shively, C. A., Brammer, G. L., Kaplan, J. R., Raleigh, M. J., & Manuck, S. B. (Dept. of Comparative Medicine, Bowman Gray School of Medicine,
Winston-Salem, NC 27103). American Journal of Primatology, 1991, 23, 99-112.
. .
There were no persistent differences in whole blood serotonin (WBS)
between clearly dominant and subordinant monkeys, despite substantial
behavioral differences and stability of social status. WBS was best
predicted by a model that included interaction between rate of grooming and social status, extreme aggressiveness, and time spent alone.
*Estimation of immunoreactive testicular androgen metabolites in the
urine of saddle-back tamarins. Epple, G., Kuderling, I., Belcher, A.
M., Schafer, A., & Lerchl, A. (Monell Chemical Senses Center, 3500 Market St., Philadelphia, PA 19104-3308). American Journal of Primatology, 1991, 23, 87-98.
. .
Hydrolysis, extraction, and radioimmunoassay techniques measure
testosterone conjugates as the major steroid metabolites in Saguinus
fuscicollis. Results suggest that estimation of testosterone metabolite levels in urine is a valid method for the assessment of testicular activity in S. fuscicollis.
*Proteins: Biologically relevant components of the scent marks of a
primate (Saguinus fuscicollis). Belcher, A. M., Epple, G., Greenfield, K. L., Richards, L. E., Kuderling, I., & Smith, A. B., III
(Address same as above). Chemical Senses, 1990, 15, 431-446.
. .
Gel electrophoresis and concurrent behavioral studies demonstrate
that proteins contribute to the scent communication system in a primate. Tamarins discriminate scent material in which proteins have
been digested from material in which the proteins are intact, suggesting that proteins are a component of what constitutes the scent image.
Reproduction
*Seasonal variation of sperm morphology in the Mayotte brown lemur
(Eulemur fulvus mayottensis). Brun, B. & Rumpler, Y. (Fac. de Medecine, Inst. d'Embroyologie, 11, rue Humann, F-67085, Strasbourg, Cedex
France). Folia Primatologica, 1990, 55, 51-56.
. .
The Mayotte brown lemur shows continuous spermatogenesis throughout
the year, but motility and concentration of spermatazoa decrease during the non-breeding season, while teratospermia increases.
*Seasonal variation of the seminiferous epithelium cycle in Mayotte's
brown lemur (Eulemur fulvus mayottensis). Rasamimanana, P., Brun,
B., Meyer, J. M., & Rumpler, Y. (Address for Y. R. same as above).
Folia Primatologica, 1990, 55, 193-199.
. .
Spermatogenic activity is not arrested during the non-breeding season.
*Pregnancy, gestation and parturition in free-ranging Tana River
crested mangabeys (Cercocebus galeritus galeritus). Kinnaird, M. F.
(Dept. of Ecology & Evolutionary Biology, Princeton Univ., Princeton,
NJ 08544-1003). American Journal of Primatology, 1990, 22,
285-289.
. .
Mean gestation for 7 animals is 180 (�4.49) days. All females
showed postconception sexual swellings after the first 2 months of
pregnancy.
*Reproductive parameters in the sooty mangabey (Cercocebus torquatus
atys). Gust, D. A., Busse, C. D., & Gordon, T. P. (Yerkes RPRC, Field
Station, 2409 Taylor Rd., Lawrenceville, GA 30243). American Journal
of Primatology, 1990, 22, 241-250.
. .
Data from 67 females over 7.5 years indicated sexual maturation at
36.4�1.1 months and first parturition at 56.5�1.8 months, indicating a
period of adolescent sterility of over 1 year. Infant survival did not
vary significantly according to the age class of the mother. Births
and the onset of perineal swellings in young females showed a seasonal
distribution, with swelling cycles and conceptions rarely occurring
from April through September.
*Reproductive performance and excretion of urinary estrogens and gonadotropins in the female pygmy marmoset (Cebuella pygmaea). Ziegler,
T. E., Snowdon, C. T., & Bridson, W. E. (Dept. of Psychology, Univ.
of Wisconsin, Madison, WI 53706). American Journal of Primatology,
1990, 22, 191-203.
. .
Colony records and urinalysis reveal that the pygmy marmoset is a
nonseasonal breeder, producing twins (76%), triplets (8%), and singlets (16%). First births occurred between 24-42 months of age, and
5-27 months post pairing. Gestational length was a mean of 141.9 days
from the postpartum LH peak, which occurred a mean of 15.6 days after
parturition.
*Confirmation of ovulation and characterization of luteinizing hormone
and progesterone secretory patterns in cycling, isosexually housed
Bolivian squirrel monkeys (Saimiri boliviensis boliviensis). Alexander, S. E., Yeoman, R. R., Williams, L. E., Aksel, S., & Abee, C. R. (1801 Williams St. #401, Denver, CO 80218). American Journal of Primatology, 1991, 23, 55-60.
. .
A much greater elevation of serum estradiol (E�) has been measured
after mating than that observed in similarly cycling monkeys that did
not mate, raising the possibility that cycling squirrel monkeys may
not ovulate during nonmated cycles. Laparoscopy of isosexually housed,
cycling monkeys confirm ovulation and follicular rupture in the
absence of mating and delineate the relationship between periovulatory
lutenizing hormone, progesterone, and E� secretory patterns.
*Reassessment of age of sexual maturity in gibbons (Hylobates spp.).
Geissmann, T. (Anthropological Inst., Univ. Zurich-Irchel, Winterthurerstr. 190, CH-8057 Zurich, Switzerland). American Journal of
Primatology, 1991, 23, 11-22.
. .
Seven case reports on captive gibbons of known age show that cap-
tive males of H. leucogenys leucogenys and of the siamang (H. syndactylus) can breed at the age of 4 and 4.3 years, respectively. Captive H. lar x H. moloch hybrid and siamang females can breed at
5.1 and 5.2 years, respectively. It is not clear whether these ages
are the same in the wild.
*Collection and quality of rhesus monkey semen. Lanzendorf, S. E.,
Gliessman, P. M., Archibong, A. E., Alexander, M., & Wolf, D. P. (D.
P. W., Oregon RPRC, 505 N.W. 185th Ave., Beaverton, OR 97006).
Molecular Reproduction and Development, 1990, 25, 61-66.
. .
Description of the collection, processing, and evaluation of two
methods of penile electroejaculation. No correlation was found between
coagula volume and sperm numbers. The authors conclude that penile
electroejaculation, combined with control of stimulus current, provides a consistent, successful, and humane method for the collection
of semen in the rhesus monkey.
*Consortships and conceptions in captive rhesus macaques (Macaca
mulatta). Small, M. F. (Dept. of Anthropology, McGraw Hall, Cornell
Univ., Ithaca, NY 14853). Primates, 1990, 31, 339-350.
. .
The term "consortship" has been traditionally accepted as a specific nonhuman primate mating pattern, but the exact nature of this
behavioral pattern and its evolutionary importance have been less
clearly understood. This study demonstrates that for rhesus macaques,
who exhibit both consort behavior and more promiscuous matings, there
is no clear reproductive advantage to long-term consortships.
*Pregnancy-induced sclerosis in the myometrial vessels of cynomolgus
monkeys (Macaca fascicularis). Hayama, S.-I., Kamiya, S., & Nigi, H.
(Dept. of Wild Animal Medicine, Nippon Veterinary & Zootechnical College, Musashino, Tokyo, 180 Japan). Primates, 1990, 31, 427-429.
. .
Sclerotic changes were found histologically in the myometrial
vessels of 27 of 33 female cynomolgus monkeys. The changes were not
observed in nulliparous cases, but only in multiparous ones, suggesting that they are a pregnancy-induced phenomenon.
*In vitro fertilization in nonhuman primates. Dukelow, W. R., Pierce,
D. L., Roudebush, W. E., Jarosz, S. J., & Sengoku, K. (Endocrine
Research Ctr., Michigan State Univ., East Lansing, MI 48824). Journal of Medical Primatology, 1990, 19, 627-639.
. .
Ovulation induction, sperm capacitation, and fertilization have
been studied for over 50 years in nonhuman primates, but it has only
been in the past 20 years that extensive studies on sizeable numbers
of embryos have been done. Findings, mainly from the squirrel monkey,
baboon, rhesus, and cynomolgus macaque, indicate that the fertilization process appears to be similar to that of man and other mammals.
*Maternal blood pressure and fetal ultrasonography in normal baboon
pregnancies. Brans, Y. W., Kuehl, T. J., Hayashi, R. H., & Reyes, P.
(Hutzel Hospital, Dept. of Pediatrics, 4707 St. Antoine Blvd.,
Detroit, MI 48201). Journal of Medical Primatology, 1990, 19,
641-649.
. .
Pregnancy in baboons is characterized by lower systolic, diastolic,
and mean blood pressures than in the nonpregnant state. As pregnancy
progresses, diastolic and mean pressures tend to increase while systolic pressure remains low. Sonographic measurements of fetal growth
follow a sigmoid pattern, but their increase in relation to length of
gestation approximates a straight line between 6 and 21 weeks of gestation.
*Birth sex ratio and infant mortality rates in captive western lowland
gorillas. Mace, G. M. (Inst. of Zoology, Zoological Soc. of London,
London NW1 4RY). Folia Primatologica, 1990, 55, 156-165.
. .
Study of over 300 births to captive females showed a significant
trend for more male offspring to be born to older mothers, while very
young mothers (less than about 8 years old) give birth to only 31%
males. It is suggested that there are selective pressures to rear
males at a time when they can provision them most effectively, as male
reproductive success probably varies more than that of females and may
be dependent upon maternal care.
*Continuous fetal heart rate monitoring and tocodynamometry in routine
obstetrical care, fetal stress and non-stress testing, and labor
induction in the chimpanzee (Pan troglodytes). Mahoney, C. J., Gordon, M., & Briggs, D. (LEMSIP, NYU School of Medicine, 550 First
Ave., New York, NY 10016). Journal of Medical Primatology, 1990,
19, 681-714.
. .
Continuous fetal heart rate monitoring in the chimpanzee can provide a valuable tool for fetal assessment and management of labor in
any attempt to reduce the unacceptably high (16%) rate of perinatal
and neonatal infant mortality in captive chimpanzees in the US.
*Procedures for improving maternal behavior in captive chimpanzees.
Hannah, A. C. & Brotman, B. (Dept. of Psychology, Univ. of Stirling,
Stirling, Scotland FK9 4LA). Zoo Biology, 1990, 9, 233-240.
. .
Ten pregnant female chimpanzees were housed together with unrelated
infant chimpanzees to adopt, or with lactating female chimpanzees and
infants to observe, or with both. All ten showed appropriate maternal
behavior when their infants were born, in contrast to a group of eight
chimpanzees who had no such experience, and whose infants had to be
removed for hand-rearing.
Taxonomy
*The taxonomy of savannah baboons: An odontomorphometric analysis.
Hayes, V. J., Freedman, L., & Oxnard, C. E. (Dept. of Anatomy & Human
Biology, Univ. of Western Australia, Nedlands, Western Australia,
6009). American Journal of Primatology, 1990, 22, 171-190.
. .
Investigation of the dental dimensions of 443 savannah baboons,
representing the species Papio anubis, P. cynocephalus, and P.
ursinus, and a few hybrids, suggest that the three morphotypes are
clearly distinct. The hybrids are not intermediate, but different. The
findings, along with other data, tend to warrant maintenance of the
present specific distinctions.
-------------------------------------------------------------------
In many cases, the original source of references in this section has been the Current Primate References prepared by The Primate Information Center, Regional Primate Research Center SJ-50, University of Washington, Seattle, WA 98l95. Because of this excellent source of references, the present section is devoted primarily to presentation of abstracts of articles of practical or of general interest. In most cases, abstracts are those of the authors.
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. (Phone: 401-863-2511)
Judith_Schrier@brown.edu
ACKNOWLEDGMENTS
The Newsletter is supported by U. S. Public Health
Service Grant RR-00419 from the Animal Resources Program,
Division of Research Resources, N.I.H.
Cover drawing of a drill (Mandrillus leucophaeus) by Dr. Robert M. George, Department of Anatomy, University of South Carolina
Copyright @1991 by Brown University
Editor: Judith E. Schrier, M. Sc.
Associate Editor: James S. Harper, D.V.M.
Consulting Editor: Morris L. Povar, D.V.M.
Copy Editor: Elva Mathiesen, B. A.
Founding Editor: Allan M. Schrier, Ph.D.
{kind=link}
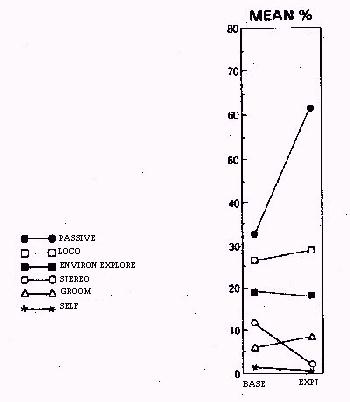{kind=link}