 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 45 NUMBER 1 JANUARY 2006
Printable (PDF) Version of this issue
Articles and Notes
Observations of Meat Eating by Captive Juvenile Macaques, by C. A. D. Nahallage & M. A. Huffman......1
Exploring the Utility of Van Schaikís Model of Male Coalitions for Female Mantled Howler Monkeys (Alouatta palliate), by C. B. Jones......4
Environmental Enrichment Objects for the Improvement of Locomotion of Caged Rhesus Macaques (Macaca mulatta), by R. Plesker, J. Heller-Schmidt, & H. Hackbarth......7
Pair Formation and Reintroduction of Temporarily Separated Partners: A Discussion on the Laboratory Animal Refinement & Enrichment Forum......11
News, Information, and Announcements
Updating Standards for the Care and Use of Laboratory Animals......3
Meeting Announcements......6
ASP 2006 Conservation Small Grant Applications......10
Announcements from Publications: Australasian Primatology......12
Travel-Associated Dengue Infections ñ United States, 2001-2004......13
Workshop Announcements......14
PASA 4th Veterinary Healthcare Workshop; Laboratory Animal Disease Workshop; Captive Breeding and Reintroduction; Teaching Research Ethics; Second Zoo Animal Training Workshop
News Briefs......15
Zoo Atlanta Gorilla Gives Birth to Twins; Research Center for Primates Opens in Korea; Tulane Primate Center Thrives and Grows; Gorilla Haven Joins the AZA; Mountain Gorilla Veterinary Project Moves; The Larry Jacobsen Primate Library!
Volunteer Opportunities......16
Gibbon Conservation Center ñ California; Hurricane Wilma Recovery ñ Florida
Resources Wanted and Available......17
IVIS Books Online; Primate Treat Recipe Cookbook Available in pdf; New Software Release by Noldus; Training and Enrichment Talks
Information Requested or Available......18
NIH is Ready for Electronic Submission; Biographical Sketches: Evolutionists and Ecologists; ìVirtual Primate Museumî; USDA Releases Report on Animal Care Program; FY2004 USDA/APHIS Animal Care Report Online; Change to USDA Procedures on Annual Reports; Exchange of Scientific Papers; Conference Report: Monkeys Old and New; More Interesting Websites
Italian Society for Evolutionary Biology......19
Educational and Research Opportunities......21
New World Primate Caregiver/Office Assistant; Summer Apprentice Program ñ Washington State
2006 Directory of Graduate Programs in Primatology and Primate Research...... 37
Awards Granted at 2005 American Society of Primatologists Meeting......46
Duane Rumbaugh, Distinguished Primatologist; Education Committeeís Student Prize Award Winners; Research and Development Grant Award Winners
SCAW Presents Harry C. Rowsell Award......46
Departments
Positions Available......20
Environmental Enrichment Specialist ñ Tulane; Quality Assurance Specialist ñ Tulane; Primate Veterinarian ñ Alice, Texas; Evolutionary Ecologist ñ Binghamton
Primates de las AméricasÖLa Página......22
Recent Books and Articles......23
* * *
Observations of Meat Eating by Captive Juvenile Macaques
Charmalie A. D. Nahallage and Michael A. Huffman
Primate Research Institute, Kyoto University
Introduction
In most wild nonhuman primate species, the diet consists mainly of fruits, berries, grains, leaves, buds, seeds, flowers, bark, and a very small amount of animal matter, mainly invertebrates. Animal matter is an important source of protein for these animals, yet they spend less time feeding on it than on plants (Harding, 1975; Hill, 1997; Teleki, 1981).
Hunting of vertebrates, unlike invertebrates, by nonhuman primates is not common; systematic hunting of vertebrates has been recorded only in baboons and chimpanzees (baboons: Harding, 1975; Strum, 1981; chimpanzees: Boesch, 1994a, 1994b; Teleki, 1973). Systematic hunting of vertebrates by wild macaques has not yet been reported, and so all of the captures described to date have been fortuitous. Major animal components in the diet of macaques includes insects and other small invertebrates (spiders, shellfish, and water snails) when available, and only very rarely small vertebrates and eggs.
There arewere numerous reports of meat-eating behavior of macaques in the wild (Estrada & Estrada, 1977; Krishnamani, 1996; Kumara et al., 2000; Suzuki et al., 1991). However, records of meat-eating in captive macaques are few, and Caine et al. (1979) stressed the need to keep records on incidents of predatory behavior in captive troops. In captivity animals have less access to the outside environment, which limits their contact with other animals.
This report describes two incidents of bird eating recorded in two captive macaque species in Kyoto Universityís Primate Research Institute (PRI), Inuyama, Japan.
Study troops
The two captive troops are housed in two separate outdoor enclosures. These enclosures are surrounded by walls about three meters high; electric wires are attached to the uppermost part of the walls to ensure that monkeys cannot escape. Since there are no overhead coverings, birds can fly into the enclosure freely to feed on monkey chow, which is given once a day. Birds that come to feed on the monkey chow are tree sparrows (Passer montanus saturatus), pigeons (Columba livia), and crows (Corvus corone). The Takahama troop, which consists of 46 Japanese macaques (Macaca fuscata) of both sexes and various ages, is housed in a 960 m2 outdoor enclosure. The rhesus (Indian Macaca mulatta) troop, 37 monkeys of both sexes and various ages, is housed in a 680 m2 outdoor enclosure.
I (C. N.) have been studying the stone-handling behavior (e.g. Huffman, 1996) of these two groups, hoping to do an inter-species comparison of that behavior, and of its seasonal and diurnal variation. I videotape focal observations of all the individuals, 5 days a week, from 7:00 a.m. until dark. In the end all individuals were sampled equally at all times of day in every season. The Takahama troop was observed from June, 2003, to November, 2005; the rhesus troop from November, 2003, to September, 2004, again starting from May, 2005, and will continue until December, 2005. The incidents described below occurred while I was doing focal observations on other individuals in each group. When I noticed what was happening, I turned the video recorder toward the events described.
Observations
Case 1: Japanese macaque (Macaca fuscata). Duration of observation: 17:46 ñ 17:59 hr.
On the 10th of June, 2003, around 17:46 hr, an adult tree sparrow fell from a tree located in the middle of the enclosure. My attention was drawn to it by the movement of monkeys in the enclosure. , The alpha male of the troop came and sniffed the bird, but did not touch it. Later, some juveniles also came to investigate, but none touched it. Then a juvenile (the two-year-old daughter of the beta female) came, pushed the bird about, and started to take pull off feathers off the bird, sometimes using her hands and sometimes her mouth. While she was performingdoing this a one- year- old female and an infant male looked on, but did not participate. None of the adults or sub-adults showed any interest ion the bird. The juvenile female dragged the bird from place to place and kept taking feathers off. The bird was trying to escape by hopping away, since it could not fly. The juvenile female always caught up with it and dragged it around the enclosure as a plaything but did notwas not observed eating its flesh. By theut next morning, however, only the feathers was remained in a corner of the enclosure. It was assumed to have been eaten by the monkeys.

Figure 1: Japanese macaque juvenile with tree sparrow.
Case 2: Rhesus macaque (Macaca mulatta). Duration of observation: 17:07 ñ 17:12 hr.
On the 1st of July, 2005, around 17:07 hr, a tree sparrow that was resting on one of the walls fell to the ground; my attention was drawn to it because monkeys ran towards it. , Tthere was a fight over the bird among juveniles and somehow onea juvenile female ran away with it onto the wooden logs at the center of the enclosure, followed by the others. As she was about to feed on the bird it slipped through her fingers. Then the three-year-old son of the alpha female took the bird and started eating it. First he bit on the head and then turned it to the other side and started eating the belly. , Hhe ate all parts of the bird except the feathers and feet. Theseis he threw away, while the other juveniles looked at him from time to time but did not approach him. , Aas in the earlier case, the adults did not show any interest ion the bird.

Figure 2: Rhesus macaque juvenile with tree sparrow.
I had seen the remains of pigeon feathers in the rhesus enclosure a few times, but had never directly observed a monkey capturing or eating pigeons. When I asked the keepers of the enclosure about this, they confirmed that the monkeys have been observed eating both tree sparrows and pigeons in the past.
Discussion
Most of the cases that have been reported for vertebrate predation in captivity were fortuitous (Caine et al., 1979; Kessler et al., 1980). The occurrence of this behavior in captive conditions could not be due to hunger, as food pellets remain in the enclosure for most of the day, but may be due to the novelty of the food item itself, or an occasional desire for variety in the diet, including animal protein.
In the other reported cases of captive macaque troops, hunting was displayed by adult dominant individuals and mainly by females, as in wild macaque species. However in the two incidents reported here, juveniles were responsible for both and adults showed no interest in the prey. Birds are regularly seen flying into these enclosures to feed on the monkey chow, and most of the time they are not disturbed by monkeys, They are not afraid of monkeys and eat near them. Occasionally a juvenile monkey may try to chase a bird away, but we had never observed any monkey chasing a bird with the intention of killing it. When we asked other researchers, one said that she had observed a juvenile Japanese monkey from the same troop taking pulling feathers off from a pigeon a few years ago (K. Chatani, personal communication). This feeding habit in captive primates housed outdoors should be closely monitored, given current global concerns about the spread of avian influenzas.
Editorís Note: In contrast, my experience with macaques, in particular rhesus, housed outside at Bowman Gray School of Medicine, North Carolina, is that they readily chased, caught, and ate any bird that entered their enclosure.Ý We also felt that this was frequently the mode of transmission of yersiniosis in these monkeys. ñ Larry Hulsebos
References
Boesch, C. (1994a). Chimpanzeesñred colobus monkeys: A predatorñprey system. Animal Behaviour, 47, 1135-1148.
Boesch, C. (1994b). Cooperative hunting in wild chimpanzees. Animal Behaviour, 48, 653-667.
Caine, N., Zanovitz, C., vanTassel, J., Yee, K., & Mitchell, G. (1979). Predatory behavior in a captive rhesus monkey. Laboratory Primate Newsletter, 18 [1], 25-26.
Estrada, A., & Estrada, R. (1977). Patterns of predation in a free-ranging troop of stump-tailed macaques (Macaca arctoides). Relations to the ecology II. Primates, 18, 633-646.
Harding, R. S. O. (1975). Meat eating and hunting in baboons (Papio anubis). In R. Tuttle, (Ed.), Sociology and psychology of primates (pp. 245-257). Den Haag: Mouton.
Hill, D. A. (1997). Seasonal variation in the feeding behavior and diet of Japanese macaques (Macaca fuscata yakui) in lowland forest of Yakushima. American Journal of Primatology, 43, 305-322.
Huffman, M. A. (1996). Acquisition of innovative cultural behaviors in nonhuman primates: A case study of stone handling, a socially transmitted behavior in Japanese macaques. In C. M. Heyes & B. G. Galef, Jr., (Eds.), Social learning in animals: The roots of culture (pp. 267-289). San Diego: Academic Press.
Kessler, M. J., Brown, R. J., & O`Neil, T. P. (1980). Predation of birds by gang-caged Rhesus monkeys. Laboratory Primate Newsletter, 19 [2], 9.
Kumara, H. N., Singh, M. E., Sharma, A. K., Singh, M. R., & Anandakumar, M. (2000). Faunal component in the diet of lion-tailed macaque. Primate Report, 58, 57-65.
Krishnamani, R. (1994). Diet composition of the bonnet macaque (Macaca radiata) in a tropical dry evergreen forest of southern India. Tropical Biodiversity, 2, 285-297.
Strum, S. (1981). Processes and products of change: Baboon predatory behavior at Gilgil, Kenya. In R. S. O. Harding & G. Teleki (Eds.), Omnivorous primates: Gathering and hunting in human evolution (pp. 255-302). New York: Columbia University Press.
Suzuki, S., Hill, D. A., Maruhashi, T., & Tsukahara, T. (1990). Frog- and lizard-eating behavior of wild Japanese macaques in Yakushima, Japan. Primates, 3, 421-426.
Teleki, G. (1981). The omnivorous diet and eclectic feeding habits of chimpanzees in Gombe National Park, Tanzania. In R. S. O. Harding & G. Teleki (Eds.), Omnivorous primates: Gathering and hunting in human evolution (pp. 303-343). New York: Columbia University Press.
------------------------------ Authorsí address: Primate Research Institute, Kyoto University, 41-2 Kanrin, Inuyama, Aichi 484, Japan [e-mail: [email protected]].
------------------------------
* * *
Updating Standards for the Care and Use of Laboratory Animals
The National Institutes of Health is exploring the need to update the laboratory animal welfare standards of the Guide for the Care and Use of Laboratory Animals (Guide) . As a first step, the Office of Scientific Affairs (OSA) is seeking to identify new scientific information that might warrant NIH issuing a contract for a new or updated edition of the Guide.
The purpose of the Guide is to assist institutions in caring for and using animals in ways judged to be scientifically, technically, and humanely appropriate. Further, recommendations of the Guide are based on published data, scientific principles, expert opinion, and experience with methods and practices that have proved to be consistent with high-quality humane care and use.
Our intent is to solicit new scientifically valid information, methods or practices, published data, or other advances in the humane care and use of laboratory animals, in order to assess the need for updating the standards in the current 1996 Guide. NIH is interested in new information and knowledge related to the four chapters of the 1996 Guide: Institutional policies and responsibilities; Animal environment, housing, and management; Veterinary medical care; and Physical plant. In particular, new scientific information related to the following topics is solicited: ∑ The macro- and microenvironment of animal facilities. ∑ Housing for laboratory animals, including space, temperature and humidity, ventilation, acoustics, and illumination. ∑ Structural and social environment of animals. ∑ Husbandry, sanitation and pest control. ∑ Disease and disease manifestations in laboratory animals. ∑ Population management of genetically modified animals. ∑ Physical plant standards. ∑ The topics listed in Appendix A, Selected Bibliography, of the 1996 Guide.
Interested parties, individuals, and organizations may submit any of the following: ∑ articles or citations for articles published in reputable peer-reviewed scientific journals since the development of the 1996 Guide; ∑ science-based information or scientific principles concerning the humane care and use of laboratory animals developed and widely accepted by the research community and not addressed in the 1996 Guide; ∑ newly published science-based standards for animal environment, housing, management and structural design not cited in the 1996 Guide.
This Request for Information (RFI) is for information and planning purposes only and shall not be construed as a solicitation or as an obligation on the part of the Government. The Government does not intend to award a contract on the basis of responses nor otherwise pay for the preparation of any information submitted or the Governmentís use of such information. Acknowledgment of receipt of responses will not be made, nor will respondents be notified of the Governmentís evaluation of the information received. However, should such a requirement materialize, no basis for claims against the Government shall arise as a result of a response to this request for information or the Governmentís use of such information as either part of our evaluation process or in developing specifications for any subsequent requirement. Responses may be disclosed under the Freedom of Information Act. Any proprietary information should be so marked.
Responses should be identified with RFI No. NOT-OD-06-011. The deadline for submittal is February 28, 2006. Please submit three copies of your response to: Dr. Margaret Snyder, Director, Office of Scientific Affairs, Office of Extramural Research, OD, NIH, 6705 Rockledge I, Suite 4184, MSC 7983, Bethesda, MD 20892-7983 [e-mail: [email protected]]. For FedEx or courier, use: Rockville, MD 20817.
* * *
Exploring the Utility of Van Schaikís Model of Male Coalitions for Female Mantled Howler Monkeys (Alouatta palliata)
Clara B. Jones
Fayetteville State University and Community Conservation, Inc.
Introduction
Van Schaik and his colleagues (2004) proposed a general model (ìthe van Schaik Modelî, hereafter ìthe Modelî) of within-group coalitionary aggression for male primates comprising five ìbasic coalition typesî: (1) rank-changing coalitions targeting individuals ranking above all coalition partners; (2) rank-changing coalitions in which higher-rankers support lower-rankers to rise to a rank below themselves; (3) non-rank-changing coalitions expected to occur whenever high-ranking males have low-ranking close relatives; (4) non-rank-changing coalitions by high-rankers against lower-ranking targets; and, (5) non-rank-changing coalitions in which all partners rank below their target and which flatten the payoff distribution.
Van Schaik et al. (2004) and Jones (2005a) tested the Model and found it to be a relatively good fit to data existing in the literature. Jones (2005a) suggested that coalitions among both males and females may be viewed not only as indicators of reproductive competition but also as mutual policing and suppression of competition within groups. Indeed, there is no a priori reason that the Model will not apply to within-group coalitionary aggression among female primates, a view with which van Schaik concurs (C. P. van Schaik, personal communication, 2005).
The purpose of this communication is to evaluate the utility of the Model for within-group coalitions among adult female mantled howler monkeys (Alouatta palliate).
Methods, Study Site, and Animals
The study was conducted from January, 1976, to February, 1977, and in January, 1978, at Hacienda La Pacífica, Cañas, Guanacaste, Costa Rica (10∞18í N, 85∞07í W). Results are based upon randomized focal (Altmann, 1974) and ad lib. observations. Modal social organization of mantled howlers is multimale-multifemale, yielding a polygynandrous mating system (Jones, 1978, 1980, 1985, 2000). Two marked groups (see Jones, 2005b) were studied in two habitats of seasonal, tropical dry forest environment, riparian and deciduous (Frankie et al., 2004). Age of animals was estimated by tooth wear (see Jones, 2005b). Coalitions among females were observed only in the riparian habitat group (Group 5, 402 h of observation: Tables 1 and 2).
| 5-7 yr | 7-10 yr | 10-15 yr | >15 yr | Unknown |
| BC, GS, GRS, GWS, UM1 | RS, PS, YS | PY, RYS | SS | LL, UM2, UM3, |
Table 1: Females observed in Group 5, by age group.
| COALITION PARTNERS | TARGET FEMALE |
| GWS , SS | RYS ? |
| GWS, SS | RYS? |
| GWS, UM2 | PS |
| GS , YS | PS |
| GRS , GWS | PS |
| GWS, SS | GRS |
| LL , UM1 | SS? |
| GS, GWS | PS |
| PS, LL | SS? |
| GS, GRS | UM3 |
| GRS, GWS | LL |
| UM1, GS, MJ | GWS? |
| UM1, MJ | RS |
| UM1, MJ | BC? |
| UM1, MJ | PY |
| UM1, LI | PY |
Table 2: Female-female coalitions and female-infant or juvenile coalitions. Arrows indicate females who emigrated from Group 5 between January 1976 and August 1977. LI = large infant; MJ = medium juvenile. See Jones (1978, 1980) for further explanation and dominance hierarchies for three periods during the study.
Results
Twenty-four coalitions were observed in Group 5, twelve between adult females, four between an adult female and an infant or juvenile, and seven between adult males (Jones, 1980, pp. 396-397; Jones, 2005a). Coalitions among adults, then, were within-sex events (intrasexual selection). Coalitions involving adult females are presented in Table 2. The four coalitions between an adult female and an infant or juvenile always involved a female target subordinate to the adult female of the coalition partnership. Another coalition involved two adult females and a medium juvenile against a female dominant to either adult female of the coalition partnership. The eleven remaining coalitions were limited to two coalition partners against a third, lower-ranking target.
Discussion
For an accurate interpretation of female-female coalitionary aggression in A. palliata it is necessary to review the pattern of hierarchical dominance relations in this species. In tropical dry forest environment, mantled howler hierarchies are linear with few reversals. The pattern of relations is ìage-reversedî (Jones, 1978, 1980) whereby young adults (D1) are highest-ranked, middle-aged (D2) individuals are medium-ranked, and older individuals (D3 and D4) are ranked at the bottom of the hierarchy. The rule for young individuals (subadult or D1) entering the hierarchy from the bottom is, ìUp or out!î (Jones, 1980). Both sexes may disperse from natal groups in Alouatta species (1978, 1980; also see Jones, 2004; Glander, 1992), and ìtargetingî is a characteristic method for group expulsion of females, sometimes effected by coalitionary aggression (see Jones, 1978, 1980, 2004; Sterck et al., 1997). Since secondary dispersal is rare or absent in A. palliata (Glander, 1992), ejection from oneís group must generally imply death for the expelled female.
For the present analysis, three patterns are displayed. The most frequent pattern (n= 11) entails both members of the female coalition dominant to the targeted female, van Schaik et al.ís (2004) type 4. In two cases (RYS and SS: Tables 1, 2; also see Jones, 1980), targeted females subsequently emigrated from Group 5. It is also important to note, however, that this most frequent pattern of coalitionary aggression suggests that lower-ranking (e.g., SS) females may help young females (e.g., GWS) to increase their dominance rank (ìrank-changingî coalitions), a variation of van Schaik et al.ís (2004) type 2. The most common pattern of results for female A. palliata might also represent ìpolicingî (suppression of selfish behavior: Frank, 2003) of a group member by the coalition partners, an interpretation that is consistent with van Schaik et al.ís (2004) type 4.
In one instance (Table 2), van Schaik et al.ís (2004) type 5 coalition was observed whereby all coalition partners ranked below their target (GWS). The targeted female, ranked #3 in the female dominance hierarchy of her group (Jones, 1980), ultimately emigrated. Although this pattern of coalitionary aggression was rarely observed, it appears to be a potentially effective agonistic tactic. For the case described here, the emigration of GWS would have resulted in a rank-change for GS and UM1, since these females were ranked #4 and #5 in the female hierarchy below GWS. In the Model, however, type 5 is classified as ìnon-rank-changing.î
The final four coalitions occurred between an adult female and an infant or juvenile. The female target was always subordinate to the adult female coalition partner. It is not possible to be confident that these events involving immatures are, in fact, cases of coalitionary aggression despite the observation that one target, BC, emigrated.
Confirmation of the functions of this possible coalitionary pattern awaits further investigation. These events, for example, may represent immatures ìcopyingî the behavior of adults, possibly their mothers. The latter possibility raises the important observation that coalition partners may be related (see Jones, 2005a). In conclusion, the Model, particularly type 4, provides a very good fit to most cases of coalitionary aggression observed for female mantled howler monkeys. Indeed, the Model, originally devised as a schema for within-group male-male coalitions, provides a better fit for coalitions among adult female than among adult male mantled howlers (see Jones, 2005a).
Future studies should provide theoretical as well as empirical tests of the Model as well as investigate the role of coalitions for within-group policing. Policing with coalitionary aggression has the potential to influence reproductive skew (the apportionment of reproduction within groups) by suppressing the selfish reproductive efforts of targets, a topic in need of study. Finally, as pointed out in Jones (2005a), it will be important to document post-coalition-responses, a category of post-punishment-responses, since targeted individuals, their kin, or onlookers may retaliate against one or more of the original coalition partners, effects that may increase the spiteful consequences of coalitionary aggression (see Jones, 2002).
References
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227-267.
Frank, S. A. (2003). Perspective: Repression of competition and the evolution of cooperation. Evolution, 57, 693-705.
Frankie, G. W., Mata, A., & Vinson, S. B. (2004). Biodiversity conservation in Costa Rica. Berkeley: University of California Press.
Glander, K. E. (1992). Dispersal patterns in Costa Rican mantled howling monkeys. International Journal of Primatology, 13, 415-436.
Jones, C. B. (1978). Aspects of reproduction in the mantled howler monkey (Alouatta palliata Gray). Unpublished PhD Dissertation, Cornell University, Ithaca, NY.
Jones, C. B. (1980). The functions of status in the mantled howler monkey, Alouatta palliata Gray: Intraspecific competition for group membership in a folivorous Neotropical primate. Primates, 21, 389-405.
Jones, C. B. (1985). Reproductive patterns in mantled howler monkeys: Estrus, mate choice, and copulation. Primates, 26, 130-142.
Jones, C. B. (2000). Alouatta palliata politics: Empirical and theoretical aspects of power. Primate Report, 56, 3-21.
Jones, C. B. (2002). Negative reinforcement in primate societies related to aggressive restraint. Folia Primatologica, 73, 140-143.
Jones, C. B. (2004). The number of adult females in groups of polygynous howling monkeys (Alouatta spp.): Theoretical inferences. Primate Report, 68, 7-25.
Jones, C. B. (2005a, July 3). A preliminary test of the Van Schaik model of male coalitions for Costa Rican mantled howler monkeys (Alouatta palliata). Laboratory Primate Newsletter, 44[3], <www.brown.edu/primate/current.html>.
Jones, C. B. (2005b). An exploratory analysis of developmental plasticity in Costa Rican mantled howler monkeys (Alouatta palliata palliata). In A. Estrada, P. A. Garber, M. Pavelka, & L. Luecke (Eds.), New perspectives in the study of Mesoamerican primates: Distribution, ecology, behavior, and conservation (pp. 265-285). New York: Springer.
Jones, C. B. (2005c, April). Condition-dependent reproductive skew in primates: Preliminary thoughts. Theoretical Primatology Project Newsletter, 3(2), Supplement, <www.robertwilliams.org/tpp/tpp_v3-2supp.html>.
Sterck, E. H. M., Watts, D. P., & van Schaik, C. P. (1997). The evolution of female social relationships in nonhuman primates. Behavioral Ecology and Sociobiology, 41, 291-309.
Van Schaik, C. P., Pandit, S. A., & Vogel, E. R. (2004). A model for within-group coalitionary aggression among males. Behavioral Ecology and Sociobiology, 57, 101-109.
------------------------------ Authorís address: Dept of Psychology, Fayetteville State Univ., 1200 Murchison Rd, Fayetteville, NC 28301 [e-mail: [email protected]].
------------------------------
* * *
Meeting Announcements
The AWEN Group, Inc., presents its 3rd Annual Assessment and Treatment of Pain (ATOP) and Distress in Animals Conference Thursday, February 9, 2006, at the Doubletree Guest Suites, Fort Lauderdale, Florida. The assessment and treatment of pain and distress are broad objectives for animal care and use programs, but the devil is often in the details. ATOP III will concentrate on some of the particulars in managing pain and distress in animals.Ý It is appropriate for IACUC members, investigators, veterinarians, facility managers and veterinary technicians involved in the review, preparation and/or application of treatments to animals. For more information, and to register, go to <www.theawengroup.com>. Registration deadline is January 17, 2006.
The Centers for Disease Control and Prevention, the American Society for Microbiology, the Council of State and Territorial Epidemiologists, the Association of Public Health Laboratories, and the World Health Organization will cosponsor the Fifth International Conference on Emerging Infectious Diseases, March 19-22, 2006, in Atlanta, Georgia. The conference will explore current research, surveillance, and prevention and control programs addressing all aspects of emerging infectious diseases. The conference will include general and plenary sessions, symposia, panels of speakers, oral and poster presentations, and exhibits. Abstracts should address new, reemerging, or drug-resistant infectious diseases that affect human health. The deadline for late-breaking abstracts is February 1, 2006. Information about abstract submission is available at <www.iceid.org/abstractsubmission.htm>. Registration information is at <www.iceid.org> and by e-mail at < [email protected]> or < [email protected]>.
All interested persons are invited to attend the Australasian Primate Societyís 24th Annual Conference, March 31st to April 2nd, 2006, at the Perth Zoo, 20 Labouchere Rd., South Perth, Western Australia 6151. More information and registration forms are available at <www.primates.on.net>, or from APS, P.O. Box 631, Mitcham Shopping Centre, Mitcham, South Australia, 5062 [e-mail: [email protected]].
The European Association of Zoo and Wildlife Veterinarians will meet at the Budapest Zoo, May 24-28, 2006. See <www.zoobudapest.com/vet/eazwv2006>.
The Delaware Valley, New Jersey, and Metro New York AALAS Branches invite you to the 2006 Tribranch Symposium, to be held in the Meadowlands Exposition Center in Secaucus, New Jersey, on June 12-14, 2006.Ý ìAchieving the Highest Standards of Animal Careî is the Symposium theme and your abstracts for poster, technical, or scientific sessions are welcome. For more information, see <www.tribranch.org>.
The 4th International Veterinary Vaccines and Diagnostics Conference will be held on June 25-29, 2006, in Oslo, Norway. Many internationally known speakers are invited to address a number of ìhotî topics. Contributions from conference delegates for poster display are most welcome. Abstracts must be submitted for both invited oral and poster presentations and will be published in the abstract book. The abstract deadline is 15 February 2006. For more information, see <www.ivvdc.org>.
* * *
Environmental Enrichment Objects for the Improvement of Locomotion of Caged Rhesus Macaques (Macaca mulatta)
Roland Plesker(1), Judith Heller-Schmidt(1), and Hansjoachim Hackbarth(2)
(1)Paul-Ehrlich-Institut (2)Institute for Animal Welfare and Behaviour, University of Hanover
Introduction and Literature
Environmental enrichment in general is a major issue of debate in laboratory animal science. It is part of the refinement of housing conditions recommended by the ì3Rî-concept of Russell and Burch (1959) for the improvement of animal experimentation, contributing significantly to the well-being of laboratory animals.
This issue is of extraordinary importance in primates, because of their high cognitive abilities in comparison to other laboratory animals (Bayne, 1991; for specific examples see Bloom & Cook, 1989; Buchanan-Smith, 1995; Lutz & Novak, 1995; Line & Hougton, 1987; Line et al., 1989; Markowitz & Line, 1989; Meunier et al., 1989; Murchison, 1991; Preilowski et al. 1988; Reinhardt, 1990, 1992, 1993, 2003; Reinhardt et. al., 1989; and Reinhardt & Reinhardt, 2003). To avoid behavioral abnormalities in these species, it is essential to supply cognitive challenges in the form of different environmental enrichment procedures regularly (Line, 1987).
The aim of this study was to examine whether the locomotion of caged rhesus macaques at the Paul-Ehrlich-Institut is reduced compared to rhesus macaques in their natural environment. Second, we investigated whether locomotion of caged rhesus macaques can be improved by using two locomotive objects: a treadmill and a rotary barrel.
Materials and methods
Animals: The study was conducted with two groups of Chinese rhesus macaques, described in Table 1, at the Paul-Ehrlich-Institut (PEI) in Langen, Germany. Some were imported; the others born here.
Housing: The groups of monkeys were housed in an indoor primate facility in two closed, tiled, and air conditioned rooms. The cages were stainless steel, 3 x 1.25 x 2.25 meters (8.44 cu-m). Temperature was controlled at 20∞ C, and humidity was between 40 and 60 percent. A twelve-hour day was automatically started at 6:00 a.m. with artificial neon light. Natural daylight came through one window. Feeding consisted of monkey pellets (ìssniff Primaten, vegetarischî, 10 mm; ‚ssniff Spezialdi‰ten GmbH, Soest, Germany) ad libitum between 8:00 and 9:00 a.m., and a variety of seasonal vegetables and fruits between 12.30 and 13.30 p.m.
In each cage, the following objects were available: three metal benches, two metal rods, a hanging PrimaHedron swing, and three branches. In addition, plastic bottles, plastic canisters, rubber balls, and cartons were supplied at irregular intervals.
| No. | Sex | Age | Age Class | Origin | |
| A | 53 | M 13 | Adult | China | |
| 58 | F | 14 | Adult | China | |
| 59 | F | 13 | Adult | China | |
| 272 | F | 2 | Juvenile | PEI | |
| 287 | F | 1 | Juvenile | PEI | |
| 303 | M | Ω | Juvenile | PEI | |
| B | 54 | F | 15 | Adult | China |
| 238 | M | 6 | Adult | PEI | |
| 265 | F | 4 | Adult | PEI | |
| 284 | M | 2 | Juvenile | PEI |
Table 1: Description of the two study groups
Enrichment objects: A metal treadmill 78 cm in diameter (Figure 1) and a rotating wooden barrel (vol. = 30 liters, Figure 2) were used. Each group received the treadmill first and the barrel later.

Figure 1: A metal treadmill.
Collection of data: First, the animals were trained for half a year, until they accepted the presence of an observer in the animal room without any change in behavior. During this time, any kind of behavior was recorded.
After this, a time budget was recorded for the time between 6:00 a.m. and 6:00 p.m., in order to compare these data with those of rhesus monkeys in their natural environment. For this purpose, one minute was used to detect the actual behavior of each individual within a group (i.e., 10/12 seconds for each individual). The observed behavior was placed in one of the six following categories: feeding, locomotion, resting, social behavior, agonistic behavior, and other behaviors.

Figure 2: A rotating wooden barrel.
For calculating the effectiveness of the treadmill and barrel, data were collected for two hours per day (10:00 a.m.-12:00 a.m.) as described in the above section. There were two control weeks before the treadmill was offered Tuesday through Friday for two weeks; then two additional control weeks. This was followed by four weeks of ìdiscontinuous object-offerî (only Tuesdays and Fridays), and another two control weeks (Figure 3). This schedule was then repeated for the barrel.
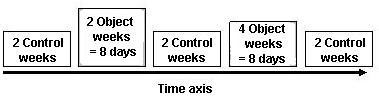
Figure 3: Study design for each of the two objects
Data calculation: First, median values were calculated for all animals and all categories. With these values, a time budget was produced. For statistical calculation, the Wilcoxon Rank Sum Test, Wilcoxon Signed Rank Test, Friedman Test, and Bonferroni-Holm adjusted Wilcoxon-Signed Rank Test were used where appropriate.
Results
Chopra, Seth & Seth (1992) published time budgets of rhesus macaques living at different places in India. In comparison to these behavioral data, especially rhesus macaques living in areas categorized by the authors as ìtempleî and ìstreetî, our two groups of caged rhesus macaques at the PEI spent an equal amount of time in locomotion (Figure 4).
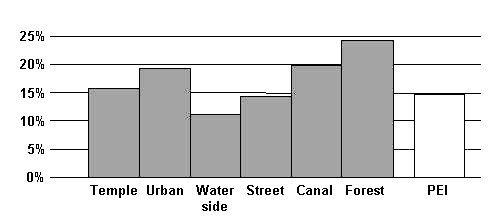
Figure 4: Amount of locomotion (in % of time) observed in rhesus macaques in India and at the Paul-Ehrlich-Institut.
Both the treadmill and rotating barrel were used for locomotion by both groups of caged rhesus macaques (Table 2). Juveniles within both groups used the objects significantly more often than the adults (Wilcoxon Rank Sum Test, one-sided hypothesis, a=0.025, p<0.0001; Table 2).
Group A adults juveniles barrel 2.9% 0.8% 1.4% 16.5% 19.7% 16.3% treadmill 0.2% 0.2% 0.1% 12.2% 13.0% 14.0% Animal number 59 53 58 287 303 272 Group B adults juveniles barrel 0.5% 1.9% 0.1% 11.0% 23.2% treadmill 0.5% 0.5% 2.1% 17.9% 29.6% Animal number 238 54 265 284 307 Table 2: Mean percentage of time which the monkeys spent with the objects. See Table 1 for animal descriptions.
The total amount of locomotion in both groups was significantly increased while the objects were present, compared to the control weeks (Friedman-Test, p<0.0001; Figure 5; locomotion with and without objects).
There was no significant difference in the use of either the barrel or the treadmill between the groups (Bonferroni-Holm adjusted Wilcoxon-Signed Rank Test, one-sided hypothesis, a=0.025, p= 0.047; Figure 6). However, individual preferences could be demonstrated (data not shown).
No significant differences were detected between a ìcontinuousî supply of the objects (4 consecutive times per week for two weeks) and a discontinuous supply (2 nonconsecutive days a week for four weeks) (Wilcoxon Signed Rank Test, one-sided hypothesis, a=0.025, p=0.0423; Figure 6).
In addition, no loss of interest in either object within the period of investigation could be demonstrated for the whole group (Figure 6); however, individual differences were seen (data not shown).
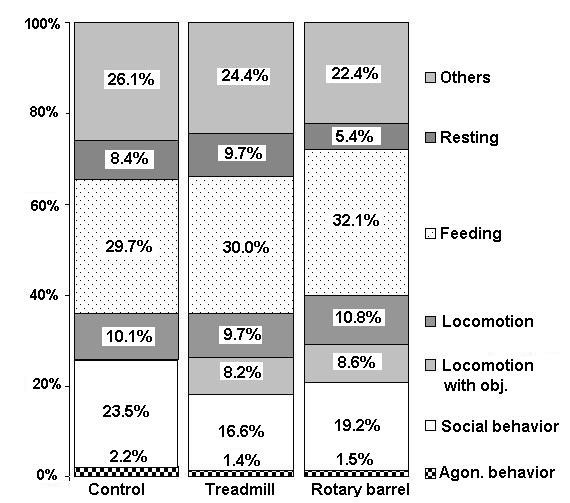
Figure 5: Mean distribution of the behavioral categories (in %) during the control weeks and weeks with locomotion object.
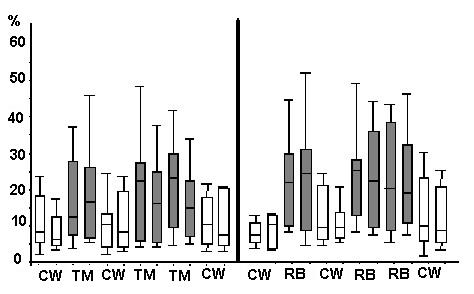
Figure 6: Plot (in %) of total locomotion (locomotion + locomotion with object) per hour. Each bar represents the data of one week (both groups together), and the horizontal lines are the median values. CW = 2 control weeks; TM = 2 weeks with treadmill; RB = 2 weeks with rotary barrel
Due to the increase in locomotion, the amount of time spent in aggressive behavior significantly decreased (Wilcoxon Signed Rank Test, one-sided hypothesis, a=0.025, p<0.0001; result: minus 0.7% / minus 0.8%, respectively). In addition, the time spent in social behavior also significantly decreased. (Wilcoxon Signed Rank Test, one-sided hypothesis, a=0.025, p<0.0001: result: minus 6.9% with the mill and minus 4.3% with the barrel) (see Figure 5).
Discussion
The hypothesis of the present study was that, due to the restricted amount of space, the caged monkeys necessarily move less than those in a natural habitat with its foraging necessity.
However, the time budget in the laboratory, compared to that described by Chopra et al. (1992), showed that, even in nature, especially in areas where the monkeys have not been encouraged to forage much for food, the amount of locomotion was comparable to what we found in our caged groups. Of course, one needs to be very careful in comparing literature data with oneís own results, since many factors can contribute to differences. Nevertheless, the relative equality in locomotion was an unexpected result of our first data collection.
On the other hand, it was not surprising that our groups of rhesus macaques used both locomotion objects within their cages. However, the difference in object use between the adults and the juveniles was unexpectedly high: while the adults almost did not use the objects at all, the youngsters spent up to nearly 30% of their time with one of them (the treadmill). The greater amount of curiosity in younger individuals might have contributed to this result. We have seen similar results when we offered other objects, such as puzzle feeders, to vervets.
Although a barrel may be used differently from a metal treadmill (e.g., chewing on the wooden barrel components), we were unable to detect significant differences in the use of either the rotating wooden barrel or the metal treadmill at the group level. However, as mentioned in ìResultsî, individual preferences for one or the other were seen during the investigation.
Although we were unable to detect any significant differences between a continuous or discontinuous offer of a locomotion object, we would recommend a discontinuous offer, since this allows the provision of another object on the other days, and therefore the opportunity for more variation in housing conditions.
At the group level, we were unable to demonstrate any loss of interest within the period of investigation. However, this period might be too short to answer a question of steady interest sufficiently, since we are housing monkeys with a life span of more than 30 years.
By offering the objects we were able to increase the amount of locomotion significantly compared to the control weeks. This effect simultaneously decreased the amount of agonistic behavior. This is, of course, a desired effect for a colony of caged rhesus macaques. Less positive is the simultaneous reduction in social interactions within the groups when the objects are present. The question is, whether these reductions stay stable when the objects are offered for a longer period of time.
References
Bayne, K. (1991). Providing environmental enrichment to captive primates. Compendium of Continuing Education for the Practicing Veterinarian, 13 [11], 1689-1692 and 1694-1695.
Bloom, K. R., & Cook, M. (1989). Environmental enrichment: Behavioural responses of rhesus to puzzle feeders. Laboratory Animal, 18 [5], 25-31.
Buchanan-Smith, H. (1995). The effect of food distribution on captive Old World primates. Shape of Enrichment, 4 [1], 12-13.
Line, S. (1987). Environmental enrichment for laboratory primates. Journal of the American Veterinary Medical Association, 190, 854-859.
Line, S. W., & Houghton, P. (1987). Influence of an environmental enrichment device on general behaviour and appetite in rhesus macaques. Laboratory Animal Science, 37, 508 (Abstract).
Line, S. W., Markowitz, H., Morgan, K. N., & Strong, S. (1989). Evaluation of attempts to enrich the environment of single-caged nonhuman primates. In J. W. Driscoll (Ed.), Animal care and use in behavioural research: Regulations, issues and applications (pp. 103-117). Beltsville, MD: National Agricultural Library.
Lutz, C. K., & Novak, M. A. (1995). Use of foraging racks and shavings as enrichment tools for groups of rhesus monkeys (Macaca mulatta). Zoo Biology, 14, 463-474.
Markowitz, H., & Line, S. (1989): Primate research models and environmental enrichment. In E. F. Segal (Ed.), Housing, care and psychological well-being of captive and laboratory primates (pp. 202-212). Park Ridge, NJ: Noyes Publications.
Meunier, L. D., Duktig, J. T., & Landi, M. S. (1989). Modification of stereotypic behavior in rhesus monkeys using videotapes, puzzle-feeders and foraging boxes. Laboratory Animal Science, 39, 479 (Abstract).
Murchison, M. A. (1992). Task-oriented feeding device for singly caged primates. Laboratory Primate Newsletter, 31[1], 9-11.
Preilowski, B., Reger, M., & Engele, H. (1988). Combining scientific experimentation with conventional housing: A pilot study with rhesus monkeys. American Journal of Primatology, 14, 223-234.
Reinhardt, V. (1990). Comparing the effectiveness of PVC perches versus wooden perches as environmental enrichment objects for singly caged rhesus monkeys. Laboratory Primate Newsletter, 29[1], 13-14.
Reinhardt, V. (1992). Space utilization by captive rhesus macaques. Animal Technology, 43, 11-17.
Reinhardt, V. (1993). Foraging enrichment for caged macaques: A review. Laboratory Primate Newsletter, 32[4], 1-4.
Reinhardt, V. (2003). Legal loophole for subminimal floor area for caged macaques. Journal of Applied Animal Welfare Science, 6, 53-56.
Reinhardt, V., Houser, W. D., & Eisele, S. (1989). Pairing previously singly caged rhesus monkeys does not interfere with common research protocols. Laboratory Animal Science, 39, 73-74.
Reinhardt, V. & Reinhardt, A. (2001). Environmental enrichment for caged rhesus macaques: A photographic documentation and literature review. Washington, DC: Animal Welfare Institute.
Russell, W. M. S., & Burch, R. L. (1959). The principles of humane experimental technique. Potters Bar: Universities Federation for Animal Welfare.
------------------------------ First authorís address: Paul-Ehrlich-Institut, Paul-Ehrlich-Str. 51-59, 63225 Langen, Germany [e-mail: [email protected]].
------------------------------
* * *
ASP 2006 Conservation Small Grant Applications
The American Society of Primatologistsí Conservation Committee is soliciting grant proposals for ASP Conservation Small Grants for 2006. These grants (up to $1,500) are designed to help fund conservation research or related projects, including conservation education. ASP members working in habitat countries are especially urged to apply or to help someone from a habitat country submit a meaningful project which can be a portion of a larger effort. Grant application guidelines may be obtained by contacting the ASP Conservation Committee Chair at the address below or at the ASP Website.
The ASP Conservation Committee will be making the Conservation Small Grant awards early again this year. This decision was made to better facilitate our getting conservation grant money to the winners in time for the ìsummerî months ñ when many of these projects get underway. The 2006 deadline for submission of grant proposals is January 16, 2006. Materials may be submitted online at the ASP Website, <www.asp.org>, beginning in early December, or sent as an e-mail attachment to the Committee Chair. Grants will be announced in late March. Please direct any questions to: Janette Wallis, Chair, ASP Conservation Committee, ABTI-American Univ. of Nigeria, Lamido Zubairu Way, Yola Township By-pass, PMB 2250, Yola, Adamawa, Nigeria [e-mail: [email protected] or [email protected]].
* * *
Pair Formation and Reintroduction of Temporarily Separated Partners
A Discussion on the Laboratory Animal Refinement & Enrichment Forum
The following discussion (which has been edited) took place from October 25 to 28, 2005, between Erik Moreau [[email protected]], Dawn Abney [[email protected]], Autumn Sorrells [[email protected]], and Viktor Reinhardt [[email protected]]. To subscribe to the Forum, send an e-mail to <[email protected]t>.
Moreau: ìHow have you been re-introducing a previously established pair after a brief period of separation?î
Reinhardt: ìMy experience with rhesus has taught me never to directly reintroduce partners after one of them has been removed from the home cage for more than 24 hours. When you simply put them together, there is a risk that the two companions do not recognize each other quickly enough at the moment of introduction but treat each other as strangers. The consequences of this misunderstanding can be VERY severe.
ìYou can avoid this risk by inserting a transparent or grated mesh divider, and then introduce the one who had been away into the empty half of the cage. Let them find out who they are, and then remove the divider. This trick always worked with rhesus, and there is no reason why it shouldnít work also for your cynos. I know there is an extra time investment but I think it pays off in the safety of the animals.î
Moreau: ìI had never had a problem rotating one of my adult rhesus between two other males. This guy was quite submissive and he showed it very clearly. This may be the reason why the other two males never started to assert themselves by means of overt aggression: the submissive one gave them no reason.î
Abney: ìI have had great success with pairing cynos in the past. For some obscure reason, adult cyno males have been easier to pair than females. You might have a little bit of trouble re-pairing them if they were separated for a month or longer with no visual contact, but I have put pairs back together, after being separated for two weeks, with no problem. We use a clear panel placed between the two cages so the pair can maintain visual contact while separated, but even when thatís not possible the pair typically goes back together just fine. The only animals I have had problems re-pairing after more than one month-long separations were adult male rhesus.
ìIf your cyno males are separated for one or two days only, I really donít think you have anything to worry about when it comes to re-pairing them without preliminaries. Cynos donít always group well, but it is my experience that they make pretty good pairs distinguished by a long-lasting, firm compatibility!î
Reinhardt: ìYour message is very encouraging because you make it quite clear that adult cyno males CAN be paired without undue risks. Many people were made to believe that adult cyno males are better not paired because they were assumed to be intolerant of each other. Obviously this does not need to be true, depending on how you pair the animals and how you keep them after the pair has been established.
ìI should perhaps point out that there are published articles that are in line with your observations:
Sorrells: ìPlease do let us know what technique you use to match up male partners. We have great difficulty pairing 3- to 7-year-old male cynos. Females never give us a problem.î
Abney: ìI am a little surprised that your 3- and 4- year-old males donít go together well. With animals that young, I usually just put them directly together. They are typically compatible without any ado. I never had problems with young males.î
Reinhardt: ìI also introduced young rhesus macaques directly without encountering problems. When they are over three years old, rhesus ó especially males ó often show strong dominance status ambitions, so I first allowed them to establish their rank relationships during a non-contact familiarization period before introducing them as a pair. This eliminated the risk of injurious fighting over dominance at the moment of introduction.î
Abney: ìWith our cyno males older than 5 years, I definitively take no chances but always start off with a transparent panel between the two intended partners. From this I can usually gauge how the socialization will go: Attacking the panel = bad; Lipsmacking or showing curiosity = good. Most of the time I know within the first 30 minutes whether or not things will work out when I eventually give them full access to each other by removing the panel. Needless to say, I separate partners the moment they show signs of imminent fighting; otherwise I leave them together. Compatible partners are not separated during the night.î
Sorrells: ìYes, this is the same technique we use. Yet, many of the animals tested showed lipsmacking for about 20 minutes and then started sham-attacking the partner behind the panel over and over again for up to two hours. We do not take the risk of removing the dividing panel in these cases, but even other partners who give the impression of being more at ease with each other often end up fighting and injuring each other when we remove the panel. This isnít to say we have never paired male cynos, but we havenít had great luck. We keep trying though, much to the PIsí dismay!î
Reinhardt: ìAre you pairing the males in a male-only environment with no females being around?î
Sorrells: ìYes, we are.î
Reinhardt: ìDid you try introducing partners whom you deem compatible in a different cage? When I started pairing adult rhesus, I familiarized potential partners with the help of a grated cage divider, which I simply removed once the pair had established a clear dominance-subordinance relationship. This usually worked, but on one occasion both partners engaged in serious territorial aggression, and this was the end of the pairing attempt. From then on, I always took the trouble to introduce the two partners in a different double cage, and if needed return them to their original location a few days later.î
Sorrells: ìYes, we do this too, but I think it does not make a big difference. Our males just donít work out very well in a pair situation.î
Reinhardt: ìTo sum up, it seems advisable to monitor the reintroduction of temporarily separated partners very carefully, and if possible, to allow the animals to first recognize each other before they are reunited as a pair. There is no good reason to believe that male cynos are less suitable for isosexual pair-housing than rhesus. If and how experimentally-induced pathophysiological processes affect the compatibility of pairs needs to be explored.î
* * *
Announcements from Publications: Australasian Primatology
Submissions (scientific reports, articles, photos, news) are requested for the next edition of Australasian Primatology, a biannual journal produced by the Australasian Primate Society. Scientific reports will be peer-reviewed upon request. Members and nonmembers are welcome to submit electronic copies to <[email protected]>. Please visit the Website <www.primates.on.net> or our discussion group <groups.yahoo.com/group/ausprimatesociety>.
* * *
Travel-Associated Dengue Infections ñ United States, 2001-2004
Dengue is a mosquito-transmitted, acute viral disease caused by any of the four dengue virus serotypes (DEN-1, DEN-2, DEN-3, and DEN-4). Dengue is endemic in most tropical and subtropical areas of the world and has occurred in U.S. residents returning from travel to such areas. The risk for dengue infection among travelers can be reduced by use of repellents and by avoiding exposure to mosquitoes.
From 2001 to 2004 serum samples from 366 patients who had suspected dengue on the basis of clinical presentation and onset of symptoms were submitted to CDC from 37 states and the District of Columbia. Of the 366 patients, 77 were laboratory-diagnosed as acute dengue infections. Of these 77 patients, 67 had dengue infection diagnosed by elevated anti-dengue IgM antibodies, and 10 patients had infection diagnosed after isolation of dengue virus from their serum. Of the 77 acute infections, eight were diagnosed as primary infections, and 12 were secondary infections. For the remaining 57 cases, whether the infection was the patientís first dengue infection or a subsequent infection could not be determined either because a convalescent sample (collected more than 5 days after symptom onset) was not submitted, or both samples were collected during the convalescent phase of infection. Among the 366 suspect cases, dengue testing was negative in 183 patients. A total of 22 patients had elevated IgG titers, suggesting that a flavivirus infection or vaccination (e.g., yellow fever) had occurred in the past but that infection was not the cause of the acute symptoms. For 88 patients, the result of dengue testing was indeterminate because a convalescent sample for serologic testing was unavailable.
The most commonly reported symptoms were fever, headache, myalgias, chills, and rash. Fourteen patients had at least one hemorrhagic symptom, and nine had elevated liver transaminases. Because of incomplete reporting, whether any of the laboratory-diagnosed cases met the clinical criteria for dengue hemorrhagic fever (DHF) could not be determined; however, 15 patients required hospitalization, including one who died. The fatal case occurred in an adult in otherwise good health who had recently returned from a month-long visit to Mexico. Travel destinations were available for 66 patients; 20 patients reported recent travel to a Caribbean island during the two weeks before illness onset, 14 to Pacific islands, 11 to Asia, 10 to Central America, 10 to South America, and one to Africa. Ten patients acquired their dengue infections during travel to areas of the United States in which dengue is endemic (Puerto Rico and U.S. Virgin Islands) or to a U.S. location where an outbreak was occurring (Hawaii from May 27, 2001, to January 30, 2002).
Dengue is transmitted to humans by Aedes mosquitoes. Prevention of dengue among traveling U.S. residents is possible by avoiding exposure to mosquitoes. Prevention not only benefits the traveler but also prevents establishment of autochthonous dengue transmission in areas of the United States in which a competent vector is abundant but dengue virus is absent. During an outbreak that began on May 27, 2001, a total of 122 Hawaii residents tested positive for recent dengue infection after establishment of autochthonous dengue transmission. This outbreak resulted in 45 more cases of laboratory-diagnosed dengue infections than were reported among U.S. travelers from the remaining 49 states for the period 2001-2004. Autochthonous transmission of dengue was also documented in Texas in 1999, when 66 patients with laboratory-diagnosed dengue were identified.
The incubation period for dengue has a range of 3-14 days (in the majority of cases, 4-7 days). Dengue virus infection can be asymptomatic or cause illnesses ranging from mild undifferentiated fever to severe disease, including hemorrhagic manifestations and shock. DHF is characterized by fever, minor or major bleeding phenomena, thrombocytopenia (<100,000 platelets/micro-L), and evidence of increased vascular permeability (e.g., hemoconcentration [hematocrit increased by >20% from baseline], pleural or abdominal effusions, or hypoproteinemia). Previous dengue infection increases the risk for DHF in a patient with subsequent dengue infection. Dengue shock syndrome is DHF with signs of circulatory failure, including narrow pulse pressure (<20 mm Hg), hypotension, or shock, and can result in a case-fatality rate of approximately 10%.
Persons traveling to areas in which dengue is endemic should avoid exposure to mosquitoes by using repellents, wearing protective clothing, and remaining in well-screened or air-conditioned areas. No vaccine is available for preventing dengue infection. Health-care providers should consider dengue in the differential diagnoses of illness for all patients who have fever and a history of travel to tropical and subtropical areas within two weeks before the onset of symptoms. Supportive measures should be administered, and only acetaminophen is recommended for management of pain and fever. Acetylsalicylic acid (i.e., aspirin) and other nonsteroidal anti-inflammatory agents are contraindicated because of their anticoagulant properties and, in the case of children, because of their association with Reye Syndrome. Patients with dengue should be monitored for signs of DHF, especially hypotension, because prompt fluid therapy reduces morbidity and mortality.
Acute-phase (0-5 days after onset of symptoms) and convalescent-phase (6-30 days after onset of symptoms) serum samples obtained for viral isolation and serologic diagnosis, respectively, should be sent through state or territorial health departments to CDCís Dengue Branch, Div. of Vector-Borne Infectious Diseases, National Center for Infectious Diseases, 1324 Calle Cañada, San Juan, PR 00920-3860 [787-706-2399; fax: 787-706-2496]. Serum samples should be accompanied by a summary of clinical and epidemiologic information, including date of disease onset, date of sample collection, and a detailed recent travel history. Additional information for health-care providers regarding dengue case reporting and instructions for specimen shipping are available at <www.cdc.gov/ncidod/dvbid/dengue/dengue-hcp.htm>. ñ From Morbidity and Mortality Weekly Report, 2005, 54, 556-558
* * *
Workshop Announcements
PASA 4th Veterinary Healthcare Workshop
The Pan African Sanctuary Alliance (PASA) will continue its commitment to raising the level of primate care in Africa by staging the PASA 2006 Veterinary Healthcare Workshop, March 19-25, at the Tacugama Chimpanzee Sanctuary in Sierra Leone. The workshop, which will be the fourth since PASA expanded its annual workshop schedule in 2003, is designed to help veterinary staffs at PASA member sanctuaries through seminars, case studies, and practical work. In addition, the 2006 workshop will focus on issues such as transmissible and zoonotic diseases and the development of effective and comprehensive animal health screening programs that will help sanctuaries as they progress towards reintroduction.
PASA is an organization of primate sanctuaries that includes 17 facilities in 12 African countries. For more information regarding PASA or the Veterinary Healthcare Workshop, please contact < [email protected]>, and see <www.panafricanprimates.org>.
Laboratory Animal Disease Workshop
The Midwest Division of the Charles Louis Davis, D.V.M. Foundation, in co-sponsorship with the Biologic Resources Laboratory (BRL) of the University of Illinois at Chicago, will present a Workshop and Symposium on Laboratory Animal Diseases, April 19-22, 2006. Speakers for the symposium include: Dr. Keith Mansfield, Associate Professor of Pathology at Harvard Medical School and Associate Director at the New England Regional Primate Center; and Dr. Annette Gendron-Fitzpatrick, Laboratory Director and Pathologist for the Research Animal Resources Center at the University of WisconsinñMadison. They will present seminars on ìDiseases of Nonhuman Primatesî.
The BRLís collection of study materials will be available for review, at 1840 West Taylor St.Ý The 2 x 2 slide collection includes 14,000 kodachromes, many of which have been digitized, on laboratory animal diseases and management.Ý In addition 3,000 glass micropathology slides with histories and sixty-six T60 video tutorials of the Foundationís Independent Study Center at The Biologic Resources Laboratory will be available for individual and/or group study.Ý Microscopes, projectors (2x2 and LCD), and VCRs will be available at the BRL.
Members of the senior staff of the BRL will give a Simulated Practical Examination in the morning on April 21st, and seminars covering Rodent Pathology, Primate Taxonomy, Other Rodents, and Large Animal Parasites in the afternoon.
For the complete text of the announcement and registration information contact Jim Artwohl [312-996-1217; e-mail: [email protected]] or the C.L. Davis Foundation [e-mail: [email protected]].
Captive Breeding and Reintroduction
A workshop on captive breeding and reintroduction of native species will be held at the Zoological Society of London, May 9, 2006. For information, contact Deborah Body, Regentís Park, NW1 4RY London, U.K. [+20 7449 6610; Fax: +20 7586 2870].
Teaching Research Ethics
Indiana Universityís thirteenth annual Teaching Research Ethics Workshop will convene at the Indiana Memorial Union on the campus of Indiana University in Bloomington, Indiana, May 10-13, 2006. Session topics will include an overview of ethical theory, trainee and authorship issues, conflicts of interest, using human subjects in clinical and non-clinical research, and responsible data management. Many sessions will feature techniques for teaching and assessing the responsible conduct of research.
For more information contact Glenda Murray, Program Associate, Poynter Center, Indiana Univ., 618 E. Third St, Bloomington IN 47405-3602 [812-855-0262; fax: 812-855-3315; e-mail: [email protected]]. Information and registration are also available at <lpoynter.indiana.edu>.
Second Zoo Animal Training Workshop
The Second Zoo Animal Training Workshop will be held June 19-22, 2006, at Paignton Zoo Environmental Park, Devon, U.K. For information, contact N. Dorey, Paignton Zoo Environmental Park, Totnes Road, Paignton, Devon, TQ4 7EU, U.K.
* * *
News Briefs
Zoo Atlanta Gorilla Gives Birth to Twins
A gorilla at Zoo Atlanta has given birth to twins ñ only the sixth time thatís happened in a North American zoo since 1966. Zoo officials say Kuchi, a 21-year-old female western lowland gorilla, gave birth to twins on October 31. The twins are the first offspring of Zoo Atlantaís newest silverback, Taz, and the third and fourth offspring for Kuchi.
There are about 350 western lowland gorillas in captivity in North America. Experts say fewer than 75,000 survive in the wild, partly because their habitat is being destroyed by logging and farming, but also because of the illegal bushmeat trade in Africa, where wild animals are slaughtered for human consumption.
Zoo Atlantaís team of primate experts and veterinary staff plan to monitor Kuchi and the twins closely. Of the six sets of gorilla twins born since 1966, only three survived.
Fifteen gorillas have been born and raised at Zoo Atlanta since 1988. With 22 western lowland gorillas, Zoo Atlanta has the second-largest collection in North America. And officials say an additional three gorillas at the zoo are expecting. ñ Associated Press, November 2, 2005
Research Center for Primates Opens in Korea
A research center opened yesterday that will conduct stem cell experiments on primates. Primates are genetically similar to humans and are used prior to clinical tests on humans when developing stem cell therapies for hard-to-cure diseases.
The state-run Korea Research Institute of Bioscience and Biotechnology launched the National Primates Research Center in North Chungcheong Province with an investment of 7.8 billion won.
Chang Kyu-tae, the head of the center, said the institute will save enormous amounts of money which would otherwise be spent by Korean scientists on overseas tests. ìIt costs 100 million won for overseas tests on one primate,î said Chang. ìThe establishment of the center will also prevent the possible outflow of Koreaís advanced stem cell technology.î
The center has a breeding colony of 74 monkeys and plans to increase the number to 1,500 by 2010. ñ Korea Herald, November 8, 2005
Tulane Primate Center Thrives and Grows
The Tulane National Primate Research Center (NPRC) isnít just surviving, itís thriving. The Center received notice that the National Center for Research Resources of the National Institutes of Health awarded two grants of $4 million each for new construction and expansion of the current breeding facility. In addition, the primate center received more than $1.6 million to support research training in experimental medicine and pathology to prepare veterinarians for careers in biomedical research. The grant proposals were submitted before Hurricane Katrina hit Louisiana.
The first facility grant will fund construction of a facility that will house monkeys assigned to a variety of research studies and state-of-the-art nursery facilities for infant monkeys. The new housing and procedure areas will replace existing square footage in the 40-year-old facility that contains both laboratory and animal housing.
The new facility will be located on the Covington Campus of the Tulane NPRC, and will functionally separate animal areas from laboratory areas, addressing security, access, and biosafety issues that currently exist with the commingling of laboratories and animal housing.
The second grant funds a plan to create a national nonhuman primate breeding colony resource that will also be built on the Covington Campus.
Improvements to land currently owned by Tulane University will include utilities, roadways, drainage, security, and fencing. In addition to corral housing for animals, buildings will be constructed for staging and quarantine of animals as they are received from or shipped to other facilities. ñ From an October 19 press release
Gorilla Haven Joins the AZA
On September 20, Jane T. R. Dewar wrote: A new update has been posted at <www.gorilla-haven.org>, announcing our recent accreditation into AZA (American Zoo and Aquarium Association) as a ìRelated Facilityî ñ one of only 16 such places in North America! Congratulations, Jane and Steuart!
Mountain Gorilla Veterinary Project Moves
Morris Animal Foundation has announced a new partnership between their Mountain Gorilla Veterinary Project (MGVP) and the Maryland Zoo in Baltimore. The partnership will move the MGVP to the Maryland Zoo and effectively establishes the MGVP as its own nonprofit organization.
Morris Animal Foundationís MGVP has been providing veterinary care to mountain gorillas since 1986. The program began when Dian Fossey approached Morris Animal Foundation to help save the species from extinction. At that time, only 248 mountain gorillas were known to exist in the Virunga Mountains; however, in the early 1990s, another gorilla population was confirmed in the Bwindi Impenetrable Forest. Today, the population of this highly endangered species includes about 380 gorillas in the Virungas and another 320 in the Bwindi Impenetrable Forest.
Since its inception, the MGVP has grown from one veterinarian in Rwanda to seven in Rwanda, Uganda, and the Democratic Republic of the Congo. Dr. Mike Cranfield, The Maryland Zooís director of animal health, research, and conservation, has served as the MGVPís project director for seven years and will continue to do so.
Last year, Morris Animal Foundation funded the MGVP with more than $417,670, which helped Project veterinarians to evaluate and monitor more than 50 cases of illness or injury among the mountain gorillas. Half of these cases required direct intervention. Through the Employee Health Program, the MGVP team organized health exams for more than 150 employees of various mountain gorilla organizations. This preventive approach is critical to guarding the gorillas against potentially contagious human diseases. Morris Animal Foundation will continue to provide funding to the MGVP as it establishes itself as its own entity. The Maryland Zoo, which has experience in managing international projects, will take over the administrative side, working with other institutions.
The Larry Jacobsen Primate Library!
Joseph W. Kemnitz,, Director of the Wisconsin National Primate Research Center, announced on December 12 that the Primate Center Library will be named, at a ceremony on December 16, in honor of Larry Jacobsen.
* * *
Volunteer Opportunities
Gibbon Conservation Center ñ California
The Gibbon Conservation Center (GCC), formerly the International Center for Gibbon Studies, is a non-profit organization in southern California, founded in 1976 by Alan Mootnick. It is devoted exclusively to the conservation, propagation, and study of gibbons, for the betterment of the species. We disseminate our findings through educational tours and publications, and by traveling the world assisting zoos and rescue centers.
1. Primate Keeper Duties: animal care and maintenance of the facilities and grounds. Depending on the time of the year, work is 10-12 hours/day, beginning in early morning, 7 days per week. May also include Clerical and Center Assistant work (see below).
2. Center Assistant Duties (depending on skills, needs of the Center, and personal preference): maintenance of grounds, behavioral observations, cleaning of food preparation area, library research, fund-raising, and word processing. Center Assistants must keep a minimum distance of two meters from all enclosures that house gibbons.
3. Clerical Assistant Duties (depending on skills, needs of the Center, and personal preference): word processing, fund-raising, library research, and behavioral observations.
GCC provides free lodging to resident volunteers in an older, basic, travel trailer with access to the bathroom, kitchen, and laundry in the house. Volunteers buy their own food and personal items, which cost approximately $100ñ200 per month at the local supermarkets (depending on the tastes of the volunteer).
Training, lasting approximately one week, is essential at GCC because the volunteers have to follow strict procedures that are mandated by law and our own goals of cleanliness, gibbon welfare and breeding, and safety for people and gibbons. The training involves working hands-on with an experienced primate keeper.
Local transportation is by train or bus, unless one of the staff happens to be going into town.
Applications are available on our Website, <www.gibboncenter.org/volunteering.htm>, or from the address, e-mail or phone numbers below. Return the completed application with a cover letter, resume, and two letters of recommendation. Contact: Patricia Dahle [661-943-4915] or Erin Bell [661-296-9495], Volunteer Coordinators [e-mail: [email protected]; fax: 661-296-1237]; or write to Gibbon Conservation Center, Volunteer Coordinator, P. O. Box 800249, Santa Clarita, CA 91380.
For additional information see <www.gibboncenter.org>. Please mention ZooNews Digest should you apply. ñ Posted to ZooNews Digest
Hurricane Wilma Recovery ñ Florida
The DuMond Conservancy and Monkey Jungle, in Miami, Florida, have suffered extensive damage due to the effects of Hurricane Wilma. Conservancy spokesperson Willy Cotto reports that the owl monkey canopy has been decimated and that there has been extensive damage to primate housing. All animals are safe, but there is an urgent need for a generator to provide fresh water and refrigeration. Dr. Sian Evans, Willy Cotto, and their staff have been very creative in caring for the monkeys, but running water is necessary to clean the primatesí enclosures.
Volunteers are needed for all aspects of cleanup, repair, and restoration. The DuMond Conservancy for Primates and Tropical Forests is a 501 (c)(3) non-profit organization providing a home for confiscated and retired laboratory primates. In addition, the Conservancy serves as an educational and scientific resource for students interested in learning more about our closest relatives.
The DuMond Conservancy is located in subtropical South Florida, near Miami. The warm climate allows us to keep our primates out-of-doors year round, either free-ranging in a lush, 4-acre forest, or in large enclosures planted with natural vegetation and located in secluded woods.
The primate facility, Monkey Jungle, adjacent to the Conservancy, also suffered similar damages and is in need of similar help. Contact Dr. Sian Evans or Willy Cotto, DuMond Conservancy, 14805 Southwest 216th St, Miami, FL 33170 [305-323-9079; 305-238-9981; e-mail: [email protected]]. ñ Posted to Alloprimate, October 26
* * *
Resources Wanted and Available
IVIS Books Online
International Veterinary Information Service (IVIS) is publishing Laboratory Animal Medicine and Management, edited by J. D. Reuter and M. A. Suckow. Chapters are added to the Website, <www.ivis.org>, as they are ready. Present contents include a Book preview, by J. D. Reuter & M. A. Suckow; Regulatory compliance, by J. D. Reuter; Environmental enrichment for laboratory animals, by K. L. Stewart & K. Bayne; and an Appendix: Regulations on laboratory animal use, by J. D. Reuter. Chapters in preparation include Biology and medicine of non-human primates, by D. D. Myers; and Occupational health, by C. Hankenson.
New chapters have been added to the e-book, A Concise Review of Veterinary Virology, written by G. Carter, D. Wise, and E. Flores. These chapters, ìOrthomyxoviridaeî and ìBunyaviridae and Bornaviridaeî, are available at <www.ivis.org/new.asp>.
Access to the material is free, but restricted to licensed veterinarians, veterinary students, researchers, and professionals in the animal health industry only. You will be required to register.
Primate Treat Recipe Cookbook Available in pdf
The Catarrhine Cafe Cookbook, compiled by Rita Bellanca and Diella Koberstein, includes ìTricks of the Tradeî, recipes, and examples of enrichment items. A pdf version of this resource is available at <www.wanprc.org/wanprc/cookbook-forExternal.pdf>, part of the Website of the Psychological Well-being Program of the Washington NPRC: <www.wanprc.org/wanprc/psych-wellbeing.htm>.
New Software Release by Noldus
Noldus Information Technology has released The Observer XT, a totally renewed software package for behavioral observation, video recording and multimodal measurements. The new software was developed in close collaboration with researchers. Important innovations include the integration and automatic synchronization of physiological measurements, the support of multiple videos, more flexible and powerful coding options, and an integrated view of behavioral data with physiology and video. New coding options include building and fine tuning your coding scheme while observing, the use of rating scales, start-stop coding, hierarchical coding schemes, multiple modifiers, and more. Setting up a study has become very quick and easy.
The development team focused on ease of use, and on interoperability. The result is a software that can interface with all sorts of equipment, including a wide range of camera types, MPEG encoders, data acquisition devices, and computer programs. The new software is also compatible with Pocket Observer, the event recorder that runs on handheld computers, and with earlier versions of The Observer. For more information, please see <www.noldus.com/observerxt>; or contact Albert Willemsen [e-mail: [email protected]].
Training and Enrichment Talks
Amy Kerwin announces that the Animal Behavior Management Alliance <www.membersolutions.net/ organization/about.asp> has posted its ìTraining and Enrichment Talksî on the Primates Inc. Website.
Go to <www.primatesinc.com> and click on ìresourcesî. There, you can download the following pdfs:
* * *
Information Requested or Available
NIH is Ready for Electronic Submission
The National Institutes of Health will soon require all competing research grant applications to come in electronically via the Web portal of <Grants.gov> on a new SF 424 Research and Related (R&R) application.
ìWe are phasing in the changes by type of grant program (mechanism), beginning with the December 1, 2005, submission date for small business applicants and culminating in May, 2007, when all grant programs will be submitted electronically on the new form. See <era.nih.gov/ElectronicReceipt> for dates and other information. Applicants should carefully note the transition date for the grant mechanism for which they wish to apply. Once a grant mechanism is in the electronic mode and the grant opportunity is posted on <Grants.gov>, applicants will be able to download and begin working on their application package.
ìApplicants must be aware that until the transition, any applications submitted for that grant mechanism should be submitted on paper forms at <grants2.nih.gov/grants/funding/phs398/phs398.html>. NIH systems will not be ready to receive those applications electronically until the transition date. Paper applications will not be accepted after the transition date for a grant mechanism.
ìPrepare to use electronic submission by studying <era.nih.gov/ElectronicReceipt/preparing.htm>, and regularly visit this Website to keep pace with changes in NIHís electronic receipt program. Also see <era.nih.gov/ElectronicReceipt/faq.htm> for more details on electronic submission.î
Biographical Sketches: Evolutionists and Ecologists
Charles H. Smith, Professor and Science Librarian, Western Kentucky University, Bowling Green [e-mail: [email protected]], has announced that he has just finished making operational a new Website service entitled ìSome Biogeographers, Evolutionists and Ecologists: Chrono-Biographical Sketchesî, which provides about 250 profiles of leading naturalists of the past at <www.wku.edu/~smithch/chronob/homelist.htm>.
This new site is meant to stand on its own as a biographical source, but it is also linked from the entries at the already existing ìEarly Classics in Biogeography, Distribution, and Diversity Studiesî sites, at <www.wku.edu/~smithch/biogeog> and <www.wku.edu/~smithch/biogeog/index2.htm>.
ìVirtual Primate Museumî
Dr. Bernhard Wiesemueller of the Universit‰t Gˆttingen has created a ìVirtuelles Primatenmuseum ñ The Virtual Primate Museumî, at <primatenmuseum.de>. People who wish to contact him regarding this project can send an e-mail to < [email protected]>.
USDA Releases Report on Animal Care Program
The USDA has released a report entitled Audit Report, APHIS Animal Care Program Inspection and Enforcement Activities, Report No: 33002-3-SF. This report has been issued by the Agriculture Departmentís Office of the Inspector General and specifically covers APHISí Animal Care Program. The report is available at <www.usda.gov/oig/new.htm>. ñ National Association for Biomedical Research, October 20
FY2004 USDA/APHIS Animal Care Report Online
The Fiscal Year 2004 Annual Report for the Animal Care division of USDA/APHIS is now available online.Ý The report shows that 1,101,958 AWA-covered animals were involved in research in 2004, continuing a long trend of fewer AWA-covered animals in research, down from 2,134,182 as recently as 1992. See <www.aphis.usda.gov/ac/awreports/awreport2004.pdf>.
Change to USDA Procedures on Annual Reports
The National Association for Biomedical Research (NABR) announced that they had learned that, effective Wednesday, October 26, annual reports submitted to USDA by research facilities will be released with signature blocks. A ruling by the Department of Justice states that signatures can no longer be redacted. The Animal Welfare Act requires that the CEO or Institutional Official sign and certify the reports.
ìPlease do not hesitate to contact NABR regarding this issue [202-857-0540; e-mail: [email protected]].
Exchange of Scientific Papers
The International Group of Exchange of Medical and Biological Articles has been created to allow exchange of scientific papers among students, researchers and professionals around the world. The intention is to provide access to medical and biological literature to all who want it. Messages must be in English or Spanish. See <br.groups.yahoo.com/group/IGEMBA>.
Conference Report: Monkeys Old and New
The New York Consortium in Evolutionary Primatology convened its first international conference in New York, October 28-30, 2005. Entitled ìMonkeys Old and Newî, the conference provided a venue for discussion of topics in the evolution, behavior, genetics, paleontology and morphology of extant and extinct monkeys from the New and Old Worlds. A report is available at <www.nycep.org/monkey-symposium>. Abstracts of talks are available at <www.nycep.org/monkey-symposium/assets/TALK_ABSTRACTS_10-27-05.pdf>, and of posters at <www.nycep.org/monkey-symposium/assets/POSTER_ABSTRACTS_10-21-05.pdf>.
More Interesting Websites
* * *
Italian Society for Evolutionary Biology
The Italian Society for Evolutionary Biology (for information, see the Societyís temporary Website:
<web.unife.it/progetti/genetica/CongBiolEvol>)
has been formed to encourage the teaching of evolutionary theory in all Italian schools and universities, and to defend such teaching from the recent attacks of creationist movements. The Society also aims to stimulate research in evolutionary biology by encouraging collaborations among Italian evolutionary biologists in Italy and abroad, and between Italian researchers and their foreign colleagues.
The steering committee would also like to announce that the first Italian Congress of Evolutionary Biology will take place at the end of August, 2006, in Florence. The steering committee is composed of: Giorgio Bertorelle, University of Ferrara, President < [email protected]>; Gisella Caccone, Yale University, Vice-President < [email protected]>; Francesco Scalfari, University of Asti, Secretary-Treasurer < [email protected]>; Renato Fani, University of Firenze, Counselor <[email protected]>; Marco Ferraguti, University of Milano, Counselor < [email protected]>; Alessando Minelli, University of Padova, Counselor < [email protected]>; Mariella Rasotto, University of Padova, Counselor < [email protected]>; Francesco Santini, University of Toronto, Counselor < [email protected]>; and Ivan Scotti, INRA, French Guyana, Counselor < [email protected]>.
Membership in the Society is open to everyone interested in evolutionary biology. For membership information, please contact the Societyís secretary [e-mail: [email protected]].
* * *
Positions Available
Environmental Enrichment Specialist ñ Tulane
Tulane National Primate Research Center is seeking an Environmental Enrichment Specialist to perform a variety of tasks related to the implementation and maintenance of the environmental enrichment program for nonhuman primates and other laboratory species as directed by the Enrichment Program Coordinator. This person assists with the social management of new and existing social groups; identifies, reports, and implements interventions for primates with behavioral problems; assists with the design and evaluation of enrichment devices and other behavioral management strategies; implements positive reinforcement training; facilitates the daily implementation of enrichment procedures; maintains documentation of program implementation; and generates progress reports.
Qualifications include a BS degree with a background in behavioral sciences and 2 yearsí experience in laboratory animal care; or 4 yearsí experience in laboratory animal care plus certification as an AALAS LAT; or 6 years of experience in laboratory animal care; PLUS the ability to follow oral and written instructions and the ability to lift items weighing up to 43 lbs.
Salary will be commensurate with experience for this permanent full-time position. Contact Kate Baker, Tulane NPRC, 18703 Three Rivers Rd, Covington, LA 70433 [985-871-6578; fax: 985-871-6328; e-mail: [email protected]].
Quality Assurance Specialist ñ Tulane
The Tulane National Primate Research Center is also seeking a Quality Assurance Specialist to provide quality assurance for their Animal Care and Use Program by ensuring that all aspects of animal husbandry, animal care, and maintenance of the animal facility comply with all applicable regulatory agencies and internal standard operating procedures. This person will be required to work weekends and after hours and will be required to wear a pager/cell phone for emergencies. S/he will maintain documentation in support of regulatory compliance.
Qualifications include a BS degree in Animal Science or a related field (preferred); experience in developing and maintaining a Quality Assurance Program in industry or academic settings, or 6 years of laboratory animal care experience, or 4 years of laboratory animal care experience and AALAS Laboratory Animal Tech certification; the ability to follow oral and written instructions; computer literacy; and the ability to lift items weighing up to 50 lbs. Salary will be $28,500 ñ 41,800 per year, commensurate with experience.
Please send resume to Tulane NPRC, 18703 Three Rivers Rd, Covington, LA 70433 [fax: 985-893-1352; e-mail: [email protected]].
Primate Veterinarian ñ Alice, Texas
Covance Research Products (CRP), a drug development company with facilities in 18 countries worldwide, is seeking a veterinarian with nonhuman primate experience for their Texas facility, Covance Research Products. Specific duties and responsibilities include: * developing and performing standard operating procedures; * assisting in animal welfare and regulatory compliance, including AAALAC, ISO, OLAW, and federal and state regulations; * supporting overall animal health initiatives; * supporting our Texas site through appropriate client contact, societal meeting attendance, and participation in site visits; * training animal care staff; and * supporting CRP and Covance veterinary initiatives.
Requirements include * a DVM or VMD degree; * licensing in the Commonwealth of Texas; * strong surgical skills and pharmacometrics experience; * demonstrated interest in program development; and * client service orientation.
Apply online at <www.covance.com/careers>, or e-mail resume to <[email protected]>.
Evolutionary Ecologist ñ Binghamton
The Department of Biological Sciences at Binghamton University (one of the four SUNY University Centers) invites applications for a new position at the assistant professor level in Evolutionary Ecology. Ý We seek someone who can establish a strong, extramurally-funded, empirical research program informed by evolutionary theory. Research that integrates evolutionary ecology with other disciplines (e.g., ecosystem ecology, genetics) will receive special attention. This position is designed to contribute to cross-departmental integration through EvoS [see <bingweb.binghamton.edu/~evos>], our campus-wide evolutionary studies program.Ý Applicants should submit a CV, a statement of research and teaching interests, and a sample of reprints to Dr. David Sloan Wilson, Chair, Evolutionary Ecology Search Committee, Dept of Biological Sciences, Binghamton University, SUNY, Binghamton, NY 13902-6000; and have three letters of reference sent to Dr. Wilson. Review of applications will begin on January 1, 2005, and will continue until the position is filled.
Binghamton University is an equal opportunity/affirmative action employer.
* * *
Educational and Research Opportunities
New World Primate Caregiver/Office Assistant
Pacific Primate Sanctuary, Maui, Hawaii, is seeking someone with a background and interest in animal husbandry, welfare, conservation, exotic veterinary medicine, biology, and related fields, who is a mature team player with respect for others. We need a good, clear communicator (written and spoken English) and coordinator with a minimal personal agenda, who seeks mutual growth for all and is able to make a long-term commitment to the primates and the staff. The position requires a self-starter and hard worker, humble, compassionate, self-reliant, organized, professional, computer literate, open to learning, capable of problem solving and of completing a task. Our Resident Intern needs to be physically fit, with no communicable diseases or criminal record, with a driverís license and an enjoyment of a rural lifestyle with simple amenities. A one-year commitment is requested. Citizens of foreign countries must secure their own visas and permits independently.
Responsibilities of an animal caregiver involve: providing daily care, enrichment, and nurturing to about 60 monkeys; administering medications; preparing food; cleaning enclosures; and maintaining the Sanctuary facility. Office assistance would include: administering the volunteer program (scheduling, processing of applications, advertising), data entry (Mac-Filemaker Pro, Excel, Word), correspondence, record-keeping, fundraising, grant writing, and public relations. After a training period, managerial duties would be added and more technical training may be available. The position requires an average of 40 hours per week, including on-call duties.
Pacific Primate Sanctuary is a nonprofit organization (501(C)(3)). Our staff is 15 volunteer caregivers and a support system of veterinary and other professionals. Since the well-being of the monkeys is our primary focus, they are not on exhibit, the facility is not open to the public, and they are not subjected to any medical research. As a result, our organization is not eligible for funding from any governmental agencies nor do we receive funds from admissions. Therefore, we must rely solely upon donations from compassionate individuals and organizations. Our operating budget is extremely small. Financial independence for living expenses would be required. If you are considering the possibility of seeking grant support on your own, supplementary funds for needs other than housing could be built into your budget.
We provide housing and utilities. An organic garden and fruit orchard exist on-site. A new 24-foot yurt is adjacent to the Sanctuary on a large piece of rainforest property with its own bathroom facilities. You would be trained by experienced members of our staff in all aspects of New World primate care (Callithrix and Cebus). Observational research could be conducted on this colony of marmosets, tamarins, and capuchin monkeys in a naturalistic environment.ÝÝ
Weíd greatly appreciate receiving your resume, CV, and letters of reference by e-mail. Please visit our Website, <www.pacificprimate.org>. ñ Lucy L. Wormser, Founder and President, Pacific Primate Sanctuary, Inc., 500 AÝHaloa Rd, Haiku, Maui, HawaiiÝ 96708 [phone and fax: 808-572-8089; e-mail: [email protected]].
Summer Apprentice Program ñ Washington State
The Chimpanzee & Human Communication Institute (CHCI) is taking applications for our 10-week Summer Apprentice Program (June 18 to August 26, 2006). Graduates, undergraduates, and postgraduates from any nation and from various academic backgrounds (e.g. anthropology, biology, psychology, linguistics, philosophy, etc.) are encouraged to apply. This program is open to students from all universities and to post-graduates.
The research at CHCI involves a group of four chimpanzees who use the signs of American Sign Language (ASL). Three of the four were part of the cross-fostering research that began with Drs. R. A. & B. T. Gardner. Each chimpanzee was raised in an enriched environment in which his or her human family members used only ASL, much like the environment in which a deaf human child grows up. The fourth chimpanzee, Loulis, was adopted by Washoe in 1978 and learned his signs from other chimpanzees as a focus of research done by the co-directors of CHCI, Dr. Roger and Deborah Fouts. Currently, the chimpanzees reside at the CHCI on the campus of Central Washington University in Ellensburg, Washington, in a large state-of-the-art facility.
Apprentices are at the institute daily ñ cleaning enclosures, preparing meals and enrichment, making observations of the chimpanzees, and participating in one or more research projects. The first week is intensive training in laboratory jobs and chimpanzee behaviors. The philosophy of CHCI is that the needs of the chimpanzees come first. Apprentices are trained in humane care and research techniques. After several weeks each apprentice becomes more autonomous and has responsibilities in the research project.
The program fee is $1800 and there is a non-refundable $25 application processing fee. This does not include housing and transportation. Inexpensive housing is available on campus. A course in ASL is highly recommended but not required. For more information see <www.cwu.edu/~cwuchci/apprentice.html>; or contact Dr. Mary Lee Jensvold, CHCI, CWU, Ellensburg, WA 98926 [e-mail: [email protected]]. The deadline to apply is March 17, 2006.
* * *
Primates de las Américas... La Página
Estimados lectores, hemos decidido hacer esta entrega adicional. En ella se incluyen un resumen desde Argentina, además de informaciones sobre la naciente Sociedad Argentina de Primatología (SARPRIM), un libro de Colombia y un útil programa estadístico. Saludos cordiales, Tania Urquiza-Haas ([email protected]) y Bernardo Urbani ([email protected]).
Estrategias de optimización del forrajeo en el mono aullador negro y dorado (Alouatta caraya) en el noreste argentino. Verónica Ventura. Sección de Etología, Fac. de Ciencias, Univ. de la República, Montevideo, Uruguay ([email protected]). Tesis de pregrado, 2004. Orientadora: Silvia Corte. Instituto de Biología, Universidad de la República, Uruguay.
Los primates que incluyen frutos en su dieta, para los cuales el alimento está agrupado en parches discretos y dispersos entre sí, como es el caso de los aulladores, se espera que maximicen la ingesta energética consumiendo los frutos al comienzo de la jornada, ya que representan energía de acceso rápido después de muchas horas de inactividad, y es cuando las hojas contienen la menor concentración de fotosintatos (y por consiguiente menor nivel proteico y glucídico). También se espera que minimicen la energía gastada en la búsqueda del alimento siguiendo rutas no al azar en las cuales se minimice la distancia viajada entre los parches de alimento. Se pretendió evaluar si los ejemplares de Alouatta caraya residentes en la Provincia de Corrientes, poseían comportamientos coincidentes con estas estrategias de optimización del forrajeo. Para esto se tomaron registros comportamentales de dos grupos por el método de barrido cada 5 minutos durante 19 y 20 días respectivamente, abarcando toda la jornada de forrajeo (completando en total 393 horas de observación) y se cuadriculó el territorio de cada grupo en cuadrantes de 20 x 20 m, a efectos de mapear las rutas seguidas por los animales durante sus movimientos diarios. La ingestión de la mayor proporción de frutos coincide con la de locomoción (la cual implica la búsque-da activa del alimento) en la segunda mitad del día (U=27.5; a=0.05), lo cual se opone a la primer hipótesis propuesta. Esto puede ser explicado asumiendo que en la mañana ingieren los alimentos mas cercanos disponibles, quizás debido a un déficit energético después del largo ayuno nocturno. Los datos confirman además un patrón espacial de forrajeo no coincidente con el azar, ya que utilizan una pequeña fracción del territorio disponible (19 y 37%) constituida por diferentes sectores visitados con una alta variabilidad. Los altos valores del &IACUTE;ndice de Circularidad, por otra parte, son opuestos al principio de minimización de distancia, ya que llegan a viajar por día 40 veces el desplazamiento neto, moviéndose de forma repetitiva a través de los mismos árboles, lo cual podría estar relacionado con estrategias para minimizar los costos de la detoxificación vegetal. Las posibles causas para que no se cumplan las hipótesis propuestas estarían relacionadas con la estrategia digestiva particular de los aulladores, por lo que los determinantes críticos de los patrones comportamentales y alimenticios podrían ser mayormente internos y no externos. El grupo con más integrantes poseía menor superficie (y alimento) disponible, y viajó una menor distancia diaria, con un patrón de movimiento menos azaroso, altamente circular y repetitivo, mostrando una explotación más intensiva del recurso disponible, quizás resultado de una mayor presión para optimizar el forrajeo. El estudio de cómo los animales lidian con los problemas impuestos por el entorno y de la plasticidad con la que lo hacen resulta fundamental para entender las fuerzas evolutivas que guían los procesos adaptativos.
Sociedad Argentina de Primatología (SARPRIM)
Jóvenes investigadores en primatología de la Argentina están creando la Sociedad Argentina de Primatología (SAPRIM). Sus objetivos son vincular a la comunidad primatológica de aquel país y buscar espacios de encuentro con otros, y principalmente estimular a la nueva generación de estudiantes para que se integren y potencien esta disciplina. Se tendría pensado preparar un noticiero online trimestral. Si tiene interés en suscribirse al SAPRIM puede enviar la siguiente información; 1) Nombre y apellido; 2) Lugar de trabajo; 3) Correo electrónico; 4) Posición académica; 5) &AACUTE;reas de interés; 6) Especies de interés; y 7) Sugerencias. Para enviar la información anterior o solicitar cualquier adicional escriba al <[email protected]>.
Los Micos de Colombia
El siguiente libro primatológico de Colombia ha sido publicado, a saber,
Bennett, S. (2003). Los micos de Colombia. Santafé de Bogotá: Instituto de Investigación de Recursos Biológicos Alexander von Humboldt/Fundación Tropenbos. 260 pp. (ISBN: 958-8151-21-X).
Información adicional puede encontrarse en esta: <www.humboldt.org.co/humboldt/mostrarpagina.php?codpage=600022&ano=2003>.
Programa para Análisis en Primatología
ìPoptoolsî es un excelente pequeño programa estadístico (aproximadamente 2 MB) que es fácil de bajar y se autoinstala en Excel. Una vez allí puede realizar diversos análisis básicos de estadística utilizando de manera extremadamente sencilla la plantilla de Excel. Este programa fue diseñado por los ecólogos Greg Hood y Jean DeBord en Camberra, Australia. Es absolutamente recomendable tenerlo y puede ser bajado gratuitamente en: <www.cse.csiro.au/poptools> o directamente en <www.cse.csiro.au/poptools/download.htm>.
* * *
Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
Books
* Commensalism and Conflict: The Human-Primate Interface. Special Topics in Primatology, Volume 4. J. D. Paterson & J. Wallis (Eds.). J. Wallis (Series Ed.). American Society of Primatologists, 2005. [Price: $60]
. . . Contents; Foreword, by L. D. Wolfe; Preface, by J. D. Paterson; Perceptions of pests: Human attitudes to primates, conflict and consequences for conservation, by P. C. Lee & N. E. C. Priston; By-product mutualism: Conservation implications amongst monkeys, figs, humans, and their domesticants in Honduras, by F. Burton & A. Carroll; People, crops, and primates: A conflict of interests, by C. M. Hill; Human social issues, disease, and sympatric apes in the Central African Republic, by A. A. Lilly; Residents and immigrants: Reactions and perceptions of crop raiding in Masindi District, Uganda, by J. Paterson; Primates in the forest: Sakalava ethnoprimatology and synecological relations with black lemurs at Ambato Massif, Madagascar, by I. C. Colquhoun; Tanala synecological relations with lemurs in southeastern Madagascar, by P. C. Wright, B. R. Andrianamihaja, & S. Raharimiandra; Tourist impacts on the behavior of black howling monkeys (Alouatta pigra) at Lamanai, Belize, by A. Treves & K. Brandon; Monkey forests and human landscapes: Is extensive sympatry sustainable for Homo sapiens and Macaca fascicularis on Bali? by A. Fuentes, M. Southern, & K. G. Suaryana; Characterizing primate pet ownership in Sulawesi: Implications for disease transmission, by L. Jones-Engel, M. Schillaci, G. Engel, U. Paputungan, & J. Froehlich; Farmer attitudes toward the conservation of ìpestî monkeys: The view from Nepal, by M. K. Chalise & R. L. Johnson; Rhesus commensalism in India: Problems and prospects, by C. H. Southwick, I. Malik, & M. F. Siddiqi; City monkeys (Macaca mulatta): A study of human attitudes, by A. Srivastava & F. Begum; Coexistence of bonnet monkeys (Macaca radiata radiata Geoffroy) with planters in the cardamom (Elettaria cardamum Maton) and coffee (Coffea arabica L.) plantations of Karnataka, South India: Hospitable or hostile? by A. K. Chakravarthy & N. E. Thyagaraj; Habitat destruction, population compression and overbrowsing by the Zanzibar red colobus monkey (Procolobus kirkii), by K. S. Siex; Round up the usual suspects ñ Conflict between monkeys and farmers in east and west Africa, by J. Chism; The population status of the ursine colobus (Colobus vellerosus) at Boabeng-Fiema, Ghana, by T. Saj, J. Teichroeb, & P. Sicotte; Tibetan macaques, visitors, and local people at Mt. Emei: Problems and countermeasures, by Q.-K. Zhao; Recent expansion of the range of Japanese macaques, and associated management problems, by K. Watanabe & Y. Muroyama; Applied conditioned taste aversion and the management and conservation of crop-raiding primates, by D. L. Forthman, S. C. Strum, & G. M. Muchemi; and Managing monkeys and mangos, by M. Baker & A. Schutt.
* Handbook of Primate Husbandry and Welfare. S. Wolfensohn & P. Honess. Ames, IA: Blackwell Publishing, 2005. 176 pp. [Price: paperback, $69.99]
. . . Contents: 1. Primates: Their characteristics and relationship with man. 2. The physical environment. 3. Staff management and health and safety. 4. Nutrition. 5. Physical well-being. 6. Psychological well-being. 7. Training of primates. 8. Breeding. 9. Sourcing and transporting primates.
Magazines and Newsletters
* Connection, Fall, 2005. [AAALAC International, 11300 Rockville Pike, Suite 1211, Rockville, MD 20852-3035]
. . . Includes: ìPreparing for the semiannual reviewî.
* Animal Research News & Analysis Newsletter, October 2005. [The Humane Society of the United States, 2100 L St, NW, Washington, DC 20037; <www.hsus.org/ animals_in_research/general_information_on_ animal_research/subscribe_to_the_animal_ research_news_analysis_newsletter>].
* CC Update, Summer 2005, 16[2]. [Community Conservation, Inc., 50542 One Quiet Lane, Gays Mills, WI 54631; <[email protected]>].
* Gorilla Journal: Journal of Berggorilla & Regenwald Direkthilfe, June, 2005, No. 30. English: <www.berggorilla.de/gj30e.pdf>; German: <www.berggorilla.de/gj30d.pdf>; French: <www.berggorilla.de/gj30f.pdf>. December, 2005, No. 31. English: <www.berggorilla.de/gj31e.pdf>; etc.
* NCRR Reporter, Spring 2005, 29[2]
. . . Contents include ìMonkey genes find expressionî and ìBringing veterinarians into biomedical researchî.
* Neotropical Primates: A Journal and Newsletter of the IUCN/SSC Primate Specialist Group, August 2004, 12[2].
. . . Contents: The use of date palms (Phoenix sp.) as resting and sleeping sites by Callithrix jacchus in northeastern Brazil, by M. O. Teles de Menezes; On the occurrence of the owl monkey (Aotus azarai) in Cerro Leon, Chaco, Paraguay, by J. M. Campos, I. Benitez, & D. A. Meritt, Jr.; The use of camera-traps in a survey of the buff-headed Capuchin monkey, Cebus xanthosternos, by M. C. Martins Kierulff, G. Rodrigues dos Santos, G. Canale, C. E. Guidorizzi, & C. Cassano; Preservation and extraction of DNA from feces in howler monkeys (Alouatta caraya), by L. I. Oklander, M. Marino, G. E. Zunino, & D. Corach; InfecÁáo por endoparasitas em um grupo de bugios-pretos (Alouatta caraya) em um fragmento florestal no estado do Mato Grosso do Sul, Brasil, by K. C. I. Godoy, A. Odalia-Rimoli, & J. Rimoli; A muriqui (Brachyteles hypoxanthus) with a broken leg at the EstaÁao Biológica de Caratinga, Minas Gerais, Brazil, by F. P. Paim, M. F. Iurck, S. L. Mendes, & K. B. Strier; A survey of black howler (Alouatta pigra) and spider (Ateles geoffroyi) monkeys along the Rio Lacantún, Chiapas, Mexico, by A. Estrada, S. Van Belle, & Y. García del Valle; Primate species at the Tiputini Biodiversity Station, Ecuador, by L. D. Marsh; Survey of a gallery forest primate community in the cerrado of the Distrito Federal, central Brazil, by R. P. Barros Henriques & R. J. Cavalcante; Resultados da enquete sobre ocorrÍncua de orunatas no Rio Grande do Sul, Brasil, by T. L. Codenotti & V. Martins da Silva; and In memoriam: Pekka Soini: 1941-2004, by R. A. Mittermeier, E. W. Heymann, J. Salo, & M. Pyh‰l‰.
* Primate Report, July 2005, 71.
. . . Contents: Morphology, behaviour and molecular evolution of giant mouse lemurs (Mirza spp.) Gray, 1870, with description of a new species, by P. M. Kappeler, R. M. Rasoloarison, L. Razafimanantsoa, L. Walter, & C. Roos; The use of several microsatellite loci applied to 13 neotropical primates revealed a strong recent bottleneck event in the woolly monkey (Lagothrix lagotricha) in Colombia, by M. Ruiz-Garcia; Relative humidity, ambient temperature, and urine washing behavior in Bolivian squirrel monkeys, Saimiri boliviensis boliviensis, by J. C. Ruiz; and Adenocarcinoma of the uterus in a common marmoset (Callithrix jacchus), by J. Kaspareit, S. Friderichs-Gromoll, E. Buse, & G. Habermann.
Monographs
* The emergence of social cognition in three young chimpanzees. M. Tomasello & M. Carpenter. Monographs of the Society for Research in Child Development, Serial No. 279, 70[1]. Boston: Blackwell, 2005.
. . . A report on a series of 10 studies, previously conducted with human infants. Taken together, the findings support the idea that the early ontogeny of human social cognition comprises two distinct trajectories, each with its own evolutionary history: one for understanding the basics of goal-directed action and perception, common to all apes, and another for sharing psychological states with others in collaborative acts involving joint intentions and attention, unique to the human species.
Special Journal Issues
* Advances in field behavioral endocrinology. K. B. Strier & T. E. Ziegler (Guest Eds.). American Journal of Primatology, 2005, 67[1].
. . . Contents: Introduction: Advances in field-based studies of primate behavioral endocrinology, by K. B. Strier & T. E. Ziegler; Effects of reproductive and social variables on fecal glucocorticoid levels in a sample of adult male ring-tailed lemurs Lemur catta) at the Beza Mahafaly Reserve, by L. Gould, T. E. Ziegler, & D. J. Wittwer; Social and reproductive factors affecting cortisol levels in wild female golden lion tamarins (Leontopithecus rosalia), by K. L. Bales, J. A. French, C. M. Hostetler, & J. M. Dietz; Behavioral strategies and hormonal profiles of dominant and subordinate common marmoset (Callithrix jacchus) females in wild monogamous groups, by M. B. Cordeiro Sousa, A. C. Sales da Rocha Albuquerque, F. da Silva Albuquerque, A. Araujo, M. E. Yamamoto, & M. de Fatima Arruda; Behavioral indicators of ovarian phase in white-faced capuchins (Cebus capucinus), by S. D. Carnegie, L. M. Fedigan, & T. E. Ziegler; Variation in the resumption of cycling and conception by fecal androgen and estradiol levels in female northern muriquis (Brachyteles hypoxanthus), by K. B. Strier & T. E. Ziegler; Coming of age: Steroid hormones of wild immature baboons (Papio cynocephalus), by L. R. Gesquiere, J. Altmann, M. Z. Khan, J. Couret, J. C. Yu, C. S. Endres, J. W. Lynch, P. Ogola, E. A. Fox, S. C. Alberts, & E. O. Wango; Female testosterone, dominance rank, and aggression in an Ethiopian population of hybrid baboons, by J. C. Beehner, J. E. Phillips-Conroy, & P. L. Whitten; Radioimmunoassay of estrone conjugates from urine dried on filter paper, by C. D. Knott; Reproductive endocrinology of wild female chimpanzees (Pan troglodytes schweinfurthii): Methodological considerations and the role of hormones in sex and conception, by M. E. Thompson; and Fecal steroid research in the field and laboratory: Improved methods for storage, transport, processing, and analysis, by T. E. Ziegler & D. J. Wittwer.
* Special Section: Animal Behaviour. Current Science, 2005, 89[7].
. . . Contents: The animal mind and conservation of species: Knowing what animals know, by D. K. Candland; Behaviour genetics in the post-genomics era: From genes to behaviour and vice versa, by A. Joshi; Zeitgebers (time cues) for biological clocks, by V. K. Sharma & M. K. Chandrashekaran; Neurobiology and behaviour: A network of connections, by R. Balakrishnan; Ecology proposes, behaviour disposes: Ecological variability in social organization and male behavioural strategies among wild bonnet macaques, by A. Sinha, K. Mukhopadhyay, A. Datta-Roy, & S. Ram; Integrating proximate and ultimate causation: Just one more go! by B. Thierry; Role of parasites and commensals in shaping host behaviour, by M. Jog & M. Watve; Variation in male mating behaviour within ungulate populations: Patterns and processes, by K. Isvaran; Social and reproductive behaviour in elephants, by T. N. C. Vidya & R. Sukumar; Midnightís children? Solitary primates and gregarious chiropterans, by S. Radhakrishna; Managing primates in zoos: Lessons from animal behaviour, by A. Mallapur; Mate choice and its implications for conservation and management, by S. Quader; and Behavioural studies: A necessity for wildlife management, by M. Singh & W. Kaumanns.
* Programme of the 1. Congress of the European Federation for Primatology (EFP) and 9. Tagung der Gesellschaft f¸r Primatologie (GfP) August 09-12, 2005. Primate Report, August 2005, 72.
Anatomy and Physiology
* Effects of age and sex on hematologic and serum biochemical values of vervet monkeys (Chlorocebus aethiops sabaeus). Sato, A., Fairbanks, L. A., Lawson, T., & Lawson, G. W. (Div. of Lab. Animal Med., David Geffen School of Med., UCLA, Box 951718, Los Angeles, CA 90095-1718). Contemporary Topics in Laboratory Animal Science, 2005, 44, 29-34.
. . . Hematologic and serum biochemical values are of great importance in assessing animal health. Normal reference ranges for vervet monkeys have seldom been reported, making it difficult for clinicians to interpret blood values. Blood samples were collected from 140 healthy vervet monkeys of Caribbean origin (60 females; 80 males). Male and female data were displayed separately within six life-stage categories (yearlings, juveniles, adolescents, young adults, adults, and aged). The effects of sex and age on these values were examined statistically. Significant age-related factors included red blood cell count, hemoglobin, hematocrit, mean corpuscular volume, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration, alkaline phosphatase, albumin, total protein, globulin, direct bilirubin, blood urea nitrogen, creatinine, glucose, calcium, phosphorus, potassium, albumin/globulin ratio, blood urea nitrogen/creatinine ratio, and sodium/potassium ratio values. Significant sex-related values included red blood cell count, hemoglobin, hematocrit, mean corpuscular hemoglobin, mean corpuscular hemoglobin concentration, neutrophil count, total bilirubin, direct bilirubin, creatinine, glucose, calcium, phosphorus, total carbon dioxide, chloride, potassium, and sodium/potassium ratio values.
* Wild chimpanzees show population-level handedness for tool use. Lonsdorf, E. V., & Hopkins, W. D. (W. D. H., Div. of Psychobiology, Yerkes NPRC, Emory Univ., Atlanta, GA 30322 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2005, 102, 12634-12638.
. . . ìWhether nonhuman primates exhibit population-level handedness remains a topic of considerable theoretical and empirical debate. One continued subject of discussion is whether evidence of population-level handedness in primates is confined to studies in captive animals or whether it is in both captive and wild subjects. Here, we report evidence of population-level handedness in wild chimpanzees for a tool-use task known as ëtermite-fishing.í We also compared the handedness for termite-fishing with other published reports on handedness for nut-cracking and wadge-dipping and found task-specific differences in handedness. Last, combing all of the published data on tool use in wild chimpanzees, we show that hand preferences are heritable. Contrary to previous claims, our results demonstrate that population-level handedness is evident in wild chimpanzees and suggest that the antecedents of lateralization of function associated with hand use were present at least 5 million years ago, before the PanñHomo split.î
* Dental senescence in a long-lived primate links infant survival to rainfall. King, S. J., Arrigo-Nelson, S. J., Pochron, S. T., Semprebon, G. M., Godfrey, L. R., Wright, P. C., & Jernvall, J. (Dept of Anthro., Stony Brook Univ., Stony Brook, NY 11794 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2005, 102, 16579-16583.
. . . ìPrimates tend to be long-lived, and, except for humans, most primate females are able to reproduce into old age. Although aging in most mammals is accompanied by dental senescence due to advanced wear, primates have low- crowned teeth that wear down before old age. Because tooth wear alters crown features gradually, testing whether early dental senescence causes reproductive senescence has been difficult. To identify whether and when low-crowned teeth compromise reproductive success, we used a 20-year field study of Propithecus edwardsi, a rainforest lemur from Madagascar with a maximum lifespan of >27 years. We analyzed tooth wear in three dimensions with dental topographic analysis by using Geographical Information Systems (GIS) technology. We report that tooth wear exposes compensatory shearing blades that maintain dental function for 18 years. Beyond this age, female fertility remains high; however infants survive only if lactation seasons have elevated rainfall. Therefore, low-crowned teeth accommodate wear to a point, after which reproductive success closely tracks environmental fluctuations. These results suggest a tooth-wear-determined, but rainfall-mediated, onset of reproductive senescence. Additionally, our study indicates that even subtle changes in climate may affect reproductive success of rainforest species.î
* Experience induces structural and biochemical changes in the adult primate brain. Kozorovitskiy, Y., Gross, C. G., Kopil, C., Battaglia, L., McBreen, M., Stranahan, A. M., & Gould, E. (C. G G., Dept of Psychology, Princeton Univ. Princeton, NJ 08544 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A.., 2005, 102, 17478-17482.
. . . Primates exhibit complex social and cognitive behavior in the wild. In the laboratory, however, the expression of their behavior is usually limited. A large body of literature shows that living in an enriched environment alters dendrites and synapses in the brains of adult rodents. To date, no studies have investigated the influence of living in a complex environment on brain structure in adult primates. We assessed dendritic architecture, dendritic spines, and synaptic proteins in adult marmosets housed in either a standard laboratory cage or in one of two differentially complex habitats. A month-long stay in either complex environment enhanced the length and complexity of the dendritic tree and increased dendritic spine density and synaptic protein levels in the hippocampus and prefrontal cortex. No differences were detected between the brains of marmosets living in the two differentially complex environments. Our results show that the structure of the adult primate brain remains highly sensitive even to modest levels of experiential complexity. For adult primates, living in standard laboratory housing may induce reversible dendritic spine and synapse decreases in brain regions important for cognition.
Animal Models
* Using siRNA in prophylactic and therapeutic regimens against SARS coronavirus in rhesus macaque. Li, B.-j., Tang, O., Cheng, D., Qin, C., Xie, F. Y., Wei, O., Xu, J., Liu, Y., Zheng, B.-J., Woodle, M. C., Zhong, N., &Ý Lu., P. Y. (P. Y. L., Intradigm Corp, 12115 K Parklawn Dr., Rockville, MD 20852 [e-mail: [email protected]]). Nature Medicine, 2005, 11, 944-951.
. . . Development of therapeutic agents for severe acute respiratory syndrome (SARS) viral infection using short interfering RNA (siRNA) inhibitors exemplifies a powerful new means to combat emerging infectious diseases. Potent siRNA inhibitors of SARS coronavirus (SCV) in vitro were further evaluated for efficacy and safety in a rhesus macaque (Macaca mulatta) SARS model using clinically viable delivery while comparing three dosing regimens. Observations of SARS-like symptoms, measurements of SCV RNA presence and lung histopathology and immunohistochemistry consistently showed siRNA-mediated anti-SARS efficacy by either prophylactic or therapeutic regimens. The siRNAs used provided relief from SCV infection-induced fever, diminished SCV viral levels and reduced acute diffuse alveoli damage. The 10?40 mg/kg accumulated dosages of siRNA did not show any sign of siRNA-induced toxicity. These results suggest that a clinical investigation is warranted and illustrate the prospects for siRNA to enable a massive reduction in development time for new targeted therapeutic agents.
* HIV Gag protein conjugated to a Toll-like receptor 7/8 agonist improves the magnitude and quality of Th1 and CD8+ T cell responses in nonhuman primates. Wille-Reece, U., Flynn, B. J., Loré, K., Koup, R. A., Kedl, R. M., Mattapallil, J. J., Weiss, W. R., Roederer, M., & Seder, R. A. (R. A. S., Cellular Immunology Section, Vaccine Research Center, NIH, Bethesda, MD 20892 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2005, 102, 15190-15194.
. . . ìInduction and maintenance of antibody and T cell responses will be critical for developing a successful vaccine against HIV. A rational approach for generating such responses is to design vaccines or adjuvants that have the capacity to activate specific antigen-presenting cells. In this regard, dendritic cells (DCs) are the most potent antigen-presenting cells for generating primary T cell responses. Here, we report that Toll-like receptor (TLR) agonists and ligands that activate DCs in vitro influence the magnitude and quality of the cellular immune response in nonhuman primates (NHPs) when administered with HIV Gag protein. NHPs immunized with HIV Gag protein and a TLR7/8 agonist or a TLR9 ligand [CpG oligodeoxynucleotides (CpG ODN)] had significantly increased Gag-specific T helper 1 and antibody responses, compared with animals immunized with HIV Gag protein alone. Importantly, conjugating the HIV Gag protein to the TLR7/8 agonist (Gag-TLR7/8 conjugate) dramatically enhanced the magnitude and altered the quality of the T helper 1 response, compared with animals immunized with HIV Gag protein and the TLR7/8 agonist or CpG ODN. Furthermore, immunization with the Gag-TLR7/8 conjugate vaccine elicited Gag-specific CD8+ T responses. Collectively, our results show that conjugating HIV Gag protein to a TLR7/8 agonist is an effective way to elicit broad-based adaptive immunity in NHPs. This type of vaccine formulation should have utility in preventive or therapeutic vaccines in which humoral and cellular immunity is required.î
* Protection of macaques from vaginal SHIV challenge by vaginally delivered inhibitors of virus-cell fusion. Veazey, R. S., Klasse, P. J., Schader, S. M., Hu, Q., Ketas, T. J., Lu, M., Marx, P. A., Dufour, J., Colonno, R. J., Shattock, R. J., Springer, M. S., & Moore, J. P. (J. P. M., Dept of Microbiology & Immunology, Weill Med. College of Cornell Univ., New York, NY 10021 [e-mail: [email protected]. edu]). Nature, 2005, 438, 99-102.
. . . ìHuman immunodeficiency virus type 1 (HIV-1) continues to spread, principally by heterosexual sex, but no vaccine is available. Hence, alternative prevention meth-ods are needed to supplement educational and behavioral-modification programs. One such approach is a vaginal microbicide: the application of inhibitory compounds before intercourse. Here, we have evaluated the microbicide concept using the rhesus macaque ëhigh doseí vagi-nal transmission model with a CCR5-receptor-using simian-human immunodeficiency virus (SHIV-162P3) and three compounds that inhibit different stages of the virus-cell attachment and entry process. These compounds are BMS-378806, a small molecule that binds the viral gp120 glycoprotein and prevents its attachment to the CD4 and CCR5 receptors; CMPD167, a small molecule that binds to CCR5 to inhibit gp120 association; and C52L, a bacterially expressed peptide inhibitor of gp41-mediated fusion. In vitro, all three compounds inhibit infection of T cells and cervical tissue explants, and C52L acts synergistically with CMPD167 or BMS-378806 to inhibit infection of cell lines. In vivo, significant protection was achieved using each compound alone and in combinations. CMPD167 and BMS-378806 were protective even when applied 6Ýh before challenge.î
Animal Welfare
* A note on the influence of visitors on the behaviour and welfare of zoo-housed gorillas. Wells, D. L. (School of Psychology, Queenís University Belfast, Belfast BT7 1NN, Northern Ireland, U.K. [e-mail: [email protected]]). Applied Animal Behaviour Science, 2005, 93, 13-17.
. . . Visitors to zoos can be a source of potential stress to some captive-housed primates. To date, the effect of the human audience on the welfare of captive gorillas has been overlooked. This study examined the behavior of six zoo-housed gorillas during periods of both high and low visitor density, to establish whether there was any effect of audience numbers on the animalsí well-being. Low visitor density encouraged behavior suggestive of relaxation, with gorillas spending a significantly greater proportion of time resting. High visitor density, by contrast, encouraged significantly more intragroup aggression, stereotypies, and autogrooming. The findings suggest that gorillas, like many other species of primate, are excited by high numbers of visitors.
* Hair pulling: A review. Reinhardt, V. (6014 Palmer Dr., Weed, CA 96094 [e-mail: [email protected]]). Lab-oratory Animals, 2005, 39, 361-369.
. . . Hair pulling has been reported in humans, six different nonhuman primate species, mice, guinea pigs, rabbits, sheep and muskoxen, and dogs and cats. This behavior seems to occur only in subjects who are confined in an artificial environment. It has been classified as a mental disorder in humans, and as a behavioral pathology in animals. The hair is not only pulled but also, in most species, ingested. Hair pulling can be both self-directed and partner-directed, contains elements of aggression, manifests more often in females than in males, is associated with psychogenic distress, and resists treatment. Research data collected from affected animals are probably not normative, hence scientifically unreliable. The preemptive correction of husbandry deficiencies causing long-term stress may prevent the development of this bizarre behavior in healthy subjects.
Behavior
* Conformity to cultural norms of tool use in chimpanzees. Whiten, A., Horner, V., & de Waal, F. B. M. (Centre for Social Learning & Cognitive Evolution, Sch. of Psychol-ogy, Univ. of St Andrews, St Andrews, Fife KY16 9JP, U.K. [e-mail: [email protected]]). Nature, 2005, 437, 737-740.
. . . ìRich circumstantial evidence suggests that the extensive behavioral diversity recorded in wild great apes re-flects a complexity of cultural variation unmatched by species other than our own. However, the capacity for cultural transmission assumed by this interpretation has remained difficult to test rigorously in the field, where the scope for controlled experimentation is limited. Here we show that experimentally introduced technologies will spread within different ape communities. Unobserved by groupmates, we first trained a high-ranking female from each of two groups of captive chimpanzees to adopt one of two different tool-use techniques for obtaining food from the same ëPan-pipeí apparatus, then reintroduced each female to her respective group. All but two of 32 chimpanzees mastered the new technique under the influence of their local expert, whereas none did so in a third population lacking an ex-pert. Most chimpanzees adopted the method seeded in their group, and these traditions continued to diverge over time. A subset of chimpanzees that discovered the alternative method nevertheless went on to match the predominant approach of their companions, showing a conformity bias that is regarded as a hallmark of human culture.î
* First observation of tool use in wild gorillas. Breuer, T.,ÝNdoundou-Hockemba, M., &ÝFishlock, V. (Max Planck Inst. for Evolutionary Anthropology, Leipzig, Germany [e-mail: [email protected]]). PLoS Biology, 2005, 3[11], e380, <www.plosbiology.org>.
. . . ìDescriptions of novel tool use by great apes in response to different circumstances aids in understanding the factors favoring the evolution of tool use in humans. This paper documents what we believe to be the first two observations of tool use in wild western gorillas (Gorilla gorilla). We first observed an adult female gorilla using a branch as a walking stick to test water deepness and to aid in her attempt to cross a pool of water at a swampy forest clearing in northern Congo. In the second case we saw another adult female using a detached trunk from a small shrub as a stabilizer during food processing. She then used the trunk as a self-made bridge to cross a deep patch of swamp. In contrast to information from other great apes, which mostly show tool use in the context of food extraction, our observations show that in gorillas other factors, such as habitat type, can stimulate the use of tools.î
* The social nature of primate cognition. Barrett, L., & Henzi, P. (School of Biol. Sciences, Univ. of Liverpool, Crown St, Liverpool L69 7ZB, U.K. [e-mail: [email protected]]). Proceedings of the Royal Society of London, B, 2005, 272, 1865-1875.
. . . The hypothesis that the enlarged brain size of the primates was selected for by social, rather than purely eco-logical, factors has been strongly influential in studies of primate cognition and behavior over the past two decades. However, the Machiavellian intelligence hypothesis, also known as the social brain hypothesis, tends to emphasize certain traits and behaviors, like exploitation and deception, at the expense of others, such as tolerance and behavioral coordination, and therefore presents only one view of how social life may shape cognition. This review outlines work from other relevant disciplines, including evolutionary economics, cognitive science, and neurophysiology, to illustrate how these can be used to build a more general theoretical framework, incorporating notions of embodied and distributed cognition, in which to situate questions concerning the evolution of primate social cognition.
* Chimpanzees are indifferent to the welfare of unrelated group members. Silk, J. B., Brosnan, S. F., Vonk, J., Henrich, J., Povinelli, D. J., Richardson, A. S., Lambeth, S. P., Mascaro, J., & Schapiro, S. J. (Dept of Anthropology, UCLA, Los Angeles, CA 90095 [e-mail: [email protected]]). Nature, 2005, 437, 1357-1359.
. . . ìHumans are an unusually prosocial speciesówe vote, give blood, recycle, give tithes and punish violators of social norms. Experimental evidence indicates that people willingly incur costs to help strangers in anonymous one-shot interactions, and that altruistic behavior is motivated, at least in part, by empathy and concern for the welfare of others (hereafter referred to as other-regarding preferences). In contrast, cooperative behavior in nonhuman primates is mainly limited to kin and reciprocating partners, and is virtually never extended to unfamiliar individuals. Here we present experimental tests of the existence of other-regarding preferences in nonhuman primates, and show that chimpanzees (Pan troglodytes) do not take advantage of opportunities to deliver benefits to familiar individuals at no material cost to themselves, suggesting that chimpanzee behavior is not motivated by other-regarding preferences. Chimpanzees are among the primates most likely to demonstrate prosocial behaviors. They participate in a variety of collective activities, in-cluding territorial patrols, coalitionary aggression, cooperative hunting, food sharing, and joint mate guarding. Consolation of victims of aggression and anecdotal accounts of solicitous treatment of injured individuals suggest that chimpanzees may feel empathy. Chimpanzees sometimes reject exchanges in which they receive less valuable rewards than others, which may be one element of a ësense of fairnessí, but there is no evidence that they are averse to interactions in which they benefit more than others.î
* What perceptual rules do capuchin monkeys (Cebus apella) follow in completing partly occluded figures? Fujita, K., & Giersch, A. (Dept of Psychology, Grad. School of Letters, Kyoto Univ., Sakyo, Kyoto 606-8501, Japan [e-mail: [email protected]]). Journal of Experimental Psychology: Animal Behavior Processes 2005, 31, 387-398.
. . . Capuchin monkeys matched a variety of rodlike figures, distinguishable only by their central portions. The monkeys were then tested for perceptual completion rules by examining which comparison figures they would choose as matching the sample with the critical central portion occluded by a horizontal belt. In Experiment 1, the monkeys overwhelmingly chose a straight rod over disconnected rods and rods having irregular shapes at their centers, irrespective of the presence-absence of common motion of visible parts of the samples. The monkeys chose a connected rod for relatable rods and disconnected rods for nonrelatable ones in Experiment 2. In Experiments 3 and 4, some of the monkeys chose rods specified by the global regularity of the contour, whereas others did not. Experiment 5 showed that humans and capuchin monkeys basically follow similar perceptual rules in completing occluded contours, but the global regularity rule may be stronger in humans.
* Auditory-visual cross-modal perception of communicative stimuli in tufted capuchin monkeys (Cebus apella). Evans, T. A., Howell, S., & Westergaard, G. C. (Language Research Center, Georgia State Univ., 3401 Panthersville Rd, Decatur, GA 30342 [e-mail: [email protected]]). Journal of Experimental Psychology: Animal Behavior Processes, 2005, 31, 399-406.
. . . Research on cross-modal performance in nonhuman primates is limited to a small number of sensory modalities and testing methods. To broaden the scope of this research, the authors tested capuchin monkeys for a seldom-studied cross-modal capacity in nonhuman primates, auditory-visual recognition. Monkeys were simultaneously played two video recordings of a face producing different vocalizations and a sound recording of one of the vocalizations. Stimulus sets varied from naturally occurring conspecific vocalizations to experimentally controlled human speech stimuli. The authors found that monkeys preferred to view face recordings that matched presented vocal stimuli. Their preference did not differ significantly across stimulus species or other stimulus features. However, the reliability of the latter set of results may have been limited by sample size. From these results, the authors concluded that capuchin monkeys exhibit auditory-visual cross-modal perception of conspecific vocalizations.
* Can squirrel monkeys (Saimiri sciureus) plan for the future? Studies of temporal myopia in food choice. McKenzie, T., Cherman, T., Bird, L. R., Naqshbandi, M., & Roberts, W. A. (W. A. R., Dept of Psychology, Univ. of Western Ontario, London, ON N6A 5C2, Canada [e-mail: [email protected]). Learning & Behavior, 2004, 32, 37k7-390.
. . . In seven experiments, two squirrel monkeys were given choices between arrays of food that varied in the quantity offered. In Experiments 1-5, the monkeys were offered choices between quantities of the same food that varied in a 2:1 ratio. The squirrel monkeys failed to show the temporal myopia effect or a decrease in preference for the larger quantity as the absolute number of food items offered increased. Even when given choices of 8 versus 16 peanuts and 10 versus 20 peanuts, both monkeys signifi-cantly preferred the larger quantity. An examination of the monkeysí rates of consumption indicated that 20 peanuts were consumed over a 1- to 2-h period, with eating bouts separated by periods of nonconsumption. In Experiments 6A, 6B, and 7, food was either pilfered or replenished 15 min after an initial choice, so that choice of the smaller quantity led to more total food in the long run. These manipulations caused both monkeys to reduce choice of the larger quantity, relative to baseline choice. The results suggest that squirrel monkeys anticipated the future consequences of their choices.
Care
* Effects of isosexual pair-housing on biomedical implants and study participation in male macaques. Roberts, S. J., & Platt, M. L. (Dept of Neurobiology, Box 3209, Duke Univ. Med. Ctr, Durham, NC 27710 [e-mail: [email protected]). Contemporary Topics in Laboratory Animal Science, 2005, 44[5], 13-18.
. . . ìSocial housing has been shown to contribute to the psychological well-being and physical health of captive primates, and this factor has led to United States Department of Agriculture guidelines requiring facilities to address the social needs of primate species known to live socially in nature as long as doing so does not endanger the animals or interfere with research goals. Although pair-housing is the best way to provide social enrichment, many researchers and facilities are hesitant to implement it, particularly in biomedical research contexts where implanted devices or behavioral performance might be compromised. In order to study the effects of pair-housing on biomedical implants and study participation, we collected data from a group of isosexually pair-housed male macaques (adult and subadult) with 1) cranial and eye implants and 2) controlled access to water as means of motivating subjects to participate in psychophysical studies. Implants, study participation, and weight gain were not adversely affected by pair-housing. Our results support the use of pair-housing as social enrichment for macaques with biomedical implants and controlled access to water.î
* Cortisol response to relocation stress in Garnettís bushbaby (Otolemur garnettii). Watson, S. L., McCoy, J. G., Stavisky, R. C., Greer, T. F., & Hanbury, D. (Dept of Psychology, Univ. of So. Mississippi, Hattiesburg, MS 39402-5025). Contemporary Topics in Laboratory Animal Science, 2005, 44, 22-24.
. . . ìRelocation of research animals, either within a facility or from one facility to another, is assumed to be stressful. Development of appropriate research methodologies may be facilitated by understanding the extent and duration of the physiological response to relocation stress and whether the stress can be buffered by environmental or social factors, such as the presence of a cagemate. To characterize the response to relocation stress in Garnettís bushbaby, we assessed cortisol concentrations in nine female and six male bushbabies during relocation to a different facility; six of the animals were pair-housed at the time of the move and were moved with their respective cagemates. Fecal cortisol was assessed at three time points: 1) baseline (1 day prior to moving); 2) relocation (the day the animals were relocated); and 3) post-relocation (7 days after relocation). Cortisol concentrations were higher at the relocation time point than at baseline and post-relocation, which did not differ. Cortisol concentration did not differ as a function of having a cagemate during relocation. Although relocation resulted in a significant (P < 0.05) increase in cortisol excretion concentration, the levels returned to baseline within 7 days after the stressor.î
* Toxic epidermal necrolysis in two rhesus macaques (Macaca mulatta) after administration of rituximab. Allen, K. P., Funk, A. J., & Mandress, T. D. (Dept of Comp. Med., Univ. of Tennessee Health Science Ctr, Memphis, TN 38163). Comparative Medicine, 2005, 55, 377-381.
. . . ìA 2- and a 7-year-old rhesus macaque developed toxic epidermal necrolysis (TEN) after administration of rituximab. Rituximab, a chimeric monoclonal antibody (mAb) directed against the CD20 antigen on B lymphocytes, is used to treat certain B cell neoplasias. The macaques were part of a gene therapy study that involved administering an adeno-associated viral vector encoding human factor IX (hFIX) to the animals. Both animals developed antibody against hFIX, which eliminated expression of the protein. Rituximab was administered to deplete the population of B cells producing antibodies against the protein. Two days after treatment, the 7-year-old animal developed erythemic skin lesions that rapidly progressed in severity, resulting in epidermal sloughing and ulceration. Despite aggressive treatment with analgesics, antibiotics, and corticosteroids, the animal had to be euthanized 5 days later. The 2-year-old macaque had no reaction to the initial dose of rituxi-mab and received a second infusion 2 weeks later. Two days after drug administration, skin lesions developed; aggressive analgesic, antibiotic, and corticosteroid treatment was initiated, and the lesions resolved. A third rituximab dose was given approximately two months after the second. Skin lesions developed and were treated. The animal made a full recovery. In both cases, skin biopsies were taken and histopathologic findings were consistent with TEN. A severe, life-threatening condition, TEN manifests as an intolerance reaction in the skin. The most common cause of TEN is a response to previous drug administration. To our knowledge, this condition has not been reported in association with rituximab administration in macaques.î
* The efficacy of diazepam treatment for the management of acute wounding episodes in captive rhesus macaques. Tiefenbacher, S., Fahey, M. A., Rowlett, J. K., Meyer, J. S., Pouliot, A. L., Jones, B. M., & Novak, M. A. (New England Primate Research Center, Harvard Med. School, Southborough, MA 017721). Comparative Medicine, 2005, 55, 387-392.
. . . The spontaneous development of self-injurious behavior (SIB) in singly housed monkeys poses a challenge for their management and well-being in captivity. Relatively little information is available on effective treatments for SIB. This study examined the effects of diazepam (Valium) on self-wounding and other abnormal behaviors in eight individually housed male rhesus monkeys (Macaca mulatta). Each monkeyís response to an anxiolytic dose of diazepam (1 mg/kg or greater orally) was compared with the animalís behavior during drug-free periods. When examined across all animals, treatment with diazepam did not significantly alter wounding frequency or rates of self-directed biting without wounding. However, closer examination of the data revealed that four of the animals showed significant decreases in self-biting and wounding frequency (positive responders, PR group), whereas the remaining monkeys showed a trend towards increased wounding frequency (negative responders, NR group). Subsequent examination of colony and veterinary records demonstrated that compared with NR monkeys, PR monkeys had spent significantly more years in individual cage housing and had experienced a greater number of minor veterinary procedures. PR animals also were significantly less likely to have a documented history of self-biting behavior. These findings suggest that SIB is not a homogeneous disorder in rhesus monkeys; rather, distinct subtypes exist that require different treatment approaches.
Conservation
* Global hotspots of species richness are not congruent with endemism or threat. Orme, C. D. L., Davies, R. G., Burgess, M., Eigenbrod, F., Pickup, N., Olson, V. A., Webster, A. J., Ding, T.-S., Rasmussen, P. C., Ridgely, R. S., Stattersfield, A. J., Bennett, P. M., Blackburn, T. M., Gaston, K. J., & Owens, I. P. F. (I.P.F.O., Div. of Biology, Imperial College London, Silwood Park, Ascot, Berkshire SL5 7PY, U.K. [e-mail: [email protected]]). Nature, 2005, 436, 1016-1019.
. . . Biodiversity hotspots have a prominent role in conservation biology, but it remains controversial to what extent different types of hotspot are congruent. It is demonstrated here that hotspots of species richness, threat, and endemism do not show the same geographical distribution. Only 2.5% of hotspot areas are common to all three aspects of diversity, with over 80% of hotspots being idiosyncratic. More generally, there is a surprisingly low overall congruence of biodiversity indices, with any one index explaining less than 24% of variation in the other indices. These results suggest that, even within a single taxonomic class, different mechanisms are responsible for the origin and maintenance of different aspects of diversity. Consequently, the different types of hotspots also vary greatly in their utility as conservation tools.
* Population census of the white-headed langur (Trachypithecus leucocephalus) at Longrui Karst Hills, Guangxi, China. Wang, D., ÝRan, W., Jin, T., &ÝPan, W. (Giant Panda & Wildlife Conservation Research Center, College of Life Sciences, Peking Univ., Beijing, 100871, PRC [e-mail: [email protected]]). Primates, 2005, 46, 219-222.
. . . ìA population census of white-headed langurs was conducted using sleeping-site counts and the line-transect method from July to August 2002Ýat the Longrui Karst Hills. This area had been the largest habitat of the langurs across their range before the 1990s. While our survey revealed 146 old sleeping-sites, no white-headed langurs or fresh sleeping-sites were found in this area. Our study indicated that there had previously been a large population of langurs at Longrui Karst Hills, but now the langurs are possibly locally extinct in this area. If langurs still exist within the area, the population density must be very low. Interviews with local people confirmed that the number of white-headed langurs has been decreasing since the 1980s. Poaching was very common in the past and continues to be a problem at present, and it is likely that this has caused the decrease in the white-headed langur population at Longrui Karst Hills.î
* Towards a culture of biodiversity conservation. Halffter, G. (Inst. de Ecología, A.C., Apdo. Postal 63, 91000 Xalapa, Veracruz, México [e-mail: [email protected]]). Acta Zoológica Mexicana (n.s.) , 2005, 21(2), 133-153.
. . . This article is intended as a synthesis of the ideas, previously presented by the author, on the conservation of biodiversity as well as new proposals, heretofore unpublished. The author focuses on the central theme that in each society the ideas held about the use and the conservation of biodiversity and, in more general terms, about nature, are an integral part of the culture of that society, and as such cannot be separated from its ethical and aesthetic values, or from its socio-economic reality. Three broad approaches to the protection and use of species richness are considered: rustic use, the establishment of protected areas, and land use policy. Three types of protected areas are examined: hunting or game reserves, national parks and biosphere reserves. These conservation strategies appeared at different points in history in response to different needs and are therefore not mutually exclusive. They share the goal of protecting the exceptional richness of animals and plants within a delimited area. The problems facing these approaches, given the increasing evidence of spatial exchange of species, i.e. beta diversity, are examined. A single area is not enough to protect beta diversity, even when it is a big one. In response to this dilemma a completely new alternative is proposed, one that is complementary to the three currently in use ñ that of archipelago reserves.
Disease
* Septicemia and peritonitis in a colony of common marmosets (Callithrix jacchus) secondary to Klebsiella pneumoniae infection. Pisharath, H. R., Cooper, T. K., Brice, A. K., Cinaciolo, R. E., Pistorio, A. L., Wachtman, L. M., Mankowski, J. L., & Newcomer, C. E. (Dept of Comp. Med., Johns Hopkins Univ. Sch. of Med., Ross 4th Floor, 720 Rutland Ave., Baltimore, MD 21205). Contemporary Topics in Laboratory Animal Science, 2005, 44, 35-37.
. . . Six common marmosets from a colony of 50 died over a period of 3 weeks, with the predominant finding of gram-negative bacterial septicemia. Four of these animals died peracutely; the other two were found when they were moribund, and they subsequently died despite clinical intervention. Gram-negative bacterial rods were present in the blood vessels of stained tissues from five of the six marmosets. Three marmosets also had severe fibrinopurulent peritonitis. In addition, one of the marmosets with peritonitis also had purulent mesenteric lymphadenitis with large colonies of gram-negative bacterial rods within dialated colonic crypts. Klebsiella pneumoniae was isolated from multiple organs in three of the marmosets. Clinical evaluation of the entire colony identified four marmosets with anorexia, nasopharyngeal discharge, and diarrhea. These marmosets were treated with enrofloxacin immediately, and they responded well. K. pneumonia could not be cultured from nasal or fecal samples obtained from the colony animals. Because of the peracute nature of the disease, animals often die before exhibiting clinical symptoms, and antibiotics are seldom helpful. In this outbreak both of the major forms of Klebsiella infection in common marmosets were seen: the peracute form with bacteremia and minimal inflammatory reaction around blood vessels, and the chronic form with bacteremia, fibrinopurulent peritonitis, and mesenteric lymphadenitis.
* Laboratory wash resistance of long-lasting insecticidal nets. Gimnig, J. E., Lindblade, K. A., Mount, D. L., Atieli, F. K., Crawford, S., Wolkon, A., Hawley, W. A., & Dotson, E. M. (Div, of Parasitic Diseases, CDC, Mailstop F42, 4770 Buford Hwy, Atlanta, GA 30341 [e-mail: [email protected]]). Tropical Medicine & International Health, 2005, 10, 1002-1029.
. . . Long-lasting insecticidal nets (LLINs) may eliminate the need for retreatment of mosquito nets used for the control of malaria and other vector-borne diseases. The efficacy of LLINs after repeated washing under laboratory conditions has been used to predict long-lasting efficacy under field conditions. We evaluated under laboratory conditions the wash resistance of two LLINs (PermaNetÆ 1.0, Vestergaard-Frandsen, Denmark; OlysetÆ, Sumitomo Chemical Co., Japan), two candidate LLINs (DawaÆ, Siamdutch Mosquito Netting Co., Thailand; Insector, Athanor, France), and a net treated with a process designed to increase its wash resistance, and compared them with conventionally treated nets (deltamethrin, 25Ýmg/m2). Nets of all six types were washed using a standard protocol and tested weekly using WHO cone bioassays with Anopheles gambiae (Kisumu strain). The PermaNetÆ 1.0 was the most wash resistant with >50% mosquito mortality in WHO cone bioassays after as many as 20 washes. The DawaÆ net also retained some activity after repeated washing but exhibited wide variation in insecticide retention and biological activity. The remaining nets lost >90% of their biological activity after six washes as measured by 24-h mortality of A. gambiae in WHO cone tests. After 20 washes, all nets lost >50% of their initial insecticide concentrations except for the OlysetÆ net. After 20 washes, nets were heated for 4Ýh at 60Ý∞C to determine whether biological activity could be restored by heat-assisted regeneration. Only the OlysetÆ net was regenerated by heating, with average mosquito mortality and knockdown in WHO cone tests rising to >90% after heating for 4Ýh at 60Ý∞C. However, regeneration of the biological activity of OlysetÆ nets that had been washed three times did not occur at 30Ý∞C or 35Ý∞C after 12Ýweeks. The wash resistance of these LLINs corresponded well to their retention of biological activity observed in a field trial, suggesting that wash resistance may be a good predictor of the longevity of insecticidal activity of LLINs under field conditions.
* Repellent properties of celery, Apium graveolens L., compared with commercial repellents, against mosquitoes under laboratory and field conditions. Tuetun, B., Choochote, W., Kanjanapothi, D., Rattanachanpichai, E., Chaithong, U., Chaiwong, P., Jitpakdi, A., Tippawangkosol, P., Riyong, D. & Pitasawat, B. (B. P., Dept of Parasitology, Fac. of Med., Chiang Mai Univ., Chiang Mai 50200, Thailand [e-mail: [email protected]]). Tropical Medicine & International Health, 2005,Ý10,Ý1190-1198.
. . . ìIn our search for new bioactive products against mosquito vectors, we reported the slightly larvicidal and adulticidal potency, but remarkable repellency of Apium graveolens both in laboratory and field conditions. Repellency of the ethanolic preparation of hexane-extracted A. graveolens was, therefore, investigated and compared with those of 15 commercial mosquito repellents including the most widely used, DEET. Hexane-extracted A. graveolens showed a significant degree of repellency in a dose-dependent manner with vanillin added. Ethanolic A. graveolens formulations (10 25% with and without vanillin) provided 2 5Ýh protection against female Aedes aegypti. Repellency of the most effective repellent, 25% of hexane-extracted A. graveolens with the addition of 5% vanillin, was comparable to that obtained from 25% of DEET with 5% vanillin added. Moreover, commercial repellents, except formulations of DEET, showed lower repellency than that of A. graveolens extract. When applied on human skin under field conditions, the hexane-extracted A. graveolens plus 5% vanillin showed a strong repellent action against a wide range of mosquito species belonging to various genera. It had a protective effect against Aedes gardnerii, A. lineatopennis, Anopheles barbirostris, Armigeres subalbatus, Culex tritaeniorhynchus, C. gelidus, C. vishnui group and Mansonia uniformis. The hexane-extracted A. graveolens did not cause a burning sensation or dermal irritation when applied to human skin. No adverse effects were observed on the skin or other parts of the human volunteersí bodies during 6Ýmonths of the study period or in the following 3Ýmonths, after which time observations ceased. Therefore, A. graveolens can be a potential candidate for use in the development of commercial repellents that may be an alternative to conventional synthetic chemicals, particularly in community vector control applications.î
* Wave-like spread of Ebola Zaire. Walsh, P. D., Biek, R., & Real, L. A. (Max Planck Inst. for Evolutionary Primatology, Leipzig, Germany [e-mail: [email protected]]). PLoS Biology, 2005, 3[11], e371, <biology.plosjournals.org>.
. . . ìIn the past decade the Zaire strain of Ebola virus (ZEBOV) has emerged repeatedly into human populations in central Africa and caused massive die-offs of gorillas and chimpanzees. We tested the view that emergence events are independent and caused by ZEBOV variants that have been long resident at each locality. Phylogenetic analyses place the earliest known outbreak at Yambuku, Democratic Republic of Congo, very near to the root of the ZEBOV tree, suggesting that viruses causing all other known outbreaks evolved from a Yambuku-like virus after 1976. The tendency for earlier outbreaks to be directly ancestral to later outbreaks suggests that outbreaks are epidemiologically linked and may have occurred at the front of an advancing wave. While the ladder-like phylogenetic structure could also bear the signature of positive selection, our statistical power is too weak to reach a conclusion in this regard. Distances among outbreaks indicate a spread rate of about 50 km per year that remains consistent across spatial scales. Viral evolution is clocklike, and sequences show a high level of small-scale spatial structure. Genetic similarity decays with distance at roughly the same rate at all spatial scales. Our analyses suggest that ZEBOV has recently spread across the region rather than being long persistent at each outbreak locality. Controlling the impact of Ebola on wild apes and human populations may be more feasible than previously recognized.î
* Bushmeat hunting, deforestation, and prediction of zoonoses emergence. Wolfe, N. D., Daszak, P., Kilpatrick, A. M., & Burke, D. S. (Johns Hopkins Bloomberg School of Public Health, G15 N. Wolfe St/E5038, Baltimore, MD 21205 [e-mail: [email protected]]). Emerging Infectious Disease, 2005, 11[12],
<www.cdc.gov/ncidod/EID/vol11no12/04-0789.htm>.
. . . ìUnderstanding the emergence of new zoonotic agents requires knowledge of pathogen biodiversity in wildlife, human-wildlife interactions, anthropogenic pressures on wildlife populations, and changes in society and human behavior. We discuss an interdisciplinary approach combining virology, wildlife biology, disease ecology, and anthropology that enables better understanding of how deforestation and associated hunting leads to the emergence of novel zoonotic pathogens.î
* Fruit bats as reservoirs of Ebola virus. Leroy, E. M., Kumulungui, B., Pourrut, X., Rouquet, P., Hassanin, A., Yaba, P., Délicat, A., Paweska, J. T., Gonzalez, J.-P., & Swanepoel, R. (Centre Internat. de Recherches Méd. de Franceville, BP 769 Franceville, Gabon [e-mail: [email protected]]). Nature, 2005, 438, 575-576.
. . . ìThe first recorded human outbreak of Ebola virus was in 1976, but the wild reservoir of this virus is still un-known. Here we test for Ebola in more than a thousand small vertebrates that were collected during Ebola out-breaks in humans and great apes between 2001 and 2003 in Gabon and the Republic of the Congo. We find evidence of asymptomatic infection by Ebola virus in three species of fruit bat, indicating that these animals may be acting as a reservoir for this deadly virus. Human infection directly from fruit bats might in part be countered by education, as these animals are eaten by local populations living in the outbreak regions.î
Evolution, Genetics, and Taxonomy
* Analysis of the Macaca mulatta transcriptome and the sequence divergence between Macaca and human. Magness, C. L., Fellin, P. C., Thomas, M. J., Korth, M. J., Agy, M. B., Proll, S. C., Fitzgibbon, M., Scherer, C. A., Miner, D. G., Katze, M. G., & Iadonato, S. P. (S. P. I., Illumigen Biosciences Inc., Suite 450, 2203 Airport Way South, Seattle, WA 98134). Genome Biology, 2005, 6, <genomebiology.com/2005/6/7/r60>.
. . . ìWe report the initial sequencing and comparative analysis of the Macaca mulatta transcriptome. Cloned sequences from 11 tissues, nine animals, and three species (M. mulatta, M. fascicularis, and M. nemestrina) were sampled, resulting in the generation of 48,642 sequence reads. These data represent an initial sampling of the putative rhesus orthologs for 6,216 human genes. Mean nucleotide diversity within M. mulatta and sequence divergence among M. fascicularis, M. nemestrina, and M. mulatta are also reported.î
* The highland mangabey Lophocebus kipunji: A new species of African monkey. Jones, T., Ehardt, C. L., Butynski, T. M., Davenport, T. R., Mpunga, N. E., Machaga, S. J., & De Luca, D. W. (Udzungwa Mountains National Park, Box 99, Mangíula, Tanzania [e-mail: [email protected]]. Science, 2005, 308, 1161-1164.
. . . ìA distinct species of mangabey was independently found at two sites 370 kilometers apart in southern Tanzania (Mount Rungwe and Livingstone in the Southern Highlands and Ndundulu in the Udzungwa Mountains). This new species is described here and given the name ëhighland mangabeyí Lophocebus kipunji sp. nov. We place this monkey in Lophocebus, because it possesses non-contrasting black eyelids and is arboreal. L. kipunji is distinguished from other mangabeys by the color of its pelage; long, upright crest; off-white tail and ventrum; and loud call. This find has implications for primate evolution, African biogeography, and forest conservation.î
* Macaca munzala: A new species from western Arunachal Pradesh, northeastern India. Sinha, A., Datta, A., Madhusudan, M. D., & Mishra, C. (Nature Conservation Foundation, 3076/5, 4th Cross, Gokulam Park, Mysore, India [e-mail: [email protected]]). International Journal of Primatology, 2005, 26, 977-989.
. . . ìMacaca, comprising 20 well-characterized species, represents the largest and one of the most ecologically and socially diverse of all the nonhuman primate genera. We report the discovery of a macaque that is new to science from the high altitudes of western Arunachal Pradesh, a biodiversity-rich state in northeastern India. We propose the scientific name Macaca munzala and the vernacular name Arunachal macaque for the species. It shares morphological characteristics independently with the Assamese macaque (Macaca assamensis) and with the Tibetan macaque M. thibetana; like them, it appears to belong to the sinica species-group of the genus. However, the species is distinctive in relative tail length, which is intermediate between those of Tibetan and Western Assamese macaques, the subspecies with which it is sympatric. It is also unique in its altitudinal distribution, occurring largely at altitudes between 2000 and 3500 m. We provide a morphological characterization of the species, report preliminary data on its field biology and discuss possible taxonomic identity in relation to the other closely-related species of Macaca.î
* Evidence from opsin genes rejects nocturnality in ancestral primates. Tan, Y., Yoder, A. D., Yamashita, N., & Li, W.-H. (W.-H. L., Dept of Ecology & Evolution, Univ. of Chicago, 1101 East 57th Street, Chicago, IL [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2005, 102, 14712-14716.
. . . ìIt is firmly believed that ancestral primates were nocturnal, with nocturnality having been maintained in most prosimian lineages. Under this traditional view, the opsin genes in all nocturnal prosimians should have undergone similar degrees of functional relaxation and accumulated similar extents of deleterious mutations. This expectation is rejected by the short-wavelength (S) opsin gene sequences from 14 representative prosimians. We found severe defects of the S opsin gene only in lorisiforms, but no defect in five nocturnal and two diurnal lemur species and only minor defects in two tarsiers and two nocturnal lemurs. Further, the nonsynonymous-to-synonymous rate ratio of the S opsin gene is highest in the lorisiforms and varies among the other prosimian branches, indicating different time periods of functional relaxation among lineages. These observations suggest that the ancestral primates were diurnal or cathemeral and that nocturnality has evolved several times in the prosimians, first in the lorisiforms but much later in other lineages. This view is further supported by the distribution pattern of the middle-wavelength (M) and long-wavelength (L) opsin genes among prosimians.î
* Further evidence for small-bodied hominins from the Late Pleistocene of Flores, Indonesia. Morwood, M. J., Brown, P., Jatmiko, Sutikna, T., Wahyu Saptomo, E., Westaway, K. E., Rokus Awe Due, Roberts, R. G., Maeda, T., Wasisto, S., & Djubiantono, T. (Archaeol. & Palaeoanth., School of Human & Environ. Studies, Univ. of New England, Armidale, New South Wales 2351, Australia [e-mail: [email protected]]). Nature, 2005, 437, 1012-1017.
. . . ìHomo floresiensis was recovered from Late Pleisto-cene deposits on the island of Flores in eastern Indonesia, but has the stature, limb proportions and endocranial volume of African Pliocene Australopithecus. The holo-type of the species (LB1), excavated in 2003 from Liang Bua, consisted of a partial skeleton minus the arms. Here we describe additional H. floresiensis remains excavated from the cave in 2004. These include arm bones belonging to the holotype skeleton, a second adult mandible, and postcranial material from other individuals. We can now reconstruct the body proportions of H. floresiensis with some certainty. The finds further demonstrate that LB1 is not just an aberrant or pathological individual, but is representative of a long-term population that was present during the interval 95-74 to 12 thousand years ago. The excavation also yielded more evidence for the depositional history of the cave and for the behavioral capabilities of H. floresiensis, including the butchery of Stegodon and use of fire.î
* Basal anthropoids from Egypt and the antiquity of Africaís higher primate radiation. Seiffert, E. R., Simons, E. L., Clyde, W. C., Rossie, J. B., Attia, Y., Bown, T. M., Chatrath, P., & Mathison, M. E. (Dept of Earth Sciences, Univ. of Oxford, Parks Rd, Oxford OX1 3PW, U.K. [e-mail: [email protected]]). Science, 2005, 310, 300-304.
. . . ìEarly anthropoid evolution in Afro-Arabia is poorly documented, with only a few isolated teeth known from before ~35 million years ago. Here we describe craniodental remains of the primitive anthropoid Biretia from ~37-million-year-old rocks in Egypt. Biretia is unique among early anthropoids in exhibiting evidence for nocturnality, but derived dental features shared with younger parapithecids draw this genus, and possibly >45-million-year-old Algeripithecus, into a morphologically and behaviorally diverse parapithecoid clade of great antiquity.î
* Evolution of human-chimpanzee differences in malaria susceptibility: Relationship to human genetic loss of N-glycolylneuraminic acid. Martin, M. J., Rayner, J. C., Gagneux, P., Barnwell, J. W., & Varki, A. (A. V., Dept of Cellular and Molecular Medicine, UCSD, La Jolla, CA 92093 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2005, 102, 12819-12824.
. . . ìChimpanzees are the closest evolutionary cousins of humans, sharing >99% identity in most protein sequences. Plasmodium falciparum is the major worldwide cause of malaria mortality. Plasmodium reichenowi, a morphologically identical and genetically very similar parasite, infects chimpanzees but not humans. Conversely, experimental P. falciparum infection causes brief moderate parasitization and no severe infection in chimpanzees. This surprising host specificity remains unexplained. We modified and enhanced traditional methods for measuring sialic acid (Sia)-dependent recognition of glycophorins by merozoite erythrocyte-binding proteins, eliminating interference caused by endogenous Sias on transfected cells, and by using erythroleukemia cells to allow experimental manipulation of Sia content. We present evidence that these remarkable differences among such closely related host-parasite pairs is caused by species-specific erythrocyte-recognition profiles, apparently related to the human-specific loss of the common primate Sia N-glycolylneura-minic acid. The major merozoite-binding protein erythrocyte-binding antigen-175 of P. falciparum apparently evolved to take selective advantage of the excess of the Sia N-acetylneuraminic acid (the precursor of N-glycolylneura-minic acid) on human erythrocytes. The contrasting preference of P. reichenowi erythrocyte-binding antigen-175 for N-glycolylneuraminic acid is likely the ancestral condition. The surprising ability of P. falciparum to cause disease in New World Aotus monkeys (geographically isolated from P. falciparum until arrival of peoples from the Old World) can be explained by parallel evolution of a human-like Sia expression pattern in these distantly related primates. These results also have implications for the prehistory of hominids and for the genetic origins and recent emergence of P. falciparum as a major human pathogen.î
* Radiocarbon dating of interstratified Neanderthal and early modern human occupations at the Chatelperronian type-site. Gravina, B., Mellars, P., & Ramsey, C. B. (P. M., Dept of Archaeology, Cambridge Univ., Downing St, Cambridge CB2 3DZ, U.K. [e-mail: [email protected]]). Nature, 2005, 438, 51-56.
. . . ìThe question of the coexistence and potential interaction between the last Neanderthal and the earliest intru-sive populations of anatomically modern humans in Europe has recently emerged as a topic of lively debate in the archaeological and anthropological literature. Here we report the results of radiocarbon accelerator dating for what has been reported as an interstratified sequence of late Neanderthal and early anatomically modern occupations at the French type-site of the Chatelperronian, the Grotte des Fées de Ch‚telperron, in east-central France. The radio-carbon measurements seem to provide the earliest secure dates for the presence of Aurignacian technology ñ and from this, we infer the presence of anatomically modern human populations ñ in France.î
* Genetic diversity and reproductive success in mandrills (Mandrillus sphinx). Charpentier, M., SetchellÝ, J. M., Prugnolle, F., Knapp, L. A.,ÝWickings, E. J., Peignot, P., & Hossaert-McKey, M. (Centre díEcologie Fonctionnelle et Evolutive, UMR 5175, CNRS, 1919 Route de Mende, 34293 Montpellier Cedex 5, France [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2005, 102, 16723-16728.
. . . ìRecent studies of wild animal populations have shown that estimators of neutral genetic diversity, such as mean heterozygosity, are often correlated with various fitness traits, such as survival, disease susceptibility, or reproductive success. We used two estimators of genetic diversity to explore the relationship between heterozygosity and reproductive success in male and female mandrills living in a semifree-ranging setting in Gabon. Because social rank is known to influence reproductive success in both sexes, we also examined the correlation between genetic diversity and social rank in females, and acquisition of alpha status in males, as well as length of alpha male tenure. We found that heterozygous individuals showed greater reproductive success, with both females and males producing more offspring. However, heterozygosity influenced reproductive success only in dominant males, not in subordinates. Neither the acquisition of alpha status in males, nor social rank in females, was significantly correlated with heterozygosity, although more heterozygous alpha males showed longer tenure than homozygous ones. We also tested whether the benefits of greater genetic diversity were due mainly to a genome-wide effect of inbreeding depression or to heterosis at one or a few loci. Multilocus effects best explained the correlation between heterozygosity and reproductive success and tenure, indicating the occurrence of inbreeding depression in this mandrill colony.î
* Phylogenetic relationships among some Ateles species: the use of chromosomic and molecular characters. Nieves, M., Ascunce, M. S., Rahn, M. I., & Mudry, M. D. (Grupo de Investigación en Biología Evolutiva, Dept. de Ecología, Genética y Evolución, Fac. de Ciencias Exactas y Naturales, Univ. de Buenos Aires, Ciudad Universitaria, Pabellón II, 4to Piso Lab.: 46-47, C1428EHA Buenos Aires, Argentina [e-mail: [email protected]]). Primates, 2005, 46, 155-164.
. . . ìAs with most platyrrhines, the systematics of Ateles is under discussion. In order to help clarify its systematic, we employed chromosomic and molecular characters to analyze the phylogenetic relationship among some species of the genus Ateles. Chromosomic studies were conducted on 14 atelid specimens: eight Ateles from A. paniscus, A. chamek, A. belzebuth and A. geoffroyi, and six Alouatta caraya. Ateles paniscus showed 2N=32, whereas A. chamek, A. belzebuth and A. geoffroyi presented 2N=34, XX/XY (with a submetacentric X and a variable Y) corroborated by male meiosis. Nucleotide sequence variation at the mitochondrial cytochrome c oxidase subunit II gene (COII) was analyzed in ten New World monkey specimens. Parsimony trees showed consistent phylogenetic relationships using both chromosomic forms and mitochondrial COII gene sequences as characters. Particularly, chromosomic phylogenies showed A. hybridus as a divergent taxon from the remaining group, whereas A. chamek, A. belzebuth and A. marginatus form an unresolved clade with A. geoffroyi as sister group.î
* Subspecies composition and founder contribution of the captive U.S. chimpanzee (Pan troglodytes) population. Ely, J. J., Dye, B., Frels, W. I., Fritz, J., Gagneux, P., Khun, H. H., Switzer, W. M., & Lee, D. R. (Alamogordo Primate Facility, Building 1303, P.O. Box 956, Holloman AFB, NM 88330-0956 [e-mail: [email protected]]). American Journal of Primatology, 2005, 67, 223-241.
. . . Chimpanzees are presently classified into three subspecies: Pan troglodytes verus from west Africa, P. t. troglodytes from central Africa, and P .t. schweinfurthii from east Africa. A fourth subspecies (P. t. vellerosus), from Cameroon and northern Nigeria, has been proposed. These taxonomic designations are based on geographical origins and are reflected in sequence variation in the first hypervariable region (HVR-I) of the mtDNA D-loop. Although advances have been made in our understanding of chimpanzee phylogenetics, little has been known regarding the subspecies composition of captive chimpanzees. We sequenced part of the mtDNA HVR-I region in 218 African-born population founders and performed a phylogenetic analysis with previously characterized African sequences of known provenance to infer subspecies affiliations. Most founders were P .t. verus (95.0%), distantly followed by the troglodytes schweinfurthii clade (4.6%), and a single P. t. vellerosus (0.4%). Pedigree-based estimates of genomic representation in the descendant population revealed that troglodytes schweinfurthii founder representation was reduced in captivity, vellerosus representation increased due to prolific breeding by a single male, and reproductive variance resulted in uneven representation among male P. t. verus founders. No increase in mortality was evident from between-subspecies interbreeding, indicating a lack of outbreeding depression. Knowledge of subspecies and their genomic representation can form the basis for phylogenetically informed genetic management of extant chimpanzees to preserve rare genetic variation for research, conservation, or possible future breeding.
Field Studies
* Dispersal patterns among three species of squirrel monkeys (Saimiri oerstedii, S. boliviensis and S. sciureus): I. Divergent costs and benefits. Boinski, S., Kauffman, L., Ehmke, E., Schet, S., & Vreedzaam, A. (Dept of Anthropology, 1112 Turlington Hall, P.O. Box 117305, Univ. of Florida, Gainesville, FL 32611-7305 [e-mail: [email protected]]). Behaviour, 2005, 142,Ý525-632.
* Dispersal patterns among three species of squirrel monkeys (Saimiri oerstedii, S. boliviensis and S. sciureus): II. Within-species and local variation. Boinski, S., Ehmke, E., Kauffman, L., Schet, S., & Vreedzaam, A. (Address same as above). Behaviour, 2005, 142,Ý633-677.
* Dispersal patterns among three species of squirrel monkeys (Saimiri oerstedii, S. boliviensis and S. sciureus): III. Cognition. Boinski, S. (Address same as above). Behaviour, 2005, 142,Ý679-699.
. . . ìHere we present the first report on dispersal in a wild population of the Neotropical primate Saimiri sciureus (Primates: Cebidae). Long-term observations (1998-2001) of this squirrel monkey represent part of a broader study of the forest community at Raleighvallen in the Central Suriname Nature Reserve. These new dispersal records for S. sciureus are combined with comparable information from congeners, S. boliviensis in Peru and S. oerstedii in Costa Rica. The resulting three-way compilation includes the ecological, social and mating context for each congener. Further enhancing the inherent phylogenetic control of a within-genus comparison, these data were collected with the explicit intent of joint analyses, and the study sites for these small, arboreal social mammals are three of the least disturbed extant Neotropical forests in the historical record. Saimiri appears to merit description as the genus with the most diverse set of species dispersal patterns yet documented among mammals. (1) S. sciureus of both sexes undertake dispersal on several to many occasions during their lifetime. Females and immatures commonly transfer between troops. The large portion of male S. sciureus spend their adult years as solitary or peripheral males. Few males attain secure residence in a mixed-sex troop, a prerequisite for mating success. (2) On attainment of sexual maturity, male S. boliviensis emigrate with their same-age cohort, first joining all-male bands, and eventually entering mixed-sex troops with this same natal male birth cohort. Natal female S. boliviensis are philopatric and form cohesive matrilines. Within-troop competition determines each matrilineís priority of access to fruit resources. (3) In contrast to both S. sciureus and S. boliviensis, S. oerstedii males are philopatric and maintain tight affiliation with same-age-cohort males. Natal female S. oerstedii emigrate as juveniles prior to their first mating season, and may undertake secondary dispersals in subsequent years.î
* Biogeographical and floristic predictors of the presence and abundance of mantled howlers (Alouatta palliata mexicana) in rainforest fragments at Los Tuxtlas, Mexico. Cristóbal-Azkarate, J., Veý, J. J., Asensio, N., & Rodríguez-Luna, E. (Apo Postal 199, C.P. 91001, Xalapa, Veracruz, México [e-mail: [email protected]]). American Journal of Primatology, 2005, 67, 209-222.
. . . ìThis research focuses on identifying the principal habitat characteristics that influence the presence and abundance of mantled howlers in forest fragments. We provide information on the demography of several fragmented Alouatta palliata mexicana subpopulations at Los Tuxtlas, Mexico, and relate this to the biogeographical and floristic characteristics of the forest fragments inhabited. The most important habitat characteristics related to the presence and abundance of howlers in the fragments were fragment size and floristic diversity. On the other hand, some evidence suggests that given the conditions under which howlers in our study area live (i.e., small and degraded fragments with high densities), secondary vegetation may be beneficial for the survival of the howlers. Finally, we discuss the possibility that the very low immature-to-female ratio (IFR) in the groups, and the lack of juveniles found in many of the study groups, may be due to high mortality rates in immatures. A reduction in food availability because of the high population densities of these groups may be responsible for this process.î
General
* A unique biomedical resource at risk. VandeBerg, J. L., & Zola, S. M. (Southwest NPRC, San Antonio, TX 78245). Nature, 2005, 437, 30-32.
. . . Research using chimpanzees has been crucial in the fight against human diseases such as hepatitis. VandeBerg, Zola, and colleagues on the U.S. National Chimpanzee Resource Committee urge that this now dwindling resource be sustained.
Instruments and Techniques
* A disposable vaginal speculum for the chimpanzee (Pan troglodytes). Hasselschwert, D. L., & Fontenot, M. B. (Univ. of Louisiana at Lafayette New Iberia Research Center, New Iberia, LA 70560). Contemporary Topics in Laboratory Animal Science, 2005, 44, 50-52.
. . . The unanticipated under-utilization of chimpanzees by the National Institutes of Health (NIH) for current NIH-supported research has prompted the application of a variety of contraceptive forms. Among the many methods attempted, the intrauterine device (IUD) has been described as having an efficacy similar to that described for humans. In addition, the device is both a financially sound and practical solution to prevention of overpopulation, while minimizing disruption of normal cyclic hormonal fluctuations. However, variations in the size and shape of the sexual skin of the female chimpanzee and the depth of the vaginal vault have posed physical constraints on the use of instrumentation developed for humans for assisting with insertion of the device. In addition the literature is lacking in specific methodology for pelvic examination in chimpanzees. Previously, the University of Louisiana at Lafayette New Iberia Research Center used a 3-ml plastic syringe case as a vaginal speculum to accommodate requirements of an approved research protocol in African green monkeys (Chlorocebus aethiops). Similarly, a simple and effective disposable vaginal speculum was developed for the chimpanzee. The closed tip of a plastic syringe case of appropriate size (20 to 60 ml) was removed to provide an open tube; momentary heating smoothed the cut edge. This simple speculum allowed for sufficient visualization of the cervix for assessment and assistance in the insertion of the IUD. Variously sized speculums were prepared to accommodate differences in animal size. This simple and effective speculum was clean, disposable, and inexpensive.
* Flow cytometric sorting of fresh and frozen-thawed spermatozoa in the western lowland gorilla (Gorilla gorilla gorilla). OíBrien, J. K., Stojanov, T., Crichton, E. G., Evans, K. M., Leigh, D., Maxwell, W. M. C., Evans, G., & Loskutoff, N. M. (Centre for Advanced Technologies in Animal Genetics and Reproduction, Fac. of Vet. Science, Univ. of Sydney, Sydney, NSW 2006, Australia [e-mail: [email protected]]). American Journal of Primatology, 2005, 66, 297-315.
. . . ìWe adapted flow cytometry technology for high-purity sorting of X chromosome-bearing spermatozoa in the western lowland gorilla. Our objectives were to develop methodologies for liquid storage of semen prior to sorting, sorting of liquid-stored and frozen-thawed spermatozoa, and assessment of sorting accuracy. In study 1, the in vitro sperm characteristics of gorilla ejaculates from one male were unchanged (P>0.05) after 8 hr of liquid storage at 15∞C in a non-egg yolk diluent (HEPES-buffered modified Tyrodeís medium). In study 2, we examined the efficacy of sorting fresh and frozen-thawed spermatozoa using human spermatozoa as a model for gorilla spermatozoa. Ejaculates from one male were split into fresh and frozen aliquots. X-enriched samples derived from both fresh and frozen-thawed human semen were of high purity, as determined by fluorescence in situ hybridization (FISH; 90.7%+/-2.3%, overall), and contained a high proportion of morphologically normal spermatozoa (86.0%+/-1.0%, overall). In study 3, we processed liquid-stored semen from two gorillas for sorting using a modification of methods for human spermatozoa. The sort rate for enrichment of X-bearing spermatozoa was 7.3+/-2.5 spermatozoa per second. The X-enriched samples were of high purity (single-sperm PCR: 83.7%) and normal morphology (79.0%+/-3.9%). In study 4 we examined frozen-thawed gorilla semen, and the sort rate (8.3+/-2.9 X-bearing sperm/sec), purity (89.7%), and normal morphology (81.4%+/-3.4%) were comparable to those of liquid-stored semen. Depending on the male and the type of sample used (fresh or frozen-thawed), 0.8-2.2% of gorilla spermatozoa in the processed ejaculate were present in the X-enriched sample. These results demonstrate that fresh or frozen-thawed gorilla spermatozoa can be flow cytometrically sorted into samples enriched for X-bearing spermatozoa.î
* Species and sex identification of western lowland gorillas (Gorilla gorilla gorilla), eastern lowland gorillas (Gorilla beringei graueri) and humans. Matsubara, M., Basabose, A. K., Omari, I., Kaleme, K., Kizungu, B., Sikubwabo, K., Kahindo, M., Yamagiwa, J., & Takenaka, O. (Lab. of Human Evolution Studies, Kyoto Univ., Sakyo-ku, KyotoÝ 606-8502, Japan [e-mail: [email protected]]). Primates, 2005, 46, 199-202.
. . . Methods for the identification of the sex and species of individuals from samples non-invasively taken from humans and gorillas were established. Amplification of a segment of amelogenin (AMG), which is an XñY homologous gene, using two pairs of primers from human AMG, revealed both X- and Y-specific bands. The possibility of sex identification was examined by typing the AMG gene using hair and fecal samples from captive western lowland gorillas in Japan and hair samples from wild eastern lowland gorillas in the Kahuzi-Biega National Park, Democratic Republic of Congo, which were sexed by direct observation. Species-specific bands of AMG in gorillas and humans were identified by restriction fragment length polymorphisms analysis. These tests could be used for sexing unidentified individuals of wild western and eastern lowland gorillas, and screening contamination of human DNA from non-invasively acquired samples.
* Assessing dietary protein of colobus monkeys through faecal sample analysis: A tool to evaluate habitat quality. Chapman, C. A., Webb, T., Fronstin, R., Wasserman, M. D., & Santamaria, A. M. (Anthropology Dept, 855 Sherbrooke St West, McGill Univ., Montreal H3A 2T7, Canada [e-mail: [email protected]]). African Journal of Ecology, 2005, 43, 276-278.
. . . ìHere we evaluate the suitability of using the protein content of colobus monkey fecal samples as an index of the protein content in foods eaten. This method would change the tedious task of collecting many potential food items to the relatively easy task of collecting fecal samples. Previous studies of lagomorphs and ruminants have documented a positive correlation between the protein content of foods and fecal protein; however, there are a number of potentially important differences in the digestive physiology of these animals and colobus monkeys. We made this evaluation through a series of feeding trials on Angolan black-and-white colobus (Colobus angolensis), in which we altered the protein content of their foods. Subsequently, we described the protein content of fecal samples of populations of Abyssinian black-and-white (C. guereza) and red colobus (Piliocolobus tephrosceles) found in or near Kibale National Park, Uganda. Samples were collected from a nutritionally stressed population, living in a forest fragment, and an unstressed population from within the national park.î
Reproduction
* Female age-specific reproductive rates, birth seasonality, and infant mortality of ring-tailed lemurs on St. Catherines Island: 17-year reproductive history of a free-ranging colony. Parga, J. A., & Lessnau, R. G. (Univ. of Texas at Austin, Dept of Anthropology, 1 University Station C3200, Austin, TX 78712-0303 [e-mail: [email protected]. edu]). Zoo Biology, 2005, 24, 295-309.
. . . ìTo compare reproductive parameters of a managed population of free-ranging provisioned ring-tailed lemurs (Lemur catta) to wild L. catta populations, we analyzed birth and mortality records collected over the 17-year history of the St. Catherines Island ring-tailed lemur colony. The majority of births in this colony (approximately 80%) occurred in March, which indicates that most females at this site conceived during the first estrus cycle of the breeding season, as do wild ring-tailed lemur females. Females in this provisioned colony reach the age at first parturition (primiparous age) earlier (at 2 years) than do wild ring-tailed lemurs, which typically begin to reproduce at 3 years of age. Female fecundity is lowest at 2 years of age (34%), but increases to 67-100% for females age 3 and older. Survival analyses showed that infant survivorship for very young (2-3 years) and very old females (>10 years) are similar, and are lower than that for reproductively prime-aged females (4-9 years), although this trend is not statistically significant (chi-sq=3.18, df=2, n=125, P=0.203). There was a large amount of variability in infant survival rates among the oldest females, and a few older females (ages 13 and 14) still had high infant survival rates despite their old age. When compared to wild populations, provisioning in this island colony generally caused a decrease in the female primiparous age, a decrease in infant mortality, and an increase in multiple-offspring births (twins or triplets).î
* Maternal age, parity, and reproductive outcome in captive chimpanzees (Pan troglodytes). Roof, K. A., Hopkins, W. D., Izard, M. K., Hook, M., & Schapiro, S. J. (W. D. H., Div. of Psychobiology, Yerkes NPRC, Emory Univ., 954 Gatewood Rd, Atlanta, GA 30322 [e-mail: [email protected]]). American Journal of Primatology, 2005, 67, 199-207.
. . . As early as the 1970s, it was suggested that nonhuman primates may serve as models of human reproductive senescence. In the present study, the reproductive outcomes of 1,255 pregnancies in captive chimpanzees were examined in relation to parity and its covariate, maternal age. The results show that the percentage of positive pregnancy outcomes was negatively correlated with increasing parity. In addition, spontaneous abortions, stillbirths, and caesarian sections were positively correlated with increasing parity. Maternal age, rather than parity, was found to be the most important predictor of negative birth outcome. This study supports research demonstrating reproductive decline and termination in nonhuman primates, and is the first to quantitatively account for this phenomenon in captive female chimpanzees.
------------------------------
We would like to acknowledge Primate-Science as a source for information about new books.
------------------------------
* * * 2006 Directory of Graduate Programs in Primatology and Primate Research
* * *
Awards Granted at 2005 American Society of Primatologists Meeting
Duane Rumbaugh, Distinguished Primatologist
Duane Rumbaugh, a pioneer in comparative psychology and the study of primate learning, intelligence and language, has been honored by ASP as a ìDistinguished Primatologistî. Congratulations!
Education Committeeís Student Prize Award Winners
Honorable Mentions: Brian Kelly: ìPerformance of Rhesus Monkeys (Macaca mulatta) on a scale model taskî (Oral Presentation). Rachel Dvoskin: ìFunctional variation of the Macaca mulatta CRH gene is associated with temperament and reproductive outcomeî (Paper Presentation). Paper Winner: Mitchell Irwin: ìLiving in forest fragments reduces group cohesion in diademed sifakas (Propithecus diademi) in Eastern Madagascar by reducing patch size of food resourcesî. Poster Winner: Dorothy Mandell: ìComputer training procedures and software for use with nonhuman primate infantsî.
Research and Development Grant Award Winners
* * *
SCAW Presents Harry C. Rowsell Award
Nelson L. Garnett, DVM, and W. Ron DeHaven, DVM, MBA, are the Scientists Center for Animal Welfareís (SCAW) 2005 Harry Rowsell Award winners. The Rowsell Award, established by SCAW in 1992, is given to honorees known for their commitment to good science and the humane treatment of animals used in research, testing and teaching.
Nelson Garnett is currently the Director of the Office of the Johns Hopkins University Animal Care and Use Committee.Ý He retired from the U.S. Public Health Service and his position as Director of the National Institutes of Health Office of Laboratory Animal Welfare.
Ron DeHaven is the Administrator for the U.S. Department of Agricultureís Animal and Plant Health Inspection Service (APHIS). He obtained a DVM degree from Purdue University in 1975 and an MBA from Millsaps College in 1989. After graduating from veterinary school, he spent four years in the U.S. Army Veterinary Corps before beginning his career with APHIS in 1979.
The award is given in honor of Harry Rowsell, OC, DVM, PhD. Dr. Rowsell was the founder and executive director of the Canadian Council on Animal Care and he was a member of the SCAW Board of Trustees from 1983 to 1986. He was also the first recipient of the award.
* * *
Contents of Volumes 40-42 (2001-2003)
Contents of Volumes 43-44 (2004-2005)
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; FAX: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
Current and back issues of the Newsletter are available on the World Wide Web at
http://www.brown.edu/primate
The Newsletter is supported by Brown University
Cover photograph of a cotton-top tamarin (Saguinus Oedipus) at Roger Williams Park Zoo, by Mark Abbott
Copyright (c) 2006 by Brown University
Last updated: December 21, 2005