 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 44 NUMBER 2 APRIL 2005
Printable (PDF) Version of this issue
Articles and Notes
Environmental Enrichment for Monkeys Using Plants, by A. S. Chamove......1
Report on the Extent and Character of Primate Use in Scientific Procedures across Europe in 2001, by A. E. Rennie & H. M. Buchanan-Smith......6
Forage Box as Enrichment in Single- and Group-Housed Callitrichid Monkeys, by A. S. Chamove & L. Scott......13
News, Information, and Announcements
Information Requested or Available......5
. . .
AAALAC Student Information; More Interesting Websites
Grants Available: Aging Dissertation Awards to Increase Diversity......12
News Briefs......18
. . .
Perforation Caused Orangutan�s Death; New Orangutan Moves to Great Ape Trust; Bronx Zoo to Close Island Preserve; New Monkey Discovered in Northeastern India; Right to Name New Monkey Sells for $650G; Gorillas Cross to Rwanda; Sylvia Taylor; Primate Research Center for HIV/AIDS Opened; Proposed Standards of Care for Chimp Sanctuaries; Hollywood Apes Retiring to Sanctuary; Viet Nam Discovers Special Langur Monkeys; Guardian of Ghana�s Monkey Village Dies; Kenya Seizes Smuggled Baby Chimps; More Organisms Targeted for Genome Sequencing; Wolff Director of Compliance Oversight, OLAW; NCRR Associate Director of Comparative Medicine; Chimps Escape, Attack Visitors at Animal Sanctuary
Announcements from Publications......21
. . .
Call for Papers - JAAWS; PNAS Invites Submissions
Meeting Announcements......22
Volunteer Opportunities......22
. . .
Veterinarian - Sierra Leone; Kalaweit Rehabilitation Program
Resources Wanted and Available......24
. . .
Primate Info Net Seeking Digital Images; ACVP and ASVCP Proceedings Available On-Line; Veterinary Virology The Monkey Toy Store
Workshop Announcement: PASA Annual Management Workshop......25
Research and Educational Opportunities......25
. . .
Behavior and Ecology Courses - Costa Rica; Primatology/Ecology Field School - Kenya; CLASS Lab Animal Science Seminar
Calls for Award Nominations......27
. . .
AAALAC International Technician Fellowship Award; 2nd Annual Lab Products Animal Technician Award; HSUS 2005 Russell and Burch Award Nominations
Departments
Positions Available......23
. . .
Campus Veterinarian - NHLBI; Associate Operations Manager - Yerkes; Licensed Veterinary Technician/Technologist
Primates de las Am�ricas�La P�gina......26
Recent Books and Articles......28
* * *
Environmental Enrichment for Monkeys Using Plants
Arnold S. Chamove
Massey University School of Psychology
Introduction
It is commonly believed that making an enclosure more natural is an improvement and thereby an enrichment. The addition of plants to primate enclosures is generally believed to make the setting more pleasing for observers (Price et al., 1994) and presumably for the occupants as well. I could find no data on the effectiveness of plants in primate enclosures to evaluate this latter assumption. Published research (Chamove & Rohrhuber, 1989) suggests that in large outdoor enclosures, the cover provided by leaves is attractive to small primates. Most enrichment studies look at food-relevant (Chamove, 1989) or food-irrelevant (Renner et al., 2000) objects; plants are intermediate along such a continuum.
It is likely that the presence of protective cover is important for small diurnal primates. Observations of free-ranging common marmosets (Callithrix jacchus) in semi-natural conditions revealed that they prefer to travel and to forage through the densest of vegetation, in dense networks of thin, flexible, non-woody tangles wherever these are located (Chamove & Rohrhuber, 1989). Dense tangles are important, too, in the ecology of the red-crested tamarin (Garber, 1984). These tamarins (Saguinus oedipus geoffroyi) spend about 3 hours/day feeding and foraging. About 70% of this time involves searching for and capturing insects, and this is done primarily on branches under 3 cm in diameter; most climbing and jumping also occurs in pursuit of insects. About 24% of their feeding/foraging time involves plant material (Garber, 1980, 1984), and 20% of that time (37% of total time) is spent feeding on �fragile� branches under .8 cm in diameter and in dense vegetation, while 40% of this time on fragile branches is spent hanging from the branch.
Tamarins avoid vertical supports during travel. Wild callitrichids carry out only about 4% of their locomotor activities on vertical supports and these supports tend to be over 32 cm (Garber, 1980). When they do use vertical branches, they prefer to ascend on vertical branches between 2.5 and 12 cm. Most ranging is accomplished by long leaps beginning and ending on thin flexible non-vertical branches.
All of the marmosets and tamarins (Callitrichidae) are both arboreal and diurnal, with a natural diet consisting of approximately equal parts insects, fruits, and tree and plant exudates (Epple, 1975; Garber & Sussman, 1984), suggesting that the distribution of branches might be important to their activity patterns. The �claw-like� nails possessed by both tamarins and marmosets (Garber, 1980 p.186) allow these primates to move along supports ranging from under 0.8 to over 32 cm in diameter (Garber, 1984). However, the orientation, spacing, and size of the branches would be expected to have important effects on energy expenditure in these monkeys. Although metabolic costs of locomotion are relatively high in small primates (Martin, 1979), it is much easier for smaller animals to climb than larger ones if one considers the maximum muscle pull and tendon stressors (Coimbra-Filho & Magnanini, 1972; Collins, 1983).
I sought to evaluate the behavioral effect of growing plants on the walls of our primate enclosure by hanging mesh and growing plants up the mesh.
Methods
To see if adding plants and supports next to wall areas would alter behavior, the 30 subjects in these studies were chosen from cotton-top tamarins (Saguinus oedipus oedipus) living in five large family groups, all housed in Stirling University Psychology Primate Unit, Scotland (Price & McGrew, 1990). They were fed a high protein diet early mornings and a fruit diet in the early afternoon. The families had from 9 to 15 members; 6 animals were selected from each family to be the focal animals, although some data was collected from all animals. The selected subjects were the two parents, two oldest adult twin offspring, and the two youngest independent offspring. The animals were chosen so that the different ages of the subjects could be compared with regards to the patterns of locomotion performed; the older animals were expected to be less active (Baker, 2000). Since the oldest offspring and the parents in each family were approximately the same weight, we hoped to examine the effect of age on locomotion while weight was constant (see Table 1).
All testing was carried out in the 33-m3 home cages of the subjects, which, although varying slightly, measured approximately 310 x 320 x 333 cm. The cages contained a variety of branches, most suspended from the mesh of the ceiling. The branches had been cut green from willow and poplar trees around the campus and varied in size, orientation, and flexibility. They lost their leaves soon after being cut and rapidly dried. They were placed so routine husbandry was not obstructed, so few were in the center and few were in the bottom third of the cage. There was a total of 44 linear meters of branches. Of those, 36 linear meters were over 10 mm in width, and 24 meters were both non-vertical and over 10 mm wide.
| Juvenile | Young Adult | Parent | Mean | ||
| Weight | Mean Range | 404 400-408 | 491 489-493 | 531 495-567 | 475 |
| Age | Mean Range | 10 10-10 | 36 36-36 | >75* 75 |
Table 1: Mean ages (months) and mean weight (grams), range in italics, for the three categories of tamarin observed (n = 30).
* Importation records suggest these wild-born animals were at least 6.2 years old.
Apart from the branches, the enclosures also contained nest boxes, feeding platforms, horizontal perches, and some small mesh cages, which were sited against the back wall of the enclosure. These structures were counted as �other� parts of the cage for the purpose of data collection and totaled approximately 4 m2 in horizontal surface area per enclosure. All of the home cages had a full 10.3 m2 vertical mesh front. The other three walls were made of smooth concrete and could not be climbed by the monkeys. Details of husbandry can be found elsewhere (McKenzie et al., 1986; Price & McGrew, 1990).
Figure 1: Trellis with only dead branches.
Three types of trellis were put in each room, side by side in a random order along one wall. Each type was approximately 0.9 m wide and 2.9 m high. The two flexible mesh types were made of dark green plastic, one with 15 mm holes, the other with larger 50 mm holes. As the monkeys could not pass through either of the flexible mesh types, six larger holes were cut at irregular intervals in each sheet. The inflexible trellis was made of 30 mm-wide rigid strips of wood with 150 mm square holes. All were designed for supporting plants and purchased from garden centers. The experiment was to investigate the effect of plants and not of different trellises. The three trellis types produce minor findings in this study, and are only included because it may be of some interest in choosing trellises for small monkeys.
The three trellis sheets were mounted 100 mm from the walls and empty grow-bags placed at their base. The animals were allowed to adapt to their presence for a month before the first period of data recording began (�MESH�), more than enough for adaptation (Chamove, 1996). A fortnight later and when the plants were growing well in their bags and ready to be put in the enclosures, a second test was done with only the three types of mesh present, also called �MESH.�
Figure 2: Trellis with plants growing.
Plants had been started into growth in grow-bags and were moved into the rooms when the plants needed support, at about 200 mm high. After three weeks and when the strongest growing plants had reached about 3 m in height, the second collection of data took place (�MESH + plants�). One month later when all of the growing plants had reached the top of the trellis, the third collection took place (�MESH + PLANTS�). Two months later when most of the plants had died, all of the stems were removed from the mesh and the final data collection session took place with the again empty grow-bags still in place but no plants (�MESH�). When the three sets of data collected from the mesh-only conditions were compared (before plant growth, just before plants were put into rooms, and after plants were removed from rooms), and when no differences were found, they were combined for analysis and are termed the �MESH� condition.
The following plants were assessed: the annuals runner beans, nasturtium, canary creeper (Tropaeolum speciosum), climbing green beans, peas, sweet peas, black-eyed Susan (Thunbergia alata), cathedral bells (Cobaea scandens), cucumber, morning glory (Ipomoea), melon, pumpkin, Scotch (sic) marigold (Calendula), and the perennial climber Russian vine (Polygonum baldschuanicum). The only plants making substantial growth were the runner beans, the two tropaeolum (nasturtium and canary creeper), and the Russian vine. The others presumably suffered from low light levels.
The motor activity seen on and off the mesh was recorded. These activities were classified into six types, the first three active, the last three inactive:
During each of the experimental conditions, the activity (described above) of the subjects was recorded for six animals in each of five families of cotton-top tamarins using focal-animal sampling; the order of testing subjects was strictly random within families. In two groups there was only a single oldest and single youngest offspring and not the usual twins. In one of these the three oldest offspring were used. In each of these conditions, 30 animals were observed, each for two 5-min sessions on two different days, during which the six categories of activity were recorded. Activities involving the mesh and plants were differentiated from those elsewhere in the enclosures.
In addition, behavior on the plants was recorded by observing all independently locomoting animals in the family for two 30-min sessions and noting the time spent on the mesh/plants, the number of passages through the mesh, the time spent behind the mesh, and instances of manipulations of the plant material. It was thought that the animals might hide behind the plants and inspect them for insects. All of the above were recorded as frequency per 10-sec period, except for passages through the mesh and manipulations of the plant material, which were recorded every time they were observed. Because of the novel exploratory nature of the research, it was decided that time should be spent in sampling from a large number of animals for a rather short period of time rather than from fewer animals for longer periods.
Analysis of data was done using multivariate analysis of variance. The analysis used percent of total possible time as the measures. The analysis used the three ages of monkey, the three conditions, two mesh locations-on or off-, two categories of activity, and six levels within each category. Alpha was set at 0.05 throughout.
Results
Figure 3: Percentage of time spent in six activities in the presence of bare mesh twice before and once after testing with plants (MESH), with mesh supporting smaller plants (MESH + plants), and with mesh and taller plants (MESH + PLANTS). Top six behaviors are those shown when off the mesh; bottom six (below 6%) are behaviors shown while on the mesh.
When the walls had no mesh on them, the monkeys spent under 1% of their day on them, mostly bouncing off them. When the mesh had no plants on it, the monkeys spent about 15% of their day on it. When the plants were growing on the mesh, this reduced to about 10% of the day. This percentage compares with other data reporting 19% of observations on vertical mesh in marmosets (Dettling & Pryce, 1998). As illustrated in Figure 1 (lower 6 behaviors), the presence of plants reduced both active (leap, run, climb) and inactive (hang, sit, walk) behaviors on the mesh, F6,98 = 6.5, p < 0.0001. As expected, the presence of the mesh also reduced the behavior off the mesh (6 behaviors in the upper half of figure), especially the less active behavior patterns, F10,127 = 3.2, p = 0.001. Leaping was rarely seen onto the large-holed wooden and medium-holed plastic mesh, and hanging was about halved on these substrates in comparison with the small-holed plastic mesh, F6,98 = 36.5, p < 0.0001. Climbing, too, was more frequent on the two plastic mesh types than on the wooden trellis.
Figure 4: Percentage of time on large-holed wooden, medium-holed plastic, and small-holed plastic mesh for three ages of monkey separated into time behind the mesh (right) and total time on the mesh (left).
Overall, all the recorded behaviors decreased in the presence of plants. The active behaviors such as �walk� and �run� decreased from about 11% to 9% when animals were observed off the mesh. They were even less common (decreasing to 3%) when on the mesh. The active behaviors off the mesh of hanging and climbing decreased from 23% and 10% to 14% and 5% when plants were growing; while on the mesh the same behaviors dropped from 4% and 4.5% down to 1.5% and 2.5% respectively.
There was a clear preference by the monkeys for the large-holed wooden trellis, followed by the medium-holed plastic mesh, F2,34 = 58.1, p < 0.0001. While the animals spent least time on the small-holed plastic mesh, the plants grew best on this variety of mesh; plants grew poorest on the wooden mesh. We think the animals preferred the wooden mesh as they could pass through it at any point and they could easily sit on it; we think the plants grew best on the plastic mesh as it gave closer support and had no sharp, rigid edges.
There were few significant interactions of mesh size with age or size of monkey. The right half of Figure 4 shows that parents spend significantly less time behind the mesh than the younger animals, F4,34 = 4.1, p < 0.01. The left half shows the stronger preference by all sub-groups for the large wooden mesh, F4,34 = 3.2, p = 0.02.
Discussion
The presence of growing plants did not have the dramatic effect expected by their desirability to both the public and staff in progressive zoos around the world. Any cost-benefit analysis based on our data here would suggest an unfavorable ratio where plant maintenance costs are high. In our situation even growing the plants was problematic and costly, and did not appear to make a clear and interpretable difference in the behavior of the monkeys. Other dead plant material seems a better bet in this situation (e.g., McKenzie et al., 1986).
One salient difference when comparing the wild, a zoo, and a laboratory is the lack of anything living in the environment of the last. The advantages of materials of plant origin in zoos and laboratories are well documented (Chamove, 1989; Fajzi, et al., 1989). However, it seems from this limited study that growing plants in such a hostile environment is not worth the cost if the goal is to measurably enrich the lives of small arboreal monkeys.
It was clear that live plants were an improvement for the monkeys, even though the change in behavior was not great. One behavior which clearly differentiates enrichment from stress is locomotion. In stressful situations, high-level locomotion such as running and low-level locomotion such as walking increase; with enrichment both behaviors decrease (Chamove, 1997). With plants all active behaviors decreased, both when on and off the mesh, suggesting enrichment, albeit at a low level.
A more substantial benefit from the vertical mesh was that a part of the cage that is normally inaccessible to the monkeys, namely the bare smooth walls, was now accessible and was used, on average, 10% of the day, and the vertical mesh area of the cage was increased by 75%. When the large mesh was present, they spent 30% of the time on it.
References
Baker, K. C. (2000). Advanced age influences chimpanzee behavior in small social groups. Zoo Biology, 19, 111-19.
Chamove, A. S., & Rohrhuber, B. (1989). Moving Callitrichid monkeys from cages to outside areas. Zoo Biology, 8, 151-163.
Chamove, A. S. (1989). Environmental enrichment: A review. Animal Technology, 40, 155-178.
Chamove, A. S. (1996). Cage cleaning: Interest or intrusion? Australasian Primatology, 11, 2-5.
Chamove, A.S. (1997). How do we decide what works: Evaluating enrichment research. Australasian Primatology, 11, 4-5.
Collins, B. H. (1983). Climbing and some effects of arboreality on the natural history of primates. Doctoral Thesis, University College, London.
Coimbra-Filho, A., & Magnanini, A. (1972). On the present state of Leontepithecus and some data about new behavioral aspects and management. In D. A. Bridgewater (Ed.), Saving the lion marmoset (pp. 59-69). Wheeling, WV: Wild Animal Propagation Trust.
Dettling, A., & Pryce, C. R. (1998). Physical environment and its influence on behaviour in captive common marmosets (Callithrix jacchus). In European Marmoset Research Group (Ed.) , Marmosets: A handbook for fundamental and applied research (pp. 282-309). New York: Academic Press.
Epple, G. (1975). The behavior of marmoset monkeys (Callithricidae). In L. A. Rosenblum (Ed.), Primate behavior, Vol. 4 (pp. 195-239). New York: Academic Press
. Fajzi, K., Reinhardt, V., & Smith, M. D. (1989). A review of environmental enrichment strategies. Laboratory Animal, 18, 23-35.
Garber, P. A. (1980). Locomotor behavior and feeding ecology of the Panamanian tamarin (Saguinus oedipus geoffroyi, Callitrichidae, Primates). International Journal of Primatology, 1, 185-201.
Garber, P. A. (1984). Use of habitat and positional behavior in a neotropical primate, Saguinus oedipus. In P. S. Rodman & J. G. H. Cant (Eds.), Adaptations for foraging in nonhuman primates: Contributions to an organismal biology of prosimians, monkeys, and apes (pp. 112-133). New York: Colombia University Press.
Garber, P. A., & Sussman, R. W. (1984). Ecological distinctions between sympatric species of Saguinus and Sciurus. American Journal of Physical Anthropology, 65, 135-146.
Martin, R. D. (1979). Phylogenetic aspects of prosimian behavior. In G. A. Doyle & R. D. Martin (Eds.), The study of prosimian behavior (pp. 307-357). New York: Academic Press.
McKenzie, S. M., Chamove, A. S., & Feistner, A. T. C. (1986). Floor coverings and hanging screens alter arboreal monkey behavior. Zoo Biology,5, 339-348.
Price, E. C., Ashmore, L. A., & McGivern, A.-M., (1994). Reactions of zoo visitors to free-ranging monkeys. Zoo Biology, 13, 355-373.
Price, E. C., & McGrew, W. C. (1990). Cotton-top tamarins (Saguinus (o.) oedipus) in a semi-naturalistic captive colony. American Journal of Primatology, 20, 1-12.
Renner M. J., Feiner, A. J., & Orr, M. G. (2000). Environmental enrichment for New World primates: Introducing food-irrelevant objects and direct and secondary effects. Journal of Applied Animal Welfare Science, 3, 23-32.
* * *
Information Requested or Available
AAALAC Student Information
A new feature of AAALAC International�s redesigned Website is a section specifically for students looking for general information on animal research and testing. The section contains links to materials from many organizations, answering questions most commonly received from students. This is a resource for anyone with questions about how and why animals are used in medical research: <www.aaalac.org/resources/studentinfo.cfm>.
More Interesting Websites
* * *
Report on the Extent and Character of Primate Use in Scientific Procedures across Europe in 2001
Anita E. Rennie and Hannah M. Buchanan-Smith
Department of Psychology, University of Stirling
Introduction
In this report we present and discuss statistical data available from those European countries that use non-human primates (henceforth �primates�) in research. Primate-user countries in the European Union include Austria, Belgium, Finland, France, Germany, Italy, the Netherlands, Spain, Sweden, and the U.K.; all are included in the analysis as far as possible. We also consider primate-use statistics for Switzerland, a country which, although not a member of the European Union and not subject to the provisions of European Directive 86/609/EEC (European Union, 1986), has used a number of primates in research in the past and has published statistical data regarding its use of animals in scientific procedures in 2001.
Countries which are members of the European Union are legally required to provide statistical data on the numbers of animals they use in scientific research and information about the origin of those animals and the purpose of procedures in which they are used (European Union, 1986). This information is published by the Secretary General of the Council of Europe and is available to members of the public throughout the world. These statistical data must be presented in accordance with Appendix B of the European Convention on the Protection of Animals Used in Experimental and Other Scientific Procedures (ETS 123 - European Commission, 1986, 1992). The Swiss statistical data on the use of animals in scientific procedures are also published by the Council of Europe in the same format. In some countries (including Germany) implementation of the new recording procedures has required amendments to national legislation, thus delaying compliance with the new regulations (European Commission, 1992). It is partially for this reason that some data for 1999 and 2001 remain unavailable. In accordance with the Convention (European Commission, 1986), the statistics published by the Council of Europe include only experimental animals and not those kept in laboratories for breeding purposes or those kept as stock animals. The term �procedure� is defined in the European Convention (1986) as: �any experimental or other scientific use of an animal which may cause it pain, suffering, distress or lasting harm, including any course of action intended to, or at least liable to, result in the birth of an animal in any such conditions, but excluding the least painful methods accepted in modern practice (that is �humane� methods) of killing or marking an animal� (European Commission 1986, Article 1, Paragraph 2c).
For the purposes of this review, statistical data on the number of nonhuman primates used in scientific procedures in each primate-user country were extracted from animal use statistics for 2001, published in 2003 (Council of Europe, 2002; Home Office, 2002) and the uses and sources of these primates are examined. It should be noted that statistics for Austria, Italy, and Spain, countries that have used a number of primates in research in the past (Commission of the European Communities [�CEC�], 2003), are as yet unavailable for 2001. The statistics discussed in this report do not include data from these countries, but do include data from the majority of primate-user countries in Europe. Statistics from 2001 are compared with those from 1999, the most recent complete set of statistics on animal use across Europe (CEC, 2003), published in January, 2003.
Numbers of Primates Used in 2001
Based on statistics available for 2001, a total of 11,081 primates were used in scientific research across Europe (Council of Europe, 2002; Home Office, 2002). It should be noted that, until data for all European primate-user countries become available, this number represents an under-estimation of the total number of primates used. Data on the taxonomic breakdown of primates used are also incomplete and, for the purposes of this review, data will be considered proportions (expressed as percentages) of the total number of primates or taxonomic group of primates used in science. From available data, 75% of primates used in research across Europe were Old World monkeys, 15% were New World species, and 10% were prosimians. One ape (0.01% of primates used) was used in the Netherlands. The proportion of each group of primates used in 2001 is comparable with figures for 1999, in which 72% were Old World species while 19% were New World monkeys and 9% were prosimians. In 1999, six apes (0.07%) were used, again in the Netherlands. These data are presented in Figure 1.
Figure 1: Number of primates of each taxonomic group used in Europe as a proportion of the total number of primates used that year in a) 1999 and b) 2001.
Despite the similar spread of primate use across taxonomic groups in the years 1999 and 2001, the overall number used increased by around 2000 animals (9097 and 11081 primates for 1999 and 2001 respectively - Council of Europe, 2002; Home Office, 2002; CEC, 2003) and final figures for 2001 (including estimated numbers used in Austria, Italy and Spain, based upon previous data) are likely to bring the total increase to nearer 2500. Also, primate species account for a larger proportion of the total number of animals used in research in 2001 compared to 1991 (CEC, 1994), 1996 (CEC, 1999) and 1999 (CEC, 2003) (0.07%, 0.09%, 0.09% and 0.12% for 1991, 1996, 1999, and 2001 respectively). Because European statistics on animal use are not reported every year, it is impossible to determine whether the increase is a real trend or simply representative of yearly fluctuations in primate use. In order to provide some information about the sort of trends and fluctuations seen in the use of primates, statistical data from the U.K. (one of the main primate-user countries in Europe) from 1991-2002 were examined (Home Office, 1992, 1993, 1994, 1995, 1996, 1997, 1998, 1999, 2000, 2001, 2002, 2003). These data are presented in Figure 2. The data show that increasing and decreasing trends in primate use have occurred over periods of three to four years, but yearly fluctuations are also apparent (Figure 2a). It is likely that these short- and longer-term fluctuations represent changes in research approaches and the availability of funding, rather than an overall tendency to increase or decrease primate use. In Figure 2b it can be seen that there has been a slight but consistent decline in the number of New World monkeys used (mainly the common marmoset (Callithrix jacchus) as a proportion of total primate use in the U.K. since 1993. Conversely, the number of Old World monkeys used (mainly long-tailed [Macaca fascicularis] and rhesus [Macaca mulatta] macaques) again as a proportion of total primate use, has gradually increased. In 2001, most European countries used more Old World than New World monkeys, which is consistent with the trends observed in the U.K. over the last decade.
Figure 2: a) Total number of primates used in research in the U.K. between 1991 and 2002. b) The number of prosimians and New and Old monkeys used, as a proportion of the total number of primates used in the U.K., between 1991 and 2002.
The use of primates in experimental procedures must be justified from an ethical perspective and valid from a scientific perspective. The number of primates used in each experimental procedure must be determined by balancing the need to minimize both harm (Refinement) and the number of animals used (Reduction). In some cases more primates may be used in order to reduce the costs of the procedure on each individual primate. Thus, an increase or decrease in the number of primates used does not necessarily reflect an overall increase or decrease in welfare costs. Because of this interplay between Reduction and Refinement, the justification and validity of each study using primates is perhaps more important than the actual number of animals used.
France, Germany and the U.K. have traditionally been the most prolific primate-user countries in Europe and this is still evident in 2001 (Council of Europe, 2002; Federal Ministry of Consumer Protection, Nutrition and Agriculture, 2003; Home Office, 2002). In 2001, more than 84% of all primates used in research across Europe were used in France, Germany and the U.K. See Table 1 for details.
| 1999 | 2001 | |||
| Country | no. | % | no. | % |
| U.K. | 3191 | 35 | 3342 | 30 |
| France | 2322 | 25.5 | 3840 | 35 |
| Germany | 2084 | 23 | 2115 | 19 |
| Netherlands | 320 | 3.5 | 393 | 3.5 |
| Belgium | 490 | 5.4 | 710 | 6.4 |
| Switzerland | * | * | 512 | 4.6 |
| Italy | 512 | 5.6 | * | * |
| Czech Republic | * | * | 92 | 0.8 |
| Sweden | 66 | 0.7 | 77 | 0.7 |
| Spain | 96 | 1.1 | * | * |
| Austria | 7 | 0.1 | * | * |
| Finland | 9 | 0.1 | 0 | 0 |
| Total | 9097 | 100 | 11081 | 100 |
Table 1: The use of primates in each user country in 1999 and 2001, expressed both as a total for each country and as a percentage of the total number of primates used in Europe of that year. * = data unavailable
Extent to which Primates are Re-Used
Parties to the Convention agreed that for the purposes of Article 27 and 28 of the Convention (1986, ETS 123), the term �re-use� should be defined and recorded as follows: �If in the framework of one single procedure it is necessary for scientific reasons to make more than one intervention on the same animal, this is not regarded as re-use. However, if in the framework of two or more unrelated and independent procedures, the same animal is used whereas a different animal could equally well have been chosen, this second or subsequent use is regarded as re-use� (European Commission 1992, paragraph 9).
The Parties consider that this information should be collected, but without specification of the number of times that the same animal has been re-used, so that re-use of an animal is counted only once in each year in which it is re-used. Data concerning re-use are very incomplete for 2001, with only three European countries reporting this statistic correctly or at all (Belgium, France, the Netherlands). Of the countries that did report on the extent of re-use of primates, the Netherlands is the only country that has consistently re-used more than half of laboratory-housed primates, 66% in 1999 and 58% in 2001. While Belgium re-used 17% of primates in experiments in 1999, only 9% were re-used in this country in 2001; and France, the greatest user of primates in 2001, was also the country that re-used animals the least (1.0% and 1.9% of primates re-used in 1999 and 2001 respectively). No total figure for re-use of primates is available for 2001; however in 1999, 17% of primates were re-used overall in Europe (CEC, 2003). The decision to re-use animals in more than one experiment, or to use greater numbers of animals in only one experiment, is likely to be primarily dictated by the purpose of the procedures concerned and is controversial on both ethical and welfare grounds. Re-use of animals is an issue about which there is no universal agreement (Fentener van Vlissingen, 1999). The re-use of healthy animals ensures that the need for na�ve animals is reduced. The animal is kept longer in a laboratory environment and there is therefore more opportunity to work with, develop a relationship with, and, potentially, train him or her. Thus, the adverse effects of the procedures themselves and of maintenance of the animal in the laboratory can be reduced and welfare improved overall (Bayne et al., 1993; Reinhardt, et al., 1995; Bayne, 2002; Prescott & Buchanan-Smith, 2003). Conversely, the cumulative harm experienced by an individual exposed to repeated experimental interventions and maintained in a relatively barren environment might be considered to constitute unreasonable suffering. The scientific validity of the animal as a model may also be compromised by its exposure to previous procedures. The ethical and scientific acceptability of re-use should therefore be considered on a case-by-case basis, taking into account the relative harm associated with the use of na�ve animals and that of re-use and the effects of re-use on scientific validity (Fentener van Vlissingen, 1999).
Purposes of Procedures in Which Primates Are Used
The statistics were also examined in order to determine the purposes of procedures in which primates are used in science across Europe, according to the categories required in the Convention (1986, ETS 123) (European Commission, 1986). The statistics show that experiments on primates are restricted to five categories of research as follows; diagnosis of disease, fundamental biological research, applied studies in human and veterinary medicine (not including safety evaluation), toxicology and other safety evaluations (including those related to studies in human and veterinary medicine), and other (Council of Europe 2002; Home Office, 2002). These statistics are presented differently by the U.K. Home Office. In U.K. Home Office publications, the category �applied studies in human and veterinary medicine� includes toxicological evaluation and efficacy testing, while the category �protection of man, animals and environment (toxicology)� includes other toxicology, but not that related to human and veterinary medicine. According to the Convention (ETS 123), all statistics on toxicological tests should be recorded together (European Commission, 1986). The statistics given by the U.K. are recalculated for presentation in the format required by the Convention (ETS 123) (CEC, 2003); however the revised �European� presentation of data from the U.K. have not yet been published for 2001. The Czech Republic presents these data in the same way as the U.K. Home Office. For the purposes of this review, the data on the categories of experiments will reflect data from countries other than the U.K. and the Czech Republic. Data from the U.K. and the Czech Republic are considered separately in the text.
Figure 3: Uses of primates in France, Germany and the Netherlands in 2001 according to category: (top) for all primates as a proportion of the total number of primates used; (center) for New World monkeys as a proportion of all New World monkeys used; and (bottom) for Old World monkeys as a proportion of all Old World monkeys used.
Overall, primates are used with almost equal frequency in applied studies in human and veterinary medicine and in toxicological studies (43 and 40% respectively) (Council of Europe, 2002). 16% of primates are used in studies of a fundamental nature. 1% of primates are used to diagnose disease or in �other� studies. These data are presented in Figure 3a. Prosimians are only used in Germany and France: 55% of these were used in fundamental studies, and the remainder were used in applied studies and in toxicology (24% and 21% respectively). As can be seen in Figure 3b, 33% of New World primates were used in fundamental studies, 44% in applied studies and 21% in toxicological studies. In contrast, 57% of Old World primates were used in toxicological research, while only 6% were used in fundamental studies. A significant proportion of Old World primates (36%) were also used in applied studies for human and veterinary medicine (Figure 3c).
France was the only country that used the majority of primates in toxicological studies (68%). In all other countries, primates were used mainly in applied studies for human and veterinary medicine (Belgium, 95%; Czech Republic, 100%; Germany, 72%; the Netherlands, 65%; Sweden, 66%; Switzerland, 72%; U.K., 77%).
Data from the Czech Republic and the U.K. show that most toxicological and safety evaluations carried out on primates are associated with applications in human and veterinary medicine (including toxicological evaluation) (100% and 77% respectively). In the U.K., primates were not used in any other toxicological safety tests: 20% of primates were used in studies of a fundamental nature and 3% were used to diagnose disease. The majority of experiments using Old World monkeys were applied studies (87%), while 56% of New World primates were used in these studies. 34% of New World primates were used in fundamental studies.
Sources of Laboratory Primates
Breeding establishments for laboratory animals in countries that are party to the Convention (European Commission, 1986) and regulated by Council Directive 86/609/EEC (European Union, 1986) must be registered in accordance with Article 15 of the Directive and must be afforded a minimum standard of care in accordance with Article 5 of the Directive. In order to be registered, a competent member of the staff, who is ultimately responsible for the care of the animals in the establishment, must be named (Article 16 of the Directive). Under Articles 17 and 18 of the Directive, each animal must be permanently identified, and records on origin, arrival and departure dates, destinations, and deaths must be kept for all animals. Under Article 19 of the Directive, animals used in research must be obtained from registered breeding or supplying establishments, unless exemption has been obtained from the authority controlling experiments in the establishment using them. For many species of laboratory animals, this law has resulted in animals being bred �in-house� and most are easily available within EU Member States. These animals are therefore protected by the Directive and the European Convention (1986, ETS 123). This has reduced the number of animals that must be transported from outside the European Union and therefore any welfare issues associated with such transportation (Wolfensohn, 1997). Although this must be considered an improvement, it should be noted that transportation within the European Union continues and in most cases is likely to have an adverse affect on animal well-being.
Old World monkeys however, are less likely to be bred �in house� and the statistics for 2001 show that the majority of Old World monkeys are obtained from sources not covered by European legislation (Council of Europe, 2002), although it should be noted that these sources may be subject to other legislation. In total (using data for Belgium, France, the Netherlands, Sweden, U.K.) 65% of laboratory-housed primates were obtained from countries not covered by European legislation in 2001, while 15% of primates were bred within the establishment in which they were used. Under Article 15 of the European Directive, establishments that breed primates in source countries and supply them to European Countries that are Party to the Convention must be approved by the European authorities or must be registered. In order to obtain approval, these establishments must comply with the very basic provisions of the Directive (Articles 5-14). However, the breeding and supplying establishments themselves are not compelled by law to maintain such standards and exemption from Article 15 of the Directive can be obtained under Articles 19(4) and 21 of the Directive.
Figure 4: Sources of primates used in research in the U.K. since 1995.
There are differences in the ease with which different species of primate are bred in laboratories. The common marmoset, the species of New World monkey that is most frequently used in laboratories, can be bred relatively easily (Poole, et al., 1999). In 2001, 48% of the New World monkeys used in science (using data from Belgium, France, the Netherlands, and Sweden) were bred within the user establishment, and in total 57% were obtained from countries regulated by Council Directive 86/609/EEC (1986). There is, however, wide variation in the extent of in-house breeding between primate-user countries. In the Netherlands, in 2001, 93% of New World monkeys used in science were bred within the user establishment, while in France, 42% of New World monkeys were bred on-site. In total 53% were bred in countries subject to European legislation. However, the remaining 47% of New World primates used in France in 2001 were obtained from countries that are not subject to European law.
In contrast to New World monkeys, 87% of Old World monkeys used in science in Europe in 2001 were obtained from countries that are not regulated by Council Directive 86/609/EEC (1986) (again using data available from Belgium, France, the Netherlands, and Sweden). However, it is likely that this total gives a somewhat distorted view of the source of Old World primates, as detailed data on sources of primates were not reported by Germany and the U.K., two of the three main primate-using countries. France, the third main user of primates in Europe, obtained 90% of its Old World monkeys from countries that are not regulated by Council Directive 86/609/EEC (1986). In 1999, the U.K. obtained more than 84% of Old World primates from within the EU (CEC, 2003). Thus the 87% of Old World monkeys obtained from countries that are not regulated by Council Directive 86/609/EEC (1986) reported above is likely to be artificially high. Home Office data on the sources of all primates used in research in the U.K. were considered in order to show how the sources of animals may change over time. These data are presented in Figure 4 from 1995 to 2002, as data are unavailable before this time. It appears from these data that the number of primates being bred within U.K. research establishments or being obtained from other sources in the U.K. is declining, while the number obtained from countries not subject to European legislation is increasing - but this probably is because the U.K. appears to be using an increasing proportion of Old World monkeys.
Summary and Conclusions
Examination of all the currently available statistical data on the use of laboratory animals in 2001 in European primate-user countries indicates that at least 11,081 primates were used in science in 2001, and that the majority of these were Old World monkeys. This total is higher than that for 1999, although Old World monkeys were also used more frequently than New World species in 1999. Examination of data from the U.K. for the years 1991 to 2002 shows that the number of primates used was subject to short term (1 year) and longer term (3-4 years) fluctuations and no overall trend was apparent in the data. On the basis of this evidence, the apparent rise in primate use in science in Europe between 1999 and 2001 cannot be considered to be indicative of an increasing trend in primate use overall, but rather is likely to be indicative of yearly fluctuations in primate use. Annual increases or decreases in the total number of primates used may not be reflective of changes in welfare overall, but simply the balance between reducing numbers, minimizing harm, and the importance of scientific validity. Data from the U.K. for 1991-2002 indicate that the use of Old World primates is comparatively higher and has been increasing over the last 10 years, while the use of New World species is declining slightly in this country. The U.K., France, and Germany were the main users of primates in 2001 and also in 1999, but other countries continue to use a small but significant number of primates. The re-use of primates occurred most frequently in the Netherlands, where over 50% of primates were re-used in 1999 and 2001. While the re-use of primates in science remains controversial, it is suggested that the decision to re-use must be based upon the relative costs and benefits of the re-use, the use of naive animals, and the scientific validity of the animal as a model. The statistics show that in 2001 primates were used mainly in applied studies and toxicology and safety evaluations, but also in fundamental studies. The data also show that more Old World monkeys were used in toxicological research than New World monkeys and that New World monkeys were used mainly in fundamental research and in applied studies. From data available so far it was found that more than half of all primates used in research in 2001 were obtained from countries outside the EU, where they are not protected by the European Directive (86/609/EEC ETS 123) and the European Convention. Data from the U.K. show that the number of primates obtained from within EU-regulated countries has consistently declined over the last five years.
This report is based upon an incomplete set of statistics on the use of animals in science across Europe for 2001, and the conclusions that can be drawn from it must be taken with caution. However, the report provides a useful picture of the current extent and character of primate use in Europe, against which discussion and prioritization of welfare and other issues can be made. Although the standardization of the collection and presentation of statistics, as agreed by Parties to the Convention in 1997 (European Commission, 1992), has gone some way to improve the accessibility of the data, inconsistencies remain. These inconsistencies will be resolved as the Parties to the Convention change national practice to come into line with current and future European requirements. An in-depth review of the content and presentation of statistics on animal use by the U.K. Home office is in progress. The outcome and recommendations of the review should provide a useful reference for consideration by the EU.
References
Bayne, K. A. L., Dexter, S. L., & Strange, G. M. (1993). The effects of food treat and provisioning and human interaction on the behavioural well-being of rhesus monkeys (Macaca mulatta). Contemporary Topics in Laboratory Animal Science, 32, 6-9.
Bayne, K. (2002). Development of the human-research animal bond and its impact on animal well-being. ILAR Journal, 43, 4-9.
Commission of the European Communities (1994). First report from the Commission to the Council and the European Parliament on the statistics on the number of animals used for experimental and other scientific purposes in the member states of the European Union. Brussels: European Commission.
Commission of the European Communities (1999). Second report from the Commission to the Council and the European Parliament on the statistics on the number of animals used for experimental and other scientific purposes in the member states of the European Union. Brussels: European Commission.
Commission of the European Communities (2003). Third report from the Commission to the Council and the European Parliament on the statistics on the number of animals used for experimental and other scientific purposes in the members states of the European Union. Brussels: European Commission.
Council of Europe (2002). Statistical data on the use of laboratory animals in Belgium 2001. Strasbourg.
Council of Europe (2002). Statistical data on the use of laboratory animals in the Czech Republic 2001. Strasbourg.
Council of Europe (2002). Statistical data on the use of laboratory animals in Denmark 2001. Strasbourg.
Council of Europe (2002). Statistical data on the use of laboratory animals in Finland 2001. Strasbourg.
Council of Europe (2002). Statistical data on the use of laboratory animals in France 2001. Strasbourg.
Council of Europe (2002). Statistical data on the use of laboratory animals in Greece 1999, 2001 and 2002. Strasbourg.
Council of Europe (2002). Statistical data on the use of laboratory animals in the Netherlands 2001. Strasbourg.
Council of Europe. 2002. Statistical data on the use of laboratory animals in Sweden 2001. Strasbourg.
Council of Europe (2002). Statistical data on the use of laboratory animals in Switzerland 2001. Strasbourg.
European Commission (1986). European Convention for the protection of vertebrate animals used for experimental and other scientific purposes. ETS 123. Strasbourg. <conventions.coe.int/treaty/en/ treaties/html/123.htm>
European Commission (1992). Multilateral consultation of parties to the European Convention for the protection of vertebrate animals used in experimental and other scientific procedures (ETS 123). Resolution on the interpretation of certain provisions and terms of the Convention.
European Union (1986). Council Directive 86/609/EEC. Paris, France.
Federal Ministry of Consumer Protection, Nutrition and Agriculture (2003). Number of laboratory animals and purposes of experiments for 2001. Germany.
Fentener van Vlissingen, J. M. (1999). The re-use of animals for research: A humane endpoint. In C. F. M. Hendriksen and D. B. Morton (Eds.), Humane endpoints in animal experiments for biomedical research: Proceedings of the international conference, 22-25 November 1998, Zeist, the Netherlands. (pp. 145-147) London: Royal Society of Medicine Press.
Home Office (1992). Statistics of scientific procedures on living animals Great Britain 1991. HMSO, U.K.
Home Office (1993). Statistics of scientific procedures on living animals Great Britain 1992. HMSO, U.K.
Home Office (1994). Statistics of scientific procedures on living animals Great Britain 1993. HMSO, U.K.
Home Office (1995). Statistics of scientific procedures on living animals Great Britain 1994. HMSO, U.K.
Home Office (1996). Statistics of scientific procedures on living animals Great Britain 1995. HMSO, U.K.
Home Office (1997). Statistics of scientific procedures on living animals Great Britain 1996. HMSO, U.K.
Home Office (1998). Statistics of scientific procedures on living animals Great Britain 1997. HMSO, U.K.
Home Office (1999). Statistics of scientific procedures on living animals Great Britain 1998. HMSO, U.K.
Home Office (2000). Statistics of scientific procedures on living animals Great Britain 1999. HMSO, U.K.
Home Office (2001). Statistics of scientific procedures on living animals Great Britain 2000. HMSO, U.K.
Home Office (2002). Statistics of scientific procedures on living animals Great Britain 2001. HMSO, U.K.
Home Office (2003). Statistics of scientific procedures on living animals Great Britain 2002. HMSO, U.K.
Poole, T., Hubrecht, R., & Kirkwood, J. K. (1999). Marmosets and tamarins. In T. Poole (Ed.), The UFAW handbook on the care and management of laboratory animals (pp. 559-574). Oxon: Blackwell Science Ltd.
Prescott, M. J., & Buchanan-Smith, H. M. (2003). Training nonhuman primates using positive reinforcement techniques. Journal of Applied Animal Welfare Science, 6, 157-162.
Reinhardt, V., Liss, C., & Stevens, C. (1995). Restraint methods of laboratory non-human primates: A critical review. Animal Welfare, 4, 221-238.
Wolfensohn, S. E. (1997). Brief review of scientific studies of the welfare implications of transporting primates. Lab Animal, 31, 303-305.
* * *
Grants Available: Aging Dissertation Awards to Increase Diversity
The National Institute of Aging (NIA) announces the availability of dissertation awards (R36) in all areas of research within the Institute�s mandate to increase the diversity of the research workforce on aging. These awards are available to qualified predoctoral students in accredited research doctoral programs in the United States (including Puerto Rico and other U.S. territories or possessions). NIA expects to award $250,000 to $300,000 annually beginning in FY 2006 to support 6 to 8 dissertation awards. You may submit an application if your organization has the following characteristics:
Complete information is available at <grants.nih.gov/grants/guide/pa-files/PAR-05-061.html>, and the application form is at <grants.nih.gov/grants/funding/phs398/phs398.html>.
* * *
Forage Box as Enrichment in Single- and Group-Housed Callitrichid Monkeys
A. S. Chamove and L. Scott
Massey University Psychology Department and Chemical & Biological Defence Establishment
Introduction
When different enrichment techniques have been compared, foraging consistently is preferred by animals to other activities such as object manipulation (Bryant et al., 1988; Line & Morgan, 1991; Harper, 2001), leads to greater improvement in behavior (Chamove, 1989; Pyle et al., 1996), and is more cost-effective (Chamove et al., 1982; Chamove, 2001). The behavioral improvement is characteristically a reduction in abnormal and aggressive patterns, and an increase in species-typical forms. Perhaps this is because foraging takes up so much of a monkey�s time in the wild (Garber, 1980b; Terborgh, 1983).
Most published studies of foraging in nonhuman primates have dealt with socially housed animals (Chamove, 1989), many of which were in zoos (Fajzi et al., 1989; Harper, 2001). It can be argued that individually housed monkeys are in more need of enrichment since they lack partners with which to interact (Brinkman, 1996).
Unfortunately, the approach, recommended by some (Chamove et al., 1982), of using woodchip litter as a floor covering is impractical for situations such as individual caging with mesh floors. Researchers have demonstrated that foraging can be successfully scaled down for socially-housed (Boccia, 1989; Rosenblum & Smiley, 1984) or individually-housed (Bryant et al., 1988; Bayne et al., 1991, 1992; Murchinson, 1994) terrestrial monkeys maintained in laboratory conditions. A number of alternative approaches and a range of substrata in which to forage have been tested. These include foraging/grooming boards (Bayne et al., 1991), artificial turf (Bayne et al., 1992), and puzzle balls (Crockett et al., 2001).
The present study was designed to investigate the propensity of two arboreal callitrichid species, the common marmoset (Callithrix jacchus) and the cotton-top tamarin (Saguinus oedipus), for foraging. A forage-box task was chosen for the evaluation because of the results of a pilot study by the senior author with group-housed cotton-top tamarins, which showed that this device occupied the animals for more time than hole searching (see also Birke, 1988), retrieval of hanging food, ground foraging, string pulling, or using a play wheel. Food-irrelevant objects appear to have little effect on behavior of marmosets and tamarins (Chamove & Anderson, 1989; Renner et al., 2000). The goal was to reach an enrichment level of foraging comparable to the levels of Callithrix jacchus (Rylands & de Faria, 1993), Saguinus imperator (Terborgh, 1983), S. oedipus geoffroyi (Garber, 1980b), and S. fuscicollis (Yoneda, 1984) reported in the wild: namely time budgets of daily activity of 55%, 46%, 34%, and 32% respectively. This report summarizes the results of two related studies, describing investigations using a simple Perspex forage box for singly-housed marmosets and group-living marmosets and tamarins.
Foraging appears to be motivated by factors other than merely food: monkeys, like various other animals (e.g., starlings [Englis & Ferguson, 1985]), will continue to forage for food even when the same food items are freely available. Nevertheless they will forage over twice as much when food-motivated. They will also forage in substrata where there is no food at all, although only about one-third as much as when they are able to find food (Anderson & Chamove, 1984). Without using food deprivation, it is easy to increase food motivation by using desirable items that are not commonly available to the animals. Food preferences were assessed prior to testing the foraging apparatus.
Study 1: Individually Housed Marmosets
Subjects: Food preference was assessed for eight individually housed common marmosets, four of each sex. The animals were captive bred, with ages ranging from 15 to 27 months. They had been individually housed for at least 6 months and lived in .22 m3 cages (75 x 48 x 60 cm) in a colony room where they could see other animals at about 1 m distance. Temperature was 22o-23oC, humidity 50-60%, and artificial lighting was on for 12 hours. Animals were normally fed at 1600 h on a primate diet (Labsure CPD X), supplemented with fruit and vitamins. Housing details can be found in Scott (1991). All testing of marmosets occurred between 1000 and 1500 h.
Each monkey was offered 3-10 items from a list of eight foods. Each food type was given on a different day and presented by itself in a metal bowl similar to those in which food was normally given. The test foods were Tesco malt loaf, Rice Krispies, fresh apple, acacia gum, sultana raisins, Bioserv brand 45-mg banana pellets, millet, and live meal worms. We measured latency to first contact, duration of first contact, number of contacts, number eaten, and total contact time with the food items.
From total contact time, and confirmed by the other measures, the above order was the average order of preference for the eight foods, malt loaf being most preferred and meal worms the least. So, in all tests with marmosets below, food items were used in the following pattern: 4 pieces of malt loaf, 10 Rice Krispies, 5 pieces of apple, 2 piece of Acacia gum, 5 sultanas, and 3 meal worms. Except for mealworms, food items were about the same size.
Apparatus: The forage box, shown in Figure 1, was constructed of clear 0.5 cm-thick Perspex-brand plastic sheeting, and was 29 cm long, 13 cm wide and 12.5 cm deep. A false bottom was inserted to produce a functional depth of 8 cm so that animals could reach to the bottom if a lid were in place. A barred wire top with 1.25 cm-wide openings could be fitted to the top; and a grid floor could also be inserted over the base within the box. This floor grid had 36 2.7-cm2 honeycomb sections 5 cm deep made of 5 mm thick clear plastic. It was anticipated that the barred lid and Perspex floor grid would increase the difficulty of the task in that they would prohibit head and whole body from entering the box, encourage hand-only foraging, and thus increase the duration of this activity. In previous foraging tasks monkeys typically find and eat all the food rather quickly, commonly within the first hour, leaving nothing to find for the remainder of the day.
Figure 1: Three tamarins using the Perspex forage box.
Procedure-Box Design: To evaluate different components of the forage box apparatus, a Latin square design was used to present four conditions to the monkeys. The conditions were as follows: the forage box filled with sawdust and (1) no floor grid, no top lid; (2) no grid but the lid present; (3) grid in place but no lid; (4) both grid and lid in place. Animals were given a single 1-hr presentation of each of the four arrangements on different days. An 8 cm-deep layer of sawdust was placed in the box and food items were buried at varying depths beneath the surface.
�Foraging� was defined as active searching through the substrate or physical contact with the forage box.
Results: The plain forage box (no-lid/no-grid) was the easiest and also the most effective condition for the monkeys, individuals spending an average of 63% of the hour foraging. This was reduced by about 2% in the presence of a lid, by about 6% with the floor grid inserted, and by 19% when both the lid and grid were in place.
The monkeys averaged just over one approach to the box per minute per animal during the hour test and the contacts averaged about 46 sec when neither grid nor lid was used. Both this rate of approach and contact time were lower when both grid and lid were in place. Forage-related hand movements averaged about 7 per min in both the box alone and box + grid conditions. This decreased to an average of about 5 per minute in the box + lid and to about 2.5 per minute for the box + lid + grid.
The purpose of the lid and grid was to prohibit head and whole body entries into the box, to encourage hand-only foraging, to make the task more difficult, and thereby increase the total duration of foraging. Forage-related hand movements averaged 420 per animal per hour with no lid or grid, but actually decreased significantly by 60% to 167/animal/hr with both lid and grid in place. Thus the lid and grid did not have the desired effect of increasing the duration of foraging by making foraging more difficult and reducing the probability of success. Instead their presence decreased the duration of foraging. The plain forage box was used thereafter in these studies.
Procedure-First Hour: To assess the stability of foraging behavior over the first hour of the day, the eight marmosets were retested with the forage box presented without its grid or lid. These animals had developed stable patterns of foraging over a period of weeks and we expected no changes in foraging over subsequent days. Each monkey was tested on 13 separate days, and each test was 60 min. long.
Figure 2: Mean percent time spent foraging through sawdust in a forage box (no floor grid, no top lid) during 1 hour for 7 singly housed marmosets from a group of 8, and the individual of the 8 that foraged most (top line) and the one that foraged second least (bottom line) of the group of 8. One monkey of the group of 8 is not included in this figure for reasons detailed in the text.
Results: Figure 2 shows the mean foraging rate over the hour-long test for seven of the eight animals and the performance of two extreme animals of that seven. Seven of the 8 animals showed similar patterns of foraging over time, the most active of these 7 showing an asymptote of foraging around 75% and the least active of the 7 around 35% (Figure 2). One animal showed an unusual pattern, clearly different from all the others, steady at about 13% throughout the hour, and consequently is not shown in the figure. As illustrated, there were two clear reductions in foraging, the first around 20 min. and a second around 45 min.; both of these fluctuations were shown by all 7 of the animals referred to above, although all were tested independently and on different days. Most animals were foraging for about 50% of the time during the second half of the hour. This is slightly below the findings of Dettling and Pryce (1998) for a group of the same species, discussed more fully below.
Procedure-Over Days: Four animals, two of each sex, were monitored for 4-hour periods over 15 days to evaluate the stability of foraging behavior over a longer time period. Monkeys were observed once at points 1, 2, 3, and 4 hours after the forage box was put into the home cage, and a record made of whether they were foraging at each point.
Results: Results showed that the pattern of foraging was similar for the full four hours to that seen during detailed observation during the first hour. That is, over 50% of the animals were observed actively foraging at the 4-hour mark. While values after 1 hour were similar to those after 4 hours, the values after 2 hours and after 3 hours were slightly lower, when just over 40% of the animals were foraging at each of these two point samples.
This compares with an extensive sample of behavior taken by the senior author of 11 individually housed marmosets housed in standard single cages containing a nest box and a wooden perch. Animals were fed protein around 1230 h, fruit around 1600 h, and cleaning took place around 1400 h. Three categories of behavior were recorded by point sampling 5 times per hour between 0900 and 1600 to give 35 samples per animal per day at approximate 12-minute intervals, spread over a two-month period, for 20 days of testing which excluded weekends, giving 700 samples per animal. Location was divided into: on floor, front mesh surface, perch, or inside nest box. Behavior was divided into groom, huddle, play, abnormal, feed/drink, gnaw. Activity was divided into inactive, looking, sit but active, walk, or run (defined in Millar et al., 1988).
In such a refined environment, the predominant activity is just looking (65%). The animals are usually on the perch or door (83%), rarely (15%) on the cage floor. The monkeys rarely engage in active behavior such as walking (10%) or running (3%), and other activities are not common (feeding [7%] and grooming [4%]). Abnormal behavior is seen in 2% of observations. These behavior patterns are similar to those seen in group-housed marmosets (Millar et al., 1988), except that in family groups huddling is over 10 times more common and mostly directed towards other animals rather than self-directed. Abnormal behavior was never seen (see also Maier et al., 1982), and only 36% of the time was spent inactive or looking.
It appears that our individually housed marmosets are more active when they have access to a forage box, replacing inactivity with foraging. Group-housed marmosets (Dettling & Pryce, 1998) do not replace inactivity with foraging, rather replacing active behaviors with foraging when given the chance to do so.
Study 2: Group-Housed Marmosets and Tamarins
Subjects: Four family groups of cotton-top tamarins, composed of from 5 to 11 (total of 35) individuals, were used to observe foraging in a group situation. Two family groups of common marmosets, composed of 6 or 7 individuals, were also observed. The animals lived in 56 m3 rooms (3.5 x 4 x 4 m), with wooden branches, ropes, wooden and mesh platforms, woodchips covering the floors, and many objects (details in Price & McGrew, 1990). Animals were captive bred and their ages ranged from 0.5 to 2 years except for the breeding pairs, who were much older.
Procedure: During testing, the animals received their normal rations of food: high protein foods at 0945 h, mixed fruit at 1200 h, and apple at 1615 h. An additional 10 gm of food per monkey was buried in woodchips in the forage box. The food consisted of Farley�s Rusk, grapes, peanuts, and raisins presented in approximately equal amounts by volume. All these are foods that these animals particularly liked and rarely received.
After extensive habituation to the box with food buried in chips, it was presented (see Figure 1) on two consecutive days at 1000 h during which data were collected for 6 hours, using a video camera directed at the box. To analyze the data, the video was replayed on the �fast forward� setting. Point sampling every 7 sec corresponded to samples every 60 sec in real time. The number of animals foraging in the box was noted at these 1 min points.
Results: As marmosets and tamarins did not differ in any behavior analyzed, their data were combined. The animals showed two peaks of foraging in the box, the first during the first hour (5.8 visits per animal per hr) and the second peak during the fifth hour (5.1); and two lows during the third hour (2.8) and the sixth hour (3.6). This pattern was similar to that shown by individually housed marmosets, who showed peaks during the first and fourth hours, and compares with behavior of wild tamarins (Terborgh, 1983; Yoneda, 1984), in which there are two similar peaks of foraging, one during the first hour after waking (700 h) and one at 1500 h; there are also two lows, one during the third or fourth hours and another at 1400 h.
Foraging during the first hour averaged 10.4% of scans. This value is much lower than the 75% reported during the first half hour for a group of common marmosets housed in a smaller (5 m3) cage and offered plant pots with freshly killed crickets and groundnuts mixed into sawdust and suspended from the ceiling (Dettling & Pryce, 1998). The 75% also includes foraging for food in sawdust on the ground, which, when presented without the elevated pots, averaged about 15%. Even subtracting 15% still leaves a remarkable 60% of the time foraging in the first half hour for their group-housed marmosets. Their rate is comparable to the 55% in our individually-housed marmosets.
Over our 6 hours of observation, any given monkey was observed foraging on 7.4% of scans. We observed 4.4 visits to the box per subject per hour, a total of about 26 visits each. We can compare this with normal feeding patterns in the laboratory situation during the remainder of the daytime but after the first feed of the day. In this post-feeding period the monkeys spent 1.9% of the day or paid 1.1 visits over the 6-hr period to the food tray. We can also look at the total time normally spent feeding, but including the first feed of the day plus any visits to the food tray during the following 6 hours. When this is computed, the normal or baseline daily forage pattern rises to 3.3% or 2.3 visits from just under 2% of time and just over 1 visit to the food. So foraging time is doubled over all baseline feeding time by using the forage box, or trebled if the comparison is only with the time spent feeding after the initial and rather invariable hour of breakfast feeding. This 7% at the forage box compares to 12% of time in the wild spent foraging on fruit, 4% on vegetable matter, and 16% on insect matter in other tamarin species (Garber, 1980a, 1980b; Terborgh, 1983; Yoneda, 1984), a grand total of 32% of the day foraging under wild conditions.
Discussion
This pair of studies shows that a small forage box will encourage foraging in singly housed marmosets living in cages that must be impoverished environments. The marmosets foraged over 60% of the time during the first hour and about 40% of the rest of the day in these boxes. Marmosets and tamarins living in large family groups in less impoverished environments increased their foraging from a baseline of 2% by an additional 7% over the day. While this is not an impressive increase for the callitrichid family groups, the increase to 60% of the day is substantial for the individually housed marmosets. One goal of enrichment is to provide activities that match those found in the wild. This increase in group foraging does not compare favorably with a foraging rate of 30% in captive group-housed terrestrial macaques given grain in woodchips or with 50% for individually housed macaques given artificial turf (Bayne et al., 1992). It is, however, comparable to studies giving arboreal callitrichids foraging opportunities at floor level (Chamove et al., 1982).
When presented with food, common marmosets will more often take it directly into their mouths than use their hands. This might explain why in the conditions where a lid was used, there were fewer forage-related hand movements than might have been expected. Making extraction more difficult, by forcing them to forage with their hands through the openings of a lid, does not increase their foraging time, suggesting that it is the movements, rather than the task, that control the motivation.
Foraging through a floor grid seems to have unpredictable consequences. The provision of a lid and floor grid to our box reduced foraging by almost 20%, but did not eliminate it. Marmosets readily (over 60% of time) foraged through the floor of the extension we provided. Macaques also readily forage through floor mesh (5 cm2 mesh) but not through vertical front bars (5 cm) into an outside bin (Brinkman, 1996). Murchinson (1994) found that young pig-tail macaques increased their feeding from 1% to 11% of the first hour when he reduced the size of holes in standard chow feeder boxes from 5 to 3 cm.
Although it is now standard practice to house monkeys in compatible pairs or groups, there are occasions when single housing for at least part of the day is essential to the experimental protocol or therapeutic regime, or required to alleviate social incompatibilities. In such cases, cage extensions with provision for foraging can be used, and are now routinely used in the laboratory of the junior author.
References
Anderson, J. R., & Chamove, A. S. (1984). Allowing captive primates to forage. In Standards in laboratory animal management (Vol. 2, pp. 253-256). Potters Bar: Universities Federation for Animal Welfare.
Bayne, K., Mainzer, H., Dexter, S., Campbell, G., & Yamada, F. (1991). Reduction of abnormal behaviors in individually housed rhesus monkeys (Macaca mulatta) with a foraging/grooming board. American Journal of Primatology, 23, 23-33.
Bayne, K., Dexter, S., Mainzer, H., McCully, C., Campbell, G., & Yamada, F. (1992). The use of artificial turf as a foraging substrate for individually housed rhesus monkeys (Macaca mulatta). Animal Welfare, 1, 39-54.
Birke, L. (1988). Better homes for laboratory animals. New Scientist, 120, 56-60.
Boccia, M. L. (1989). Long-term effects of a natural foraging task on aggression and stereotypies in socially housed pigtail macaques. Laboratory Primate Newsletter, 28[2], 18-19.
Brinkman, C. (1996). Toys for the boys: Environmental enrichment for singly housed adult male macaques (Macaca fascicularis). Laboratory Primate Newsletter, 35[2], 4-9.
Bryant, C. E., Rupniak, N. M. J., & Iversen, S. D. (1988). Effects of different environmental enrichment devices on cage stereotypies and autoaggression in captive cynomolgus monkeys. Journal of Medical Primatology, 17, 257-267.
Chamove, A. S. (2001). Floor-covering research benefits primates. Australasian Primatology, 14, 16-19.
Chamove, A. S. (1981). Establishment of a breeding colony of stumptailed monkeys. Laboratory Animals, 15, 251-259.
Chamove, A. S. (1989). Environmental enrichment: A review. Animal Technology, 40, 155-178.
Chamove, A. S., & Anderson, J. R. (1989). Examining environmental enrichment. In E. Segal (Ed.), The psychological well-being of primates (pp. 183-202). Philadelphia: Noyes Publications.
Chamove, A. S., Anderson, J. R., Morgan-Jones, S. C., & Jones, S. P. (1982). Deep woodchip litter: Hygiene, feeding, and behavioral enhancement in eight primate species. International Journal for the Study of Animal Problems, 3, 308-318.
Crockett, C. M., Bellanca, R. U., Heffernan, K. S., Ronan, D. A., & Bonn, W. F. (2001). Puzzle Ball foraging device for laboratory monkeys. Laboratory Primate Newsletter, 40[1], 4-7.
Dettling, A., & Pryce, C. R. (1998). Physical environment and its influence on behaviour in captive common marmosets (Callithrix jacchus). In European Marmoset Research Group (Eds.), Marmosets: A handbook for fundamental and applied research. New York: Academic Press.
Fajzi, K., Reinhardt, V., & Smith, M. D. (1989). A review of environmental enrichment strategies for singly caged nonhuman primates. Laboratory Animal, 18, 23-35.
Garber, P. A. (1980a). Locomotor behavior and feeding ecology of the Panamanian tamarin (Saguinus oedipus geoffroyi, callitrichidae, primates). University Microfilms International No. 81-03, 681.
Garber, P. A. (1980b). Locomotor behavior and feeding ecology of the Panamanian tamarin (Saguinus oedipus geoffroyi, Callitrichidae, Primates). International Journal of Primatology, 1, 185-201.
Harper, P. (2001). Eight years of environmental enrichment for Adelaide Zoo�s adult male orangutan. Australasian Primatology, 15, 15-23.
Inglis I. R., & Ferguson, N. J. K. (1985). Starlings search for food rather than eat freely available identical food. Animal Behaviour, 34, 614-616.
Line, S. W., & Morgan, K. N. (1991). The effects of two novel objects on the behavior of singly caged adult rhesus monkeys. Laboratory Animal Science, 41, 365-399.
Maier, W., Alonso, C., & Langguth, A. (1982). Field observations on Callithrix jacchus jacchus L. Zeitschrift f�r S�ugetierkunde, 47, 334-346.
Millar, S. K., Evans, S., & Chamove, A. S.. (1988). Older offspring contact novel objects soonest in callitrichid families. Biology of Behavior, 13, 82-96.
Murchinson, M. (1994). Forage feeder box for single animal cages. Laboratory Primate Newsletter, 33[1], 1-3.
Price, E. C., & McGrew, W. C. (1990). Cotton-top tamarins (Saguinus (o.) oedipus) in a semi-naturalistic captive colony. American Journal of Primatology, 20, 1-12.
Pyle, D. A., Bennett, A. L., Zarcone, T. J., Turkkan, J. S., Adams, R. J., & Hienz, R. D. (1996). Use of two food foraging devices by singly housed baboons. Laboratory Primate Newsletter, 35[2], 10-14.
Renner, M. J., Feiner, A. J., & Orr, M. G. (2000). Environmental enrichment for New World primates: Introducing food-irrelevant objects and direct and secondary effects. Journal of Applied Animal Welfare Science, 3, 23-32.
Rosenblum, L. A., & Smiley, J. (1984). Therapeutic effects of an imposed foraging task in disturbed monkeys. Journal of Child Psychology & Psychiatry & Allied Disciplines, 25, 485-497.
Rylands, A. B., & de Faria, D. S. (1993). Habitats, feeding ecology, and home range size in the genus Callithrix. In A. B. Rylands (Ed.), Marmosets and tamarins: Systematics, behaviour, and ecology (pp. 262-271). Oxford: Oxford University Press.
Scott, L. (1991). Environmental enrichment for single housed common marmosets. In H. O. Box (Ed.), Primate responses to environmental change (pp. 265-275). London: Chapman & Hall.
Terborgh, J. (1983). Five New World primates: A study in comparative ecology. Princeton: Princeton University Press.
Yoneda, M. (1984). Ecological study of the saddle backed tamarin (Saguinus fuscicollis) in northern Bolivia. Primates, 25, 1-12.
* * *
News Briefs
Perforation Caused Orangutan�s Death
Final medical tests found that a terminal condition that led to the death of Indah, a Des Moines orangutan, was caused by a perforation in her bladder. Tests by veterinarians from the Great Ape Trust of Iowa and Iowa State University (ISU) veterinarians and private colleagues confirmed that Indah�s bladder had been damaged �many months� before she was transferred to Des Moines from the National Zoo, the Great Ape Trust of Iowa reported. Infection from the perforation and intestinal damage discovered during surgery led veterinarians to euthanize Indah on November 11. She was taken to ISU originally because she was severely constipated.
A physical examination, including X-rays, before Indah was transported from the National Zoo to the Des Moines research center, didn�t show the problems, which are difficult to view on screens, veterinarians said. The illnesses were not connected to Indah�s care at the Des Moines facility, they added.
Indah�s brother, Azy, is the sole occupant of the Des Moines facility, which is searching for more orangutans and plans to add bonobos in the spring. Eventually, it will also have gorillas and chimpanzees. The Great Ape Trust of Iowa, located in extreme southeast Des Moines, is conducting language and cognitive research on great apes. - Des Moines Register, December 23, 2004
New Orangutan Moves to Great Ape Trust
Azy the orangutan is getting a new friend at Great Ape Trust of Iowa (see <www.greatapetrust.org>). Knobi, a 25-year-old female orangutan, is scheduled to arrive at the research center in Des Moines today. Robert Shumaker, the director of the orangutan project, is picking her up from the Henry Doorly Zoo in Omaha, Nebraska, where she was born. Shumaker says Knobi will be a wonderful companion for Azy and an ideal candidate for their research into the intelligence and behavior of great apes.
Azy and his sister, Indah, were the first two apes at the research center. They arrived last September from the National Zoo in Washington, DC. Indah was euthanized in November after becoming ill. The search for Azy�s companion started, and they found Knobi. She�ll be in quarantine for 30 days. - Associated Press, February 8
Bronx Zoo to Close Island Preserve
The Bronx Zoo is closing a large animal preserve on a 14,000-acre undeveloped island off the coast of Georgia, where for 30 years zoologists have studied - among other things - the mating habits of wildlife, including lemurs, hartebeests, zebras, tortoises, gazelles and several species of exotic birds.
The preserve, known as the Wildlife Survival Center, is on St. Catherine�s Island, 12 miles long and 3 miles wide, about 50 miles south of Savannah. Alison Power, a spokeswoman for the Wildlife Conservation Society, which oversees the zoo, said the society has run the preserve since 1974 but has decided to close it because many of its �original objectives have been met.�
The preserve was established so scientists could study several species of birds and mammals in a habitat as close to wilderness as possible, Ms. Power said. She added that the island�s vast spaces, its large enclosures and its private grounds, to which no visitors are allowed, permitted zoologists to study how animals would act in �herd situations� or determine what would happen if captive beasts were reintroduced to the wilderness.
Through work at the island, scientists working with the zoo �learned invaluable information on how to maintain large herds of hoof stock,� Ms. Power said, as well as �how to create breeding groups to improve chances for successful breeding.�
Richard L. Lattis, the general director of living institutions for the Wildlife Conservation Society, said in a statement released yesterday: �For 30 years, studies at St. Catherine�s Island have helped us break new ground in the care and conservation of important species. The original program goals have been met.�
Ms. Power said the zoo would spend as much as a year relocating the 435 animals of 46 species from the island preserve to other zoos and wildlife parks around the country. Some of the animals will come to the Bronx, she said, particularly a group of lemurs, which will be moved into a special exhibit on the wildlife of Madagascar that is scheduled to open in the Bronx in 2006. - New York Times, December 29, 2004
New Monkey Discovered In Northeastern India
A species of monkey previously unknown to science has been discovered in the remote northeastern region of India. Named after the Indian state of Arunachal Pradesh where it was found, the Arunachal macaque (Macaca munzala) - a relatively large brown primate with a comparatively short tail - is described in a forthcoming issue of the International Journal of Primatology. The last species of macaque to be discovered in the wild, the Indonesian Pagai macaque, was described in 1903.
�This new species comes from a biologically rich area that is perhaps India�s last unknown frontier,� said Wildlife Conservation Society conservation scientist Dr. M. D. Madhusudan, who was part of the discovery team. �The discovery of a new species of monkey is quite rare. What is also remarkable about our discovery is that few would have thought that with over a billion people and retreating wild lands, a new large mammal species would ever be found in India, of all places.� The new species is one of the highest-dwelling primates in the world, occurring between 1600 and 3500 meters above sea level.
Although the monkey is new to science, the animal is well known to the residents of the Himalayan districts of Tawang and West Kameng, where the species occurs. The monkey�s species name, mun zala, means �deep-forest monkey� in the vernacular of the Dirang Monpa people. - Wildlife Conservation Society news release, December
Right to Name New Monkey Sells for $650G
The right to name a new species of titi monkey sold Thursday for $650,000 in an online auction, with the funds going to protect the Bolivian habitat where the species lives. The winning bidder chose to remain anonymous for now.
The monkey - which is about a foot tall and has a brown body, a golden crown, orange cheeks and a white-tipped tail - was discovered by a Wildlife Conservation Society scientist in Madidi National Park in Bolivia last year, the Conservation Society said. - Associated Press, March 4
Gorillas Cross to Rwanda
Gorilla tracking in Mgahinga National Park in south-western Uganda has been suspended after a group of habituated gorillas in the park crossed to Rwanda. The gorillas crossed into Rwanda about two months ago and wildlife authorities are puzzled at the apes� delayed return. Sources at the Uganda Wildlife Authority (UWA) suspect that the gorillas could have fled their wild counterparts, which could have invaded their territory. The gorillas had been taken through a delicate process to make them used to human presence in their habitat in a bid to encourage gorilla tourism. The sources said the apes usually cross the borders for a week or two and come back, but this time they have taken three months without returning.
Damian Akankwasa, the director of tourism, said UWA was concerned that the apes had overstayed across the border. �We pray that they come back because we have suspended gorilla viewing in the park,� he said, adding that UWA was losing revenue. Under the revised rates, UWA charges $360 per person to track gorillas in Bwindi and Mgahinga for foreign tourists and a maximum of six people track gorillas daily. Gorilla trackers destined to Mgahinga are being diverted to other habituated groups of gorillas in Bwindi Impenetrable Forest National Park.
Bwindi contains half of the 670 global population of mountain gorillas. - January 5, 2005, Gerald Tenywa, Kampala, reported on primate-net
Sylvia Taylor
Dr. Sylvia Taylor, a veterinarian with the USDA Animal Care Division, died suddenly January 6, 2005, in Tampa, Florida. Sylvia was dedicated in her quest to provide the best captive care environment for all species of animals, especially nonhuman primates. She had the good fortune to realize her dream of traveling all over the world in a professional capacity as well as to pursue personal interests. Sylvia was there to aid in the epidemic of foot and mouth disease in the United Kingdom, Exotic Newcastles Disease in California, and Avian Influenza in Virginia. She supported sanctuaries for many types of animals throughout the world and was a superb networker - she was a knowledgeable resource and would direct you to the right person with a particular expertise. - posted to CompMed by Elysse A. Orchard, DVM
Primate Research Center for HIV/AIDS Opened
India�s first primate research center will address the needs of scientific research mainly in the areas of HIV/AIDS and stem cell biology, Union Health and Family Welfare minister Anbumani Ramadoss today said. Speaking after laying the foundation stone for the Center in Vassai, Ramadoss said the National Centre for Primate Breeding and Research would be supported by the government without any fund constraints.
The U.S. Office of AIDS Research of the National Institute of Health (NIH) has given three million U.S. dollars in the form of equipment and other accessories for the Centre under an Indo-U.S. research program.
Ramadoss said the primate species which will be used at the Centre would serve as good experimental models for research if a humane approach is adopted while handling them. �We should, therefore, ensure their well-being and make all efforts to minimize the pain and distress to them during the experimentation,� he said. Scientists also have to ensure that the animals are used for experimentation only when no alternative models are available and that the number of animals used are restricted to the minimum, he said. With creation of the facility, scientists would be able to address health-related problems effectively.
Speaking on the occasion Jack Whites Tarver, Director of the Office of AIDS Research, NIH, said the primate center would open up many more opportunities between India and the U.S. His country would provide additional help if required. - from <www.newkerala.com>, January 9, 2005
Proposed Standards of Care for Chimp Sanctuaries
In the January 11 edition of the Federal Register <www.gpoaccess.gov/fr>, the National Institutes of Health published a notice of proposed rulemaking on standards of care for chimpanzees held in the Federal Sanctuary System. These proposed standards have been issued in accordance with provisions of the Chimpanzee Health Improvement, Maintenance, and Protection Act (CHIMP Act) of 2000. The proposed regulations will only be applicable to facilities that receive federal funding as part of the federal chimpanzee sanctuary system.
Hollywood Apes Retiring to Sanctuary
Eleven chimpanzees and six orangutans owned by Los Angeles animal trainer Bob Dunn will be out of the entertainment business in March and headed to a life of permanent sanctuary retirement in Wauchula, Florida. Dunn, owner of Bob Dunn�s Animal Services, has for some 30 years been the main supplier of great apes for Hollywood and other entertainment, providing chimpanzees and orangutans to movies, television shows, and advertisements. Dunn is leaving the ape business and donating all his working and breeding apes to the Center for Great Apes, a private, nonprofit sanctuary.
Located on 100 acres of tropical forest and orange groves in central Florida, the Center for Great Apes was founded by Patti Ragan, who left a business career in order to provide long term care for orangutans and chimpanzees needing permanent care, the mission of the Center. With the assistance of the Arcus Foundation, the Center for Great Apes is building new habitats for the retired apes, who will move to Florida as soon as the facilities are ready. At that time, the Center will remove from Dunn�s property all cages, nighthouses, transfer crates, and other equipment used for these apes.
The sanctuary, which is supported by individual memberships and private donations, is not open to the public as an attraction. For more information see <www.CenterForGreatApes.org>.
Viet Nam Discovers Special Langur Monkeys
Scientists have discovered two groups of special langurs, long-tailed monkeys, which they call Trachypithecus permani, in Kien Luong distirct, Kien Giang province bordering Cambodia. The first group of 23 was found on the Bai Voi (Elephant Ground) limestone mountain and the second group, which has not yet been counted, was discovered on the nearby Chua Hang (Cave Pagoda) mountain.
Scientists are looking for ways to remove the monkeys for their protection as the Elephant Ground limestone mountain is to be mined for building materials soon. - from <www.vnanet.vn>, January 26
Guardian of Ghana�s Monkey Village Dies
Mr. Daniel Kwaku Akowuah, an eminent conservationist and caretaker of Buabeng-Fiema Monkey Sanctuary in the Nkoranza District, Ghana, died on December 19, after a short illness.
Akowuah, who was 70, is remembered for the pioneering role he played in the establishment of the Monkey Sanctuary as he single-handedly tamed the animals and brought them closer to humans. Mr. Akowuah, a retired teacher, was appointed the first Game Warden of the Monkey Sanctuary and through his untiring efforts helped it to attain international status as a viable tourist project. - posted to alloprimate
Kenya Seizes Smuggled Baby Chimps
Kenya made its biggest seizure of primates in the battle to stop trade in endangered wildlife with the discovery of six baby chimpanzees crammed into a crate at an airport, Kenya Wildlife Service (KWS) said. The chimps, with a black market value of $20,000 each, and four Guenons, a type of long-tailed African monkey, were discovered abandoned and hungry on January 31 but have since been taken into care by the KWS and given toys to play with.
�The six babies were squashed in a small crate together with four guenons, they were in a very pathetic condition,� Eric Kalla, an assistant director at the KWS told Reuters. �They had begun eating their own fecal matter due to hunger and unfortunately one baby chimp died due to stress and starvation,� he said. He said the animals, en route from Egypt to Nigeria via Kenya�s Jomo Kenyatta International Airport, belonged to a woman who had claimed the crate was a kennel containing dogs. - Reuters, Feb 15
The infants were permanently transferred to the Sweetwaters Chimpanzee Sanctuary near Mount Kenya on March 5. The guenons remain in the care of the Kenya Wildlife Service.
More Organisms Targeted for Genome Sequencing
The National Human Genome Research Institute (NHGRI) announced today that the Large-Scale Sequencing Research Network will begin sequencing 12 more strategically selected organisms.
Two of the sequencing projects are aimed at gaining new insights into model organisms utilized in research on drug development and disease susceptibility. They are: sequencing the genome of a fellow primate, the marmoset (Callithrix jacchus); and identification of genetic variations (in the form of single nucleotide polymorphisms) in eight strains of rats.
The marmoset is a key model organism used in neurobiological studies of multiple sclerosis, Parkinson�s disease, and Huntington�s disease. The marmoset is also an important model for research into infectious disease and pharmacology. The marmoset was chosen also because of its position on the evolutionary tree, further removed from humans than other nonhuman primates already being sequenced, such as the chimpanzee (Pan troglodytes), rhesus macaque (Macaca mulatta) and orangutan (Pongo pygmaeus). Obtaining the marmoset genome sequence will provide a powerful tool to illuminate the similarities and differences among these primate genomes. - NIH News Release, March 1, 2005
Wolff Director of Compliance Oversight, OLAW
Axel Wolff has been appointed as Director, Division of Compliance Oversight, Office of Laboratory Animal Welfare (OLAW), Office of Extramural Research, NIH. Dr. Wolff has been serving in an acting capacity since May, 2004, prior to which he was a Senior Assurance Officer in OLAW�s Division of Assurances. Dr. Wolff graduated from the University of Missouri-Columbia�s College of Veterinary Medicine and holds a Master�s Degree in Zoology from the University of Wisconsin-Milwaukee for work conducted with tropical fruit bats.
NCRR Associate Director of Comparative Medicine
Franziska Grieder has been appointed Associate Director of the National Center for Research Resources� Division of Comparative Medicine (DCM), which supports programs and resources for advancing biomedical research through the use of nonhuman models. DCM-funded resources include the eight National Primate Research Centers and five Mutant Mouse Regional Resource Centers. Since 2000, Grieder has managed DCM�s Laboratory Animal Sciences Program. Since 1993, Grieder has been on the faculty at the Medical School of the Uniformed Services University of the Health Sciences. - From the NCRR Reporter, Fall, 2004
Chimps Escape, Attack Visitors at Animal Sanctuary
BAKERSFIELD, Calif. Mar 4, 2005 - A couple�s plans for a birthday party for their former pet chimpanzee turned tragic when two other chimps at an animal
sanctuary escaped from their cage and attacked. The man was critically injured with massive wounds to his face, body and limbs, and the attacking animals were shot dead by the son-in-law of the sanctuary�s owner.
St. James and LaDonna Davis were at the Animal Haven Ranch in Caliente to celebrate the birthday of Moe, a 39-year-old chimpanzee who was taken from their suburban Los Angeles home in 1999 after biting off part of a woman�s finger. Moe was not involved in Thursday�s attack, said Steve Martarano, a spokesman for the California Department of Fish and Game.
The couple had brought Moe a cake and were standing outside his cage when Buddy and Ollie, two of four chimpanzees in the adjoining cage, attacked St. James Davis, Martarano said. Officials have not determined how the chimps got out of their enclosure, he said. LaDonna Davis, 64, suffered a bite wound to the hand while trying to help her 62-year-old husband, Martarano said.
St. James Davis had severe facial injuries and would require extensive surgery in an attempt to reattach his nose, Dr. Maureen Martin of Kern Medical Center told KGET-TV of Bakersfield. His testicles and a foot also were severed, Kern County Sheriff�s Cmdr. Hal Chealander told The Bakersfield Californian.
Buddy, a 16-year-old male chimp, initiated the attack and after he was shot, Ollie, a 13-year-old male, grabbed the gravely injured man and dragged him down the road, authorities said.
Two other chimps, females named Susie and Bones, also escaped from the cage they shared with Ollie and Buddy, prompting sheriff�s deputies, animal control workers, and Fish and Game officials to launch a search. - Copyright 2005 The Associated Press.
* * *
Announcements from Publications
Call For Papers - JAAWS
The Journal of Applied Animal Welfare Science (JAAWS) is seeking manuscripts for publication. Conceived to promote the emerging field of animal welfare science, the goal of JAAWS is to publish articles and reports on methods of experimentation, husbandry, and care that demonstrably enhance the welfare of animals. Abstracts may be read at <www.societyandanimalsforum.org>, under �publications�.
A coproject of the American Society for the Prevention of Cruelty to Animals and the Society and Animals Forum, JAAWS is published by Lawrence E. Erlbaum. For administrative purposes, manuscripts are categorized in four broad areas of animal welfare science. Section Editors for the four content areas are:
PNAS Invites Submissions
The Proceedings of the National Academy of Sciences, U.S.A., invites everyone to submit manuscripts at their Website, <www.PNASCentral.org>. You are not required to submit your paper through an Academy member; the PNAS Office will find a member-editor for you.
* * *
Meeting Announcements
The American Academy of Veterinary Pharmacology & Therapeutics will hold their 14th Biennial Symposium at the Doubletree Hotel in Rockville, Maryland, on May 16-18, 2005. For more details, see <www.ivis.org/newsredir.asp?NL=23&url=n3>.
The 52nd Annual Meeting of the Japanese Association for Laboratory Animal Science will be held May 18-20, 2005, in Tokyo. For information, see <www.cs-oto-com/52jalas/English.htm>; or contact Masanori Suda, Secretary General of the 52nd Annual Meeting, Kanto Daiichi Service Co., Ltd, 1-16-13 Kitakasai, Edogawa-ku, Tokyo 134-8630 [+81-3-5696-8314; fax: +81-3-5696-8333; e-mail: [email protected]].
The 2005 Tribranch Symposium, sponsored by the Delaware Valley, Metro New York, and New Jersey branches of AALAS, will be held June 6-8 in Atlantic City, New Jersey. The meeting�s theme is �Animal Research - We Care�. See <www.tribranch.org>.
The 44th Annual Symposium of the Canadian Association for Laboratory Animal Science/Association Canadienne Pour la Science des Animaux de Laboratoire will be held in Vancouver, British Columbia, June 25-28, 2005. The theme this year, �Research in Motion, Making a Difference�, reflects the contributions and efforts the research community has made to improve the lives of both humans and animals around the world. The scientific program offers a mix of presentations on animal welfare, aquatics, ocular research, and practical training in current technical procedures with over 20 workshops available. Also offered are human resources modules and the latest information on diagnostic assessment methodology, oncology, and the building blocks necessary for good laboratory practices. Registration and more information are available at <www.calas-acsal.org>. Abstract deadline for posters is May 1, 2005.
The 2005 Animal Behavior Society meeting will be held in Snowbird, Utah, August 6-10. For information, see <www.animalbehavior.org/ABS/Program>.
The XXIXth International Ethological Conference will be held in Budapest, Hungary, August 20-27, 2005. To receive the Conference Newsletter, send an empty message to <[email protected]>. For more information, see <www.behav.org>.
The Oregon National Primate Research Center is hosting the 23rd Annual Symposium on Nonhuman Primate Models for AIDS, September 21-24, 2005, at the Hilton Portland & Executive Tower, Portland, Oregon. For information, see <www.ohsu.edu/NHPM2005>; or contact Lori Boshears, Conference Coordinator, Oregon NPRC, 505 NW 185th Ave, Beaverton, OR [503-533-2400; fax: 503-418-2719; e-mail: [email protected]].
The American Association of Zoo Veterinarians and the American Association of Wildlife Veterinarians will be holding a joint conference in Omaha, Nebraska, October 14-21, 2005. Topics for sessions will include: Nutrition, Health issues associated with mixed species and complex exhibits, Preventive medicine/reproduction and contraception, Legislative and regulatory issues, Case reports, Conservation projects in your backyard and Conservation updates, Guidelines for veterinarians working abroad and reports from the field, Wildlife epidemiology using remote imaging and sensing technologies, Anesthesia, and Emerging and zoonotic diseases of captive and free-ranging wildlife. For additional conference information, see <www.aazv.org>.
The 56th AALAS National Meeting will be held in St. Louis, Missouri, November 6-10, 2005. For information, see <www.aalas.org>.
* * *
Volunteer Opportunities
Veterinarian - Sierra Leone
The Tacugama Chimpanzee Sanctuary is currently seeking a volunteer veterinarian for a six-month residency at the orphanage in Freetown. Tacuagama cares for nearly 80 chimpanzees and is seeking a qualified veterinarian with primate (especially great ape) experience, capable of handling the day-to-day healthcare needs of the sanctuary, along with monitoring Tacugama�s veterinary protocols as it progresses towards reintroduction programs. For more information, please contact Tacugama manager Bala Amarasekaran [e-mail: [email protected]].
Kalaweit Rehabilitation Program
The Kalaweit Gibbons and Siamang Rehabilitation Program is now open for volunteers. Volunteers can take care of the gibbons and siamangs in the Kalaweit Project in Borneo or Sumatra. For more information, see <www.kalaweit.org>, or contact Chanee, Kalaweit, Jl. Pinus No 14, Palangka Raya, 73000, Indonesia [+62 536 26388; fax: +62 536 26388; e-mail: [email protected]].
* * *
Positions Available
Campus Veterinarian - NHLBI
The National Heart, Lung and Blood Institute (NHLBI), NIH, is seeking applicants for the position of Campus Veterinarian for NHLBI activities conducted on the NIH-campus in Bethesda, Maryland. This position offers the opportunity to make substantial contributions to the NHLBI, Division of Intramural Research (DIR) program. The DIR currently utilizes a wide variety of laboratory animals including nonhuman primates (macaques, tamarins and squirrel monkeys), farm animals (swine and sheep), dogs, rabbits, and rodents in its research program. As a key member of the Laboratory of Animal Medicine and Surgery (LAMS) team, the successful applicant will have responsibility for planning, directing, coordinating, and evaluating all aspects of animal care and use activities on the NIH campus.
Specific responsibilities include
Salary will be commensurate with experience. Officers in the Commissioned Corps of the U.S. Public Health Service may also apply. Minimum qualifications are a DVM or equivalent degree from an AVMA-accredited school of veterinary medicine (board eligibility or certification by ACLAM is desirable), licensure in at least one state, experience in the AAALAC accreditation process, and five years of progressively responsible experience in an animal research program. Experience with mouse breeding colonies and/or research-oriented surgical experience is also desirable.
The submission deadline is May 1, 2005, or until the position is filled. To apply for this position, submit CV and the names, addresses, and telephone numbers of four professional references to James �Buster� Hawkins, DVM, MS, DACLAM, 14 Service Rd South, Bldg 14E, Rm 105A, Bethesda, MD 20892-5570 [301-451-6743; e-mail: [email protected]]. NIH is an EEO employer.
Associate Operations Manager - Yerkes
Yerkes National Primate Research Center is seeking an experienced individual who, under general direction, will work with the Operations Manager in managing a comprehensive animal care program that maintains proper housing, sanitation, nourishment, environmental conditions and humane handling for a large and diverse population of laboratory animals at the Yerkes Main Center. This person will participate in the recruiting, hiring and orientation of new employees; lead, support and evaluate managerial and supervisory staff; supervise animal care staff directly or indirectly as appropriate; participate in the training of animal care staff on standard operating procedures; and support continuing education and technical development activities.
Preference will be given to individuals with a bachelor�s degree and relevant work and extensive supervisory experience; certification as Laboratory Animal Technologist by the American Association for Laboratory Animal Science; excellent organizational, problem solving, and communication skills; and a proven track record of good personnel management. Further details, including salary range and benefits, can be accessed at <www.emory.edu>; see SAR job vacancy #144462.
To be considered, send a letter of interest and a resume to Dr. James Else, Associate Director for Animal Resources, Yerkes NPRC, Emory Univ., 954 Gatewood Rd., Atlanta, GA 30329 [e-mail: [email protected]]. Emory is an EEO/AA University.
Licensed Veterinary Technician/Technologist
The Bronx Veterans Medical Research Foundation is seeking a veterinary technician to provide support to the Veterinary Medical Officer. The responsibilities of the technician will be surgical, nursing, animal health assessment, performance of routine laboratory diagnostic procedures, transgenic colony maintenance, and husbandry responsibilities when deemed necessary by supervisory persons. This person will perform a range of diagnostic support duties such as taking, recording, and reporting to supervisor deviations in vital signs, taking specimens, and labeling specimens for the laboratory. S/he will arrange and pass medical instruments and materials to medical/health professionals for a variety of diagnostic tests and procedures.
This person makes gross visual observations and identifies subtle changes regarding the animals� condition; uses appropriate handling procedures for hazardous specimens, chemicals, etc.; adheres to safe laboratory practices for working in bio-safety environments; properly disposes of potentially infectious waste materials or other potentially hazardous waste; prepares samples of blood, tissue, urine or other organic substances, providing specimens that effectively support test, examination, or research procedures; ensures that safe and humane processes are used with critical concern for the donor and for the quality of the specimen; uses standard aseptic procedures and safeguards; prepares slides, smears, cultures, etc., of extremely delicate or otherwise volatile specimens, using critical concern for the quality of the specimen; controls receipt, storage and preparation of specimens; and ensures appropriate accountability for all specimens received/created for test or examination by the laboratory.
Primate and transgenic experience is a plus, as are good surgical skills. Salary scale is $30,000 to $53,946. For information or to apply, contact Richard Mann, Veterinary Medical Officer (151), Dept of Veterans Affairs Med. Center, 130 West Kingsbridge Rd., Bronx, NY 10468 [718-584-9000 ext 1708; fax: 718-562-9120; e-mail: [email protected]].
* * *
Resources Wanted and Available
Primate Info Net Seeking Digital Images
The Wisconsin Primate Research Center Library is developing a new series of primate factsheets for Primate Info Net. Each individual factsheet will cover taxonomy, morphology, ecology, behavior, and conservation for (ultimately) all NHP species - and provide species-level access to many other resources found on Primate Info Net and other sites. In order to properly illustrate various points in the text, they are seeking appropriate digital images and audio files for each of the first group of species. File contributors would receive full credit on the Website and would retain copyright for the images/audio files.
They are seeking images associated with:
The species of interest now are
If you can help, please contact Matthew Hoffman, NPRC, University of Wisconsin-Madison, 1220 Capitol Ct, Madison, WI 53715 [608-263-5537; e-mail: [email protected]].
ACVP and ASVCP Proceedings Available On-Line
The Proceedings of the Annual Meeting of the American College of Veterinary Pathologists (ACVP) and the American Society for Veterinary Clinical Pathology (ASVCP) are now available on the International Veterinary Information Service Website: <www.ivis.org/proceedings/ACVP/2004/toc.asp>.
Veterinary Virology
New chapters have been added to the book, A Concise Review of Veterinary Virology, written by G. Carter, D. Wise, and E. Flores. These chapters, �Laboratory diagnosis of viral infections�; �Prevention of viral diseases, vaccines and antiviral drugs�; and �Host defenses to viruses� are available at <www.ivis.org/new.asp>.
The Monkey Toy Store
Alpha Genesis, Inc. (AGI) has developed an on-line enrichment super store: ALPHA TOYS, developed to meet the increased demand for toys and enrichment materials suited for nonhuman primates in laboratories, zoos, and other captive locales. Items featured in this on-line store have been tested for suitability for primates and include many of the favorites of our institution.
Why toys for monkeys? Toys are environmental enrichment and provide primates the ability to express species-typical behaviors. They provide occupation and make life in captivity more interesting, providing for the psychological well-being of animals of captivity. While toys don�t guarantee happiness, they do bring out natural exploration, play, and social behaviors for animals that would otherwise have little to do with their free time.
�Find us at <stores.ebay.com/Alpha-Toys>. We use /Pay Pal/, but that�s not the only way to buy items. You can send your order to <[email protected]>.�
ALPHA TOYS is owned and operated by Alpha Genesis, Inc (AGI). Greg Westergaard, an animal psychologist, is AGI�s President and CEO. AGI is an AAALAC-accredited animal breeding and research facility that maintains more than 5,000 nonhuman primates on three campuses in rural South Carolina. ALPHA TOYS is under the direction of Dr. Sue Howell [e-mail: [email protected]], who has an extensive background in animal welfare and environmental enrichment. �If you have ideas for items for the superstore, let us know!�
* * *
Workshop Announcement: PASA Annual Management Workshop
The Pan African Sanctuaries Alliance (PASA) will stage the PASA 2005 Management Workshop from June 4-8 at the Mount Kenya Safari Club in Mount Kenya, Kenya. The annual workshop, which will bring together African sanctuary managers, advisors, experts and supporters, will be the sixth since the formation of PASA in 2000.
Highlights of the workshop will include a visit to the Sweetwaters Chimpanzee Sanctuary and presentations on topics such as chimpanzee genetics, reintroduction, and non-invasive research.
PASA is a consortium of primate sanctuaries across Africa that collectively care for over 670 chimpanzees, 80 gorillas, 40 bonobos, and literally thousands of other endangered primates. For more information, please contact PASA, P.O. Box 351651, Los Angeles, CA 90035 [e-mail: [email protected]].
* * *
Research and Educational Opportunities
Behavior and Ecology Courses - Costa Rica
There will be four sessions of Primate Behavior and Ecology courses at La Suerte Biological Field Station in Costa Rica in 2005
Primatology/Ecology Field School - Kenya
Rutgers University and the National Museums of Kenya are establishing a primatology and ecology field school in Kenya, beginning this summer. This field school will give participants the opportunity to experience the diverse habitats of Kenya, and to gain understanding about biodiversity by using primate field studies as the entry point. Participants will be exposed to specific conservation problems and emerging innovative solutions that are contextually and culturally different than any they will find elsewhere. The curriculum will comprise lectures, readings, and discussions on important concepts in primatology and conservation biology. We will review important notions in primate behavioral ecology, and learn about primate behavior and ecological data collection. The core of the field school will be training and practice of field methods. Participants will learn how to census primates, study social behavior and habitat use, practice animal identification, time budget analysis by scan and focal animal sampling, and how to measure habitat use. In order to expose the participants to methods used to study primates found in different habitat types, the field school will be conducted at two ecologically different sites: a forest ecosystem in the Tana River Primate National Reserve (TRPNR) at the coast, and a savanna ecosystem at Segera Ranch in northern Kenya. For details, see <primate.rutgers.edu>.
CLASS Lab Animal Science Seminar
The Armed Forces Institute of Pathology, American Registry of Pathology, and the C. L. Davis DVM Foundation jointly sponsor the 8th Current Laboratory Animal Science Seminar (CLASS) July 31-August 1, 2005, at the Doubletree Hotel, Rockville, Maryland. This intensive two-day seminar provides a comprehensive review of selected topics in laboratory animal science and medicine. It is intended to serve the needs of veterinarians across a broad spectrum: entry-level laboratory animal medicine, clinical veterinarians, researchers, residents, training program directors, and facility directors. The seminar includes lectures on animal models, research methods, medicine and surgery, emerging diseases, occupational health issues, regulations, laws and guidelines, alternatives to laboratory animals, and facility management.
For more information contact: Course Coordinator Mark Hovland, Dept of Medical Education, Armed Forces Institute of Pathology, Washington, DC 20306-6000 [202-782-2637; 800-577-3749; e-mail: [email protected]]. You can register at <www.afip.org/Departments/edu/upcoming.htm>.
* * *
Primates de las Am�ricas...La P�gina
Estimados lectores, en esta edici�n ofrecemos dos res�menes de inter�s. El primero corresponde a una tesis de licenciatura recientemente defendida en Argentina. Seguidamente, el resumen de una tesis de postgrado en progreso en el estado de Par�, Brasil (en portugu�s). Saludos cordiales, Tania Urquiza-Haas <[email protected]> y Bernardo Urbani <[email protected]>.
Estudio del allogrooming y su posible rol social en el mono aullador negro (Alouatta caraya). Silvana Peker, Fac. de Ciencias Exactas y Naturales, Univ. de Buenos Aires, Argentina <[email protected]>. Tesis de Licenciatura. Director: Dr. Gabriel E. Zunino, Museo Argentino de Ciencias Naturales �Bernardino Rivadavia�.
La funci�n del acicalamiento entre individuos (allogrooming) fue analizada en una tropa de monos aulladores, Alouatta caraya, en la Isla Brasilera, Chaco, Argentina. El acicalamiento entre individuos en esta especie de mono platirrino tiene una funci�n social, como se ha encontrado en otras especies de monos catarrinos. Durante las 164 horas en contacto con la tropa se registraron 389 encuentros de acicalamiento usando el m�todo de muestreo de �todas las ocurrencias�. Se encontr� una correlaci�n entre el acicalamiento recibido y el tama�o corporal del receptor, pero no con la duraci�n del acicalamiento. El acicalamiento no se distribuy� al azar dentro de la tropa. Los machos adultos y el subadulto recibieron en forma desigual acicalamiento: uno de ellos fue el m�s favorecido. Las hembras adultas fueron las que acicalaron m�s activamente, se solicitaron entre ellas m�s acicalamiento que a otras categor�as y participaron en m�s sesiones rec�procas que los machos. Las c�pulas estuvieron precedidas y seguidas por allogrooming. Se examin� la funci�n higi�nica analizando la distribuci�n corporal de las solicitudes de allogrooming, de acuerdo con el grado variable de accesibilidad de las distintas partes del cuerpo. Algunos sitios accesibles tuvieron una frecuencia de solicitudes mayor a la proporci�n que ocupan en el cuerpo, lo cual no puede explicarse mera-mente por la funci�n higi�nica. Los sitios inaccesibles fueron solicitados con mayor frecuencia. La funci�n social del allogrooming est� reflejada por su rol dentro del cortejo y por la preferencia de un macho en particular como receptor de acicalamiento. La funci�n higi�nica est� expuesta por la alta concentraci�n de solicitudes en sitios inaccesibles al propio individuo. Por lo tanto, estos resultados apoyan la naturaleza multi-funcional del acicalamiento.
Ecologia e organiza��o social do cuxi� preto (Chiropotes satanas): Potencial para sua sobreviv�ncia a longo prazo no contexto de fragmenta��o de habitat na Amaz�nia oriental. Liza Veiga, Depto de Psicologia Experimental, Univ. Fed. do Par�, Brasil <[email protected]>.
Os cuxi�s (g�nero Chiropotes) s�o frug�voros especializados para a preda��o de sementes. Em geral preferem �reas relativamente grandes e n�o perturbadas de florestas altas de terra firme. O objeto deste estudo, o cuxi�-preto (Chiropotes satanas) � end�mico de uma �rea restrita em uma regi�o povoada onde o desmatamento contribui para colocar a esp�cie em perigo de extin��o. A ecologia e o comportamento social do g�nero Chiropotes s�o pouco conhecidos e, com uma �nica exce��o, todos os estudos pr�vios restringem-se basicamente a levantamentos populacionais, or�amentos de atividades, assim como ao comportamento alimentar e de forrageio. O objetivo principal desta pesquisa � estudar a ecologia comportamental e a organiza��o social do cuxi�preto dentro do contexto de fragmenta��o de seu habitat. Desta maneira, dois grupos de animais est�o sendo estudados: o primeiro em um frag-mento de floresta isolada e o outro numa �rea de floresta cont�nua, o qual serve em parte como uma forma de teste-munha. Para cada grupo ser� analisada de que maneira par�metros ecol�gicos influenciam estrat�gias comportamentais. Varia��es na produtividade de habitat, principal-mente a disponibilidade de recursos alimentares, ser�o relacionadas aos diferentes aspectos do comportamento (alimentar, social, or�amento de atividades e uso de espa�o). Os resultados obtidos no fragmento e na floresta cont�nua ser�o comparados com o objetivo de entender melhor as diferen�as ecol�gicas e comportamentais entre habitat fragmentado e n�o fragmentado. O estudo est� sendo realizado na Ilha de Jo�o do B� - P1 (20 ha), e na Base 4 - P2 (1300 ha), ambos pr�ximos � Base 4 da Eletronorte na margem direita do rio Tocantins, no reservat�rio de Tucuru�, Par� (3� 45� S, 49� 41� W). A vegeta��o predominante na regi�o � a floresta tropical �mida. A temperatura m�dia anual � pouco vari�vel ao longo do ano (entre 25� C e 28� C) e existe uma esta��o seca bem defin-ida. O comportamento dos dois grupos ser� monitorado durante um per�odo de 12 meses utilizando-se os m�todos de amostragem de varredura a cada 5 minutos, bem como amostragem de comportamentos. Os dados comporta-mentais ser�o coletados durante 5 dias consecutivos por m�s para cada grupo, o que equivale a pelo menos 110 horas de observa��o. Transec��es bot�nicas cobrindo �reas de 0,5 ha e 1 ha foram implantadas na Ilha e na Trilha 4, respectivamente. Todas as �rvores com um di�metro � altura do peito maior ou igual a 10cm foram medidas e marcadas para a realiza��o de um invent�rio flor�stico e a coleta de dados mensais de fenologia. Observa��es preliminares de comportamento obtidas durante a fase de habitua��o corroboram os resultados de outros estudos do g�nero Chiropotes e os demais estudos feitos na mesma �rea, mostrando o consumo de uma alta propor��o de sementes imaturas na dieta. Al�m do valor intr�nseco de informa��es sobre as caracter�sticas ecol�gicas do cuxi�-preto, o conhecimento detalhado acumulado neste estudo ajudar� na formula��o de planos para conserva��o e na identifica��o de fatores limitantes � viabilidade a longo prazo de popula��es remanescentes no cen�rio de fragmenta��o da floresta na Amaz�nia oriental.
* * *
Calls for Award Nominations
AAALAC International Technician Fellowship Award
AAALAC International has launched the �AAALAC International Technician Fellowship�. The program is made possible through a grant by Priority One Services, Inc., and in cooperation with the American Association for Laboratory Animal Science (AALAS), the Institute of Animal Technology (IAT), the U.K. Medical Research Council, and the National Institutes of Health.
The Fellowship recognizes two outstanding technicians-one IAT-Registered Technician (RAnTech) and one AALAS-Certified Technician (ALAT, LAT, LATG, or CMAR)-who have made (or have the potential to make) significant contributions to the field of laboratory animal care and use. As part of the Fellowship, the IAT-Registered recipient will participate in a week-long educational internship at an animal care and use program within a U.S. institution, then attend the National AALAS meeting in St. Louis, Missouri. The AALAS-certified recipient will participate in a week-long educational internship at an animal care and use program within an institution in the United Kingdom or the Republic of Ireland, then attend the IAT meeting (location still to be determined). Both recipients will write an article about their experiences for AAALAC International�s Connection publication.
Nomination packages for IAT Registered Technicians must be received by May 1, 2005, and should include: � one nomination letter (self nominations are encouraged!), � the nominee�s CV or resume, and � three letters of support. Nomination packages should be sent to: AAALAC International, The International Technician Fellowship Selection Committee, 11300 Rockville Pike, Suite 1211, Rockville, MD 20852-3035. Questions can be sent to <[email protected]>. Nominations for the AAALAC-Certified Technicians will be due October 1.
2nd Annual Lab Products Animal Technician Award
Lab Products, Inc., is pleased to announce their Animal Technician Awards Program. This program is intended to reward a deserving animal care technician from each of the eight AALAS Districts with the opportunity to attend their first National AALAS Meeting. The award will be limited to animal care personnel, with at least one year of laboratory animal care experience, who have never attended a National AALAS Meeting. The recipients of the Lab Products Animal Technician Award will each receive an award recognition plaque; airfare, hotel and registration for the National AALAS Meeting; one year membership to National AALAS; and $250 to cover incidentals while attending the AALAS Meeting.
Nominations are to be submitted by letter, detailing the technicians� work history, accomplishments, and community involvement; and describing how this travel award will benefit the technician and his/her facility. AALAS Branch membership, and involvement in branch AALAS activities, will be considered in the award selection. Supporting letters are encouraged and will be considered, but are not required. Send nominations to: Attn: Awards Selection Committee, Lab Products, Inc., P.O. Box 639, Seaford, DE 19973 [800-526-0469; fax: 302-628-4309]. Nominations must be received by June 15.
HSUS 2005 Russell and Burch Award Nominations
The Humane Society of the United States (HSUS) presents the Russell and Burch Award to scientists who have made outstanding contributions toward the advancement of alternative methods in the areas of biomedical research, testing, or higher education. Alternative methods are those that can replace or reduce the use of animals in specific procedures, or refine procedures so that animals experience less pain or suffering. The award, which carries a $5,000 prize, is named in honor of William M. Russell and Rex L. Burch, the scientists who formulated the Three Rs approach of replacement, reduction, and refinement. HSUS presents the award every three years at the triennial World Congresses on Alternatives and Animal Use in the Life Sciences. The next World Congress will be held August 2005 in Berlin, Germany (see <www.worldcongress.net>). Candidates for the award are judged on the scientific merit of their contributions to the alternatives field and on their professional commitment to this field. Applicants should have a history of laboratory work that is above reproach on humane grounds.
Send nominations by May 16, 2005, to: Russell & Burch Award, Animal Research Issues Section, The HSUS, 2100 L St, NW, Washington, DC 20037 [301-258-3042; fax: 301-258-7760; e-mail: [email protected]]. No special forms are necessary. Persons nominating themselves should submit a cover letter explaining their suitability for the award (see preceding paragraph), a CV, and representative published articles. Persons nominating others should submit a letter explaining the nominee�s suitability for the award and arrange to have supporting documents forwarded.
* * *
Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
Books
* The Missing Link in Cognition: Origins of Self-Reflective Consciousness, H. S. Terrace & J. Metcalfe (Eds.). New York: Oxford University Press, 2004. [Price: $75.00]
. . . Contents include: Progress in the study of chimpanzee recall and episodic memory, by C. Menzel; Do nonhuman primates have episodic memory? By B. L. Schwarz; Studies of uncertainty monitoring and metacognition in animals and humans, by J. D. Smith; Can rhesus monkeys discriminate between remembering and forgetting? By R. R. Hampton; and Metaconfidence judgements in rhesus macaques: Explicit versus implicit mechanisms, by L. K. Son & N. Kornell.
* The Cultured Chimpanzee: Reflections on Cultural Primatology. W. McGrew. New York: Cambridge University Press, 2004. [Price: $29.99 (paperback); $90 (hardback)]
. . . The Cultured Chimpanzee explores the variation in chimpanzee behavior across its range, which cannot be explained by individual learning, nor by genetic or environmental influences. It promotes the view that this rich diversity in social life and material culture reflects social learning of traditions, and more closely resembles cultural variety in humans than the simpler behavior of other animal species.
* Animal Rights: Current Debates and New Directions. C. R. Sunstein, & M. C. Nussbaum (Eds.). Oxford: Oxford University Press, 2004. 352 pp. [Price: $29.95]
* Primate Cytogenetics. S. M�ller (Ed.). Farmington, CT: Karger, 2005. 268 pp. [Price: EUR 87; US$111.00]. Hardcover reprint of Cytogenetic and Genome Research, 2005, 108[1-3].
* The Human Story: A New History of Mankind�s Evolution. R. Dunbar. London: Faber and Faber, 2005. 208 pp. [Price: $23]
Journal Contents
* Journal of Medical Primatology, 2005, 34[1].
. . . Expression of human IL-1a after intramarrow gene transfer into healthy non-human primate by adenoviral vector, by N. B�card, T. Revel, T. Sorg, D. Dormont, & R. Le Grand; Herpes B-virus seroreactivity in a colony of Macaca mulatta: Data from the Sabana Seca Field Station, a new Specific Pathogen-Free Program, by C. A. Sariol, T. Arana, E. Maldonado, M. Gerald, J. Gonzalez-Martinez, M. Rodriguez, & E. N. Kraiselburd; The significance of platelet-activating factor and fertility in the male primate: A review, by W. E. Roudebush, J. B. Massey, C. W. Elsner, D. B. Shapiro, D. Mitchell-Leef, & H. I. Kort; Contraception in the chimpanzee: 12-year experience at the CIRMF Primate Centre, Gabon, by O. Bourry, P. Peignot, & P. Rouquet; Normal hematological indices, blood chemistry and histology and ultrastructure of pancreatic islets in the wild Indian bonnet monkeys (Macaca radiata radiata), by M. D. Mythili, R. Vyas, S. S. Patra, S. C. Nair, G. Akila, R. Sujatha, & S. Gunasekaran; Congenital bronchiolo-alveolar airway malformation in a cynomolgus macaque (Macaca fascicularis), by B. S. Lewis, G. B. Hubbard, M. G. Mense, P. A. Frost; and Fatal inflammatory heart disease in a bonobo (Pan paniscus), by P. Jones, C. Mahamba, J. Rest, & C. Andr�.
Magazines and Newsletters
* Gorilla Journal, Journal of Berggorilla & Regenwald Direkthilfe, 2004, No. 29. [c/o Rolf Brunner, Lerchenstr. 5, 45473 Muelheim, Germany] <www.berggorilla.de/gj293.pdf>
* Positively Primates, 2004, 10. [DuMond Conservancy, 14805 S.W. 216 St, Miami, FL 33170]
* Zoo View: The Quarterly Magazine of the Greater Los Angeles Zoo Association, Winter 2005, 38[4]. This issue is dedicated to ape conservation.
Proceedings
* 16th meeting of the Italian Primatological Society, held at Convento dell�Osservanza, Radicondoli (SI), October 28-30, 2003. D. Formenti & A. Vitale (Guest Eds.). Folia Primatologica, 2004, 75, 385-414.
* African Journal of Ecology, 2004, 42[Suppl 1].
. . . This supplement consists of papers presented at a conference at Makerere University, Kampala, Uganda, August 20-21, 2001. Contents include: Forest fragmentation and primates� survival status in non-reserved forests of the �Kampala area�, Uganda, by D. Baranga; Red-tail monkey groups in forest patches outside the protected area system in the �Kampala area�, by D. Baranga; and The status and distribution of primates in Mubende-Toro woodlands and forests, by G. Isabirye-Basuta.
* Abstracts from the 5th Meeting of the Spanish Primatological Society. Folia Primatologica, 2005, 76, 45-66.
Reports
* Guidance on the transport of laboratory animals. J. Swallow, D. Anderson, A. C. Buckwell, T. Harris, P. Hawkins, J. Kirkwood, M. Lomas, S. Meacham, A. Peters, M., Prescott, S., Owen, R. Quest, R. Sutcliffe, & K. Thompson, (Pfizer Global Res. & Devel., Ramsgate Rd, Sandwich, Kent CT13 9NJ, U.K. [e-mail: [email protected]]). Laboratory Animals, 2005, 39, 1-39.
. . . Report of the Transport Working Group established by the Laboratory Animal Science Association.
Special Journal Issues
* Special Issue XVI. 21st Annual Symposium on Nonhuman Primate Models for AIDS. Journal of Medical Primatology, 2004, 33[5-6].
. . . Contents: Introduction, by P. A. Marx, S.-L. Hu, & T. R. Secrist; AIDS as a zoonosis? Confusion over the origin of the virus and the origin of the epidemics, by P. A. Marx, C. Apetrei, & E. Drucker; A real-time PCR-based method to independently sample single simian immunodeficiency virus genomes from macaques with a range of viral loads, by D. Williams & J. Overbaugh; SIV-induced activation of the blood-brain barrier requires cell-associated virus and is not restricted to endothelial cell activation, by A. G. MacLean, T. A. Rasmussen, D. N. Bieniemy, X. Alvarez, & A. A. Lackner; Perinatal transmission of SHIV-SF162P3 in Macaca nemestrina, by P. Jayaraman, D. Mohan, P. Polacino, L. Kuller, N. Sheikh, H. Bielefeldt-Ohmann, B. Richardson, D. Anderson, S.-L. Hu, & N. L. Haigwood; T-cell-mediated protective efficacy of a systemic vaccine approach in cynomolgus monkeys after SIV mucosal challenge, by Z. Michelini, D. R. M. Negri, S. Baroncelli, S. Catone, A. Comini, M. T. Maggiorella, L. Sernicola, F. Crostarosa, R. Belli, M. G. Mancini, S. Farcomeni, Z. Fagrouch, M. Ciccozzi, C. Rovetto, P. Liljestrom, S. Norley, J. Heeney, & F. Titti; Characterization of virus infectivity and cell-free capsid assembly of SIVMneCL8, by J. E. Dooher, M. J. Pineda, J. Overbaugh, & J. R. Lingappa; Cell-free systems for capsid assembly of primate lentiviruses from three different lineages, by J. E. Dooher & J. R. Lingappa; and the abstracts from the Symposium.
* Infectious disease research in the age of biodefense. ILAR Journal, 2005, 46[1] <www.nationalacademies.org/pr4601>.
. . . Articles include: Demand for nonhuman primate resources in the age of biodefense.
* Enrichment strategies for laboratory animals. ILAR Journal, 2005, 46[2] <www.nationalacademies.org/pr4601>.
. . . Contents include: USDA perspective on environ-mental enrichment for animals; Mandatory �enriched� housing of laboratory animals: The need for evidence-based evaluation; Stereotypies and other abnormal repetitive behaviors: Potential impact on validity, reliability, and replicability of scientific outcomes; Balancing animal research with well-being: Establishment of goals and harmonization of approaches; Potential for unintended consequences of environmental enrichment for laboratory animals and research results; Enrichment and nonhuman primates: First, do no harm; Environmental enrichment for nonhuman primates: Theory and application; and Behavioral management of chimpanzees in biomedical research facilities: The state of the science.
Anatomy and Physiology
* Toward an evolutionary perspective on conceptual representation: Species-specific calls activate visual and affective processing systems in the macaque. Gil-da-Costa, R., Braun, A., Lopes, M., Hauser, M. D., Carson, R. E., Herscovitch, P., & Martin, A. (A. M., Lab. of Brain & Cognition, NIMH, NIH, Bldg 10, Rm 4C-104, Bethesda, MD 20892-1366 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2004, 101, 17516-17521.
. . . Nonhuman primates produce a diverse repertoire of species-specific calls and have rich conceptual systems. Some of their calls are designed to convey information about concepts such as predators, food, and social relationships, as well as the affective state of the caller. Little is known about the neural architecture of these calls, and much of what we do know is based on single-cell physiology from anesthetized subjects. By using positron emission tomography in awake rhesus macaques, it was found that conspecific vocalizations elicited activity in higher-order visual areas, including regions in the temporal lobe associated with the visual perception of object form (TE/TEO) and motion (superior temporal sulcus) and storing visual object information into long-term memory (TE), as well as in limbic (the amygdala and hippocampus) and paralimbic regions (ventromedial prefrontal cortex) associated with the interpretation and memory-encoding of highly salient and affective material. This neural circuitry strongly corresponds to the network shown to support representation of conspecifics and affective information in humans. These findings shed light on the evolutionary precursors of conceptual representation in humans, suggesting that monkeys and humans have a common neural substrate for representing object concepts.
* Handedness in chimpanzees (Pan troglodytes) is associated with asymmetries of the primary motor cortex but not with homologous language areas. Hopkins, W. D., & Cantalupo, C. (Div. of Psychobiology, Yerkes NPRC, Emory Univ., Atlanta, GA 30322 [e-mail: [email protected]]). Behavioral Neuroscience, 2004, 118, 1176-1183.
. . . The neurobiology of hand preferences in nonhuman primates is poorly understood. In this study, the authors report the first evidence of an association between hand preference and precentral gyrus morphology in chimpanzees. Hand preferences did not significantly correlate with other asymmetric brain regions associated with language functions in humans, including the planum temporale and frontal operculum. The overall results suggest that homologous regions of the motor cortex control hand preferences in humans and apes and that these functions evolved independently of left-hemisphere specialization for language and speech.
* Asymmetries in the hippocampus and amygdala of chimpanzees (Pan troglodytes). Freeman, H. D., Cantalupo, C., & Hopkins, W. D. (W. D. H., address same as above). Behavioral Neuroscience, 2004, 118, 1460-1465.
. . . Magnetic resonance imaging was used to measure the hippocampal and amygdalar volumes of 60 chimpanzees. An asymmetry quotient (AQ) was then used to calculate the asymmetry for each of the structures. A one-sample t test indicated that there was a population-level right hemisphere asymmetry for the hippocampus. There was no significant population-level asymmetry for the amygdala. An analysis of variance using sex and rearing history as between-group variables showed no significant main effects or interaction effects on the AQ scores; however, males were more strongly lateralized than females. Several of these findings are consistent with results found in the human literature.
Animal Models
* Heart transplantation in baboons using 1,3-galactosyltransferase gene-knockout pigs as donors: Initial experience. Kuwaki, K., Tseng, Y.-L., Dor, F. J. M. F., Shimizu, A., Houser, S. L., Sanderson, T. M., Lancos, C. J., Prabharasuth, D. D., Cheng, J., Moran, K., Hisashil, Y., Mueller, N., Yamada, K., Greenstein, J. L., Hawley, R. J., Patience, C., Awwad, M., Fishman, J. A., Robson, S. C., Schuurman, H.-J., Sachs, D. H., & Cooper, D. K. C. (D. K. C. C., Starzl Transplantation Inst., Univ. of Pittsburgh Med. Ctr, Biomed. Sci. Tower East, Rm E1550A, 200 Lothrop St, Pittsburgh, PA 15261 [e-mail: [email protected]]). Nature Medicine, 2004, 11, 29-31.
. . . Hearts from 1,3-galactosyltransferase knockout pigs (GalT-KO, n=8) were transplanted heterotopically into baboons using an anti-CD154 monoclonal antibody-based regimen. The elimination of the galactose-1,3-galactose epitope prevented hyperacute rejection and extended survival of pig hearts in baboons for 2 to 6 months (median, 78 days); the predominant lesion associated with graft failure was a thrombotic microangiopathy, with resulting ischemic injury. There were no infectious complications directly related to the immunosuppressive regimen. The transplantation of hearts from GalT-KO pigs increased graft survival over previous studies.
* Marked prolongation of porcine renal xenograft survival in baboons through the use of a-1,3-galactosyltransferase gene-knockout donors and the cotransplantation of vascularized thymic tissue. Yamada, K., Yazawa, K., Shimizu, A., Iwanaga, T., Hisashi, Y., Nuhn, M., O�Malley, P., Nobori, S., Vagefi, P. A., Patience, C., Fishman, J., Cooper, D. K. C., Hawley, R. J., Greenstein, J., Schuurman, H.-J., Awwad, M., Sykes, M., & Sachs, D. H. (D. H. S., Transplantation Biology Research Center, Mass. General Hospital, Boston, MA 02129 [e-mail: [email protected]]). Nature Medicine, 2004, 11, 32-34.
. . . The use of animal organs could potentially alleviate the critical worldwide shortage of donor organs for clinical transplantation. Because of the strong immune response to xenografts, success will probably depend upon new strategies of immune suppression and induction of tolerance. Here we report our initial results using a-1,3-galactosyltransferase knockout (GalT-KO) donors and a tolerance induction approach. Life-supporting pig-to-baboon renal xenograft survivals of up to 83 days with normal creatinine levels have been achieved.
* Systemic AA amyloidosis in the common marmoset. Ludlage, E., Murphy, C. L., Davern, S. M., Solomon, A., Weiss, D. T., Glenn-Smith, D., Dworkin, S., & Mansfield, K. G. (New England PRC, Harvard Med/ School, Southborough, MA 01772-9012 [e-mail: [email protected]]). Veterinary Pathology, 2005, 42, 117-124.
. . . The common marmoset (Callithrix jacchus) is a small New World primate native to Brazil that has been used extensively in biomedical research. A retrospective analysis of archived hematoxylin and eosin-stained tissue sections and clinical records was conducted at the New England Primate Research Center on 86 marmosets more than 1 year of age that were euthanized during the past decade because of morbidity and failure to thrive. Approximately 17% (15 of 86) were found to have amyloid deposits in one or more organs, including the liver, adrenal glands, kidneys, and intestine. This material was shown by amino acid sequence analysis to be composed of serum amyloid A (SAA)-related protein. This type of amyloidosis, designated AA or �secondary,� is associated typically with an inflammatory process that induces elevated levels of the SAA amyloidogenic precursor molecule. Notably, there were no significant pathologic differences or other distinguishing features in animals with amyloid versus those without; furthermore, on the basis of the limited number of serum specimens available for analysis, the SAA concentrations in the two groups were comparable, thus suggesting the possible inheritable nature of the disorder. In this respect, the common marmoset provides a unique experimental model for study of the pathogenesis and treatment of AA and other forms of systemic amyloidosis.
* Use of primates in research: A global overview. Carlsson, H.-E. , Schapiro, S. J., Farah, I. , & Hau, J. (Dept of Neuroscience, Div. of Comp. Med., Uppsala Univ., Uppsala, Sweden [e-mail: [email protected]]). American Journal of Primatology, 2004, 63, 225-237.
. . . Studies published in 2001 in peer-reviewed journals were reviewed. The number and species of primates used, the origin of the animals, the type of study, the area of research of the investigation, and the location at which the research was performed were tabulated. Additionally, factors related to the animals that may have affected the outcome of the experiments were recorded. A total of 2,937 articles involving 4,411 studies that employed nonhuman primates or nonhuman primate biological material were identified and analyzed. More than 41,000 animals were represented in the studies published in 2001. In the 14% of studies for which re-use could be determined, 69% involved animals that had been used in previous experiments. Published studies most commonly used nonhuman primates or nonhuman primate biological material from the species Chlorocebus aethiops (19%), Macaca mulatta (18%), M. fascicularis (9%), and Papio spp. (6%). Of these studies, 54% were classified as in vitro studies, 14% as noninvasive, 30% as chronic, and 1% were considered acute. Nonhuman primates were primarily used in research areas in which they appear to be the most appropriate models for humans. The most common areas of research were microbiology (including HIV/AIDS (26%)), neuroscience (19%), and biochemistry/chemistry (12%). Most (84%) of the primate research published in 2001 was conducted in North America, Europe, and Japan. The animals and conditions under which they were housed and used were rarely described. Although it is estimated that nonhuman primates account for an extremely small fraction of all animals used in research, their special status makes it important to report the many husbandry and environmental factors that influence the research results generated. This analysis has identified that editors rarely require authors to provide comprehensive information concerning the subjects (e.g., their origin), treatment conditions, and experimental procedures utilized in the studies they publish. The present analysis addresses the use of primates for research, including the effects of a shortage of suitable nonhuman primate subjects in many research areas.
Animal Welfare
* Laboratory routines cause animal stress. Balcombe, J. P., Barnard, N. D., & Sandusky, C. (HSUS, 2100 L St. NW, Washington, DC 20037). Contemporary Topics in Laboratory Animal Science, 2004, 43, 42-51.
. . . Eighty published studies were appraised to document the potential stress associated with three routine laboratory procedures commonly performed on animals: handling, blood collection, and orogastric gavage. Handling was defined as any non-invasive manipulation occuring as part of routine husbandry, including lifting an animal and cleaning or moving an animal�s cage. Significant changes in physiologic parameters correlated with stress (e.g., serum or plasma concentrations of corticosterone, glucose, growth hormone or prolactin, heart rate, blood pressure, and behavior) were associated with all three procedures in multiple species in the studies we examined. The results of these studies demonstrated that animals responded with rapid, pronounced, and statistically significant elevations in stress-related responses for each of the procedures, although handling elicited variable alterations in immune system responses. Changes from baseline or control measures typically ranged from 20% to 100% or more and lasted at least 30 min or longer. These findings are interpreted to indicate that laboratory routines are associated with stress, and that animals do not readily habituate to them. The data suggest that significant fear, stress, and possibly distress are predictable consequences of routine laboratory procedures, and that these phenomena have substantial scientific and humane implications for the use of animals in laboratory research.
Behavior
* Socially learned preferences for differentially rewarded tokens in the brown capuchin monkey (Cebus apella). Brosnan, S. F., & de Waal, F. B. (Living Links Center, Emory Univ., Yerkes PRC, 954 N. Gatewood Dr., Atlanta, GA 30329 [e-mail: [email protected]]). Journal of Comparative Psychology, 2004, 118, 133-139.
. . . Social learning is assumed to underlie traditions, yet evidence indicating social learning in capuchin monkeys, which exhibit traditions, is sparse. The authors tested capuchins for their ability to learn the value of novel to-kens using a previously familiar token-exchange economy. Capuchins change their preferences in favor of a token worth a high-value food reward after watching a conspecific model exchange two differentially rewarded tokens, yet they fail to develop a similar preference after watching tokens paired with foods in the absence of a conspecific model. They also fail to learn that the value of familiar tokens has changed. Information about token value is available in all situations, but capuchins seem to pay more attention in a social situation involving novel tokens.
* What do Diana monkeys know about the focus of attention of a conspecific? Scerif, G., Gomez, J.-C., & Byrne, R. W. (School of Psychology, University of St Andrews, St Andrews, Fife KY16 9JU, U.K. [e-mail: [email protected]]). Animal Behaviour, 2004, 68, 1239-1247.
. . . �Converging experimental and observational evidence suggests that some nonhuman primates are able to co-orient with shifts in visual attention, of both conspecifics and humans. However, the underlying cognitive mechanisms involved are unclear. To investigate attention following in Diana monkeys, Cercopithecus diana diana, we used photographs of familiar conspecifics orienting towards one of two locations. A subject monkey was shown a photograph, and shortly afterwards a toy appeared at one location or the other. The toy�s position therefore either matched the location signaled by the head and body orientation of the photographed monkey (compatible), or was opposite to that location (incompatible). Overall, monkeys� first inspections, total duration of looking and number of looks were more likely to be directed to the compatible location, that is towards the direction of attention shown in the photograph. Furthermore, when a photograph of an adult monkey signaled attention to one location, but the toy appeared at the opposite (incompatible) location, subjects reinspected the monkey photographs more often than when the toy appeared at the compatible location, suggesting a violation of expectancy. This effect was not the case if the photograph was of an immature animal. Our results show that attention following was not limited to simple reflexive orienting by the monkeys, and that monkeys perceived a relationship existing between agent and object of attention.�
* Lemur latrines: Observations of latrine behavior in wild primates and possible ecological significance. Irwin, M. T., Samonds, K. E., Raharison, J.-L., & Wright, P. C. (Program in Anthropo. Sci., Stony Brook Univ., Stony Brook, NY 11794-4364 [e-mail: [email protected]]). Journal of Mammalogy, 2004, 85, 420-427.
. . . �Latrine behavior, or the preferential, repeated use of one or more specific defecation sites, is well known among mammals and believed to function in olfactory communication among individuals or groups in many circumstances. Primates have reduced their capacity for olfaction in favor of more developed visual systems; however, several prosimian primates regularly use olfactory communication for transmission of social signals, most often using scent gland secretions and urine. Latrine behaviors have been described rarely in primates and have traditionally not been included in reviews of primate olfactory communication, yet we found ample evidence that certain primate species habitually use latrine sites for defecation. Here we review the previous evidence for latrine use in primates and report new and more extensive observations of latrine use in 2 lemuriform primates (Lepilemur sp. and Hapalemur griseus). Based on these new observations, we present and evaluate 4 available hypotheses for the function of latrines (advertisement of sexual cycling, predation avoidance, intragroup spacing, and intergroup resource defense) in lemur taxa for which sufficient evidence of latrine use exists. In all cases, intergroup resource defense is the function most consistent with available observations.�
Conservation
* What can captive breeding do for conservation and what can behavior research do for captive breeding? Swaisgood, R. R. (CRES, Zool. Soc. of San Diego, San Diego, CA 92115 [e-mail: [email protected]]). The Conservation Behaviorist, 2004, 2[2], 3-5. <www.animalbehavior.org/ABS/Conservation/ccConsBeh.html>
. . . �How can we justify confining animals in small enclosures, often far removed from many salient features of the animal�s natural environment? This question speaks to concerns of animal welfare, and I see it as a challenge to behaviorists and managers to understand the behavioral needs of animals and develop captive environments that meet these needs. How can we justify the expenditure of money to maintain a few representatives of endangered species in captivity when the same funds could significantly enhance in situ conservation efforts? A reasonable answer must show that these expenses actually do not take away funds that otherwise could go to conservation of animals in their natural environments and that captive breeding programs contribute to in situ conservation. These questions are interrelated because minimal well-being is a prerequisite for reproduction.�
Disease
* Cytomegalovirus-associated discrete gastrointestinal masses in macaques infected with the simian immunodeficiency virus. Hendricks Hutto, E., Anderson, D. C., & Mansfield, K. G. (New England PRC, One Pine Hill Dr., Southborough, MA 01772 [e-mail: [email protected]]). Veterinary Pathology, 2004, 41, 691-695.
. . . Cytomegalovirus (CMV)-associated gastrointestinal masses have been reported in human acquired immune deficiency syndrome patients. This is the first report on CMV-associated gastrointestinal masses in simian immunodeficiency virus (SIV)-infected macaques. Two SIV-infected macaques presented at necropsy with multiple nodular or umbilicated masses within the gastrointestinal tract. In one animal, the masses were located throughout the gastrointestinal tract, whereas in the other, the masses were restricted to the proximal small intestine. Grossly, the masses were indistinguishable from those caused by neoplastic conditions such as lymphoma and, histologically, were composed of hyperplastic glandular tissue, dense neutrophilic infiltrates within the lamina propria, and multifocal proprial hemorrhage. Frequent cytomegalic cells with basophilic intranuclear inclusions were found in affected regions. Immunohistochemistry for CMV demonstrated frequent immunopositive cells within affected areas. Furthermore, immunohistochemistry for the proliferation marker Ki-67 demonstrated increased proliferation in hyperplastic glands and crypts. CMV should be considered a cause of discrete mass lesions in the gastrointestinal tract of SIV-infected macaques.
* Radiation-induced glioblastoma multiforme in two adult baboons (Papio cynocephalus anubis). Schielke, J. E., Kiem, H. P., Liggitt, D., & Bielefeldt-Ohmann, H. (Washington NPRC, Univ. of Washington, Seattle, WA 98195-1023). Comparative Medicine, 2004, 54, 327-332.
. . . A diagnosis of glioblastoma multiforme (GBM) was made for cerebral masses found at necropsy in two baboons. Case 1 was an adult (6.18 years old) male baboon that suddenly died during a physical examination as part of a clinical evaluation for a leg lameness. Case 2 was an adult (5.95 years old) female baboon that stopped breathing during anesthesia for magnetic resonance imaging to evaluate lethargy, weight loss, inappetence, and dilated pupils. Both animals had undergone total body irradiation with cobalt during a research protocol. The incidence of spontaneous brain tumors in nonhuman primates is low, but radiation-induced GBM lesions in rhesus macaques have been reported. A definitive diagnosis was made in these cases, using histopathologic criteria of cellular pleomorphism, high mitotic rate, regions of coagulation necrosis, and endothelial proliferation.
* Carcinosarcoma of the maxilla in a squirrel monkey (Saimiri sciureus). Moutsopoulos, N. M., Nikitakis, N. G., Powell, D. A., & Reynolds, M. A. (Dept of Periodontics, Univ. of Maryland Dental School, 666 W Baltimore St., Baltimore, MD 21201). Comparative Medicine, 2004, 54, 333-336.
. . . �We present the first, to our knowledge, described case of carcinosarcoma of the maxilla in a squirrel monkey. Carcinosarcomas are rare tumors of the upper aerodigestive tract, and consist of carcinomatous and sarcomatous tissue. Histologic analysis revealed a neoplasm composed of an adenocarcinomatous component (epithelial element) and a mesenchymal component (sarcomatous element). Metastatic growth was documented in the lung tissue and the submandibular lymph node. The histolopathologic findings, the pattern of metastasis, and the clinical progression closely resembled those of carcinosarcoma involving salivary glands in humans.�
* Legg-Calv�-Perthes disease in a rhesus macaque (Macaca mulatta). Smedley, J. V., Lomax, L. G., Williams, J. F., Barras, P. W., & Hasselschwert, D. L. (NCI, 9000 Rockville Pike, Bethesda, MD 20892-5550). Comparative Medicine, 2004, 54, 585-588.
. . . A juvenile rhesus macaque presented with atrophy of the musculature of its left leg. Physical examination localized the problem to the coxofemoral joint. Radiography revealed changes consistent with Legg-Calv�-Perthes (LCP) disease. Femoral head ostectomy was performed, and the femoral head was submitted for histologic examination, results of which confirmed a diagnosis of LCP.
* Animal models of ischemic stroke: Balancing experimental aims and animal care. Graham, S. M., McCullough, L. D., & Murphy, S. J. (Dept of Anesthesiology, Oregon Health & Science Univ., Portland, OR 97239). Comparative Medicine, 2004, 54, 486-496.
. . . Animal models of ischemic stroke are examples of an induced model that can present challenges from the perspectives of protocol review and animal management. The review presented here will include a brief summary of the current state of knowledge about clinical stroke; a general synopsis of important unanswered research questions that justify use of animal stroke models; an overview of various animal models of ischemic stroke, including strengths and limitations; and a discussion of animal care issues relative to ischemic stroke models. Good communication and interactive education among primary investigators, laboratory animal veterinarians and caretakers, and institutional animal care and use committee members are critical in achieving a balance between research objectives and animal care issues when using animal stroke models.
* Outbreak of Mycobacterium bovis in a conditioned colony of rhesus (Macaca mulatta) and cynomolgus (Macaca fascicularis) macaques. Garcia, M. A., Bouley, D. M., Larson, M. J., Lifland, B., Moorhead, R., Simkins, M. D., Borie, D. C., Tolwani, R., & Otto, G. (Dept of Comp. Med., Stanford Univ. School of Med., Stanford, CA 94305). Comparative Medicine, 2004, 54, 578-584.
. . . A tuberculosis outbreak was caused by Mycobacterium bovis in a conditioned colony of rhesus and cynomolgus macaques. Animals in five rooms were exposed, but most (16/27) infections were confined to the room that housed a mixed population of cynomolgus and rhesus macaques. In this room, rhesus (8/8) and cynomolgus (10/11) macaques naturally exposed to M. bovis were infected at nearly identical rates (Fisher exact test, 2-tailed P = 1). The clinical signs of disease and pathologic lesions in infected macaques, however, were moderately different between the two species. Rhesus macaques were more likely (5/8) to exhibit clinical signs of persistent coughing and inappetance, and had more severe pulmonary lesions. By contrast, clinical signs of disease were seen in only 1 of 19 cynomolgus macaques, and overall, the pulmonary lesions were often focal and less severe, although some still had severe involvement of the lungs similar to that seen in rhesus macaques. These differences should be taken into consideration when developing or evaluating a tuberculosis-screening program. On the basis of observations made during this outbreak, we recommend that alternative screening methods, such as the PRIMAGAM test and the ESAT-6 ELISA, be incorporated into the screening program to aid in the identification of infected animals.
* Comparative study of lung cytologic features in normal rhesus (Macaca mulatta), cynomolgus (Macaca fasicularis), and African green (Chlorocebus aethiops) nonhuman primates by use of bronchoscopy. Tate. M. K., Rico, P. J., & Roy, C. J. (Div. of Vet. Med., Fort Detrick, MD 20702-5011). Comparative Medicine, 2004, 54, 393-396.
. . . Invasive bronchoscopy and bronchoaveolar lavage (BAL) fluid collection represents an important tool in studies of the respiratory system of nonhuman primates. Bronchoscopy and BAL fluid collection were performed on groups of rhesus and cynomolgus macaques and African green monkeys, and the resulting comparative lavage cytologic features are described. Analysis of the BAL fluid did not reveal significant differences among species with respect to total cells recovered or differential cellular composition. This description of the method used to lavage the nonhuman primates and the resulting lung cytologic findings provide important comparative data for three species commonly used in biomedical research.
* Subacute necrotizing encephalopathy in a pig-tailed macaque (Macaca nemestrina) that resembles mitochondrial encephalopathy in humans. Beilefeldt-Ohmann, H., Bellanca, R. U., Crockett, C. M., Curnow, E., Eiffert, K., Gillen, M., Glanister, D., Hayes, E., Kelley, S., Minoshima, S., & Vogel, K. (Washington NPRC, Box 357330, Seattle, WA 98195-7330). Comparative Medicine, 2004, 54, 422-433.
. . . A male pig-tailed macaque, approximately 5 years old, was found to be vision-impaired and to have profound behavioral abnormalities, including hyperactivity and self-injurious behavior that was not amenable to amelioration by environmental enrichment. Facial and skeletal dysmorphisms also were noted. Magnetic resonance imaging (MRI) and positron emission tomography (PET) scanning revealed areas of possible infarction in the occipital lobes and megaventriculosis. At necropsy, following euthanasia for humane reasons, severe polio- and leukoencephalomalacia accompanied by megaventriculosis were seen in both occipital lobes and in several sulci of the parietal and frontal lobes. Light microscopic findings included loss of neocortical structure, with necrosis, neuronal loss, astrogliosis, vascular proliferation, mild spongiosis, and demyelination. The extent and severity of lesions were most pronounced in the occipital lobes and were greater in the left than in the right hemisphere. Other lesions included mild bilateral atrophy of the optic nerves, thymic involution, necrotizing dermatitis due to trauma, and a spectrum of spermatozoal abnormalities. The imaging and gross and light microscopic changes found in this animal resemble the mitochondrial encephalopathies of humans; this was corroborated by results of immunohistochemical analysis demonstrating decreased expression of enzymes of the mitochondrial oxidative complex ([OC]-I, -III, and -IV) in brain and muscle, and detection of fibrinogen immunoreactivity in neurons and glial cells. The spermatozoal defects may represent yet another aspect of a mitochondrial defect.
* Ampullary carcinoma in a group of aged rhesus macaques (Macaca mulatta). Usborne, A. L., & Bolton, I. D. (WiPRC, Univ. of Wisconsin, 1220 Capitol Ct, Madison, WI 53715). Comparative Medicine, 2004, 54, 438-442.
. . . Ampullary carcinoma was diagnosed in 6 rhesus macaques that ranged in age from 20 to 35 years. Signalment, premonitory signs of disease, and results of clinical biochemical and hematologic analyses varied among animals. Histologically, the neoplastic cells obliterated the ampulla, with regional spread to the duodenum in all 6 animals and to the pancreas in one animal. Two animals had metastases to the lung, and another two had metastases to the pancreoduodenal lymph nodes and liver. One animal had mesocolonic metastasis. Malignant tumors of the ampullary region are rare in domestic animals, and account for less than 5% of all cancers of the digestive tract in humans.
* Lacrimal gland myoepithelioma in a rhesus macaque (Macaca mulatta). Munday, J. S., Rodriguez, N. A., & Thomas, D. A. (Dept. of Pathology, College of Vet. Med., Univ. of Georgia, Athens, GA 30602). Comparative Medicine, 2004, 54, 443-446.
. . . An 18-year-old rhesus macaque developed ptosis of the left upper eyelid due to a mass that had first been observed 10 years previously. The 11x7x7-mm mass was surgically excised, and the ptosis resolved after 5 days. Histologic examination of the mass revealed two confluent cell populations. Most cells were spindle-shaped and were arranged in loose fascicles. Smaller numbers of cells had squamous differentiation. The spindle-shaped cells expressed smooth muscle actin. Cells with squamous differentiation did not express smooth muscle actin, but did, along with around half of the spindle-shaped cells, express pan-cytokeratin. On the basis of histologic and immunohistochemical findings, the mass was diagnosed as myoepithelioma. The neoplasm most likely originated from the palpebral lobe of the lacrimal gland, although accessory lacrimal gland origin could not be excluded. Recurrence of the neoplasm has not been observed 6 months after surgery.
* A monkey�s tale: The origin of Plasmodium vivax as a human malaria parasite. Escalante, A. A., Cornejo, O. E., Freeland, D. E., Poe, A. C., Durrego, E., Collins, W. E., & Lal, A. A. (Malaria Branch, Div. of Parasitic Diseases, CDC-P, Mail Stop F-12, 4770 Buford Hwy, Chamblee, GA 30341 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2005, 102, 1980-1985.
. . . �The high prevalence of Duffy negativity (lack of the Duffy blood group antigen) among human populations in sub-Saharan Africa has been used to argue that Plasmodium vivax originated on that continent. Here, we investigate the phylogenetic relationships among 10 species of Plasmodium that infect primates by using three genes, two nuclear (b-tubulin and cell division cycle 2) and a gene from the plastid genome (the elongation factor Tu). We find compelling evidence that P. vivax is derived from a species that inhabited macaques in Southeast Asia. Specifically, those phylogenies that include P. vivax as an ancient lineage from which all of the macaque parasites could originate are significantly less likely to explain the data. We estimate the time to the most recent common ancestor at four neutral gene loci from Asian and South American isolates (a minimum sample of seven isolates per locus). Our analysis estimates that the extant populations of P. vivax originated between 45,680 and 81,607 years ago. The phylogeny and the estimated time frame for the origination of current P. vivax populations are consistent with an �out of Asia� origin for P. vivax as hominoid parasite. The current debate regarding how the Duffy negative trait became fixed in Africa needs to be revisited, taking into account not only human genetic data but also the genetic diversity observed in the extant P. vivax populations and the phylogeny of the genus Plasmodium.�
* Trichomonad gastritis in rhesus macaques (Macaca mulatta) infected with simian immunodeficiency virus. Kondova, I., Simon, M. A., Klumpp, S. A., MacKey, J., Widmer, G., Domingues, H. G., Persengiev, S. P., & O�Neil, S. P. (S. O., Div. of Comp. Pathol., New England PRC, Harvard Med. School, One Pine Hill Dr., P.O. Box 9102, Southborough, MA 01772-9102 [e-mail: shawn_oneil@ hms.harvard.edu]). Veterinary Pathology, 2005, 42, 19- 29.
. . . In a retrospective study, 51 cases of gastritis (14%) were identified from among 341 necropsies performed on simian immunodeficiency virus (SIV)-infected rhesus macaques at the New England Primate Research Center from 1993 to 2001. Protozoa were seen in the stomach of 13 monkeys (25%) with gastritis. Two histopathologic manifestations of gastritis were observed: seven cases of lymphoplasmacytic gastritis with trichomonad trophozoites within lumens of gastric glands and four cases of necrosuppurative gastritis containing intralesional periodic acid-Schiff-positive protozoa; two cases of gastritis had morphologic features of both types of gastritis. In instances of necrosuppurative and combined lymphoplasmacytic and necrosuppurative gastritis, protozoa were 4-35 �m in diameter and round to tear-shaped. Because of the unusual morphology of the protozoa in these latter cases, transmission electron microscopy and polymerase chain reaction (PCR) were used to further identify these organisms. The protozoa were definitively identified as Tritrichomonas in all cases on the basis of ultrastructural characteristics (flagella and undulating membranes) and amplification of a 347-bp product of the 5.8S ribosomal RNA gene of Tritrichomonas foetus, T. suis, and T. mobilensis by PCR using DNA extracted from stomach tissue. On the basis of these observations, it is concluded that Tritrichomonas can be a significant cofactor in the development of necrosuppurative gastritis in SIV-infected rhesus macaques.
* Ovarian epithelioid trophoblastic tumor in a cynomolgus monkey. Giusti, A. M. F., Terron, A., Belluco, S., Scanziani, E., & Carcangiu, M. L. (Preclinical Development, Nerviano Med. Sci., Via le Pasteur 10, 20014 Nerviano, Milano, Italy [e-mail: [email protected]]). Veterinary Pathology, 2005, 42, 223-226.
. . . Epithelioid trophoblastic tumor is an unusual type of trophoblastic tumor, with features resembling carcinoma. In this study, we describe a 4-year-old cynomolgus monkey (Macaca fascicularis) showing, at necropsy, a lobulated mass replacing the left ovary and several nodular lesions within the lungs. Histologically, the mass in the ovary and lung metastases were characterized by nests of epithelioid cells, with intermingled, occasional, multinucleate tumor cells consistent with syncytiotrophoblasts and moderate amount of eosinophilic, hyaline-like material. Immunohistochemically, the tumor cells were diffusely positive for cytokeratins (AE1/AE3) and inhibin- , but only focal immunoreactivity was observed for human chorionic gonadotropin, whereas placental alkaline phosphatase was always negative. On the basis of morphology and immunohistochemical reactivity, tumor cells were identified as intermediate trophoblast.
* Ovarian choriocarcinoma in a rhesus monkey associated with elevated serum chorionic gonadotropin levels. Farman, C. A., Benirschke, K., Horner, M., & Lappin, P. (California Animal Health and Food Safety Lab., West Health Sciences Dr., UC-Davis, Davis, CA 95617-1770 [e-mail: [email protected]]). Veterinary Pathology, 2005, 42, 226-229.
. . . A clinically normal, 3-year-old female rhesus monkey (Macaca mulatta), which was part of a routine toxicology study, had a mass in the right ovary with metastases to the adjacent mesentery and lungs. The histologic features and immunohistochemistry results were consistent with the diagnosis of choriocarcinoma. Neoplastic cell types included cytotrophoblast (positive for cytokeratin), syncytiotrophoblast (positive for human chorionic gonadotropin), and extravillous trophoblast (positive for human placental lactogen). Because the neoplasm was present in the ovary, the uterus was normal, and the animal was not currently pregnant, this was considered a primary ovarian neoplasm of germ cell origin. The monkey had elevated serum levels of chorionic gonadotropin at the beginning of the study, indicating that, as in women, choriocarcinomas in monkeys can be associated with increased gonadotropin levels and that the tumor was preexisting at the start of the toxicology study.
Evolution, Genetics, and Taxonomy
* Molecular evidence for deep phylogenetic divergence in Mandrillus sphinx. Telfer, P. T., Souquire, S., Clifford, S. L., Abernethy, K. A., Bruford, M. W., Disotell, T. R., Sterner, K. N., Roques, P., Marx P. A., & Wickings, E. J. (Dept of Anthropology, New York Univ., 25 Waverly Pl., New York, NY 10003; [e-mail: [email protected]]). Molecular Ecology, 2003, 12, 2019-2024.
. . . Mandrills are forest primates indigenous to western central Africa. Phylogenetic analysis of 267 base pairs of the cytochrome b gene from 53 mandrills of known and 17 of unknown provenance revealed two phylogeographical groups, with haplotypes differentiated by 2.6% comprising seven synonymous transitions. The distribution of the haplotypes suggests that the Ogoou� River, Gabon, which bisects their range, separates mandrill populations in Cameroon and northern Gabon from those in southern Gabon. The haplotype distribution is also concordant with that of two known mandrill simian immunodeficiency viruses, suggesting that these two mandrill phylogroups have followed different evolutionary trajectories since separation.
* Craniodental variation in the African macaque, with reference to various Asian species. Pan, R., & Oxnard, C. (School of Anatomy & Human Biology, Fac. of Health Sciences, Univ. Witwatersrand, 7 York Rd., Park Town 2193, Johannesburg, South Africa [e-mail: [email protected]]). Folia Primatologica, 2004, 75, 355-375.
. . . Based on twenty-seven craniodental measurements and ratios derived from them, the relationship between the African macaque (M. sylvanus) and the Asian macaques were examined with principal components analyses (PCA) and Euclidean distance analysis based upon prior discriminant function analyses (DFA). Results based on analyses of raw measurements indicate that the variation between species lies in the first axis of PCA; the species are dispersed according to their differences in size. The variation between sexes (sexual dimorphism) lies in the second axis. In the analyses of ratio variables, though these two patterns of separation remain orthogonal, they lie at approximately forty-five degrees to each axis. Variables relating to anterior teeth were found to play an important role in variation analysis, and this may be related to the special food preferences of these monkeys: more frequent usage of the incisor teeth for processing frugivorous diets than in other primates that are mainly folivorous. The results from Euclidean distance analyses indicate that the average distance of species within the Asian group is shorter than that between Asian and African groups regardless of sex and variable type. Also, the variation between African and Asian groups is larger than that within the Asian group. Thus, it is reasonable to suggest that the African macaque has a range of measurements and ratios quite distinct from the species found in Asia (though the greatest separations result from the analyses of ratio data). These results support the view that M. sylvanus may be regarded as an independent species group in the genus Macaca, as proposed by Delson.
* Accelerated evolution of nervous system genes in the origin of Homo sapiens. Dorus, S., Vallender, E. J., Evans, P. D., Anderson, J. R., Gilbert, S. L., Mahowald, M., Wyckoff, G. J., Malcom, C. M., & Lahn, B. T. (B. T. L., Howard Hughes Medical Inst., Dept of Human Genetics, Univ. of Chicago, Chicago, IL 60637 [e-mail: [email protected]]). Cell, 2004, 119, 1027-1040.
. . . �Human evolution is characterized by a dramatic increase in brain size and complexity. To probe its genetic basis, we examined the evolution of genes involved in diverse aspects of nervous system biology. We found that these genes display significantly higher rates of protein evolution in primates than in rodents. Importantly, this trend is most pronounced for the subset of genes implicated in nervous system development. Moreover, within primates, the acceleration of protein evolution is most prominent in the lineage leading from ancestral primates to humans. Thus, the remarkable phenotypic evolution of the human nervous system has a salient molecular correlate, i.e., accelerated evolution of the underlying genes, particularly those linked to nervous system development. In addition to uncovering broad evolutionary trends, our study also identified many candidate genes - most of which are implicated in regulating brain size and behavior - that might have played important roles in the evolution of the human brain.�
* Early Pliocene hominids from Gona, Ethiopia. Semaw, S., Simpson, S. W., Quade, J., Rennes, P. R., Butler, R. F., McIntosh, W. C., Levin, N., Dominguez-Rodrigo, M., & Rogers, M. J. (CRAFT Stone Age Inst., Indiana Univ., 1392 W. Dittemore Rd, Gosport, IN 47433-9531 [e-mail: [email protected]]). Nature, 2005, 433, 301-305.
. . . Comparative biomolecular studies suggest that the last common ancestor of humans and chimpanzees, our closest living relatives, lived during the Late Miocene-Early Pliocene. Fossil evidence of Late Miocene-Early Pliocene hominid evolution is rare and limited to a few sites in Ethiopia, Kenya and Chad. Here we report new Early Pliocene hominid discoveries and their paleoenvironmental context from the fossiliferous deposits of As Duma, Gona Western Margin (GWM), Afar, Ethiopia. The hominid dental anatomy (occlusal enamel thickness, absolute and relative size of the first and second lower molar crowns, and premolar crown and radicular anatomy) indicates attribution to Ardipithecus ramidus. The combined radioisotopic and paleomagnetic data suggest an age of between 4.51 and 4.32 million years for the hominid finds at As Duma. Diverse sources of data (sedimentology, faunal composition, ecomorphological variables and stable carbon isotopic evidence from the paleosols and fossil tooth enamel) indicate that the Early Pliocene As Duma sediments sample a moderate rainfall woodland and woodland/grassland.
* Mitochondrial DNA variation in Chinese and Indian rhesus macaques (Macaca mulatta). Smith, D. G., & McDonough, J. (Dept of Anthropology, U.C.-Davis, Davis, CA 95616 [e-mail: [email protected]]). American Journal of Primatology, 2005, 65, 1-25.
. . . DNA was extracted from the buffy coats or serum of 212 rhesus macaques sampled throughout the species� geographic range. An 835 base pair (bp) fragment of mitochondrial DNA (mtDNA) was amplified from each sample, sequenced, aligned, and used to estimate genetic distances from which phylogenetic trees were constructed. A tree that included sequences from rhesus macaques whose exact origins in China are known was used to determine the regional origin of clusters of haplotypes, or haplogroups, defined by the trees. Indian rhesus sequences formed one large homogeneous haplogroup with very low levels of nucleotide diversity and no geographic structure, and a second much smaller haplogroup apparently derived from Burma. The sequences from Burma and eastern and western China were quite divergent from those in the major haplogroup of India. Each of these sequences formed separate clusters of haplotypes that exhibited far greater nucleotide diversity and/or population structure. Correspondingly, sequences from Indian rhesus macaques that are considered to represent different subspecies (based on morphological differences) were intermingled in the tree, while those from China reflected some, but not all, aspects of subspecific taxonomy. Regional variation contributed 72% toward the paired differences between sequences in an analysis of molecular variance (AMOVA), and the average differences between the populations of eastern and western China were also statistically significant. These results suggest that Indian and Chinese rhesus macaques were reproductively isolated during most, if not all, of the Pleistocene, during which time Indian rhesus macaques experienced a severe genetic bottleneck, and that some gene flow westward into India was subsequently reestablished. Samples from breeding centers in three different provinces of China included sequences from rhesus macaques that originated in both eastern (or southern) and western China, confirming anecdotal reports that regional breeding centers in China exchange breeding stock. Genetic differences among rhesus macaques (even those acquired from the same regional breeding center) that originate in different geographic regions and are employed as subjects in biomedical experiments can contribute to phenotypic differences in the traits under study.
* Stratigraphic placement and age of modern humans from Kibish, Ethiopia. McDougall, I., Brown, F. H., & Fleagle, J. G. (Res. School of Earth Sci., ANU, Canberra, ACT 0200, Australia [e-mail: ian.mcdougall@ anu.edu.au]). Nature, 2005, 433, 733-736.
. . . �In 1967 the Kibish Formation in southern Ethiopia yielded hominid cranial remains identified as early anatomically modern humans, assigned to Homo sapiens. However, the provenance and age of the fossils have been much debated. Here we confirm that the Omo I and Omo II hominid fossils are from similar stratigraphic levels in Member I of the Kibish Formation, despite the view that Omo I is more modern in appearance than Omo II. 40Ar/39Ar ages on feldspar crystals from pumice clasts within a tuff in Member I below the hominid levels place an older limit of 198�14 kyr (weighted mean age 196�2 kyr) on the hominids. A younger age limit of 104�7 kyr is provided by feldspars from pumice clasts in a Member III tuff. Geological evidence indicates rapid deposition of each member of the Kibish Formation. Isotopic ages on the Kibish Formation correspond to ages of Mediterranean sapropels, which reflect increased flow of the Nile River, and necessarily increased flow of the Omo River. Thus the 40Ar/39Ar age measurements, together with the sapropel correlations, indicate that the hominid fossils have an age close to the older limit. Our preferred estimate of the age of the Kibish hominids is 195�5 kyr, making them the earliest well-dated anatomically modern humans yet described.�
Instruments and Techniques
* Interpretation of two-stage experiments in animal studies. Horgan, G. W. (Biomath. & Statistics Scotland, Rowett Research Inst., Aberdeen AB21 9SB, U.K. [e-mail: [email protected]]). Laboratory Animals, 2005, 39, 75-79.
. . . Two-stage experiments allow results to be analyzed at the end of the first stage and the second stage to be omitted if the preliminary conclusions are clear-cut, offering the potential to reduce the number of experimental animals. However, using standard P values to assess the significance of results at the end of either stage will lead to an increase in risk of false positive conclusions. This paper presents a possible protocol for two-stage experiments and a method for adjusting P values. It is shown that, for experiments with reasonable power (>80%), the expected reduction in animal numbers will be at least 20%.
* Temperament correlates with training success in adult rhesus macaques. Coleman, K., Tully, L. A., & McMillan, J. L. (Oregon NPRC, 505 NW 185th Ave., Beaverton, OR 97006 [e-mail: [email protected]]). American Journal of Primatology, 2005, 65, 63-71.
. . . �In recent years there has been a marked increase in awareness of issues involving the psychological well-being of nonhuman primates (NHPs) used in biomedical research. As a result, many facilities are starting to train primates to voluntarily cooperate with veterinary, husbandry, and research procedures, such as remaining still for blood draws or injections. Such training generally reduces the stress associated with these procedures, resulting in calmer animals and, ultimately, better research models. However, such training requires great investments in time, and there can be vast individual differences in training success. Some animals learn tasks quickly, while others make slower progress in training. In this study, we examined whether temperament, as measured by response to a novel food object, correlated with the amount of time it took to train 20 adult female rhesus macaques to perform a simple task. The monkeys were categorized as exploratory (i.e., inspected a novel object placed in the home cage within 10 sec), moderate (i.e., inspected the object within 10-180 sec), or inhibited (i.e., did not inspect the object within 3 min). We utilized positive reinforcement techniques to train the monkeys to touch a target (PVC pipe shaped like an elbow) hung on their cage. Temperament correlated with training success in this study (Pearson 2=7.22, df=2, P=0.03). We easily trained over 75% of the animals that inspected the novel food (i.e., exploratory or moderate individuals) to touch the target. However, only 22% of the inhibited monkeys performed the task. By knowing which animals may not respond to conventional training methods, we may be able to develop alternate training techniques to address their specific needs. These results will also allow us to screen monkeys to be assigned to research projects in which they will be trained, with the goal of obtaining the best candidates for those studies.�
Reproduction
* Two breeding females within free-living groups may not always indicate polygyny: Alternative subordinate female strategies in common marmosets (Callithrix jacchus). Arruda, M. F., Ara�jo, A., Sousa, M. B. C., Albuquerque, F. S., Albuquerque, C. S. R., & Yamamoto, M. E. (P.O. Box 1511, Campus UFRN, Natal, RN 59078-970, Brasil [e-mail: [email protected]]). Folia Primatologica, 2005, 76, 10-20.
. . . �The mating system of callitrichids has been reported to be monogamous, polygynous, and polyandrous. In Callithrix jacchus, groups with two breeding females and groups with one breeding female have been reported. Our purpose was to evaluate the occurrence of occasional reproduction by subordinate females in free-ranging C. jacchus groups characterized as monogamous. Four groups were monitored at a field site of IBAMA-Brazil for between 20 and 72 months. We recorded the birth of 7 sets of twins to subordinate females that had never reproduced before. Sexual activities were recorded opportunistically: dominant females copulated with only the resident male, while subordinates copulated with extra-group males. We suggest these were essentially monogamous groups that occasionally had two reproductive females. Between-group copulations seem to be an alternative strategy used by the subordinates. Despite the costs, there would probably be benefits as a result of scanning for vacancies for reproductive positions in neighboring groups and the establishment of ties with extra-group males that might become reproductive partners in the future.�
* Reproductive parameters and life-history variables in captive golden-bellied mangabeys (Cercocebus agilis chrysogaster). Walker, S. E., Strasser, M. E., & Field, L. P. (Dept of Sociology & Anthropology, Southwest Missouri State Univ., Springfield, MO 65804 [e-mail: [email protected]]). American Journal of Primatology, 2004, 64, 123-131.
. . . �An understanding of the reproductive physiology of captive primates is vital for their successful management. We report on reproductive parameters and life-history variables collected at the Sacramento Zoo for five female golden-bellied mangabeys over a 7-year period. For each female, we collected data on their sexual skins, menses, gestation, and other pregnancy-related factors, such as postconception bleeding and swelling. We used these data to estimate life-history variables, such as age at onset of estrus, menses, and conception, as well as the duration of intervals between births, and between parturition and the resumption of cycling. Cercocebus agilis chrysogaster is comparable to other Cercocebus species in terms of reproductive parameters, although variability is exhibited within and among female subjects. In some cases, it appears that stressful incidents altered cycle length or halted cycling altogether. We suggest the use of husbandry practices that include consistent data collection to enhance regularity in reproductive cycles, and maximize captive breeding success for this rare species.�
* Neuroendocrine response to female ovulatory odors depends upon social condition in male common marmosets, Callithrix jacchus. Ziegler, T. E., Schultz-Darken, N. J., Scott, J. J., Snowdon, C. T., & Ferris, C. F. (Wisconsin NPRC, 1220 Capitol Ct, Madison, WI 53715). Hormones and Behavior, 2005, 47, 56-64.
. . . �Male mammals show rapid behavioral and hormonal responses to signals from sexually receptive females. However, rapid endocrine responses to female signals have not been observed in a nonhuman primate. Here, we tested the behavioral and hormonal response of male common marmosets to isolated scent secretions from ovulatory females or to vehicle control scent. Fifteen males were tested in their home cage for behavioral and hormonal responses. These males showed increased investigative and arousal behaviors to the ovulatory scent compared to the vehicle scent. Time sniffing the scent substrate and the duration of erections were significantly elevated in relation to the vehicle scent. Thirty minutes after presentation of ovulatory scent, males showed a significant increase in testosterone compared to the vehicle, but there was no difference in cortisol values. To better control for scent presentation, 15 additional males were tested under a controlled scent exposure. Current social housing condition influenced the male's testosterone response to the ovulatory scent. Single and paired males showed significant increases in testosterone levels with the ovulatory scent but did not increase cortisol levels. Single males also showed the highest change in testosterone with the ovulatory scent, but fathers showed no changes. These results indicate that a rapid hormonal response to sexually arousing cues occurs in marmosets, and the data suggest that a male's social condition influences how he responds to sexually relevant cues.�
* * *
------------------------------
We would like to acknowledge Primate-Science as a source for information about new books.
------------------------------
* * *
Contents of Volumes 40-43 (2001-2004)
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; FAX: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
Current and back issues of the Newsletter are available on the World Wide Web at
http://www.brown.edu/primate
The Newsletter is supported by U. S. Public Health Service Grant RR-00419 from the Comparative Medicine Program, National Center for Research Resources, N.I.H.
Cover illustration of Hylobates lar by Anne Richardson, in memory of Beanie.
Copyright (c) 2005 by Brown University
Last updated: March 23, 2005
{kind=link}
{kind=link}
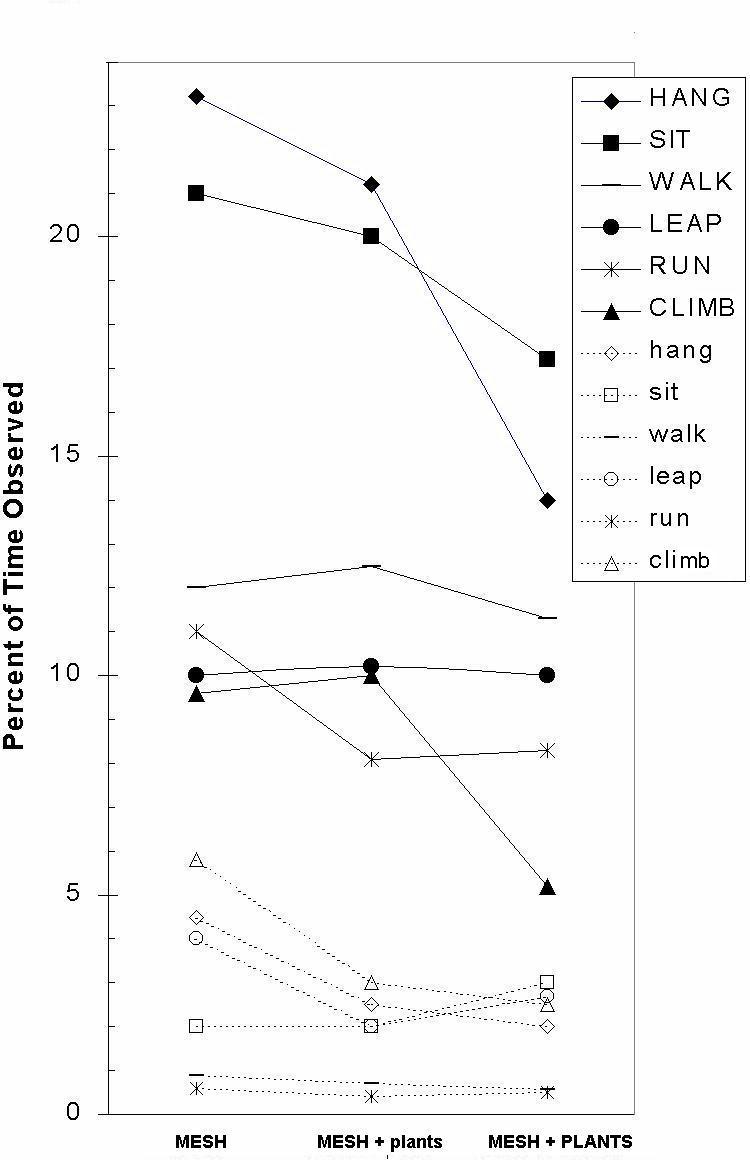{kind=link}
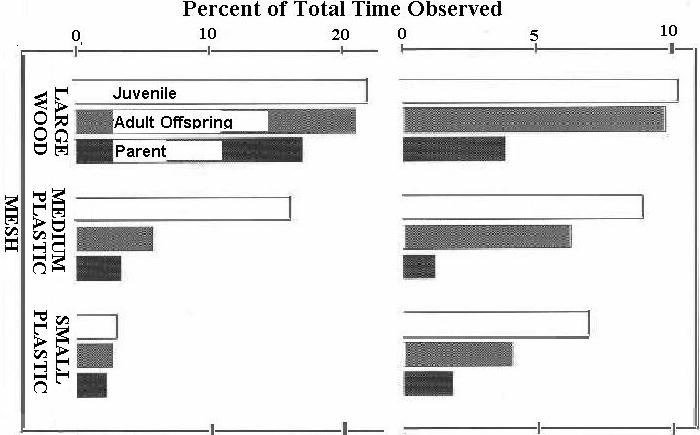{kind=link}
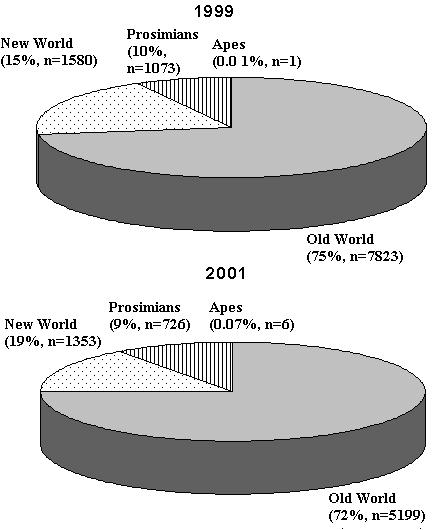{kind=link}
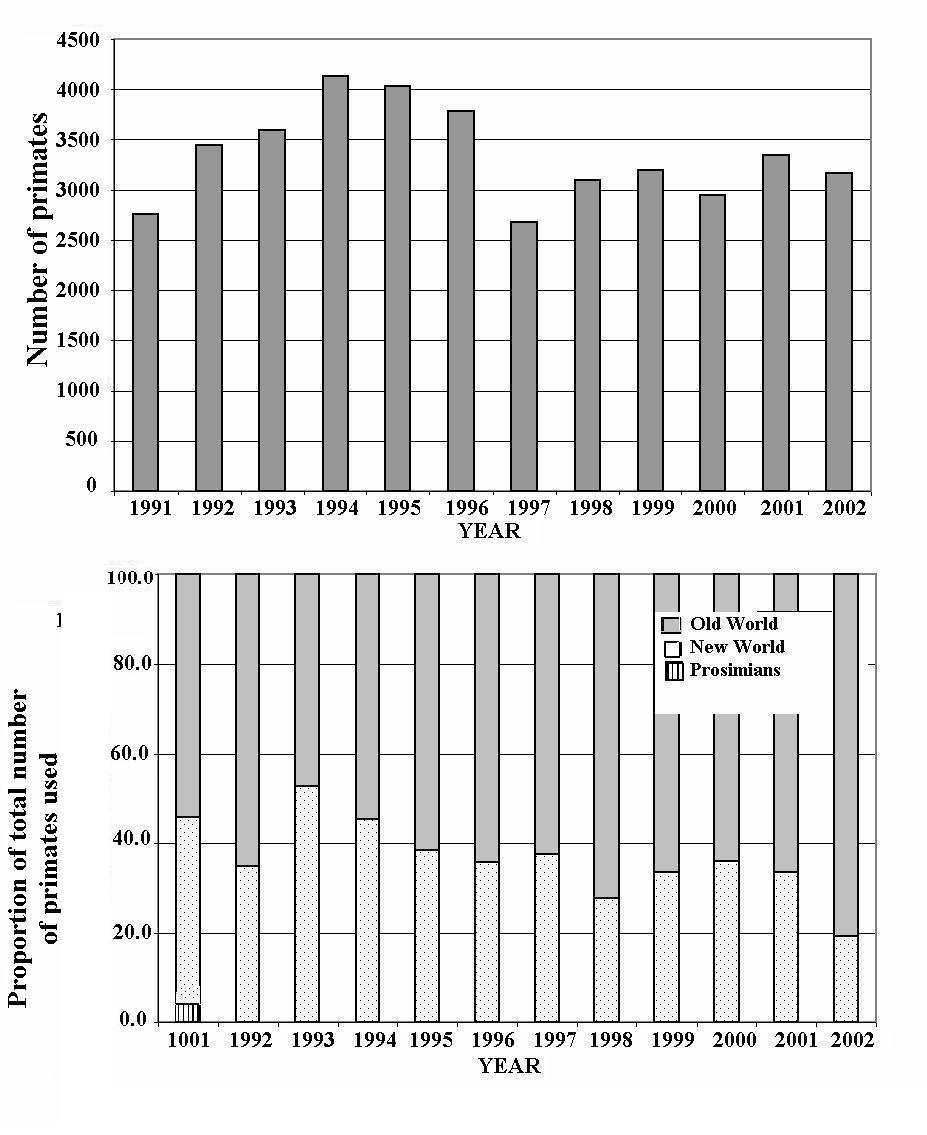{kind=link}
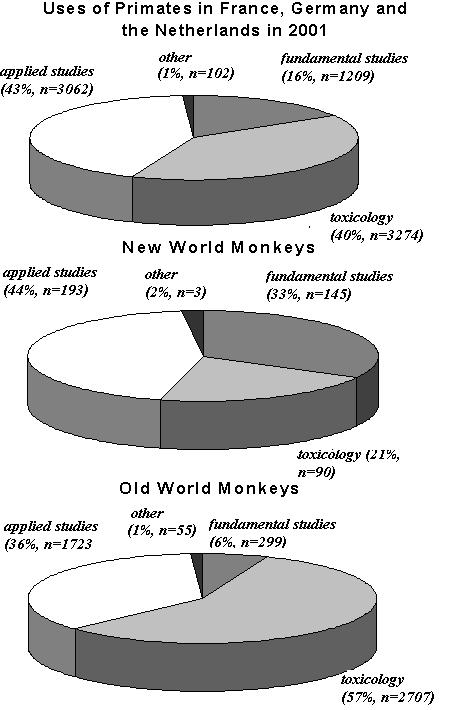{kind=link}
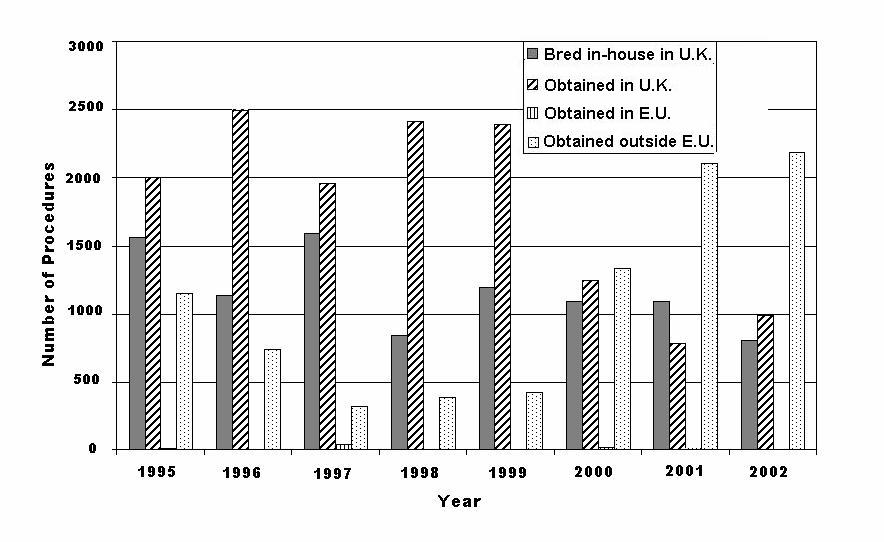{kind=link}
{kind=link}
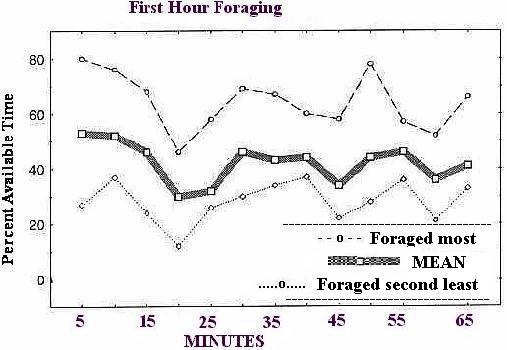{kind=link}