 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 47 NUMBER 2 APRIL 2008
Printable (PDF) Version of this issue
Articles and Notes
Reduced Cage Mesh Size Interferes with Puzzle Ball Manipulation, by J. M. Sanders, G. H. Lee and C. M. Crockett ......1
Systemic Alopecia Resulting from Hyperadrenocorticism in a Japanese Monkey, by T. Kimura ......5
Effect of Environmental Enrichment on Behavioral and Endocrine Aspects of a Captive Orangutan (Pongo pygmaeus), by C. S. Pizzutto, M. Nichi, M. G. F. G. Sgai, S. H. R. Corrŕa, P. Viau, A. M. Beresca, C. Alvarenga de Oliveira, R. C. BarnabÚ, and M. A. de B. V. GuimarŃes ......10
News, Information, and Announcements
Workshop Announcements ......4
. . . Charles Louis Davis, D.V.M. Foundation Workshop; Callitrichid Behavioral Husbandry and Management; Estimation of Primate Density or Abundance; Environmental Enrichment Training; Neotropical Primates
Volunteers Wanted: Monkey Town Primate Centre ......9
Communication Between Caregivers and Primates: A Survey, by Joanne Davis ......14
Announcements from Publications: Evolution: Education and Outreach ......14
Resources Wanted and Available ......15
. . . Noldus Catalog of Software/Hardware Solutions; New Tree Analysis Program Available Free; Galagos Wanted; Learning about Facility Inspections; Great Ape Distribution Maps; EUPRIM-Net Primate Material Bank
News Briefs ......16
. . . Stella Brewer Marsden; Appointments at OLAWľ Taylor and Hampton; OBE for Shirley McGreal; Railway to Put Gibbons Back on Track; Aye-Aye Born at Bristol Zoo; Federal Bill Will Keep Chimps in Sanctuary; Austriaĺs Supreme Court ľ Chimp Not Person; Malaysian Ministry Drops Plan to Export Monkeys; Ban on Export of Malaysian Macaques Reinstated; Bill to Conceal Names of Animal Researchers; Pruetz Wins National Geographic Society Award; Texas Judge Decides Fate of Chimpanzees
Meeting Announcements ......19
Information Requested or Available ......21
. . . AVMAĺs Responses to Questions about HSVMA; Gorilla Haven Update; More Interesting Websites
Enrichment Event ......21
Educational Opportunities ......23
. . . Training in Veterinary Pathology; Medical and Experimental Mammalian Genetics
Departments
Positions Available ......22
. . . Veterinarian ľ Davis, California; Research Scientist ľ Panama; Assistant Professor ľ Central Washington; Primate Clinical Veterinarian ľ NIH
Recent Books and Articles ......24
* * *
Reduced Cage Mesh Size Interferes with Puzzle Ball Manipulation
Jamie M. Sanders, Grace H. Lee and Carolyn M. Crockett
National Primate Research Center, University of Washington
In the wild, nonhuman primates dedicate much of their time to locating, obtaining, and eating food. When animals are housed in captivity and provided with regular meals, they no longer have the need to forage. Foraging devices can extend feeding time and encourage normal manipulative behavior (Lutz & Novak, 2005). Providing mentally stimulating foraging opportunities to laboratory primates has been an important part of the Washington National Primate Research Centerĺs (WaNPRC) Environmental Enhancement Plan for many years, following Animal Welfare Regulations (USDA, 1991).

Figure 1: Puzzle Ball foraging device fabricated from a Boomer Ball « and stainless steel hardware.
To provide regular foraging opportunities, staff at the WaNPRC in Seattle developed the Puzzle Ball in 1998 (Figure 1). The Puzzle Ball is attached permanently to the outside of an animalĺs cage and can be provisioned easily with peanuts or cereal which the monkey can retrieve from three holes. It is always accessible to be manipulated whether provisioned or not. It is cleaned easily during routine cage washing. By December 2004, Puzzle Balls had been installed on nearly every cage at the Center.
Initial installation of the Puzzle Balls at WaNPRC was associated with significant reductions in abnormal behaviors (Crockett et al., 2001). The efficacy of the Puzzle Ball as an enrichment device, however, depends on an animalĺs ability to manipulate the device. This study was undertaken to assess the impact of a new cage type on one of the WaNPRCĺs most widely used environmental enrichment devices. The main research question was whether the animalsĺ use of the device was impaired by a reduction in the size of the space between horizontal cage bars. Informal observations by WaNPRC staff suggested that the Puzzle Balls installed on the new cages were not being used effectively.
Puzzle balls are normally attached to the front of each cage (see Figure 1). The spacing between bars at the front of the older ôSeattle-styleö cages is 2.75 inches high and, depending on rod size, ranges in width from 0.75 to 0.88 inches (rectangular mesh; cages for larger animals haveálarger diameter rod). On the newly acquired cages, the spacing between bars is 0.75 inches to 0.88 inches for both width and height on all sides of the cage (square mesh, approximately 1 inch by 1 inch, center-on-center). This is equivalent to the square mesh spacing found on one side of the older Seattle-style cages (Bielitzki et al., 1990), which also ranges from 0.75 to 0.88 for both width and height. Figure 2 illustrates the mesh spacing on the front and side of Seattle-style caging used in this study.

Figure 2: Example of Seattle-style caging used in the study..
We hypothesized that monkeys would be less successful in extracting cereal treats from the Puzzle Ball in the reduced spacing condition. We also predicted that monkeys experiencing more difficulty in obtaining treats would display more tension behavior. We took advantage of the availability of both the ôstandardö mesh size and the ôreducedö mesh size on our Seattle-style cages to compare extraction success and tension behaviors in a within-subject design.
Methods
Subjects: Subjects were male (n=6) and female (n=8) pigtailed macaques (Macaca nemestrina) ranging in age from 4.46 to 8.14 years old. All animals were housed at the WaNPRC in individual Seattle-style cages sized appropriately for their weight according to Animal Welfare regulations (USDA, 1991). Some were singly housed and some were in individual cages providing grooming contact (Crockett et al., 1997). Subjects with grooming contact could manipulate the Puzzle Ball without interference from the partner. Only animals whose cages could be observed clearly from the front and square-mesh side were chosen to participate. Cages included a perch, at least one manipulable toy inside the cage, and a Puzzle Ball permanently attached to the outside of the cage. Subjects were fed monkey biscuits twice daily at 9 a.m. and 2 p.m. and water was provided ad libitum. Animals received various food treats according to the Environmental Enhancement Plan.
Procedure: We used a removable Puzzle Ball equipped with a metal clip, allowing for easy attachment and detachment. The experimental Puzzle Ball was attached at the same distance from the top of the cage as the permanently attached Puzzle Ball. In the reduced-mesh condition, the device was attached to the square-mesh side of the Seattle-style cage to simulate the reduced bar spacing of the new cages. In this way, the same subject could be tested with both mesh designs in the same cage. Animals were tested using an ABBA design where A was the reduced mesh condition and B the normal condition. Animals were tested in conditions AB on one day and conditions BA at a later time. All animals had access to their normally provisioned, but now empty, Puzzle Ball throughout the study.
At the beginning of the trial, the observer (JMS) attached the detachable Puzzle Ball to either the front or the side of the cage. Five Cheerios cereal pieces were then loaded into the Puzzle Ball. A standard wristwatch timer was used to record engagement times, defined as physical contact with, or visual exploration of, the device, to the nearest second. If an animal stopped engagement for more than 30 seconds, the timer was stopped and then resumed when the animal reengaged in the activity. The trial was terminated when the animal had extracted and/or dropped all the treats, or after 5 minutes of no engagement. Thus, recorded times reflect active engagement with the device. On a data sheet developed by the observer, success rates were recorded for number of treats extracted and eaten, dropped and unsuccessful, or those still left in the Puzzle Ball. Behavioral codes for tension and activities relevant to the task were adapted from WaNPRCĺs Psychological Well-Being Social Contact Evaluation Data Sheet, and each occurrence was recorded during the timed period. Tension behaviors included yawning, teeth grinding or chomping, vigorous scratching, shaky or violent movements, cage shaking, hitting or biting the Puzzle Ball, eyelid flashing, and head bobbing.
Analysis: Analyses were performed using Microsoft Excel and Data Desk (Velleman, 1997). Within subjects and mesh spacing condition, engagement durations were averaged across presentations. Tension displays were tallied, divided by engagement time to obtain a rate of display, averaged across presentations per condition. Wilcoxon signed-rank matched pairs tests (with adjustment to variance for ties), pairing normal vs. reduced mesh, were performed on engagement times, success (Cheerios successfully extracted and eaten), and rate of tension displays. Within mesh-spacing conditions, we tested whether engagement times decreased from the first to the second presentation. All tests involved 14 pairs. Statistical significance was set at an alpha level of .05, two-tailed.
Results
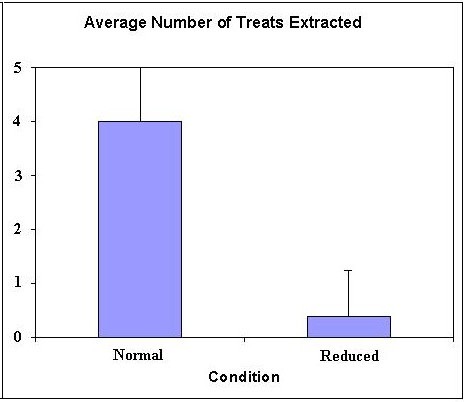
Figure 3: Average number of treats (Cheerios) extracted from Puzzle Balls placed on normal mesh spacing versus reduced mesh spacing (mean and SD). Maximum number is 5 pieces of cereal.
The number of treats extracted and eaten in the reduced mesh condition (M=0.39, SD=.84) was significantly lower than in the normal condition (M=4.00, SD=1.41), Wilcoxon test, p ≤ .001 (Figure 3). Only one of 14 subjects showed evidence of increasing skill by successfully extracting and eating 4 of 5 Cheerios on the second trial after eating none on the first trial with the reduced mesh. One other subject improved from 0 to 1. The duration of engagement with the Puzzle Ball was significantly longer in the reduced condition (M=7.70 minutes, SD=3.72) compared to the normal condition (M=0.97 minutes, SD=0.70), Wilcoxon test, p ≤ .0001. The duration of engagement did not change significantly between first and second presentation on either reduced or normal mesh (Wilcoxon tests, p > .05). The rate of tension displays per minute of engagement was significantly greater in the reduced condition (M=0.33, SD=.21) compared to the normal condition (M=0.04, SD=0.13), Wilcoxon test, p ≤ .0001 (Figure 4).
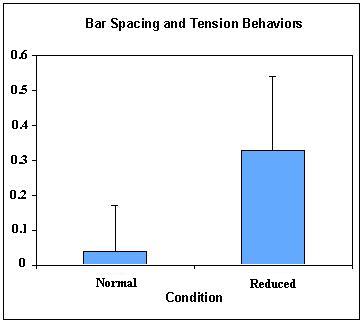
Figure 4: Mean and SD of tension displays per minute of engagement time, comparing normal and reduced mesh sizes.
Discussion
Results from this study support the contention that a reduction in the spacing of cage mesh impairs the ability of adult pigtailed macaques to forage from the Puzzle Ball. In the new caging, there appears to be inadequate access to the Puzzle Ball. Although the reduced access resulted in longer engagement times, significantly greater rates of tension behaviors provides evidence for detrimental effects of using the foraging device on cages that do not provide an animal with adequate access. It is known that monkeysĺ proficiency with the device improves over time (Lloyd et al., 2005), and it is possible that they would learn or devise more effective methods of accessing the Puzzle Ball. However, they might simply give up through frustration. Only one monkey showed notable improvement over the two trials with reduced mesh.
The reduced-mesh condition was confounded with the novelty of a new location for the Puzzle Ball (side of cage). However, this did not seem to deter the subjects as they readily manipulated the device and interacted with it longer. The results of this study also illustrate the paradox of relatively brief device manipulation reinforced by successful extraction of treats versus longer engagement time but associated with frustration, as evidenced by increased tension behaviors. Which is the more enriching experience? We believe that a foraging device should allow successful foraging and that an effective environmental enrichment program should strive to minimize tension and stress among laboratory primates.
This experience has been sobering and insightful. In selecting the new caging, we focused on aspects providing flexible social contact options. It unfortunately did not occur to us that the new caging would compromise the utility of a proven foraging device. A survey of environmental enhancement programs at 22 primate research facilities found that only about half of them provided devices to more than half of their nonhuman primates (Baker et al., 2007). Prior to the acquisition of the new cages, Puzzle Balls had been provided to almost all WaNPRC primates. Now primates in the new cages have reduced foraging options, such as frozen treats, which do not involve a foraging device.
Because the Puzzle Ball is an inexpensive, easily maintained and continuously available environmental enrichment device that allows for sufficient foraging activities as mandated by the USDA (1991), it is an important component of enrichment provided at WaNPRC. To continue to provide animals with this kind of foraging opportunity under the new cage constraints, modifications to the device or caging must be explored. We think that the most feasible solution is to remove some horizontal rod sections to emulate the mesh spacing of the Seattle-style cages, although this will be expensive. Another possibility is to investigate another type of foraging device. Any modifications, though, would need to be tested in order to ascertain whether they would result in engaging the animals in an enriching activity.
References
Baker, K. C., Weed, J. L., Crockett, C. M., & Bloomsmith, M. A. (2007). Survey of environmental enhancement programs for laboratory primates. American Journal of Primatology, 69, 377-394.
Bielitzki, J., Susor, T. G., Elias, K., & Bowden, D. M. (1990). Improved cage design for single housing of social nonhuman primates. Laboratory Animal Science, 40, 428-431.
Crockett, C. M., Bellanca, R. U., Bowers, C. L., & Bowden, D. M. (1997). Grooming-contact bars provide social contact for individually caged laboratory macaques. Contemporary Topics in Laboratory Animal Science, 36, 53-60.
Crockett, C. M., Bellanca, R. U., Heffernan, K. S., Ronan, D. A., & Bonn, W. F. (2001). Puzzle Ball foraging device for laboratory monkeys. Laborataory Primate Newsletter, 40[1], 4-7.
Lloyd, C. R., Lee, G. H., & Crockett, C. M. (2005). Puzzle-ball foraging by laboratory monkeys improves with experience. Laboratory Primate Newsletter, 44[1], 1-3.
Lutz, C. L., & Novak, M. A. (2005). Environmental enrichment for nonhuman primates: Theory and application. Institute for Laboratory Animal Research Journal, 46[2], 178-191.
U. S. Department of Agriculture. (1991). Animal Welfare, Standards, Final Rule (Part 3, Subpart D: Specifications for the humane handling, care, treatment, and transportation of nonhuman primates). Federal Register, 56, 6495-6505.
Velleman, P. F. (1997). Data Desk: The New Power of Statistical Vision. Ithaca, NY: Data Description Inc.
-------------------------------------------
Carolyn Crockett, National Primate Research Center, Box 357330, Univ. of Washington, Seattle, WA 98195-7330 [e-mail: [email protected]]. Research supported by NIH RR00166.
-------------------------------------------
* * *
Workshop Announcements
Charles Louis Davis, D.V.M. Foundation Workshop
A CL Davis Topics in Laboratory Animal Medicine Workshop will be held at the North Carolina State Universityĺs College of Veterinary Medicine, May 15-18, 2008.á May 15-16 will be presentations on various species or relevant topics.á May 17 will be the mock American College of Laboratory Animal Medicine (ACLAM) exam.á NIEHS Comparative Medicine slide sets will be available for review in the CVM library through the evening of May 18.
The full agenda and registration information can be obtained at the CL Davis Website: <www.cldavis.org/courses/upcoming.html#77>. The workshop has been approved for a maximum of 19.5 hours of continuing education by the American Association of Veterinary State Boardsĺ Registry of Approved Continuing Education program.
Information on this and other ACLAM mock exams can be obtained at the Laboratory Animal Boards Study Group Website: <www.labsg.org/2008Mock.html>. Contact David Kurtz [e-mail: [email protected]] if you have any questions.
Callitrichid Behavioral Husbandry and Management
The 7th Annual Callitrichid Behavioral Husbandry and Management Workshop will be held May 17-18, 2008, hosted by the Los Angeles Zoo and Botanical Gardens, Los Angeles, California. This is a workshop for those working with tamarins and marmosets and will include formal presentations, posters, invited speakers, open discussions, and training demonstrations. It will cover a variety of topics such as husbandry, training, enrichment, and conservation. E-mail <[email protected]> for pre-registration form and abstract guidelines. For other information contact Michelle Farmerie [412-365-2385; e-mail: [email protected]].
Estimation of Primate Density or Abundance
A workshop on ôCollection and analysis of field data for estimation of primate density or abundanceö will be given by the Centre for Research into Ecological and Environmental Modelling (CREEM), July 30 to August 1, 2008, prior to the 2008 IPS Congress, on the campus of the University of St. Andrews, one hour by train from Edinburgh. The concepts of distance sampling will be explained and the assumptions of the methods discussed. The focus of the workshop will be on practical application of the methods.
The workshop will start with an introduction to wildlife population assessment methods and a demonstration of how line transect methods are generalizations of sample count methods. The underlying theory and assumptions of line transect sampling will be covered. Computer sessions will follow, training participants in the fundamental use of the software program ôDistanceö.
More sophisticated analysis techniques, such as adjustment for size bias and stratification will be discussed. Time will be made available for discussion of special topics, selected by participants to discuss issues of special interest. Good survey design is an essential ingredient of a successful survey, so design issues and field methods will be covered in detail.
For further information on the workshop see <www.creem.st-and.ac.uk/ocs/index.php/primates/primates08>.
Environmental Enrichment Training
Regional Environmental Enrichment Conferences will sponsor a pre-IPS 2008 Training Workshop July 30 to August 3, 2008, at Edinburgh Zoo, Edinburgh, Scotland. The theme is ôImproving captive primate welfare through good behavioural husbandryö. See <www.reec.info/IPS2008.htm>.
Neotropical Primates
Brookfield Zoo is pleased to announce that it will host a Neotropical Primate Husbandry, Research, and Conservation Conference, Tuesday, October 13 through Thursday, October 15, 2009. This conference will focus on a variety of topics pertaining to neotropical primates and will bring together staff from zoological parks, sanctuaries, and universities, as well as field researchers and range country biologists to share the most current information on husbandry, conservation, and emergent issues pertaining to captive and wild populations of neotropical primates. The workshop will include three days of presentations and a poster session, as well as an icebreaker, silent auction, and banquet. Additional information regarding registration fees, travel information, and submission of abstracts will be made available in late 2008. Please contact Vince Sodaro [708-688-8707; e-mail: [email protected]] for additional information.
* * *
Systemic Alopecia Resulting from Hyperadrenocorticism in a Japanese Monkey
Tohru Kimura
Center for Experimental Animals, National Institutes of Natural Sciences, Aichi, Japan
Introduction
Hyperadrenocorticism is a common disease of dogs caused by chronic overproduction of cortisol by the adrenal cortices. Characteristic clinical features include lethargy, depression, weight loss, persistent anestrus, ascites, polydispsia, polyuria, polyphagia, and pica, as well as alopecia. This disease is well recognized in both human and veterinary medicine (Peterson, 2007; Chiaramonte & Greco, 2007). However, there is little information dealing with hyperadrenocorticism of nonhuman primates (Wilkinson et al., 1999), especially the Japanese monkey (Macaca fuscata). Although simian alopecia may result from hairless mutations, psychogenic alopecia (hair pulling [trichotillomania] and overgrooming) arises from abnormal behaviors (Ahmad et al., 2002; Reinhardt, 2005; Steinmetz et al., 2006). Little systematic research has reported on hair growth and bilateral symmetric alopecia in nonhuman primates. The connections of systemic alopecia with hyperadrenocorticism have not been fully discussed in the skin diseases of nonhuman primates.
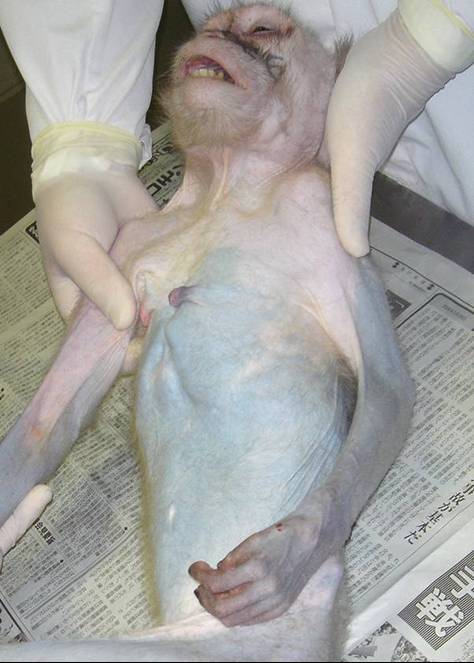
Figure 1: Systemic alopecia on the front of a Japanese monkey.
Recently, I became aware of a female Japanese monkey (Monkey A) suffering from progressive alopecia in the Center for Experimental Animals. This case study identifies, clinically and clinicopathologically, systemic alopecia in Monkey A. Additionally, after the autopsy of the monkey, tissue samples were taken for histopathological examination.
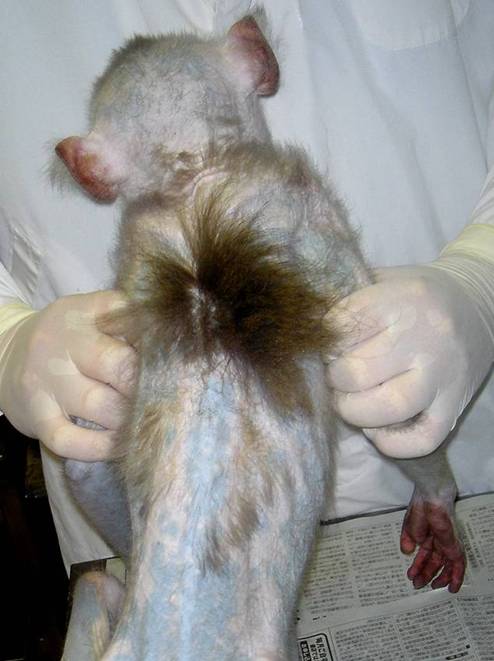
Figure 2: Systemic alopecia on the back, except between the shoulder blades, of a Japanese monkey.
Materials and Methods
Animals: For the previous two years, Monkey A had been used for behavioral studies. She was not given medical or surgical treatment, except for a few symptomatic treatments.
Our monkeys are individually housed in stainless steel cages (60 Î 70 Î 160 cm) in an animal room controlled at 25▒1░ C and 50▒10% relative humidity with 10 to 15 exchanges of 100% fresh air/hr and a 12-hr light (6 a.m. to 6 p.m.), 12-hr dark (6 p.m. to 6 a.m.) cycle. They are fed a commercial primate diet (PS, Oriental Yeast Co., Ltd., Tokyo, Japan), provided ad libitum and supplemented with a variety of fresh fruit, vegetables, and other treats daily. Water is provided through an automatic watering system furnished to each cage.
Procedures: On the day of introduction, our monkeys are anesthetized with a medetomidine (30 μg/kg, Domitol, Meiji Seiyaku Co., Ltd., Tokyo) / midazolam (0.3 mg/kg, Midazolam Injection 0.5 % [F], Fuji Pharmaceutical, Co., Ltd., Tokyo, Japan) / ketamine (2.5 mg/kg, Ketaral 50, Sankyo Co., Ltd., Tokyo, Japan) combination. Complete physical examinations are performed and blood is drawn for hematology and serum biochemistry.
Blood sample collection: Blood samples are collected from the cephalic vein of the animals using no anticoagulant. At 30 minutes after collection of blood samples, sera are separated by centrifugation at 1,500 g for 10 minutes for biochemical analysis and endocrinological examination. For hematological samples, blood is collected into tubes containing K2EDTA.
Hematology: The following parameters are examined using an automated cell counter pocH-iV (Sysmex Co. Ltd, Kobe, Japan): red blood cell count (RBC), hemoglobin concentration (Hb), packed cell volume (PCV), mean corpuscular volume (MCV), mean corpuscular hemoglobin (MCH), mean corpuscular hemoglobin concentration (MCHC), white cell count (WBC), and platelet count (PLT).
A differential WBC count is performed by staining a blood smear with a Romanowsky stain and then examining and classifying 200 WBCs. The classification of WBCs is as follows: neutrophils (band cells [Band] and segmented cells [Seg]), lymphocytes (Lym), monocytes (Mono), eosinophils (Eos), and basophils (Baso).
Serum biochemistry: The following parameters are measured using a blood chemistry analyzer (Dry Chem 3500: Fuji Film Co. Ltd, Tokyo, Japan): total protein (TP), albumin (Alb), albumin/globulin (A/G) ratio, total bilirubin (T-Bil), blood urea nitrogen (BUN), creatinine (Cre), glucose (Glu), triglycerides (TG), total cholesterol (T-Cho), asparate aminotransferase (AST), alanine aminotransferase (ALT), alkaline phosphatase (ALP), lactate dehydrogenase (LDH), creatine kinase (CK), amylase (AMS), C-reactive protein (CRP), electrolytes (Mg, Na, K, Cl, Ca), and inorganic phosphorus (IP).
These procedures were done to Monkey A at her introduction, January 20, 2004, and again, on April 18, 2006, after clinical manifestations of this disease. Endocrinological examinations were also carried out at that time.
Endocrinological examination: For Monkey A, the following parameters were measured in April, 2006, using commercial laboratories: thyroxine (T4), triiodothyronine (T3), free thyroxine (Free T4), free triiodothyronine (Free T3), estradiol, progesterone, adrenocorticotrophic hormone (ACTH), cortisol, catecholamine (adrenalin, noradrenalin and dopamine), and dehydroepiandrosterone sulfate (DHEA-S).
Histological examination: Immediately after her death on May 2, Monkey A was necropsied and tissue samples were taken for histopathological examination.
The tissue specimens were fixed in 10% neutral buffered formalin, and 4-μm paraffin sections were stained with hematoxylin and eosine (HE), and by van Giesonĺs (vG) staining.
Results
| Para- meters | Units | Hyperadrenocorticism | Reference values (n = 75) in NINS | ||
|---|---|---|---|---|---|
| Before | After | ||||
| RBCs | Î1012/l | 4.51 | 5.14 | 5.02 ▒ 0.38 | |
| Hb | g/l | 108 | 130 | 136 ▒ 12 | |
| PCV | l/l | 0.34 | 0.43 | 0.41 ▒ 0.03 | |
| MCV | fl | 76.3 | 83.3 | 81.9 ▒ 4.3 | |
| MCH | pg/cell | 24.0 | 25.3 | 27.1 ▒ 1.5 | |
| CHC | g/l | 314 | 304 | 331 ▒ 15 | |
| PLT | Î109/l | 351 | 281 | 297 ▒ 81 | |
| BCs | Î109/l | 13.7 | 5.1 | 12.5 ▒ 3.8 | |
| Stab | Î109/l | 0.4 | 0.1 | N. D. | |
| Seg | Î109/l | 9.2 | 4.2 | N. D. | |
| Lym | Î109/l | 3.6 | 0.7 | N. D. | |
| Mono | Î109/l | 0.5 | 0.1 | N. D. | |
| Eos | Î109/l | 0 | 0 | N. D. | |
| Baso | Î109/l | 0 | 0 | N. D. | |
Table 1: Hematological findings. Reference values: means ▒ S. D. NINS: National Institutes of Natural Sciences. N. D.: Not done.
Clinical findings: At the time of her introduction (January 20, 2004), there were no dermatological abnormalities. For the next two years, bilaterally progressive symmetric alopecia was gradually observed on her skin surfaces (Figures 1 and 2). During this time, I did not observe behavior disorders such as trichotillomania. A year after her introduction, hair loss began over bony prominences and areas of friction. Except between the shoulder blades, she developed systemic alopecia including the trunk, flanks, chest, arms, legs, and scalp. She showed characteristic clinical features such as lethargy, depression, weight loss, persistent anestrus, ascites, polydispsia, polyuria, polyphagia, and pica (coprophagy and eating wooden environmental enrichments). From these clinical signs and symptoms, she was diagnosed as having systemic alopecia derived from endocrinopathy. The second clinicopathological examination was done on April 18, 2006. Her condition took a turn for the worse, and she died on May 2, 2006.
Hematological findings: Hematological profiles are shown in Table 1. Marked hematological changes occurred in the monkey after the onset of hyperadrenocoticism. Although WBCs decreased by about half of the initial values, the proportion of the segmented cells increased in differential WBC counts. The other patterns of WBCs included no eosinophils and a decline in the number of lymphocytes (0.7 Î 109/l, < 15% on differential WBC counts). Erythrocytic parameters and platelet counts were within normal limits.
| Para- meters | Units | Hyperadrenocorticism | Reference values (n = 75) in NINS | ||
|---|---|---|---|---|---|
| Before | After | ||||
| TP | g/l | 73 | 63 | 69 ▒ 6 | |
| Alb | g/l | 33 | 33 | 43 ▒ 6 | |
| A/G | 0.8 | 1.1 | 1.7 ▒ 0.4 | ||
| T-Bil | μmol/l | 1.7 | 3.4 | 3.4 ▒ 1.7 | |
| BUN | mmol/l | 7.1 | 5.5 | 6.2 ▒ 2.0 | |
| Cre | μmol/l | 53 | 35 | 80 ▒ 35 | |
| Glu | mmol/l | 5.2 | 2.4 | 5.8 ▒ 1.7 | |
| TG | mmol/l | 0.8 | 0.6 | 0.9 ▒ 0.5 | |
| T-Cho | mmol/l | 3.00 | 2.33 | 3.57 ▒ 0.75 | |
| AST | μkat/l | 0.30 | 1.77 | 0.53 ▒ 0.23 | |
| ALT | μkat/l | 0.30 | 4.29 | 0.47 ▒ 0.20 | |
| ALP | μkat/l | 8.1 | 6.7 | 10.6 ▒ 8.2 | |
| LDH | μkat/l | 2.9 | 6.1 | 4.2 ▒ 1.5 | |
| CK | μkat/l | 1.42 | 1.55 | 3.47 ▒ 2.42 | |
| AMS | μkat/l | 4.24 | 3.84 | 3.57 ▒ 1.37 | |
| CRP | nmol/l | 0.07 | < 0.03 | 0.03 ▒ 0.02 | |
| Mg | mmol/l | 0.8 | 1.0 | 0.7 ▒ 0.1 | |
| Na | mmol/l | 137 | 147 | 144 ▒ 5 | |
| K | mmol/l | 4.2 | 4.8 | 3.9 ▒ 0.3 | |
| Cl | mmol/l | 98 | 100 | 102 ▒ 4 | |
| Ca | mmol/l | 2.13 | 2.20 | 2.35 ▒ 0.18 | |
| IP | mmol/l | 1.58 | 1.32 | 1.58 ▒ 0.36 | |
Table 2: Serum biochemical findings.
Serum biochemical findings: Serum biochemical findings are summarized in Table 2. Serum biochemical panel abnormalities included marked elevations in activities of hepatocytic enzymes (AST, ALT and LDH) and a decreased glucose concentration. After the onset of this disorder, Monkey A did not show changes in serum electrolytes such as Ca, IP, Mg, Na, K and Cl, or lipemia associated with the elevated TG or T-Cho.
| Para- meters | Units | Hyperadrenocorticism | Reference values (n = 75) in NINS | ||
|---|---|---|---|---|---|
| Before | After | ||||
| T4 | nmol/l | N. D. | 72.1 | 81-160 | |
| Free T4 | pmol/l | N. D. | 12.9 | 12-22 | |
| T3 | nmol/l | N. D. | 1.63 | 1.54-2.60 | |
| Free T3 | pmol/l | N. D. | 0.04 | 0.03-0.06 | |
| Estradiol | pmol/l | N. D. | 77 | 92-734 | |
| Progesterone | nmol/l | N. D. | 1.3 | 0.6-4.8 | |
| ACTH | pmol/l | N. D. | 30 | < 8.8 | |
| Cortisol | nmol/l | N. D. | 803 | 170-550 | |
| Catecholamine | |||||
| . . . Adrenalin | pmol/l | N. D. | < 16 | < 55 | |
| . . . Noradorenalin | pmol/l | N. D. | 5 | 4-18 | |
| . . . Dopamine | pmol/l | N. D. | 5 | < 6.5 | |
| DHEA-S | μmol/l | N. D. | 1.3 | 2.7-13.5 | |
Table 3: Endocrinological findings. N. D.: Not done.
Endocrinological findings: Endocrinological parameters are summarized in Table 3. There were no abnormalitiesin thyroid hormones, catecholamine, or DHEA-S levels. Although estradiol and progesterone concentrations were mildly suppressed below the normal ranges (estradiol: 92-734 pmol/l, progesterone: 0.6-4.8 nmol/l), ACTH and cortisol were markedly elevated in serum levels (ACTH: high > 8.8 pmol/l, cortisol: high > 550 nmol/dl).
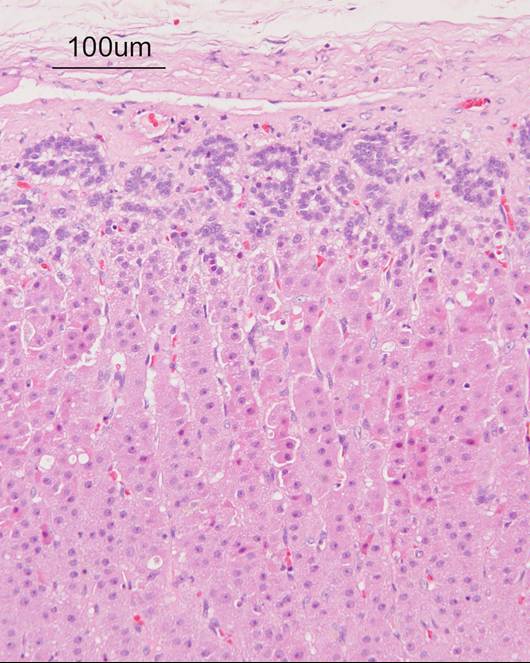
Figure 3: Systemic Microscopic photograph of the adrenal cortex. No marked hyperplasia or hypertrophy is noted in the zona fasciculata. HE stain.
Histopathological findngs: There was no marked hyperplasia or hypertrophy in the zona fasciculata of the adrenal cortex (Figure 3).
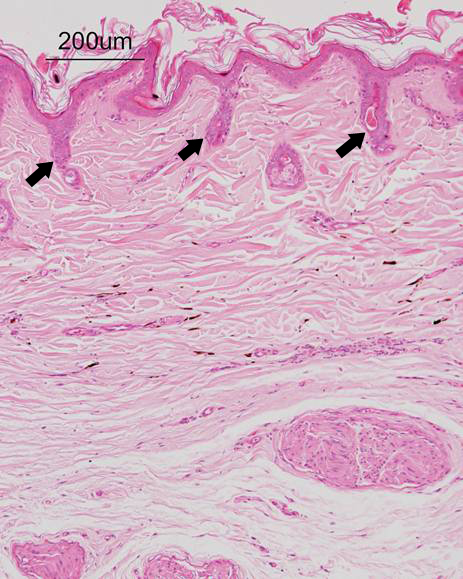
Figure 4: Microscopic photograph of the skin. Thin epidermis and hair follicle dysplasia (arrows) are seen. Hair growth cycle arrest in telogen phase is noted. No infiltration of lymphocytes or mononuclear cells is observed around and within the bulbs and root sheaths. HE stain.
Histopathological findings of the skin specimens included thin epidermis and hair follicle dysplasia with sebaceous glands (Figure 4). The epidermis consisted of 3 to 4 component cells, unaccompanied by hyperkeratosis. The hair follicles resided in telogen stages and no infiltration of lymphocytes or mononuclear cells was observed around and within the bulbs and root sheaths. Hair growth cycle arrest in telogen phase was accompanied by variable follicular atrophy and these hair follicles were uniformly absent from hair shafts.
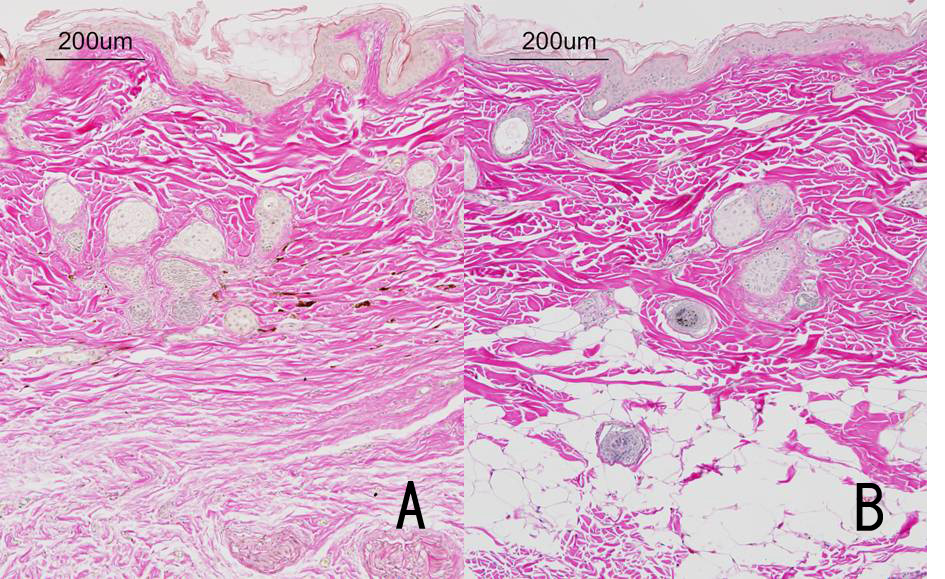
Figure 5: Microscopic photograph of the skin in Japanese monkeys. These are samples, with van Giesonĺs staining.
A: Alopecic skin in hyperadrenocorticism. Fatty tissues disappear and thin collagen fibers are seen in the deeper layer of the dermis.
B: Normal skin.
An arrangement of thin collagen fibers was found in the deeper layer of the dermis. The bundles of collagen fibers in the deep dermis were thinner than those in the superficial dermis. Fatty tissues disappeared in the entire dermis and in the connective tissues (Figure 5).
Discussion
Clinical signs and skin features in the monkey were closely consistent with those observed in dogs suffering from hyperadrenocorticism (Peterson, 2007; Scott et al., 1995, pp. 645-660). Dogs with pituitary-dependent hyperadrenocorticism resulting from pituitary adenomas or loss of feedback control tend to have signs develop grad-ually, whereas the rate of occurrence can be more rapid and unpredictable with pituitary adenocarcinoma or adrenal tumors (Scott et al., 1995, pp. 645-660). The course of this disease in Monkey A resembled that of pituitary-dependent hyperadrenocorticism. An accumulation of fluid, found within the peritoneal cavity, may have resulted from elevation of the hydrostatic pressure with hepatic congestion.
The hemogram in canine hyperadrenocorticism includes mature leukocytosis, eosinopenia, and lymphopenia. 80% of cases have eosinopenia and lymphopenia, whereas leukocytosis is seen in only 24% of cases (Ling et al, 1979). My hematological results in Monkey A accorded with a ôstress leukogramö found in dogs with hyperadrenocorticism.
In dogs affected with hyperadrenocorticism, serum biochemical examinations commonly showed high activities in AST, ALT, and ALP (high in 85 to 90% of the dogs), which was mainly due to a corticoid-induced isoenzyme. Additionally, such electrolytes as sodium, potassium, and chloride are usually within normal limits (Peterson, 2007; Scott et al., 1995, pp. 645-660). Our results agreed with those described in canine hyperadrenocorticism. Although Beardi et al. (2007) investigated alopecia areata in a rhesus monkey (Macaca mulatta), hematological and serological examinations did not show abnormal results, as all tested parameters were within the reference values. Their results revealed that clinicopathological findings in alopecia due to endocrinopathy were apparently different from those in the other types of alopecia. The terminal stage of this disorder seems to have some effects on lipid metabolism. The decreased TG, T-Cho, and Glu values may be a serum biochemical profile which is observed in the end stage of hyperadrenocorticism.
Increases in basal ACTH and cortisol concentrations in Monkey A agreed with the measurements in dogs suffering from hyperadrenocorticism (Peterson, 2007; Scott et al., 1995, pp. 645-660). Steinmetz et al. (2005) investigated coat conditions of captive rhesus macaques under different housing conditions. They reported that levels of immunoreactive cortisol metabolites (11-oxoetiocholanolone) in feces were negatively correlated with alopecia, suggesting a relationship between hypothalamic-pituitary-adrenal (HPA) axis activity and hair loss in captive rhesus macaques. The elevated ACTH and cortisol levels revealed that the pathogenesis of systemic alopecia in Monkey A differed from that of the aforementioned alopecia in captive rhesus macaques.
The histopathological findings of Monkey A were in close agreement with those described in dogs affected with hyperadrenocorticism (Scott et al., 1995, pp. 645-660; Gross et al., 2005). Representative baldness in human beings, alopecia areata, is characterized by complete or nearly complete absence of hair in one or more circumscribed areas of the scalp. The histopathological picture of alopecia areata is characterized by the infiltration of T-lymphocytes and macrophages affecting anagen hair follicles, subsequently leading to hair follicle miniaturization and defective hair shaft formation with clinically visible hair loss. Alopecia areata universalis is defined as total scalp hair alopecia accompanied by loss of body hair including eyebrows and eyelashes. Alopecia areata, including its most severe forms of alopecia areata totalis and universalis, is non-scarring inflammatory alopecia (Abel, 1997). Our histopathological findings were different from those reported in alopecia areata in human beings.
Our findings revealed that this systemic alopecia was caused by hormonal imbalances of the HPA axis itself. Our results showed that hyperadrenocorticism in Monkey A resembled pituitary-dependent hyperadrenocorticism observed generally in dogs and human beings. Unfortunately, a histopathological examination was not made of the pituitary gland of this monkey.
References
Abel, E. (1997). Alopecia areata. In: D. Elder, R. Elenitsas, C. Jaworsky, & B. Johnson, Jr. (Eds.), Leverĺs histopathology of the skin (8th ed., pp. 409-411). Philadelphia: Lippincott-Raven.
Ahmad, W., Ratterree, M. S., Panteleyev, A. A., Aita, V. M., Sundburg, J. P., & Christiano, A. M. (2002). Atrichia with papular lesions resulting from mutations in the rhesus macaque (Macaca mulatta) hairless gene. Laboratory Animals, 36, 61-67.
Beardi, B., Wanert, F., Zoller, M., Freyschmidt-Paul, P., Bodemer, W., & Kaup, F. J. (2007). Alopecia areata in a rhesus monkey (Macaca mulatta). Journal of Medical Primatology, 36, 124-130.
Chiaramonte, D., & Greco, D. S. (2007). Feline adrenal disorders. Clinical Techniques in Small Animal Practice, 22, 26-31.
Gross, T. L., Ihrke, P. J., Walder, E. J., & Affolter, V. K. (2005). Canine hyperglucocorticoidism. In: Skin diseases of the dog and cat: Clinical and histopathologic diagnosis (2nd ed., pp. 484-487). Oxford: Blackwell.
Ling, G. V., Stabenfeldt, G. H., Comer, K. M., Gribble, D. H., & Schechter, R. D. (1979). Canine hyperadrenocorticism: Pretreatment clinical and laboratory evaluation of 117 cases. Journal of American Veterinary Medical Association, 174, 1211-1215.
Peterson, M. E. (2007). Diagnosis of hyperadrenocorticism in dogs. Clinical Techniques in Small Animal Practice, 22, 2-11.
Reinhardt, V. (2005). Hair pulling: A review. Laboratory Animals, 39, 361-369.
Scott, D. W., Miller, Jr., W. H., & Griffin, C. E. (1995). Muller & Kirkĺs Small animal dermatology (5th ed.). Philadelphia: Saunders.
Steinmetz, H. W., Kaufmanns, W., Dix, I., Heistermann, M., & Fox, M. (2006). Coat condition, housing condition and measurement of faecal cortisol metabolites ľ a non-invasive study about alopecia in captive rhesus macaques (Macaca mulatta). Journal of Medical Primatology, 35, 3-11.
Steinmetz, H. W., Kaufmanns, W., Dix, I., Neimeier, K. A., & Kaup, F. J. (2005). Dermatologic investigation of alopecia in rhesus macaques (Macaca mulatta). Journal of Zoo and Wildlife Medicine, 36, 229-238.
Wilkinson, A. C., Harris, L. D., Saviolakis, G. A., & Martin, D. G. (1999). Cushingĺs syndrome with concurrent diabetes mellitus in a rhesus monkey. Contemporary Topics in Laboratory Animal Science, 38, 62-66.
-------------------------------------------
Corresponding address: Tohru Kimura, Center for Experimental Animals, National Institutes of Natural Sciences, 38, Nishigo-naka, Myodaiji, Okazaki, Aichi, 444-8585, Japan [e-mail: [email protected]].
-------------------------------------------
* * *
Volunteers Wanted: Monkey Town Primate Centre
Volunteers are needed at Monkey Town Primate Centre, near Cape Town, South Africa. They have no employed primate keepers, and only three volunteers to look after over 200 primates. They desperately need more volunteers (with experience would be great) to help run this park. For more information e-mail < [email protected]>.
* * *
Effect of Environmental Enrichment on Behavioral and Endocrine Aspects of a Captive Orangutan (Pongo pygmaeus)
Cristiane Schilbach Pizzutto1; MarcÝlio Nichi1; Manuela Gonšalvez Fraga Geronymo Sgai1; Sandra Helena Ramiro Corrŕa2; Priscila Viau1; Ana Maria Beresca2; Clßudio Alvarenga de Oliveira1; Renato Campanarut BarnabÚ1; and Marcelo Alcindo de Barros Vaz GuimarŃes1
1 Universidade de SŃo Paulo, Brasil; 2FundašŃo Parque Zoolˇgico de SŃo Paulo, Brasil
Introduction
Nonhuman primates show structural and functional similarities with humans. Many behavioral standards and emotional reactions exhibit common evolutionary and ontogenetic bases. Some behavioral and social complexities in primates are not found in other species.
For the last 20 years, studies of the behavior of captive primates have shown a steep increase (Hosey, 2005), most of these studies aiming to understand how the environment in captivity influences behavior (Hosey & Druck, 1987; Lindburg & Fitch-Snyder, 1994).
Nonhuman primates are vulnerable to the emotional effects of stress (Boere, 2002). The physiological and behavioral impact of a stressing agent is highly dependent on the perception and the response of each individual. Many acute stressing agents may cause a general increase in excitement, which, in turn, may bring physiological and psychological benefits to the animal (Natelson et al., 1987). Environments with increased social support and positive stimuli are more adequate to behavioral needs. Enriched environments in captivity are known to counter the isolation and lack of stimuli which are responsible for behavioral disorders (Herskin & Jensen, 2000). Captive orangutans may show obesity, decreased grooming, skin and coat disorders, and a tendency to remain on the floor. Opportunity for vertical movement and brachiating in high places is an important form of enrichment (Hebert & Bard, 2000). Hebert & Bard noted that enriched captive orangutans spend 62% of the time at the tops of trees, showing a preference for higher spots, where they eat their food and partially hide from the public.
Recent studies have correlated behavioral and hormonal measurements, providing new findings about different species, as well as the costs and benefits of behavioral strategies and resulting endocrine variations. This may allow a better understanding of the evolution of social behaviors.
Studying the role of each hormone requires attention to the social and ecological context of the endocrine function, as well as the relationship between behavioral variations and individual history. Studies indicate important differences in absolute values of some hormones when comparing captive and free-ranging animals (Wingfield & Moore, 1987). The reasons for those differences are being investigated using ethological analysis associated with the evaluation of hormonal levels before and after social and environmental stimulation (Ketterson & Nolan, 1992).
Here we study behavioral and hormonal changes in a single female orangutan, before and after the introduction of environmental enrichment.
Methods
From 2003 to 2004 we studied one captive adult female orangutan (Pongo pygmaeus), named Karen, kept for 25 years with no environmental enrichment at the FundašŃo Parque Zoolˇgico de SŃo Paulo.

Figure 1: The environment before structural modifications.
The study was performed in two phases: Baseline (evaluation under captive conditions, with no environmental enrichment ľ 3 months), and Enrichment (evaluation after structural modifications to the environment and the provision of enrichment ľ 3 months).
Two categories of variables were evaluated:
1) Behavioral variables: Behavior was recorded using focal sampling by interval (Altmann, 1974; Martin & Bateson, 1993), with data taken each 30 seconds in sessions of 30 minutes, totaling 40 hours, with 4,800 recordings before and after enrichment. Observations were performed in the morning, three times a week.
Based on the behavioral data, a repertoire of 8 behaviors was recorded (Table 1).
| Category | Behavior |
| Locomotion | Walking or running. |
| Stationary | Standing still, lying down, or sleeping. |
| Feeding | Manipulating and/or ingesting food. |
| Self-gooming | Cleaning the hair free of parasites, combing and cleaning the hair with fingers or mouth. |
| Interaction | Reacting to the audience in the exhibition area. |
| Manipulation of objects | Handling objects (e. g., plastic, paper) or any item introduced into the new enviroment (ropes, wood, plants, and stimulating items). |
| Scratching | Scratching parts of the body using hands or feet. |
| Others | Vocalizing, drinking water, urinating, and defecating. |
Table 1: Behavior categories of a captive orangutan.
2) Endocrine variables: Fecal metabolites of cortisol and estradiol were measured in samples collected at the beginning of the day. Metabolites were extracted according a technique described by Whitten et al. (1998) for feces of chimpanzees (Pan troglodytes).
To evaluate the results of hormonal measurements, three different response phases were considered: * Baseline: from the beginning of observations to the day before implementing enrichment techniques (3 months). * Habituation: starting immediately after implementation of enrichment techniques, with hormonal levels showing peaks (values two standard deviations higher then the mean value for the baseline phase) for at least two consecutive observations, until hormonal levels returned to basal levels. * Enrichment: Three months, from the end of the Habituation phase.
For the statistical analysis the Habituation phase was not considered.
Statistical analysis: All data were statistically evaluated using SAS System for Windows (SAS Institute Inc., Cary, NC, USA, 2000). Differences between phases were analyzed using parametric (Student t test) and non-parametric (Wilcoxon) tests, according to the residue normality (Gaussian distribution) and variance homogeneity. Whenever necessary, data were transformed in order to obey these statistical assumptions. A probability value of p < 0.05 was considered statistically significant. Results were described as untransformed means ▒ standard deviation. Dichotomic variables were analyzed using PROC GLIMMIX (logistic regression). Pearson and Spearman correlations were used to calculate the relationship between variables.
Results
Structural modifications performed to enrich the environment in which Karen was housed may be viewed in Figures 1, 2 and 4.

Figure 2: The environment after structural modifications.
A behavioral profile of Karen, as well as the place in which she was observed performing each behavior, before and after enrichment, is listed in Tables 2 and 3.
| Behavioral Category | Before | After | p |
| Locomotion | 10.12 | 7.78 | <0.0001 |
| Stationary | 61.75 | 25.56 | <0.0001 |
| Feeding | 12.87 | 10.84 | 0.0020 |
| Self-grooming | 3.96 | 0 | <0.0001 |
| Interaction | 0.50 | 0 | <0.0001 |
| Object manipulation | 6.82 | 54.75 | <0.0001 |
| Scratching | 3.61 | 0.95 | <0.0001 |
| Others | 0.37 | 0.12 | <0.0001 |
Table 2: Mean frequencies (%) of behavioral variables of a captive orangutan, before and after environmental enrichment.
The coefficients of variability intra-assay (CV intra), inter-assay (CV inter) and minimum detected sensibility from fecal metabolites of cortisol were: CV intra = 12.75%, CV inter = 2.21%, and sensibility = 2.55 ng/dl; and from fecal metabolites of estradiol: CV intra = 7.48%, CV inter = 0.67%, and sensibility = 1.44 pg/ml.
Results obtained for the assay validation using parallel analysis for the metabolites of cortisol and estradiol were r2 = 0.987, p<0.05, and r2 = 0.999, p<0.05, respectively.
| Location | Before (%) | After (%) | p |
| Floor | 95.29 | 2.36 | <0.0001 |
| Platform | 4.1 | 0 | <0.0001 |
| Net | 0 | 0 . | |
| Tree | 0 | 97.64 | <0.0001 |
Table 3: Mean frequencies (%) of the locations in which the captive orangutan was observed before and after enrichment.
Karen showed significant alterations in her hormonal profile before, during, and after the environmental enrichment, as shown in Figure 3.
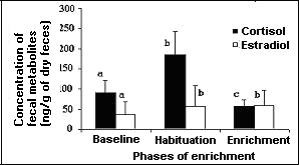
Figure. 3: Levels (mean ▒ standard deviation) of Karenĺs fecal cortisol metabolites (ng/g times 103) and estradiol (ng/g) in different phases of environmental enrichment. a, b, c: Different letters indicate statistical differences (p < 0.05).
A significant rise in fecal metabolites of cortisol was observed when Karen was introduced to the restructured and enriched environment. This level remained for 15 days, which was considered the Habituation phase. After this phase, there was a decrease in cortisol, with the levels oscillating below the Baseline phase values (Figure 3).
A statistically significant difference was found between fecal metabolites of estradiol in the Baseline period and in the Enrichment phase (Figure 3). It was also possible to verify hormonal fluctuations related to ovarian cyclicity, with evident peaks of fecal metabolites of estradiol in both phases.
Discussion
The restructuring of the environment in which Karen was housed induced changes in her behavioral profile, such as a reduction in use of the floor (from 95.9% during the Baseline phase, to 2.6% during the Enrichment phase) and an increase in time spent in higher places (Baseline = 0; Enrichment = 97.64%). These results agree with previous works in which authors described captive orangutans in an enriched environment spending a limited amount of time on the floor (Hebert & Bard, 2000), spending most of their time (62% of the observations) at the top of trees, where they ate all the food provided by the zoo and partially hid from the public.
Karen demonstrated considerable interest in stationary (trees), mobile (ropes to access the tops of the trees), and removable (boxes, plastic bottles) enrichment items. These results do not agree with those of Wilson (1972), who found that orangutans preferred stationary and mobile objects over removable objects that, according to him, do not play a role in their behavior.

Figure 4: The orangutan in her enriched environment.
One objective of the present study was to evaluate which enrichment items would be better for the species. We observed that there was a significant increase in object manipulation (6.82% to 54.75%), as observed in previous studies (e.g., Perkins, 1992), in which provision of manipulable objects and grazing items increased the occurrence of typical activities and reduced inactivity and abnormal behaviors.
A significant increase in activities involving manipulation is directly related to a marked decrease in inactivity, as seen on the ôstationaryö behavior category (Table 2). This agrees with several reports suggesting that, for primates, the quality of an enclosure is not determined only by its size. For these animals, the presence, arrangement, and organization of objects appear to play an important role (Wilson, 1982; Erwin & Sackett, 1990; Boere, 2001).
Our results demonstrate that, after enrichment, Karen was observed at the top of the trees 97.64% of the time, a value similar to that found by Gippoliti (2000) in free-ranging orangutans, who remained day and night up to 10 meters above the ground. This indicates that providing ropes and a high platform allowed the appearance of arboreal behavior, which is typical in orangutans (e.g., Perkins, 1992; Hebert & Bard, 2000).
According to Rodman & Mitani (1987), wild orangutans spend around 45% of their time eating. The same authors state that, in captivity, these animals use only 18% of their time eating, which is similar to the values found in the Baseline of the present study (10.84%).
When animals are submitted to new situations, such as environmental modifications, there is a rise in secretion of glucocorticoids and/or catecholamines (Moberg, 2000). Confronting the new environment, as well as the perception of danger or threat, leads to an immediate stimulation of the hypothalamic-pituitary-adrenal axis, which will result in a rise in blood levels of cortisol (e.g., Hennessy & Levine, 1979; Carlstead et al., 1992). Therefore, it was expected that the introduction of our animal to an enriched environment would induce significant increases in fecal metabolites of cortisol, as we found with Karen.
Karenĺs adaptation to her new environment, evidenced by the significant increase in fecal metabolites of cortisol, lasted 15 days. This period was followed by a significant reduction of cortisol metabolites below the mean levels found during the Baseline phase. These results indicate that, after the Habituation phase, the stimuli induced by the innovations no longer resulted in a stress response, suggesting an adjustment of the animal to the new environment. It is important to note that values found for the metabolites of cortisol after the Habituation phase were significantly lower than those found during Baseline, indicating an improvement in Karenĺs reaction to her environment.
To our knowledge, this is the first report on the values of fecal metabolites of cortisol and estradiol for a female orangutan (Pongo pygmaeus).
Conclusion
Our results suggest that the introduction of environmental enrichment may play an important role in modulating behavioral and endocrine aspects of captive orangutans, which may help these animals to adapt to captivity.
References
Altmann, J. (1974). Observational study of behavior: Sampling methods. Behaviour, 49, 227-267.
Boere, V. (2001). Behavior and environment enrichment. In: M. E. Fowler & Z. S. Cubas (Eds.), Biology, medicine, and surgery of South American wild animals (pp. 263-266). Ames, IO: Iowa State University Press.
Boere, V. (2002). Efeitos do estresse psicossocial cr˘nico e do enriquecimento ambiental em sagüis (Callithrix penicillata): Um estudo comportamental, fisiolˇgico e farmacolˇgico. (dissertation). SŃo Paulo, Instituto de Psicologia, Universidade de SŃo Paulo.
Carlstead, K, & Brown, J. L. (2005). Relationship between patterns of fecal corticoid excretion and behavior, reproduction, and environmental factors in captive black (Diceros bicornis) and white (Ceratotherium simum) rhinoceros. Zoo Biology, 24, 215-232. Carlstead, K., Brown, J. L., Monfort, S. L., Killens, R., & Wildt, D. E. (1992). Validation of a urinary cortisol radioimmunoassay for non-invasive monitoring of adrenal activity in domestic and nondomestic felids. Zoo Biology, 11, 165-176.
Erwin, J., & Sackett, G. P. (1990). Effects of management methods, social organization and physical space on primate behavior and health. American Journal of Primatology, 20, 23-30.
Gippoliti, S. (2000). Orang-utans in zoos: Husbandry, welfare and management in a typical arboreal solitary mammal. International Zoo News, 47, 356-368.
Gould, E., & Bres, M. (1986). Regurgitation and reingestion in captive gorilla: description and intervention. Zoo Biology, 5, 241-250. Hebert, P. L., & Bard, K. (2000). Orangutan use of vertical space in an innovative habitat. Zoo Biology, 19, 239-251.
Hennessy, J. W., Heybach, J. P., Vernikos, J., & Levine S. 1979. Plasma corticosterona concentration sensitively reflect levels of stimulus intensity in the rat. Physiology & Behavior, 22, 821-825. Hennessy, J. W., & Levine, S. (1979). Stress, arousal, and the pituitary-adrenal system: A psychoendocrine hypothesis. Progress in Psychobiology and Physiological Psychology, 8, 133-178.
Herskin, M. S., & Jensen, K. H. (2000). Effects of different degrees of social isolation on the behavior of weaned piglets kept for experimental purposes. Animal Welfare, 9, 237-249.
Hosey, G. R. (2005). How does the zoo environment affect the behaviour of captive primates? Applied Animal Behaviour Science, 90, 107-129.
Hosey, G. R., & Druck, P. L. (1987). The influence of zoo visitors on the behaviour of captive primates. Applied Animal Behaviour Science, 18, 19-29.
Ketterson, E. D., & Nolan, J. R. V. (1992). Hormones and life histories: An integrative approach. American Naturalist, 140, 33-62.
Lindburgh, D. G., & Fitch-Snyder, H. (1994). Use of behavior to evaluate reproductive problems in captive mammals., Zoo Biology, 13, 433-445.
Maple, T. L., & Finlay, T. W. (1986). Evaluating the environments of captive non-human primates. In: K. Benirschke (Ed). Primates: The road to self-sustaining populations (pp. 479-488). New York: Springer Verlag. Martin, P., & Bateson, P. (1993). Measuring behaviour: An introductory guide. Cambridge: Cambridge University Press.
Mason, J. W. (1968). A review of psychoendocrine research on the pituitary-adrenal cortical system. Psychosomatic Medicine, 30, 576-607. Moberg, G. P. (2000). Biological responses to stress: Implications for animal welfare. In: G. P. Moberg & J. A. Mench (Eds.), Biology of animal stress: Basic principles and implications for animal welfare (pp. 1-22). London: CABI Publishing.
Natelson, B. H., Creighton, D., McCarty, R., Tapp, W. N., Pitman, D., & Ottenweiler, J. E. (1987). Adrenal hormonal indices of stress in laboratory rats. Physiology and Behavior, 39, 117-125.
Perkins, L. A. (1992). Variables that influence the activity of captive orangutans. Zoo Biology, 11, 177-186.
Rodman, P. S. (1984). Foraging and social systems of orangutans and chimpanzees. In: P.S. Rodman & J. G. H. Cant (Eds.). Adaptations for foraging in nonhuman primates (pp. 134-160). New York: Columbia University Press. Rodman, P. S., & Mitani, J. C. (1987). Orangutans: Sexual dimorphism in a solitary species. In: B. B. Smuts, D. L. Cheney, R. M. Seyfarth, R. W. Wrangham, & T. T. Struhsaker (Eds.), Primate societies (pp. 146-154). Chicago: Chicago University Press.
Thorpe, S. K. S., & Crompton, R. H. (2005). Locomotor ecology of wild orangutans (Pongo pygmaeus abelii) in the Gunung Leuser Ecosystem, Sumatra, Indonesia: A multivariate analysis using log-linear modeling. American Journal of Physical Anthropology, 127, 58-78.
Whitten, P. L., Stavisky, R., Aureli, F., & Russel, E. (1998). Response of fecal cortisol to stress in captive chimpanzees (Pan troglodytes). American Journal of Primatology, 44, 57-69.
Wilson, C. C. (1972). Spatial factors and the behavior of nonhuman primates. Folia Primatologica, 18, 256-275.
Wilson, S. F. (1982). Environmental influences on the activity of captive apes. Zoo Biology, 1, 201-209.
Wingfield, J. C., & Moore, M. C. (1987). Hormonal, social, and environmental factors in the reproductive biology of free living male birds. In: D. Crews (Ed.), Psychobiology of reproductive behavior: An evolutionary perspective (pp. 149-175). Englewood Cliffs, NJ: Prentice-Hall.
-------------------------------------------
First authorĺs address: Depto de ReprodušŃo Animal, Fac. de Med. Vet. e Zootec., Univ. de SŃo Paulo, Brasil [e-mail: [email protected]].
We thank the FundašŃo Parque Zoolˇgico de SŃo Paulo for the opportunity of working with the orangutan in their collection.
We also thank the Conselho Nacional de Desenvolvimento CientÝfico e Tecnolˇgico (CNPq ľ141626/2002-0) for C. S. P.ĺs PhD scholarship, and FundašŃo de Amparo Ó Pesquisa do Estado de SŃo Paulo (FAPESP ľ 02/13966-6) for financially supporting this experiment.
This work was performed according to bioethics protocols of the Faculty of Veterinary Medicine and Zootechnology of the University of SŃo Paulo and Brazilian animal care regulations.
-------------------------------------------
* * *
Communication Between Caregivers and Primates: A Survey
Joanne Davis
University of South Australia
I am a third-year PhD student, doing a survey as part of my overall study. My PhD is about control and choice in captive primates, and I am interested in how communication between caregivers and primates affects the amount of environmental control they can get. I am also keen to learn how personality affects issues such as choice and control in the great apes. With this in mind I hope to better our understanding of welfare needs of primates in captivity, whether in zoos, sanctuaries, or laboratories.
The survey is available online, and takes about 30 minutes to complete. If that is not convenient for you, I can e-mail a copy as a Word document or snail mail a copy, upon request to my address (below).
The criteria for participation are that you are over 18 years of age and care for any species of primate. The survey can be anonymous or you may leave your name and address to request the results at the conclusion of the study.
One important component is the personality rating scales, which are included for the caretakers of orangutans, gorillas and chimps. You can request that the personality type of that subject be supplied to you in addition to the results. I particularly urge caretakers to ask their colleagues also to complete the survey for their apes if they complete one, as the personality rating scales are more accurate when there are multiple caretakers completing them for each ape. For example, it is better for four keepers to complete one survey each on one individual ape than to have four different surveys on different individuals. For other primate species this is not important as there are no personality rating scales used.
Your participation is both crucial for the study to succeed and much appreciated, and I thank you in advance for taking the time to complete the survey. It is not until we understand what our primates need, that we can give it to them!
The Web address is <www.surveymonkey.com/s.aspx?sm=WyVGJo4iVcfDYmG4B77BNA_3d_3d>. To obtain Word versions of the survey please go to <www.aarcg.org/Current%20projects.html> and download the Word document that corresponds to the species you work with (for bonobos please use the chimp document). For more details on my work, please go to <www.aarcg.org>, the site of the Applied Animal Research and Conservation Group, which sponsors this project.
-------------------------------------------
Authorĺs address: Rm C1-19, School of Psychology, University of South Australia, St Bernards Rd, Magill 5072, Australia [+618-8302 4161; fax: +618-8302 4729; e-mail: [email protected]].
-------------------------------------------
* * *
Announcements from Publications: Evolution: Education and Outreach
Springer has launched a new journal, Evolution: Education and Outreach, devoted to promoting accurate understanding and comprehensive teaching of evolutionary theory for a wide audience. Targeting K-16 students, teachers, and scientists alike, the journal publishes articles to aid members of these communities in the teaching of evolutionary theory.
ôWe actively seek contributions of (1) peer-reviewed evolutionary science articles; (2) peer-reviewed educational papers on curricula and experiences in the classroom; and (3) Ĺreportsĺ and Ĺreflectionsĺ style pieces from qualified professionals.ö
Springer will be offering four $2,500 awards: two for Best Original Paper; and two for Best Classroom Application. The Best Original Papers will be papers with topics within evolutionary biology, papers on the nature of science, and all other original research, including educational and cognitive research. The Best Classroom Applications will be lesson plans, curricula, and activities (including Powerpoint and other Internet applications) that have a direct impact on evolution education. For more information see <www.springerlink.com/content/1936-6434>. ľ From the publisherĺs announcement
* * *
Resources Wanted and Available
Noldus Catalog of Software/Hardware Solutions
Noldus Information Technology has published its first catalog of solutions for animal behavior research. This 118-page full-color book presents an overview of software and hardware products and integrated solutions, as well as examples of applications for which they can be put to use. The catalog describes solutions for research on small lab animals, wildlife and zoo animals, farm animals, fish, insects, and companion animals. To receive a free copy, fill out the form at <www.noldus.com/site/doc200407016>.
New Tree Analysis Program Available Free
TNT stands for ôTree Analysis Using New Technologyö. It is a program for phylogenetic analysis under parsimony (with very fast tree-searching algorithms; Nixon, 1999, Cladistics, 15, 407-406; Goloboff, 1999, Cladistics, 15, 407-428), as well as extensive tree-handling and diagnosis capabilities. It is a joint project by Pablo Goloboff, James Farris, and Kevin Nixon. As of November, 2007, the project was subsidized by the Willi Hennig Society, and thus the program is now being made freely available, upon agreement on the terms of the License, which is a single-user license. The most important requirements are that the program not be posted in other Websites or mirrors, and that when publishing results you acknowledge having used the Willi Hennig Society edition of TNT and cite the paper describing the program (Goloboff et. al, 2008, in press in Cladistics). For complete information, and to download TNT, see
Galagos Wanted
The Duke University Primate/Lemur Center (DLC), Durham, North Carolina,áis interested in establishing a breeding population of Galago moholi and/or Galago senegalensis. Please contact Dean Gibson, DLC Assistant Director, 3705 Erwin Rd, Durham, NCá 27705-5000 [919-489-3364, X224; e-mail: [email protected]], if your institution has these species available.
A training resource on facility inspections is a course developed by Martha Rooks, PhD, on the AALAS Learning Library: <www.aalaslearninglibrary.org>. This 10-minute course, ôThe Semi-Annual Facility Inspectionö, offers training through an interactive scenario of a typical inspection, in which compliance issues are identified in video vignettes and then addressed later in the course. The course can be accessed under the Animal Care and Use link / Regulatory and IACUC Compliance link / Working with the IACUC link.
Great Ape Distribution Maps
International Education and Resource Network (iEARN) of Australia has created distribution maps of the great apes. Here are the URLs:
EUPRIM-Net Primate Material Bank
EUPRIM-Net (European Primate Network) is a European Union-funded project networking eight European primate centers. It is aimed at advancing knowledge and competence in biological and biomedical research, and in animal keeping and breeding. The project links the centers into a network and facilitates a mutual use of existing know-how and resources. The project objectives focus on animal welfare, standardization of procedures and methods, enhanced availability of nonhuman primates, and training for those working with primates. For more information, see <www.euprim-net.eu>.
EUPRIM-Net has a primate material bank containing tissue, serum, blood, gene, and gamete samples (see <www.euprim-net.eu/biobank>). You can apply for primate material if you are an investigator at a university, research institute, or company located in a Member State of the European Union or in an Associated State. The costs for samples delivered by EUPRIM-Net are supported by the EU. According to EU regulations, a short description of your research plan is needed before samples can be delivered free of charge. Please be aware that the number of free samples is restricted. If your institution is not located in an EU Member State or an Associated State, contact < [email protected]> for fee-based orders.
* * *
After a long battle with cancer, Stella Brewer has died. Stella and her father Eddie Brewer, then Chief of Wildlife of The Gambia, founded the first-ever chimpanzee rehabilitation program in 1969. The project maintains a Website at <www.chimprehab.com>. Donations may be made in Stellaĺs honor through this Website.
In 1978 Stella wrote a book which was named ôThe Forest Dwellersö on one side of the Atlantic and ôThe Chimps of Mount Asserikö on the other. Used copies are available from <www.amazon.com>.
Stella leaves behind her husband David Marsden, two sons, and her sister Heather, also an animal lover. ľ posted to primfocus by Shirley McGreal, January 31
Appointments at OLAW ľ Taylor and Hampton
The Office of Laboratory Animal Welfare, National Institutes of Health (OLAW), is pleased to announce the appointment of Dr. Kim Taylor to the position of Senior Assurance Officer, Division of Assurances. Dr. Taylor comes to OLAW with over 13 yearsĺ experience in biomedical research involving animals.á Most recently Dr. Taylor served as the Deputy Animal Program Director for the National Institutes of Allergy and Infectious Diseases, Comparative Medicine Branch.á Prior to joining NIAID, Dr. Taylor spent 12 years in the U.S. Army where she completed a residency program in Laboratory Animal Medicine at the U.S. Army Research Institute of Infectious Diseases located in Frederick, Maryland.
Ms. Lori Hampton has joined OLAW in the Division of Policy and Education as an Animal Welfare Program Specialist. Ms. Hampton began her career at NIH in 1983. Since 1995, she has worked at the National Institute on Deafness and Other Communication Disorders, where she was a biologist studying the genetics of deafness and the function of G protein-coupled receptors and served on the NINDS/NIDCD Animal Care and Use Committee. Loriĺs twenty-four years of scientific research experience at the NIH include twenty years of working with animal models of human disease and managing laboratory operations.
OBE for Shirley McGreal
ôThe British New Yearĺs Honours list was released today. People interested in the welfare of primates will be pleased to know that Dr. Shirley McGreal has received the Order of the British Empire for her work on primate issues. She will have to learn to curtsey at Buckingham Palace!
ôMany animals have had their lives helped by Shirleyĺs work on international smuggling and trade issues. Getting a primate export ban in India was a highlight of Shirleyĺs work and enabled many animals to remain exactly where they should be ľ living wild and free and not in laboratories. Shirley has a special love of gibbons. Her gibbon facility is wonderfully maintained and well staffed. Many overseas groups struggling with rescue facilities have been helped by IPPL for many years. Prince Philip has been interested in IPPL and has supported Shirley in her work, especially with international trading issues.
ôCongratulations, Shirley! Also congratulations to the many people who have helped Shirley over the years.ö ľ posted to primfocus on December 28 by Jean Martin, of Lantzville, British Columbia
Railway to Put Gibbons Back on Track
A tribe of gibbons that was split into two by a rail track passing through their habitat is to be reunited, after several decades of separation, by two steel bridges designed to look like trees. Northeast Frontier Railway has decided to build the two bridges over an 800-meter stretch of the Gibbon Wildlife Sanctuary in Jorhat district of Assam, India, to compensate for the forced separation of the two groups. Gibbons are an exclusively arboreal species and do not set foot on the ground. When laying the rail track in the thirties, nobody in the railways or the forest department realized that the gibbon population would be bifurcated along with the stretches of forest on either side. The two steel bridges will be camouflaged to look like trees with branches and encourage the gibbon population on either side of the track to mingle.
The sanctuary, located near Mariani along the Assam-Nagaland border, has 21 gibbon families with an average of four members in each. Gibbons are an endangered species and listed in Schedule I of the Indian Wildlife Protection Act, 1972. Apart from the northeastern states, they are found in Bangladesh, Myanmar, and the southern parts of China.
It is not known how many gibbons the sanctuary had lost because of the track, since conservation efforts gained momentum only after the forest was declared a sanctuary in 1979. The sanctuary was previously known as the Hollongapar Reserve Forest. Gibbon families living on either side of the track have not increased in size because there are not enough males. Gibbons are known to be monogamous. The gibbon sanctuary is also home to six other species of primates: capped langur, pig-tailed macaque, stump-tailed macaque, Assamese macaque, rhesus macaque, and slow loris; besides leopards, pythons and elephant herds. ľ from the January 3 Calcutta ôTelegraphö, posted to primfocus by Dr. Shirley McGreal
Aye-Aye Born at Bristol Zoo
Two-month-old Raz, an aye-aye and the second of this species of lemur to be born in Britain, still fits in the palm of his keeperĺs hand. The first aye-aye born in captivity was also at Bristol Zoo Gardens in 2005, when keepers hand-reared a female called Kintana.
The aye-aye has been hunted to near extinction in its native Madagascar. Four keepers are hand-rearing Raz by giving him two-hourly feeds round the clock. Keeper of small mammals at the zoo, Caroline Brown, said: ôWe made the decision to hand-rear this infant in advance of his birth because his mother has not had much success rearing her babies. So far he is doing well. He is gaining weight and seems strong. Aye-ayes are quite slow-developing babies and require an intensive feeding regime.ö
Once thought to be extinct, the aye aye is classified as endangered. Experts say there could be as few as 1,000 left. ľ from BBC News, January 8, 2008
Federal Bill Will Keep Chimps in Sanctuary
A federal bill, sponsored by Rep. Jim McCrery (R-LA) and Senators Richard Burr (R-NC) and Mary Landrieu (D-LA) will permanently retire chimpanzees housed at Chimp Haven, a sanctuary in Keithville, Louisiana. S. 1916, signed into law by President Bush on Dec. 21, 2007, amends the Chimpanzee Health Improvement, Maintenance and Protection Act (CHIMP Act) to restrict the research permitted on retired chimpanzees housed in the sanctuary system to only noninvasive behavioral studies and medical studies based on information gathered during the course of normal veterinary care. ľ From the National Association for Biomedical Research Update, January 10, 2008
Austriaĺs Supreme Court ľ Chimp Not Person
Austriaĺs Supreme Court has dashed hopes by animal rights activists to have a chimpanzee declared a person, a January 15 statement suggested. The court recently rejected a petition to appoint a trustee for the chimp, named Matthew Hiasl Pan. The Vienna-based Association Against Animal Factories has vowed to contact the European Court of Human Rights over the matter. ľ Associated Press, January 21
Malaysian Ministry Drops Plan to Export Monkeys
Kangar: The Malaysian Natural Resources and Environment Ministry will abandon its proposal to export long-tailed macaques, a common monkey found in urban areas. Minister Datuk Seri Azmi Khalid said the decision was made after the discovery that about 80 per cent of urban monkeys had contracted diseases and were deemed unfit for export.
The cabinet had, in August last year, proposed the idea to capture and export macaques in urban areas to control their numbers.
Azmi said that about 80 per cent of the 250,000 long-tailed macaques found in urban areas were found to have diseases such as tuberculosis, malaria, hepatitis and AIDS. He said the diseases could possibly spread among the human population. ôAfter a study was conducted recently, it was found the macaques were not suitable for export because they were infected. They were supposed to fulfill the demand for exotic meat in a few countries in Asia and in the west. A study of 2,000 macaques in urban areas, which had begun several months ago, found that 80 per cent of them were infected. Only 20 per cent were healthy, and of those, only half were suitable for export.ö
He said the ministry feared that the move to export monkeys would encourage companies to hire agents to seize the monkeys, thus leading to uncontrolled hunting of the animal. Currently, no permits have been issued for the export of macaques. The ministry had earlier decided to lift a 23-year-old export ban on macaques because of the increasing cases of attacks and disturbances by the monkeys. ľ Posted to primfocus by Shirley McGreal, February 2
Ban on Export of Malaysian Macaques Reinstated
The European Coalition to End Animal Experiments (ECEAE), which represents groups in 17 countries, welcomes the recent announcement made by the Minister of Natural Resources and Environment that the government intends to reinstate the ban on the export of macaques from Malaysia.
At a time when there is growing international concern over the plight of primates in areas such as research, it is heartening to learn that Malaysia intends to maintain the important ban that it implemented in 1984. Such a move will prevent the infliction of immense suffering, misery and death to many thousands of primates who would otherwise have been exported for the food and research industries.
The ECEAE now urges the government of Malaysia to address the underlying problem of human-monkey conflict in urban areas without resorting to the killing of monkeys. Conflict between human beings and wildlife over habitat is a growing issue facing many governments. There are ways to control monkey populations, however, which are not only humane, but are also more effective. The British Union for the Abolition of Vivisection has already submitted a paper to the Malaysia Natural Resources and Environment Minister Dato Seri Hj Azmi Khalid. The paper offered suggestions on effective means of controlling the conflicts humanely, including reproduction control, relocation, and educating the public to adopt behavior that does not encourage monkeys to rely on humans for food. ľ From a February 3 ECEAE Press Release
Bill to Conceal Names of Animal Researchers
Animal rights protesters are up in arms over a bill set to hit the Utah State Senate floor next week that would conceal the names, personal addresses, and phone numbers of animal researchers at state colleges and universities.
Harold Rose, an activist with Utah Primate Freedom, said the bill is unconstitutional and undermines basic principles of democracy. ôItĺs completely ridiculous,ö Rose said. ôIĺm amazed people are not more concerned. Something is being done with public tax dollars, and itĺs been removed from all public criticism.ö
Sen. Greg Bell, R-Fruit Heights, who is sponsoring the bill, said protesters from Utah Primate Freedom have crossed the line by harassing researchers, vandalizing their homes and signing them up for unwanted magazine subscriptions. ôThe public has the right to understand what research is going on...but they donĺt need to know specific information of workers,ö Bell said. ôBasic human dignity and decency requires that we have people in good faith pursuing legitimate research, but we shouldnĺt be harassing people just because we disagree with them. But that hasnĺt worked, so we have to resort to legal means.ö
The bill would modify the Government Records Access and Management Act (GRAMA) to classify personal information of researchers who do medical or scientific research on animals at institutions of higher education as protected documents. The legislation would not only keep information from protesters, but also from journalists or other individuals who use GRAMA. Bell said such individuals would have to appeal to the GRAMA committee for the information. ľ by R. McConkie, Daily Utah Chronicle, February 1
Pruetz Wins National Geographic Society Award
Iowa State University (ISU) primatologist Jill Pruetz has won a $10,000 award from the National Geographic Society given to emerging authorities in scientific fields. Pruetz made international news in February, 2007, after publishing a study that recorded, for the first time, habitual hunting by savannah chimpanzees in Senegal using spears they fashioned from tree branches. Also of wide scientific interest was the fact that female chimpanzees exhibited the behavior more than males, perhaps because they had to improvise to find food, Pruetz said. Male chimps can catch a monkey, for example, but the females generally canĺt, she said.
The ISU Associate Professor of Anthropology found that chimps, thought to avoid water, use ponds in Senegal to cool off in the hottest parts of the day. She discovered some chimps using caves regularly to escape the heat, another behavior she added to scientific annals.
Pruetz, who is researching and teaching in Senegal this semester, appeared at a press conference at ISU Thursday via an Internet connection from a hotel in Senegal. ôI canĺt think of a better award to win,ö said Pruetz, who is thinking of using the prize money to buy a used truck. The vehicle ISU provided ôis on its last legs,ö she said. She read the e-mail informing her of the award a couple of times before she noticed there a cash prize was included. ľ from the Des Moines Register, February 7, 2008
Texas Judge Decides Fate of Chimpanzees
Keithville, Louisiana ľ On Friday, February, 15, 2008, a Bexar County, Texas, judge handed down his decision to grant a motion for summary judgment filed by Primarily Primates, Inc., to send the six chimpanzees (Sarah, Keeli, Ivy, Sheba, Harper and Emma) back to the San Antonio facility. The chimpanzees were removed from Primarily Primates in November, 2006, at the request of the Texas State Appointed Receiver and were sent to Chimp Haven where they could receive appropriate care.
Primarily Primates was seized by the Texas State Attorneyĺs office on October 13, 2006, on allegations of mismanagement of charitable funds and animals living in substandard conditions. Chimp Haven was asked to accept the chimpanzees since they were in need of medical attention and had inadequate housing, and immediate intervention was required. After consulting with the Chimp Haven Board of Directors, it was deemed in the best interest of the chimpanzees to remove them from unhealthy living conditions and bring them to the sanctuary at Chimp Haven. The chimpanzees were reunited with their family group immediately and for the past 14 months enjoyed expansive enclosures, intensive care, a well balanced diet and health care.
In April, 2007 the Texas Attorney General settled the case against Primarily Primates. Primarily Primates was joined by Friends of Animals, who funded their legal battles. They subsequently sued Chimp Haven for return of their ôpropertyö, alleging that the transfer was temporary and printing misinformation about Chimp Haven.
ôWe were happy to step forward to help these chimpanzees in need and provide them with an exceptional home for over a year. We are disappointed that the court does not consider what is in the chimpanzeesĺ best interest, but instead must rule on Ĺpropertyĺ issues,ö said Dr. Linda Brent, Chimp Havenĺs President and Director. ôOur goal is to ensure the best future for these chimpanzees, and [we] feel their welfare is best served here. We are investigating options for appeal of the decision.ö ľ Chimp Haven press release, February 15, 2008
* * *
The New York Consortium in Evolutionary Primatology (NYCEP) invites you to attend the following lectures in the New York Regional Primatology Colloquium.
The Great Ape Trust of Iowa invites you to a special scientific event, Decade of the Mind III: Emergence of Mind. This groundbreaking symposium, May 7-9, 2008, in Des Moines, Iowa, will bring together internationally recognized scientists to discuss and explore the emergence of mind and consciousness in nonhuman primates with an emphasis on great apes. Registration for the symposium is limited to 200 members of the academic community. For details about speakers and registration, see <www.greatapetrust.org/decadeofthemind>.
The 2008 Callitrichid Conference will be in Los Angeles, hosted by the Los Angeles Zoo, on May 16-18, 2008. For information, contact Michelle Farmerie [412- 365-2385; e-mail [email protected]]; for pre-registration form and abstract guidelines, contact Erin Fleming [e-mail: [email protected]].
The Norwegian Consensus/National Platform on Alternatives (Norecopa), in cooperation with the Norwegian Animal Research Authority, the Norwegian Institute for Nature Research, and the Norwegian Polar Institute, will sponsor an international consensus meeting entitled: Harmonisation of the Care and Use of Animals in Field Research, at the Clarion Hotel Oslo Airport, Gardermoen Airport, Oslo, May 21-22, 2008. Studies in the wild, or on wild animals in captivity, pose a range of challenges that do not exist under the more controlled and standardized conditions of laboratory animal research. For complete information, provisional program, and registration form, see <www.norecopa.no/sider/tekst.asp?side=8&meny=Meeting on Field Research>.
ôA symposium, Measuring Zoo Animal Welfare: Combining Approaches and Overcoming Challenges, will be held May 29-30, 2008, at the Brookfield Zoo, Brookfield, Illinois, sponsored by the Chicago Zoological Society. This symposium is intended to provide a forum for the development of an integrative approach to zoo animal welfare science. While animal welfare science has been making great strides in quantifying farm and laboratory animal well-being, the application of some of the commonly used measures to a wide variety of zoo animals requires creative and innovative solutions. By convening a colloquium of international experts in the field of animal welfare science, we will present the opportunity to discuss the most recent developments in the science of animal well-being and how to apply them to zoo animals. We will also seek to identify some of the most urgent issues for zoo animal welfare and how we can start to address them immediately given the knowledge from other fields of welfare science. For more information, see <www.brookfieldzoo.org/AWsymposium>.ö
The American College of Veterinary Internal Medicine (ACVIM) will hold its Annual Forum June 4-7, 2008, at The Henry B. Gonzales Convention Center, San Antonio, Texas. For information, see <www.acvim.org/AnnualMeetingForum>, or contact ACVIM [800-245-9081; fax: 303-231-0880; e-mail: [email protected]].
The American Society of Primatologists will hold its 31st meeting in West Palm Beach, Florida, June 18-21, 2008. The meeting will be hosted by the Palm Beach Zoo and will take place at the Palm Beach County Convention Center. The West Palm Beach Marriott hotel will be the official hotel for the meeting and they are offering extremely competitive room rates ($109 per room per night for singles, doubles, triples, and quads). Scientific sessions, exhibits, and the silent auction will take place on June 19-21 at the Convention Center. The closing banquet will be held at the Palm Beach Zoo on the evening of June 21. For details, see <www.asp.org/meetings>.
The 29th World Veterinary Congress will be held July 27-31, 2008, in Vancouver, British Columbia, Canada. Online Registration is available at <www.meet-ics.com/wvac2008/registration.html>. For brochures and general information, see <www.meet-ics.com/wvac2008>.
The Executive Committee for the International Primatological Society XXII Congress, which will be held in Edinburgh, Scotland, August 3-8, 2008, has announced that a provisional program is now available at <www.ips2008.co.uk>.á
The Second International Conference on Acoustic Communication by Animals will be held August 12-15, 2008, at Oregon State University, Corvallis, Oregon. This conference will help researchers share ideas, data, and methods in the growing and exciting field of animal acoustic communication. The aim will be to bring together senior scholars, mid-career researchers and teachers, young investigators, and students to share information and ideas across animal taxa.á For further information, please see <oregonstate.edu/conferences/animalcommunication2008>.
The 2008 meeting of the Animal Behavior Society will be held August 14-19, in Snowbird, Colorado. See <abs.animalbehavior.org>.
The International Association for the Study of Pain (IASP) has set up a Special Interest Group on pain in non-human species. Their first meeting will be a satellite meeting to the IASP meeting in Glasgow, Scotland. The satellite meeting will be August 17-18, 2008. The first day will focus on measurement of the affective component of pain, and will include seminars from Jeff Mogil, Melissa Bateson, Paul Flecknell, Marian Scott, and Lyn Wiseman. The following dayĺs theme will be measuring pain in spontaneous disease models in animals. Speakers will include Duncan Lascelles, Dan Brown, Nigel Calcutt, Tony Buffington, Alex Livingston, Bernie Hansen, and Johnny Roughan. You do not need to be an IASP member to attend, nor do you need to register for the main IASP meeting. Registration for non-members is 180 Pounds ľ this covers the two days of scientific sessions, teas, coffees, and lunch. The meeting dinner, on the evening of day 1, is 30 pounds per person. Accommodation is available in nearby halls of residence at low cost. The two days should provide a great opportunity for all who are interested in animal pain to get together and get an update on both spontaneous and induced models of pain, and how to better assess pain in animals. For more details of the program, contact Paul Flecknell [e-mail [email protected]] or Duncan Lascellesá [e-mail: [email protected]]; or see <www.iasp-sig-non-human.com/meetings.html>.
The Institute for Laboratory Animal Research will hold an international conference September 23-26, 2008, in Washington, DC, on Animal Research in a Global Environment: Meeting the Challenges. The conference will cover topics such as: how to conduct research in countries with different guidelines for animal care and use; outsourcing studies; cultural differences in veterinary care and training; and issues specific to research with mice and nonhuman primates. More information will follow.
The International Society for Comparative Psychology (ISCP) will hold its 14th Biennial Meeting, October 9-11, 2008, in Buenos Aires, Argentina. The keynote speaker will be Stephen Suomi, of the U.S. National Institutes of Health, speaking on ôRisk, Resilience, and Gene X Environment Interactions in Rhesus Monkeys and Other Primatesö. ISCP President Robert Murphey, of UC-Davis, will speak on ôLimits of Natural Selectionö. Symposia involve topics ranging from conservation of endangered species to the neuroscience of emotion, recent progress in learning models, and human causal learning. Graduate students and other junior scholars are especially encouraged to attend this meeting. For more information, see <www.comparativepsychology.org>, or contact Ruben N. Muzio, ISCP Program Chair, at < [email protected]>.
The Association of Primate Veterinarians will meet November 5-8, 2008, in Indianapolis, Indiana. See <www.primatevets.org>.
The Academy of Surgical Research Annual Meeting will be held November 6-8, 2008,áat the Indianapolis Hyatt Hotel. The program includes presentations on: pain management, cardiovascular surgery, interventional radiology, management of surgical staff, and original surgical research; and the popular Technician Roundtable; as well as workshops and a choice of wet labs. For full program information (including guidelines on submitting an abstract to become a part of the program), registration form, and hotel information, go to the Academyĺs Website: <www.surgicalresearch.org>.á
The 59th American Association for Laboratory Animal Science National Meeting will be held November 9-13, 2008, in Indianapolis, Indiana. See <nationalmeeting.aalas.org>.
The Scientists Center for Animal Welfare (SCAW) Winter Conference will be held December 8-9, 2008, in San Antonio, Texas. For information and registration, see <www.scaw.com/conference.htm>; or contact SCAW at 7833 Walker Dr., Suite 410, Greenbelt, MD 20770 [301-345-3500; fax: 301-345-3503; e-mail: [email protected]].
The 26th Annual Symposium for Nonhuman Primate Models for AIDS will be held December 9-12, 2008, at the Ritz-Carlton San Juan Hotel, San Juan, Puerto Rico, co-sponsored by the Wisconsin NPRC and the Caribbean PRC. New to the 26th meeting is a session on disease pathology, neuroAIDS, and translational applications of nonhuman primate AIDS research. See <nhp2008.primate.wisc.edu> or contact Edi Chan [608-263-3500; e-mail: [email protected]].
The 2009 International Academic and Community Conference on Animals and Society: Minding Animals will be held July 13-19, 2009, at Newcastle, Australia, hosted by the University of Newcastle and the Society and Animals (Australia) Study Group This conference will bring together animal theorists and scientists from a broad range of academic disciplines, with representatives from nongovernmental organizations, government officials from several nations, and representatives from industry, to examine the interrelationships between human and nonhuman animals from cultural, historical, geographical, environmental, representational, moral, legal and political perspectives.
The conference will have six major themes and objectives: to reassess the relationship between the animal and environmental movements in light of climate change and other jointly-held threats and concerns; to examine how humans identify and represent nonhuman animals in art, literature, music, science, and in the media and on film; how, throughout history, the objectification of nonhuman animals and nature in science and society, religion and philosophy, has led to the abuse of nonhuman animals and how this has since been interpreted and evaluated; to examine how the lives of humans and companion and domesticated nonhuman animals are intertwined, and how science, human medicine, and veterinary medicine utilize these important connections; how the study of animals and society can better inform both the scientific study of animals and community activism and advocacy; and how science and community activism and advocacy can inform the study of nonhuman animals and society. See <www.mindinganimals.com>, where you may complete an ôExpression of Interest in Attendingö form. For more information, contact Rod Bennison [+61-(0)41-491-4040; e-mail: [email protected]].
* * *
On January 14, the Humane Society of the United States (HSUS) announced a ôcorporate combinationö with the Association of Veterinarians for Animal Rights (AVAR) to form the Humane Society Veterinary Medical Association (HSVMA). The American Veterinary Medical Association (AVMA) has posted their answers to some questions it has been asked regarding this new association, at <www.avma.org/issues/animal_welfare/hsvma_faq.asp>.
Gorilla Haven Update
Jane Dewar writes: ôWe just added an update including details on Joeĺs recent procedure, Oliver and his goats, a new blog, etc., to try to keep our friends and colleagues apprised of whatĺs going on at Gorilla Haven. See <www.gorilla-haven.org> and click the Feb 3, 2008 update...
ôOne thing I didnĺt include was a construction update ... the new steel crew arrived today (Monday) and I donĺt want to jinx anything!! Fingers crossed that weĺll see some progress soon and weĺll have something to report!ö
More Interesting Websites * * *
* * *
The California National Primate Research Center (CNPRC) is seeking a highly motivated, enthusiastic veterinarian to join its primate medicine team. Position responsibilities include daily clinical care, project support, and teaching in a fast-paced biomedical research facility. The CNPRC, at the University of California, Davis (UCD), houses approximately 5000 nonhuman primates of three different species and supports a diverse variety of research programs. Salary and appointment level will be commensurate with experience.
Qualifications include a DVM/VMD from an AVMA-accredited college of veterinary medicine and a current United States license. A candidate should have excellent communication skills, a service-oriented attitude, and a desire to work in a challenging environment.
For more information on the position, contact Dr. Laurie Brignolo [530-752-2514; e-mail: llbrignolo@ primate.ucdavis.edu] or Dr. Kari Christe [530-572-2514; e-mail: [email protected]]. You may apply to UCD Human Resources Administration Bldg., Orchard Park, Davis, CA 95616, or online at <www.hr.ucdavis.edu/Emp/Careers/Application_Process>. Required application materials are available at the UCD Human Resources Administration Building, on the Internet at <www.hr.ucdavis.edu>, or at the UCD Medical Center Personnel Office.
Research Scientist ľ Panama
The Smithsonian Tropical Research Institute (STRI), headquartered in the Republic of Panama, is seeking world-class scientists to establish research programs in any current field of marine or terrestrial research, working anywhere in the tropics. These fields include biological sciences, anthropology, archeology, paleontology, geology, and soil science. Ideally, research initiated by the successful applicants will complement existing programs (see <www.stri.org>). We are especially interested in research on vertebrate biology; anthropology and archeology; paleontology; and climate change.
Panama and adjacent regions of tropical America are rich in terrestrial and marine habitats, archeological sites, and geological deposits. STRI maintains sophisticated research laboratories, a library, and support centers in Panama City, as well as diverse facilities for field research throughout the tropics. A 100ĺ vessel is available for marine and coastal research. STRI scientists maintain cooperative research programs with a world-wide network of institutions. Opportunities for mentoring young scientists are available through a vigorous fellowship program, and formal teaching is possible through programs with affiliated universities.
Applicants should have a PhD degree and postdoctoral research experience in their fields. Interested candidates should submit a summary of research accomplishments and interests, CV, five significant reprints, and the names and contact information of three potential referees.
Annual salary is commensurate with experience. Laboratory set-up and relocation expenses are provided. Housing and family education allowances may apply. Spanish-English bilingual education is available in some Panamanian schools, which follow international curricula and standards.
Review of applications will begin in April, 2008. Please send applications electronically to the Director of STRI, c/o Ms. Luz Latorraca, Office of Human Resources [e-mail: [email protected]]. Address inquiries to Dr. William Wcislo, Chair, Search Committee [e-mail: [email protected]].
STRI is an equal opportunity employer and appointments are made regardless of nationality.
Assistant Professor ľ Central Washington
The Department of Anthropology at Central Washington University (CWU) invites applications for a tenure-track faculty position at the assistant professor level. This position entails a half-time teaching assignment in the interdisciplinary Primate Behavior and Ecology program [see www.cwu.edu/~primate] and a half-time teaching assignment as Associate Director of Centralĺs Chimpanzee and Human Communication Institute [see www.cwu.edu/~cwuchci].
The successful candidate will teach undergraduate and graduate courses in the Primate Behavior and Ecology program. All university faculty are expected to engage in teaching, research, and service, with an emphasis on research collaboration with students.
As Associate Director of the Chimpanzee and Human Communication Institute, the successful candidate will oversee daily operations of the Institute and must have demonstrated experience in humane care of chimpanzees. He or she will train and supervise approximately ten staff, manage the facility and the chimpanzeesĺ daily care, and work directly with graduate and undergraduate students conducting research projects based at the Institute. Duties at the Institute will continue half-time during summers.
Minimum qualifications:
Direct questions to Lori Sheeran in the Department of Anthropology [509-963-1434; e-mail: [email protected]]. CWU is an AA/EOE/Title IX Institution.
Primate Clinical Veterinarian ľ NIH
The National Institutes of Health (NIH) has a challenging position now available in the Division of Veterinary Resources (DVR), Veterinary Medicine Branch (VMB). Major responsibilities include the provision of clinical care, direction of technical staff, protocol review and facility operations.á This position requires a strong working knowledge of laboratory animal medicine, and the ability to work with a wide spectrum of individuals in a number of capacities. You will assist the attending veterinarian care for an average census of 1000 primates.á Practical experience with primate medicine is desirable. Qualifications include a DVM/VMD degree from an AVMA-accredited school or college of veterinary medicine.á Candidates must be National Veterinary Board certified, be licensed to practice veterinary medicine in at least one state, and have one year of progressively responsible professional veterinary experience. New graduates with a strong interest in laboratory animal medicine will be considered. Graduates of veterinary schools not AVMA-accredited must have Education Commission for Foreign Veterinary Graduates certification. Salary ranges from $74,503 to $140,880 and a full package of Civil Service benefits is available.
Interested candidates should submit a current CV, bibliography, and the names and addresses of three professional references.á Applications must be received by 4/30/2008 and sent to: Dr. Judith Davis, NIH, DVR, VMB, 14 Service Rd, MSC 5590, Bldg. 14G - Rm 103, Bethesda, MD 20892-5590.
* * *
The Section on Comparative Medicine of Wake Forest University Health Sciences offers training for veterinarians seeking careers in veterinary pathology, laboratory animal medicine (LAM), nonhuman primate medicine, and research.á Those in pathology and LAM are residencies for motivated individuals interested in pursuing American College of Veterinary Pathologists (ACVP) and American College of Laboratory Animal Medicine (ACLAM) board certification, respectively.á The period of training is flexible, with appointments for one year with annual renewal up to a total of three years contingent upon mutual interest.á The nonhuman primate medicine position is a two-year residency for veterinarians seeking careers in nonhuman primate clinical medicine.á The research fellowships are three-year positions designed to provide research training leading to a PhD degree.á Every trainee will have a mentor: an ACVP Diplomate, ACLAM Diplomate, or Research Mentor, depending on the discipline.á Coursework varies according to the position but may include Medical Primatology, Primate Behavior and Psychological Well-Being, Animal Models in Biomedical Research, Diseases of Laboratory Animals, and short courses including Management of Primate Breeding Colonies, Cost Accounting for Primate Facilities, and Occupational Health.á The training faculty includes 16 faculty members, 14 of whom are veterinarians, five of whom hold board certification by ACLAM, and two of whom hold board certification by ACVP. More information on the faculty can be found at <www1.wfubmc.edu/pathresearch/training>.
Applicants for the LAM and pathology residencies must have a DVM or equivalent degree and be U.S. citizens or legal residents.á Those for the nonhuman primate medicine residencies and research fellowships must have a DVM degree from an AVMA-accredited institution and be U.S. citizens.á To apply, send a statement of career goals, resume, official college transcripts, and three letters of reference to Nancy D. Kock, Dept of Pathology, Section on Comp. Med., Wake Forest Univ. Sch. of Med., Medical Center Blvd, Winston-Salem, NC 27157-1040 [e-mail: [email protected]].
Wake Forest University Health Sciences is an affirmative action/equal opportunity employer.á
Medical and Experimental Mammalian Genetics
Applications are being accepted for The 49th Annual Short Course in Medical and Experimental Mammalian Genetics, at the Jackson Laboratory, Bar Harbor, Maine. This yearĺs course will run from July 20 to August 1, 2008, and will include daily lectures in the mornings and evenings, supplemented by afternoon mini-symposia, workshops, tutorials, and demonstrations. There is special emphasis on student-faculty interaction. The faculty is diverse in terms of disciplines and the students in terms of stage of career and fields of concentration. The content of the Short Course focuses on an up-to-date presentation of genetics in experimental animals and humans, the relationship of heredity to disease in experimental animals and humans, and the importance of molecular genetics in the diagnosis and treatment of inherited disorders. The development and uses of modern techniques in bioinformatics, mathematical genetics, genome manipulation, mutagenesis, and phenotyping are taught in lectures and in afternoon workshop sessions. The Short Course includes an implicit focus on translational biology and clinical applications. Applicants need to possess a doctoral or terminal degree or be advanced graduate or medical students.
For more information, including application instructions and a preliminary schedule, see <www.jax.org/ courses/2008/49th_short_course.html>; or contact Nancy Place, Events & Meetings Planner, Jackson Laboratory, 600 Main St, Bar Harbor, ME 04609-1500 [207-288-6257; e-mail: [email protected]]. The deadline for scholarship requests and abstracts is May 1, 2008.
* * *
Books
* Recognition and Alleviation of Distress in Laboratory Animals (Prepublication Copy). Institute for Laboratory Animal Research. Washington, DC: National Academy Press, 2008, 104 pp.; <books.nap.edu/catalog.php?record_id=11931>.
Childrenĺs Books
* Evangeline Mudd and The Golden-Haired Apes of the Ikkinasti Jungle. D. Elliott; A. Wesson (Illus.). Cambridge, MA: Candlewick Press, 2004. [Price: $5.99]
Booklets
* Biosafety in Microbiological and Biomedical Laboratoriesá(BMBL), 5th Edition. U.S. Department of Health and Human Services, Centers for Disease Control and Prevention, & National Institutes of Health. Washington: U.S. Government Printing Office, 2007, <www.cdc.gov/od /ohs/biosfty/bmbl5/BMBL_5th_Edition.pdf>.
* The NICEATM-ICCVAM Five-Year Plan (2008-2012): A plan to advance alternative test methods of high scientific quality to protect and advance the health of people, animals, and the environment. National Toxicology Program Interagency Center for the Evaluation of Alternative Toxicological Methods & ICCVAM Interagency Coordinating Committee on the Validation of Alternative Methods. January, 2008. NIH Publ. No. 08-6410. <icvam.niehs .nih.gov/docs/5yrPlan/NICEATM5YR-Final.pdf>.
Magazines and Newsletters
* American Journal of Primatology, 2008, 70[1]. * American Journal of Primatology, 2008, 70[2]. * Borneo Orangutan Survival Foundation UKĺs Newsletter, Dec., 2007,
<www.savetheorangutan.co.uk/newsletter/december_07.pdf>.
* Borneo Orangutan Survival Foundation UKĺs Newsletter, Feb., 2008,
<www.savetheorangutan.co.uk/newsletter/february_08.pdf>.
* Folia Primatologica, 2008, 79[2]. * Journal of Medical Primatology, 2008, 37[1], <www.blackwell-synergy.com/toc/jmp/37/1>. * Animal Research News & Analysis Newsletter, January, 2008. [The Humane Society of the United States, 2100 L St, NW, Washington, DC 20037; <www.hsus.org/animals_in_research/general_information_on_animal_research/subscribe_to_the_animal_research_news_analysis_newsletter>].
* NIH Extramural Nexus, January, 2008, <grants.nih.gov/grants/partners/0108Nexus.htm>.
Reports
* HSUS Pain & Distress Report, 2008, 7[4]; <www.hsus.org/web-files/PDF/ARI/pd_reports/ pdreport_jan081.pdf>.
Special Journal Issues
* Microbial quality control for nonhuman primates. ILAR Journal, 2008, 49[2]. * Association of Primate Veterinarians 34th Annual Workshop. Journal of Medical Primatology, 2008, 37 [suppl. 1], <www.blackwell-synergy.com/toc/jmp/37/s1>. Anatomy and Physiology
* Behavioral shifts and action valuation in the anterior cingulate cortex. Quilodran, R., RothÚ, M., & Procyk, E. (E. P., Inserm, U846, Stem Cell & Brain Research Inst., 69500 Bron, France [e-mail: [email protected]]). Neuron, 2008, 57, 314-325. * A voice region in the monkey brain. Petkov, C. I., Kayser, C., Steudel, T., Whittingstall, K., Augath, M., & Logothetis, N. K. (Max Planck Inst. for Biol. Cybernetics, Spemannstr. 38, 72076 TŘbingen, Germany [e-mail: [email protected]]). Nature Neuroscience, 2008, 11, 367-374. Animal Models
* Basic math in monkeys and college students. Cantlon, J. F., & Brannon, E. M. (Dept of Psych. & Neurosci., Ctr for Cognitive Neurosci., Duke Univ., Durham, NC [e-mail: [email protected]]). PloS Biol 5 (12): e328 <dx.doi.org/10.1371/journal.pbio.0050328>. * Aging and fertility patterns in wild chimpanzees provide insights into the evolution of menopause. Thompson, M. E., Jones, J. H., Pusey, A. E., Brewer-Marsden, S., Goodall, J., Marsden, D., Matsuzawa, T., Nishida, T., Reynolds, V., Sugiyama, Y., & Wrangham, R. W. (Dept of Anthropology, Harvard Univ., 11 Divinity Ave, Cambridge, MA 02138 [[email protected]]). Current Biology, 2007, 17, 2150-2156. * Face perception in monkeys reared with no exposure to faces. Sugita, Y. (Neurosci. Res. Inst., Nat. Inst. of Adv. Industrial Sci. & Tech., Teragu 1497-1, Tsukuba 300-4201, Japan [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2008, 105, 394-398. * Sensory processing disorder in a primate model: Evidence from a longitudinal study of prenatal alcohol and prenatal stress effects. Schneider, M. L., Moore, C. F., Gajewski, L. L., Larson, J. A., Roberts, A. D., Converse, A. K., & DeJesus, O. T. (Dept of Kinesiology, 2175 Med. Sci. Ctr, 1300 University Ave, Madison, WI 53706-1532 [e-mail: [email protected]]. Child Development, 2008, 79, 100-113. * Reduction of IgG in nonhuman primates by a peptide antagonist of the neonatal Fc receptor FcRn. Mezo, A. R., McDonnell, K. A., Tan Hehir, C. A., Low, S. C., Palobella, V. J., Stattel, J. M., Kamphaus, G. D., Fraley, C., Zhang, Y., Dumont, J. A., & Bitonti, A. J. (Syntonix Pharmaceuticals, Inc., 9 Fourth Ave, Waltham, MA 02451 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2008, 105, 2337-2342. * Chimpanzee autarky. Brosnan, S. F., Grady, M. F., Lambeth, S. P., Schapiro, S. J., & Beran, M. J. (Language Research Ctr, Georgia State Univ., Atlanta, GA 30303 [[email protected]]). PloS ONE, 2008, 3 [1], e1518. <www.plosone.org/doi/pone.0001518>. Animal Welfare
* Refinement of a transplantation project in the non-human primate by the use of a humane endpoint. Fante, F., Bolddrin, M., Polito, L., Ravarotto, L., Castagnaro, M., Hutabba, S., Cozzi, E., & Ancona, E. (Inst. for Animal Health Prophylaxis, Viale dellĺUniversitÓ no. 10, 35020 Legnaro, Padua, Italy [e-mail: [email protected]]. Laboratory Animals, 2007, 41, 456-469. * Transforming environmental health protection. Collins, F. S., Gray, G. M., & Bucher, J. R. (Nat. Human Genome Res. Inst., NIH, Bethesda, MD 20892 [e-mail: [email protected]]). Science, 2008, 319, 906-907. Behavior
* Embraces for infant handling in spider monkeys: Evidence for a biological market? Slater, K. Y., Schaffner, C. M., & Aureli, F. (Research Dept, Operation Wallacea Trust, Hope House, Old Bolingbroke, Lincolnshire PE234EX, U.K. [e-mail: [email protected]]). Animal Behaviour, 2007, 74, 455-461. * Payment for sex in a macaque mating market. Gumert, M. D. (Dept of Biology, Hiram College, Hiram, OH 44234 [e-mail: [email protected]]. Animal Behaviour, 2007, 74, 1655-1667. * Phylogenetic analyses of behavior support existence of culture among wild chimpanzees. Lycett, S. J., Collard, M., & McGrew, W. C. (Univ. of Liverpool, Hartley Bldg, Brownlow St, Liverpool L69 3BX, U.K. [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2007, 104, 17588-17592. * Agonistic support in juvenile Japanese macaques: Cognitive and functional implications. Schino, G., Tiddi, B., & Polizzi di Sorrentino, E. (Via Lucilio 36, 00136 Roma, Italy [e-mail: [email protected]]). Ethology, 2007, 113, 1151-1157. * Interaction location outweighs the competitive advantage of numerical superiority in Cebus capucinus intergroup contests. Crofoot, M. C., Gilby, I. C., Wikelski, M. C., & Kays, R. W. (Dept of Anthropology, Harvard Univ., Cambridge, MA 02138 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2008, 105, 577-581. * Rapid facial mimicry in orangutan play. Ross, M. D., Menzler, S., & Zimmermann, E. (Dept of Psychology, Univ. of Portsmouth, Portsmouth, Hampshire PO1 2DY, U.K. [e-mail: [email protected]]). Biology Letters, 2008, 4, 27-30. * Group size, grooming and social cohesion in primates. Lehmann, J., Korstjens, A. H., & Dunbar, R. I. M. (School of Biol. Sci., Univ. of Liverpool, Crown St, Liverpool L69 7ZB, U.K. [e-mail: [email protected]]). Animal Behaviour, 2007, 74, 1617-1629. * Dominance rank influences female space use in wild chimpanzees, Pan troglodytes: Towards an ideal despotic distribution. Murray, C. M., Mane, S. V., & Pusey, A. E. (Univ. of Minnesota, 1987 Upper Buford Circle, St. Paul, MN 55108 [e-mail: [email protected]]). Animal Behaviour, 2007, 74, 1795-1804. * When pliers become fingers in the monkey motor system. UmiltÓ, M. A., Escola, L., Intskirveli, I., Grammont, F., Rochat, M., Caruana, F, Jezzini, A., Gallese, V., & Rizzolatti, G. (G. R., Dipto di Neuroscienze, Univ. di Parma, Via Volturno 39, 43100 Parma, Italy [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2008, 105, 2209-2213. * Free-ranging rhesus monkeys spontaneously individuate and enumerate small numbers of non-solid portions. Wood, J. N., Hauser, M. D., Glynn, D. D., & Barner, D. (Dept of Psychology, Harvard Univ., 33 Kirkland St, Cambridge, MA 02138 [e-mail: [email protected]]). Cognition, 2008, 106, 207-221. * Raking it in: The impact of enculturation on chimpanzee tool use. Furlong, E. E., Boose, K. J., & Boysen, S. T. (S. T. B., Ohio State Univ., 209 Psychology Bldg, 1835 Neil Ave., Columbus, OH 43210-1222 [e-mail: [email protected]]). Animal Cognition, 2008, 11, 83-97. * Other-regarding preferences in a non-human primate: Common marmosets provision food altruistically. Burkart, J. M., Fehr, E., Efferson, C., & van Schaik, C. P. (Anthropological Inst., Univ. of ZŘrich, Winterthurerstr. 190, 8057 ZŘrich, Switzerland [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2007, 104, 19762-19766. Conservation
* Why the bonobos need a radio and other (unlikely) lessons from deepest Congo. J. Falk (text) & R. J. Ross (photography). National Geographic Adventure, 2008, No. 77, 50-67, <www.nationalgeographic.com/adventure/john-falk/bonobos-congo.html>.
Disease
* Plasmodium knowlesi malaria in humans is widely distributed and potentially life threatening. Cox-Singh, J., Davis, T. M. E., Lee, K.-S., Shamsul, S. S. G., Matusop, A., Ratnam, S., Rahman, H. A., Conway, D. J., & Singh, B. (Malaria Res. Centre, Fac. of Med. & Health Sci., Univ. Malaysia Sarawak, Kuching, Sarawak, Malaysian Borneo [e-mail: [email protected]]). Clinical Infectious Diseases, 2008, 46, 165-171. * Generation of biologically contained Ebola viruses. Halfmann, P., Kim, J. H., Ebihara, H., Noda, T., Neumann, G., Feldmann, H., & Kawaoka, Y. (Dept of Pathobiol. Sci., Sch. of Vet. Med., Univ. of Wisconsin, 2015 Linden Dr., Madison, WI 53706 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2008, 105, 1129-1133. * Independent evolution of an antiviral TRIMCyp in rhesus macaques. Wilson, S. J., Webb, B. L. J., Ylinen, L. M. J., Verschoor, E., Heeney, J. L., & Towers, G. J. (G. J. T. University College Med. Sch., Univ. Coll. London, London W1T 4JF, U. K. [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2008, 105, 3557-3562. * Independent genesis of chimeric TRIM5-cyclophilin proteins in two primate species. Virgen, C. A., Kratovac, Z., Bieniasz, P. D., & Hatziioannou, T. (T. H., Aaron Diamond AIDS Research Center, Rockefeller Univ., 455 First Ave, New York, NY 10016 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2008, 105, 3563-3568. * TRIMCyp expression in Old World primates Macaca nemestrina and Macaca fascicularis. Brennan, G., Kozyrev, Y., & Hu, S.-L. (S.-L. H., Univ. of Washington, 3000 Western Ave, Seattle, WA 98121 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2008, 105, 3569-3574. Evolution, Genetics, and Taxonomy
* Gene copy number variation spanning 60 million years of human and primate evolution. Dumas, L., Kim, Y. H., Karimpour-Fard, A., Cox, M., Hopkins, J., Pollack, J. R., & Sikela, J. M. (J. M. S., Univ. of Colorado, Aurora, CO 80045 [e-mail [email protected]]). Genome Research, 2007, 17, 1266-1277. * Producing primate embryonic stem cells by somatic cell nuclear transfer. Byrne, J. A., Pedersen, D. A., Clepper, L. l., Nelson, M., Sanger, W. G., Gokhale, S., Wolf, D. P., & Mitalipov, S. M. (S. M. M., Oregon NPRC,, Oregon Health & Science Univ., 505 N.W. 185th Ave, Beaverton, OR 97006, [e-mail: [email protected]]. Nature, 2007, 450, 497-502. * Accelerated rate of gene gain and loss in primates. Hahn, M. W., Demuth, J. P., & Han, S.-G. (Dept of Biology & School of Informatics, 1001 E. 3rd St., Indiana Univ., Bloomington, IN 47405 [e-mail: [email protected]). Genetics, 2007, 177, 1941-1949. * The role of Pleistocene refugia and rivers in shaping gorilla genetic diversity in central Africa. Anthony, N. M., Johnson-Bawe, M., Jeffery, K., Clifford, S. L., Abernethy, K. A., Tutin, C. E., Lahm, S. A., White, L. J. T., Utley, J. F., Wickings, E. J., & Bruford, M. W. (Dept of Biol. Sci., Univ. of New Orleans, New Orleans, LA 70148 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2007, 104, 20432-20436. * Do na´ve primates recognize the vocalizations of felid predators? Yorzinski, J. L., & Ziegler, T. (2320 Storer Hall, Section of Evolution & Ecology, One Shields Avenue, Davis, CA 95616 [e-mail: [email protected]]). Ethology, 2007, 113, 1219-1227. * Development and application of a phylogenomic toolkit: Resolving the evolutionary history of Madagascarĺs lemurs. Horvath, J. E., Weisrock, D. W., Embry, S. L., Fiorentino, I., Balhoff, J. P., Kappeler, P., Wray, G. A., Willard, H. F., & Yoder, A. D. (Inst. for Genome Sci. & Policy, Duke Univ., Durham, NC 27708 [e-mail: [email protected]]). Genome Research, 2008, 18, 489-499. * 4,300-year-old chimpanzee sites and the origins of percussive stone technology. Mercader, J., Barton, H., Gillespie, J., Harris, J., Kuhn, S., Tyler, R., & Boesch, C. (Dept of Archaeology, Univ, of Calgary, Calgary, AB, Canada T2N 1N4 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2007, 104, 3043-3048. Instruments and Techniques
* Evaluating orangutan census techniques using nest decay rates: Implications for population estimates. Mathewson, P. D., Spehar, S. N., Meijaard, E., Nardiyono, Purnomo, Sasmirul, A., Sudiyanto, Oman, Sulhnudin, Jasary, Jumali, & Marshall, A. J. (E. M., Nature Conservancy-East Kali-mantan Prog., Jl. Polentas 5, Markoni, Balikpapan 76122, East Kalimantan, Indonesia [e-mail: [email protected]]). Ecological Applications, 2008, 18, 208ľ221. * Cosmogenic nuclide dating of Sahelanthropus tchadensis and Australopithecus bahrelghazali: Mio-Pliocene hominids from Chad. Lebatard, A.-E., BourlŔs, D. L., Duringer, P., Jolivet, M., Braucher, R., Carcaillet, J., Schuster, M., Arnaud, N., MoniÚ, P., Lihoreau, F., Likius, A., Mackaye, H. T., Vignaud, P., & Brunet, M. (D. L. B., UnitÚ Mixte de Recherche, CNRS 6635, Univ. Paul CÚzanne, Plateau de lĺArbois, 13545 Aix-en-Provence Cedex 04, France [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2008, 105, 3226-3231. * Evaluation of iron status in lemurs by analysis of serum iron and ferritin concentrations, total iron-binding capacity, and transferrin saturation. Williams, C. V., Junge, R. E., & Stalis, I. H. (Duke Lemur Center, Duke Univ., Durham, NC 27705). Journal of the American Veterinary Medical Association, 2008, 232, 578-585. Reproduction
* Paternal effects on offspring fitness in a multimale primate society. Charpentier, M. J. E., Van Horn, R. C., Altmann, J., & Alberts, S. C. (J. A., Dept of Ecol. & Evolutionary Biol., Princeton Univ., Princeton, NJ 08544 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2008, 105, 1988-1992, <www.pnas.org/cgi/content/full/105/6/1988>. ------------------------------
We would like to acknowledge Primate-Science as a source for information about new books.
------------------------------ * * *
Contents of Volumes 40-42 (2001-2003)
Contents of Volumes 43-45 (2004-2006)
Contents of Volumes 46-47 (2007-2009)
* * *
All correspondence concerning the Newsletter should be addressed to: Current and back issues of the Newsletter are available on the World Wide Web at The Newsletter is supported by Brown University
Cover photograph of a Douc Langur (Pygathrix nemaeus), taken at the San Diego Zoo by Paul G. Wilde, June 1997.
Copyright (c) 2008 by Brown University
News Briefs
Stella Brewer MarsdenMeeting Announcements
The American Association of Physical Anthropologists annual meeting will be held April 7-13, 2008, in Columbus, Ohio. See <www.physanth.org/annmeet>.
Please contact Shara Bailey < [email protected]> if you have any questions or to request dinner information. Information Requested or Available
AVMAĺs Responses to Questions about HSVMA
Enrichment Event
The New Jersey Association for Biomedical Research and Merck & Co. will be hosting their first Enrichment Extravaganza May 8, 2008, at the Best Western Palace Hotel in Fords, New Jersey. Guest speakers will include Temple Grandin, PhD, Steve Schapiro, PhD, Joseph Garner, PhD, & Diana Reiss, PhD.á The event will also feature vendors, panel discussions, and an enrichment competition.á Registration fee is $75; the deadline for registration is April 1.á Contact Gina Savastano [e-mail: [email protected]] or Genevieve Andrews-Kelly [e-mail: [email protected]] for registration and more information.
Positions Available
Veterinarian ľ Davis, California
Preferred qualifications:
Review of applications will begin March 30, 2008. Candidates must use the online application process at <jobs.cwu.edu>. A complete application includes:
Finalists will be asked to provide names and contact information (address, phone number and e-mail) for three references. Educational Opportunities
Training in Veterinary Pathology Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
. . .
A fictional view of the life of the child of primatologists; one of a series.
. . .
A plan to * research, develop, translate, and validate new and revised non-animal and other alternative assays for integration of relevant and reliable methods into federal agency testing programs; and * identify areas of high priority for new and revised non-animal and alternative assays or batteries of those assays to create a path forward for the replacement, reduction, and refinement of animal tests, when this is scientifically valid and appropriate.
. . .
Contents: Editorial, by P. Garber; Life history traits and parental care in Lepilemur ruficaudatus, by R. Hilgartner, D. Zinner, & P. M. Kappeler; Sex, age, and family differences in the chemical composition of owl monkey (Aotus nancymaae) subcaudal scent secretions, by E. A. MacDonald, E. Fernandez-duque, S. Evans, & L. R. Hagey; Assessment of circadian rhythms throughout the menstrual cycle of female rhesus monkeys, by L. K. Barger, T. M. Hoban-Higgins, & C. A. Fuller; Reproductive success in relation to dominance rank in the absence of prime-age males in Barbary macaques, by L. Modolo & R. D. Martin; The composition of milk from Bolivian squirrel monkeys (Saimiri boliviensis boliviensis), by L. A. Milligan, S. V. Gibson, L. E. Williams, & M. L. Power; Female ovarian cycle phase affects the timing of male sexual activity in free-ranging Barbary macaques (Macaca sylvanus) of Gibraltar, by M. Heistermann, K. Brauch, U. M÷hle, D. Pfefferle, J. Dittami, & K. Hodges; Innovative social behavior in chimpanzees (Pan troglodytes), by C. Casanova, R. Mondragon-Ceballos, & P. C. Lee; The effect of male tenure and female mate choice on paternity in free-ranging Japanese macaques, by E. Inoue & O. Takenaka; Landscape attributes affecting patch occupancy by howler monkeys (Alouatta palliata mexicana) at Los Tuxtlas, Mexico, by V. Arroyo-RodrÝguez, S. Mandujano, & J. BenÝtez-Malvido; The composition of milk from free-living common marmosets (Callithrix jacchus) in Brazil, by M. L. Power, C. E. Verona, C. Ruiz-Miranda, & O. T. Oftedal; Male parenting and response to infant stimuli in the common marmoset (Callithrix jacchus), by S. R. Zahed, S. L. Prudom, C. T. Snowdon, & T. E. Ziegler; Circadian cortisol secretion and circadian adrenal responses to ACTH are maintained in dexamethasone suppressed capuchin monkeys (Cebus apella), by C. Torres-Farfan, F. J. Valenzuela, R. Ebensperger, N. MÚndez, C. Campino, H. G. Richter, G. J. Valenzuela, & M. Serˇn-FerrÚ; and Preliminary assessment of natal attraction and infant handling in wild Colobus vellerosus, by L. J. N. Brent, J. A. Teichroeb, & P. Sicotte.
. . .
Contents: Use of palm trees as a sleeping site for hamadryas baboons (Papio hamadryas hamadryas) in Ethiopia, by A. Schreier & L. Swedell; Demography, life history and migrations in a Mexican mantled howler group in a rainforest fragment, by V. Arroyo-RodrÝguez, N. Asensio, & J. Cristˇbal-Azkarate; Why do captive tufted capuchins (Cebus apella) urine wash? by K. E. Miller, K. Laszlo, & S. J. Suomi; Ontogeny of conspecific and heterospecific alarm call recognition in wild Verreauxĺs sifakas (Propithecus verreauxi verreauxi), by C. Fichtel; Behavioral variation and reproductive success of male baboons (Papio anubis Î Papio hamadryas) in a hybrid social group, by T. J. Bergman, J. E. Phillips-Conroy, & C. J. Jolly; A preliminary study of food transfer in Sichuan snub-nosed monkeys (Rhinopithecus roxellana), by Z. Zhang, Y. Su, R. C. K. Chan, & G. Reimann; Sleeping sites, sleeping trees, and sleep-related behaviors of black crested gibbons (Nomascus concolor jingdongensis) at Mt. Wuliang, Central Yunnan, China, by P.-F. Fan & X.-L. Jiang; Social facilitation of fur rubbing behavior in white-faced capuchins, by H. Meunier, O. Petit, & J.-I. Deneubourg; Social organization of black-and-white snub-nosed monkeys (Rhinopithecus bieti) at Deqin, China, by L.-W. Cui, S. Huo, T. Zhong, Z.-F. Xiang, W. Xiao, & R.-C. Quan; Survey and comparison of major intestinal flora in captive and wild ring-tailed lemur (Lemur catta) populations, by L. M. Villers, S. S. Jang, C. L. Lent, S.-C. Lewin-Koh, & J. A. Norosoarinaivo; Behaviors in rhesus monkeys (Macaca mulatta) associated with activity counts measured by accelerometer, by A. Papailiou, E. Sullivan, & J. L. Cameron; Summation and quantity judgments of sequentially presented sets by capuchin monkeys (Cebus apella), by M. J. Beran, T. A. Evans, K. A. Leighty, E. H. Harris, & D. Rice; and Natal emigration by both sexes in the La Pacifica population of mantled howlers: When do some stay? by M. R. Clarke & K. E. Glander.
. . .
Contents: Somatic variation in living, wild ring-tailed lemurs (Lemur catta), by M. L. Sauther & F. P. Cuozzo; Allonursing in tufted capuchin monkeys (Cebus nigritus): Milk or pacifier? by M. C. Baldovino & M. S. Di Bitetti; Impact of male takeover on intra-unit sexual interactions and subsequent interbirth interval in wild Rhinopithecus roxellana, by D. Zhao, B. Li, C. P. Groves, & K. Watanabe; and New records on prey capture and meat eating by bonobos at Lui Kotale, Salonga National Park, Democratic Republic of Congo, by G. Hohmann & B. Fruth.
. . .
Contents: Invariant natural killer T cells from rhesus macaque spleen and peripheral blood are phenotypically and functionally distinct populations, by B. Gansuvd, J. Goodwin, C. K. Asiedu, X. L. Jiang, U. Jargal, P. Andrades, M. A. Exley, & J. M. Thomas; Blood biochemical indicators in young and adult Cebus apella of both sexes, by H. Nunez, M. Araya, F. Cisternas, M. Arredondo, M. Mendez, F. Pizarro, A. Ortiz, R. Ortiz, & M. Olivares; Gastroesophageal reflux disease in baboons (Papio sp.): A new animal model, by E. J. Glover, M. M. Leland, E. J. Dick, & G. B. Hubbard; Pegylated interferon-α2a treatment of chronic SIV-infected macaques, by D. M. Asmuth, K. Abel, M. D. George, S. Dandekar, R. B. Pollard, & C. J. Miller; The human-ABO blood groups of free-ranging long-tailed macaques (Macaca fascicularis) and parapatric rhesus macaques (M. mulatta) in Thailand, by S. Malaivijitnond, W. Sae-Low, & Y. Hamada; Anti-CeHV1 antibodies of two cynomolgus macaques cross-react with HSV2 but not HSV1 antigens in ELISA, by E. Coutrot, M. Blancher-Sardou, & A. Blancher; Genetic diversity of longtail macaques (Macaca fascicularis) on the island of Mauritius: An assessment of nuclear and mitochondrial DNA polymorphisms, by Y. Kawamoto, S. Kawamoto, K. Matsubayashi, K. Nozawa, T. Watanabe, M.-A. Stanley, & D. Perwitasari-Farajallah; and Characterization of spontaneous mammary gland carcinomas in female baboons, by J. A. Luth, G. B. Hubbard, E. J. Dick, S. R. Frazier, & B. F. Barrier.
. . .
Contents include information about the new Public Access Policy.
. . .
Contents: Specific pathogen-free nonhuman primates; Nonhuman primate quarantine: Its evolution and practice; Development, application, and quality control of serology assays used for diagnostic monitoring of laboratory nonhuman primates; New approaches to tuberculosis surveillance in nonhuman primates; Impact of infections and normal flora in nonhuman primates on drug development; Opportunistic infections in immunologically compromised nonhuman primates; NHP infections after organ transplantation; and Macaque models of human infectious disease.
. . .
Contents: Foreword, by D. R. Lee, P. C. Nossov, & E. N. Videan; Establishing a standardized dental record-keeping system for a small investigational colony of rhesus monkeys (Macaca mulatta), by B. W. Gibson, L. McGuffey, C. P. Raflo, & B. A. Niemiec; Spontaneous pancreatic islet cell tumor in a black and white colobus monkey (Colobus guereza kikuyuensis), by D. J. Hobson & P. V. Turner; Multiple ophthalmic abnormalities in an infant rhesus macaque (Macaca mulatta), by E. P. Ribka & R. R. Dubielzig; Diagnosis and prevention of dissemination of tuberculosis in a recently imported rhesus macaque (Macaca mulatta), by S. T. Shipley, T. Coksaygan, D. K. Johnson, C. G. McLeod, & L. J. DeTolla; Dermatitis in a rhesus macaque (Macaca mulatta) experimentally infected with simian immunodeficiency virus, by J. Wilk, A. Lewis, & V. Lukas; Relationship of creatine kinase, aspartate aminotransferase, lactate dehydrogenase, and proteinuria to cardiomyopathy in the owl monkey (Aotus vociferans), by A. S. Gozalo, A. Chavera, E. J. Montoya, J. Takano, & R. E. Weller; Sudden cardiac death in 13 captive chimpanzees (Pan troglodytes), by M. L. Lammey, D. R. Lee, J. J. Ely, & M. M. Sleeper; Immunological concepts in tuberculosis diagnostics for non-human primates: A review, by P. L. Lin, J. Yee, E. Klein, & N. W. Lerche; Assessment of progestin-only therapy for endometriosis in macaque, by G. Maginnis, J. Wilk, R. Carroll, & O. D. Slayden; A case of suspected contact dermatitis in a juvenile cynomolgus monkey (Macaca fascicularis), by J. Morris & M. Etheridge; Lethargy, ulcers, bronchopneumonia and death in two aged female bonnet macaques presumed to be caused by Cercopithicine herpes virus I, by B. A. Scharf, C.-H. Wan, M. Bluth, R. Eberle, E. N. Videan, E. Smith, & J. Coplan; Cervicovaginal evaluation in macaques used as a model for topical microbicide safety studies, by D. G. Scorpio, D. S. Ruben, Z. Liao, J. E. Hildreth, & C. A. Fletcher; Reversible medetomidine/ketamine anesthesia in captive capuchin monkeys (Cebus apella), by B. R. Theriault, D. A. Reed, & M. A. Niekrasz; Cerebellar abscess in a cynomolgus macaque (Macaca fascicularis), by J. S. Villano, B. Ogden, A. Goh, L. S. Hui, & P. K. H. Chow; Reproductive abnormalities in aged female Macaca fascicularis, by M. Wilkinson, S. Walters, T. Smith, & A. Wilkinson; and 34th Annual Workshop for the Association of Primate Veterinarians, October 12-14, 2006, Park City, Utah, USA.
. . .
ôRapid optimization of behavior requires decisions about when to explore and when to exploit discovered resources. The mechanisms that lead to fast adaptations and their interaction with action valuation are a central issue. We show here that the anterior cingulate cortex (ACC) encodes multiple feedbacks devoted to exploration and its immediate termination. In a task that alternates exploration and exploitation periods, the ACC monitored negative and positive outcomes relevant for different adaptations. In particular, it produced signals specific of the first reward, i.e., the end of exploration. Those signals disappeared in exploitation periods but immediately transferred to the initiation of trials Ś a transfer comparable to learning phenomena observed for dopaminergic neurons. Importantly, these were also observed for high gamma oscillations of local field potentials shown to correlate with brain imaging signal. Thus, mechanisms of action valuation and monitoring of events/actions are combined for rapid behavioral regulation.ö
. . .
ôFor vocal animals, recognizing species-specific vocalizations is important for survival and social interactions. In humans, a voice region has been identified that is sensitive to human voices and vocalizations. As this region also strongly responds to speech, it is unclear whether it is tightly associated with linguistic processing and is thus unique to humans. Using functional magnetic resonance imaging of macaque monkeys (Old World primates, Macaca mulatta), we discovered a high-level auditory region that prefers species-specific vocalizations over other vocalizations and sounds. This region not only showed sensitivity to the Ĺvoiceĺ of the species, but also to the vocal identify of conspecific individuals. The monkey voice region is located on the superior-temporal plane and belongs to an anterior auditory Ĺwhatĺ pathway. These results establish functional relationships with the human voice region and support the notion that, for different primate species, the anterior temporal regions of the brain are adapted for recognizing communication signals from conspecifics.ö
. . .
ôAdult humans possess a sophisticated repertoire of mathematical faculties. Many of these capacities are rooted in symbolic language and are therefore unlikely to be shared with nonhuman animals. However, a subset of these skills is shared with other animals, and this set is considered a cognitive vestige of our common evolutionary history. Current evidence indicates that humans and nonhuman animals share a core set of abilities for representing and comparing approximate numerosities nonverbally; however, it remains unclear whether nonhuman animals can perform approximate mental arithmetic. Here we show that monkeys can mentally add the numerical values of two sets of objects and choose a visual array that roughly corresponds to the arithmetic sum of these two sets. Furthermore, monkeysĺ performance during these calculations adheres to the same pattern as humans tested on the same nonverbal addition task. Our data demonstrate that nonverbal arithmetic is not unique to humans but is instead part of an evolutionarily primitive system for mathematical thinking shared by monkeys.ö
. . .
ôHuman menopause is remarkable in that reproductive senescence is markedly accelerated relative to somatic aging, leaving an extended postreproductive period for a large proportion of women. Functional explanations for this are debated, in part because comparative data from closely related species are inadequate. Existing studies of chimpanzees are based on very small samples and have not provided clear conclusions about the reproductive function of aging females. These studies have not examined whether reproductive senescence in chimpanzees exceeds the pace of general aging, as in humans, or occurs in parallel with declines in overall health, as in many other animals. In order to remedy these problems, we examined fertility and mortality patterns in six free-living chimpanzee populations. Chimpanzee and human birth rates show similar patterns of decline beginning in the fourth decade, suggesting that the physiology of reproductive senescence was relatively conserved in human evolution. However, in contrast to humans, chimpanzee fertility declines are consistent with declines in survivorship, and healthy females maintain high birth rates late into life. Thus, in contrast to recent claims, we find no evidence that menopause is a typical characteristic of chimpanzee life histories.ö
. . .
Infant monkeys were reared with no exposure to any faces for 6ľ24 months. Before being allowed to see a face, the monkeys showed a preference for human and monkey faces in photographs, and they discriminated human faces as well as monkey faces. After the deprivation period, the monkeys were exposed first to either human or monkey faces for a month. Soon after, the monkeys selectively discriminated the exposed species of face and showed a marked difficulty in regaining the ability to discriminate the other nonexposed species of face. These results indicate the existence of an experience-independent ability for face processing as well as an apparent sensitive period during which a broad but flexible face prototype develops into a concrete one for efficient processing of familiar faces.
. . .
Disrupted sensory processing, characterized by over- or underresponsiveness to environmental stimuli, has been reported in children with a variety of developmental disabilities. This study examined the effects of prenatal stress and moderate-level prenatal alcohol exposure on tactile sensitivity and its relationship to striatal dopamine system function in thirty-eight 5- to 7-year-old rhesus monkeys. The monkeys were from four experimental conditions: (a) prenatal alcohol exposed, (b) prenatal stress, (c) prenatal alcohol exposed + prenatal stress, and (d) sucrose controls. Increased D2 receptor binding in the striatum, evaluated using positron emission tomography neuroimaging, was related to increased withdrawal (aversion) responses to repetitive tactile stimuli and reduced habituation across trials. Moreover, prenatal stress significantly increased overall withdrawal responses to repetitive tactile stimulation compared to no prenatal stress.
. . .
ôThe neonatal Fc receptor FcRn provides IgG molecules with their characteristically long half-lives in vivo by protecting them from intracellular catabolism and then returning them to the extracellular space. Other investigators have demonstrated that mice lacking FcRn are protected from induction of various autoimmune diseases, presumably because of the accelerated catabolism of pathogenic IgGs in the animals. Therefore, targeting FcRn with a specific inhibitor may represent a unique approach for the treatment of autoimmune disease or other diseases where the reduction of pathogenic IgG will have a therapeutic benefit. Using phage display peptide libraries, we screened for ligands that bound to human FcRn (hFcRn) and discovered a consensus peptide sequence that binds to hFcRn and inhibits the binding of human IgG (hIgG) in vitro. Chemical optimization of the phage-identified sequences yielded the 26-amino acid peptide dimer SYN1436, which is capable of potent in vitro inhibition of the hIgGľhFcRn interaction. Administration of SYN1436 to mice transgenic for hFcRn induced an increase in the rate of catabolism of hIgG in a dose-dependent manner. Treatment of cynomolgus monkeys with SYN1436 led to a reduction of IgG by up to 80% without reducing serum albumin levels that also binds to FcRn. SYN1436 and related peptides thus represent a previously uncharacterized family of potential therapeutic agents for the treatment of humorally mediated autoimmune and other diseases.ö
. . .
ôEconomists believe that barter is the ultimate cause of social wealth ľ and even much of our human culture ľ yet little is known about the evolution and development of such behavior. It is useful to examine the circumstances under which other species will or will not barter to more fully understand the phenomenon. Chimpanzees (Pan troglodytes) are an interesting test case as they are an intelligent species, closely related to humans, and known to participate in reciprocal interactions and token economies with humans, yet they have not spontaneously developed costly barter. Although chimpanzees do engage in noncostly barter, in which otherwise valueless tokens are exchanged for food, this lack of risk is not typical of human barter. Thus, we systematically examined barter in chimpanzees to ascertain under what circumstances chimpanzees will engage in costly barter of commodities, that is, trading food items for other food items with a human experimenter. We found that chimpanzees do barter, relinquishing lower value items to obtain higher value items (and not the reverse). However, they do not trade in all beneficial situations, maintaining possession of less preferred items when the relative gains they stand to make are small. Two potential explanations for this puzzling behavior are that chimpanzees lack ownership norms, and thus have limited opportunity to benefit from the gains of trade; and that chimpanzeesĺ risk of defection is sufficiently high that large gains must be imminent to justify the risk. Understanding the conditions that support barter in chimpanzees may increase understanding of situations in which humans, too, do not maximize their gains.ö
. . .
An assessment scheme was developed to establish a humane endpoint in a pig-to-primate renal xenotransplantation project, with a view to minimizing and controlling any pain or suffering conditions in the animals involved while still achieving the scientific objective. In particular, the assessment criteria for identifying the earliest endpoint are described, bearing in mind both the researcherĺs need to obtain top-quality data and the ethical need to safeguard the animals. The scheme should also be applicable to other experiments involving nonhuman primates (e.g. allotransplantation, survival after major surgery, pharmacological safety tests) because it considers reproducible general parameters together with aspects specific to each experimental model.
. . .
The U.S. Environmental Protection Agency, the U.S. National Toxicology Program, and the National Institutes of Health Chemical Genomics Center have established a collaborative research program. The studies proposed will test whether high-throughput and computational toxicology approaches can yield data predictive of results from animal toxicity studies, will allow prioritization of chemicals for further testing, and can assist in prediction of risk to humans. It is a research program that, if successful, will eventually lead to new approaches for safety assessment and a marked refinement and reduction of animal use in toxicology.
. . .
ôThe presence of young infants influences female primate social behavior, in particular social grooming, which may be exchanged for infant access within a biological market place. Although social grooming is common among Old World primate species, it is rarely observed in a number of New World primate species, including spider monkeys. We investigated whether affiliative behavior is exchanged for infant handling among wild female spider monkeys, Ateles geoffroyi yucatanesis. Occurrences of approaches, grooming and embraces received by 15 adult females were compared when they had infants less than 6 months old (Ĺmothersĺ) and at all other times. The occurrence of grooming was not influenced by the presence of infants. However, females received significantly more approaches and embraces when they were mothers compared to all other times. As mothers, they received significantly more embraces than they gave indicating that the increase in friendly behavior received was due to increased interest from other females and not simply a general increase in sociality. The exchange of embraces for infant handling was subject to a market effect as the proportion of embraces given to mothers followed by infant handling decreased when fewer infants were present in the group. Although spider monkeys use embraces as the primary means of gaining access to infants, whether embraces provide long-term benefits to the recipient, remains to be determined. Thus, embraces may only serve to signal benign intent and reassure mothers during tense interactions such as infant handling.ö
. . .
ôIn primate sexual relationships, males and females can cooperate through social trade. Market-like trading of sexual activity has been theorized, but no data have yet been presented that clearly show its existence. I collected data to test whether biological market theory could account for exchanges of male-to-female grooming and sexual activity in longtailed macaques. I explored male-to-female grooming, rates of sexual activity, and groomingľmating interchanges, which were male-to-female grooming bouts that directly involved mating. Male-to-female grooming mainly occurred when females were sexually active, and males groomed females longer per bout when mating, inspection, or presentation of female hindquarters was involved. Moreover, male-to-female grooming was associated with an increase in female rates for all forms of sexual activity, where in contrast, female-to-male grooming was associated with decreased rates of mating in the groomed males. Males did not preferentially mate with swollen females or invest more grooming in them during groomingľmating interchanges, as swellings did not seem to be a reliable indicator of female fertility. Rank status was correlated with grooming payment during groomingľmating interchanges in favour of higher-ranked males and females. In support of a biological market interpretation, the amount of grooming a male performed on a female during groomingľmating interchanges was related to the current supply of females around the interaction. The results provided evidence of a groomingľmating trade that was influenced by a mating market.ö
. . .
ôCulture has long been considered to be not only unique to humans, but also responsible for making us qualitatively different from all other forms of life. In recent years, however, researchers studying chimpanzees (Pan troglodytes) have challenged this idea. Natural populations of chimpanzees have been found to vary greatly in their behavior. Because many of these interpopulation differences cannot be readily explained by ecological factors, it has been argued that they result from social learning and, therefore, can be regarded as cultural variations. Recent studies showing social transmission in captive chimpanzee populations suggest that this hypothesis is plausible. However, the culture hypothesis has been questioned on the grounds that the behavioral variation may be explained at a proximate level by genetic differences between subspecies. Here we use cladistic analyses of the major cross-site behavioral data set to test the hypothesis that the behavioral differences among the best-documented chimpanzee populations are genetically determined. If behavioral diversity is primarily the product of genetic differences between subspecies, then population data should show less phylogenetic structure when data from a single subspecies (P. t. schweinfurthii) are compared with data from two subspecies (P. t. verus and P. t. schweinfurthii) analyzed together. Our findings are inconsistent with the hypothesis that the observed behavioral patterns of wild chimpanzee populations can be explained primarily by genetic differences between subspecies. Instead, our results support the suggestion that the behavioral patterns are the product of social learning and, therefore, can be considered cultural.ö
. . .
Agonistic support is generally considered a classical example of altruistic behaviour. It plays a critical role in the process of rank acquisition. This study investigates agonistic coalitions involving juvenile Japanese macaques (Macaca fuscata) living in a large captive group. Kinship was a good predictor of agonistic support received by juvenile macaques. Kinship also had a significant, though weaker, influence on agonistic support given by juvenile macaques. The latter was strongly influenced by the prospective rank of the individual against which the coalition was directed, i.e. by the rank relationship between the target of the coalition and the juvenileĺs mother. Juvenile macaques intervened aggressively more against prospectively subordinate individuals than against prospectively dominant individuals. These results imply agonistic support is not necessarily altruistic, and support the hypothesis that primates may understand the rank relationships that exist among their group mates, i.e., that they are aware of third-party social relationships. They also suggest that selective pressures associated with the process of rank acquisition may have favored the evolution of higher cognitive abilities.
. . .
ôNumerical superiority confers a competitive advantage during contests among animal groups, shaping patterns of resource access, and, by extension, fitness. However, relative group size does not always determine the winner of intergroup contests. Smaller, presumably weaker social groups often defeat their larger neighbors, but how and when they are able to do so remains poorly understood. Models of competition between individuals suggest that location may influence contest outcome. However, because of the logistical difficulties of studying intergroup interactions, previous studies have been unable to determine how contest location and group size interact to shape relationships among groups. We address this question by using an automated radio telemetry system to study intergroup interactions among six Cebus capucinus social groups of varying sizes. We find that the odds of winning increase with relative group size; one additional group member increases the odds of winning an interaction by 10%. However, this effect is not uniform across space; with each 100 m that a group moves away from the center of its home range, its odds of winning an interaction decrease by 31%. We demonstrate that contest outcome depends on an interaction between group size and location, such that small groups can defeat much larger groups near the center of their home range. The tendency of resident groups to win contests may help explain how small groups persist in areas with intense intergroup competition.ö
. . .
ôEmotional contagion enables individuals to experience emotions of others. This important empathic phenomenon is closely linked to facial mimicry, where facial displays evoke the same facial expressions in social partners. In humans, facial mimicry can be voluntary or involuntary, whereby its latter mode can be processed as rapidly as within or at 1 sec. Thus far, studies have not provided evidence of rapid involuntary facial mimicry in animals. This study assessed whether rapid involuntary facial mimicry is present in orangutans (Pongo pygmaeus; N=25) for their open-mouth faces (OMFs) during everyday dyadic play. Results clearly indicated that orangutans rapidly mimicked OMFs of their playmates within or at 1 sec. Our study revealed the first evidence on rapid involuntary facial mimicry in nonhuman mammals. This finding suggests that fundamental building blocks of positive emotional contagion and empathy that link to rapid involuntary facial mimicry in humans have homologues in nonhuman primates.ö
. . .
ôMost primates live in social groups in which affiliative bonds exist between individuals. Because these bonds need to be maintained through social interactions (grooming in most primates), sociality will be limited by time constraints. It has previously been shown that the time primates invest in grooming increases with group size. However, when groups become too large, individuals will not have enough time available to service all possible social relationships and group cohesion is expected to decrease. In this study, we used data from previously published studies to determine how large groups compromise on their grooming time and how ecological, phylogenetic and life history variables affect time invested in grooming (across species as well as within taxa). We used path analysis to analyze direct and indirect (via group size) effects on grooming. We showed that not only is grooming time determined by group size, but it is also affected by dispersal patterns and sex ratio. Furthermore, we found that grooming time is asymptotic when group size exceeds 40 individuals, indicating that time constraints resulting from ecological pressure force individuals to compromise on their grooming time. This was true across species, but a similar effect was also found within taxa. Cognitive constraints and predation pressure strongly affect group sizes and thereby have an indirect effect on primate grooming time. Primates that were found to live in groups larger than predicted by their neocortex size usually suffered from greater predation risk. However, most populations in our analysis were placed well within what we define as their eco-cognitive niche.ö
. . .
ôStudies from many different taxa have demonstrated that dominance rank greatly influences individual space use. While the importance of dominance among female chimpanzees, Pan troglodytes, was debated in the past, mounting evidence now shows that rank is very important. In particular, rank has been shown to influence body mass, foraging strategies, association patterns, and ultimately, reproductive success. In this study, we investigated how rank influenced female space use among chimpanzees, P.t. schweinfurthii, at Gombe National Park, Tanzania. Analyzing 10 years of data, we found that new immigrants used areas away from dominant females, and that subordinates had lower site fidelity. We also found that high-ranking females had smaller core areas and that this size difference was pronounced during periods of food scarcity when food competition is highest. These patterns suggest that dominant females outcompete subordinates, forcing them to settle elsewhere, range more widely, and shift their space use across time.ö
. . .
ôThe capacity to use tools is a fundamental evolutionary achievement. Its essence stands in the capacity to transfer a proximal goal (grasp a tool) to a distal goal (e.g., grasp food). Where and how does this goal transfer occur? Here, we show that, in monkeys trained to use tools, cortical motor neurons, active during hand grasping, also become active during grasping with pliers, as if the pliers were now the hand fingers. This motor embodiment occurs both for normal pliers and for Ĺreverse pliersĺ, an implement that requires finger opening, instead of their closing, to grasp an object. We conclude that the capacity to use tools is based on an inherently goal-centered functional organization of primate cortical motor areas.ö
. . .
Fundamental questions in cognitive science concern the origins and nature of the units that compose visual experience. Here, the capacity to individuate and store information about non-solid portions is investigated, asking in particular whether free-ranging rhesus monkeys (Macaca mulatta) quantify portions of a non-solid substance presented in discrete pouring actions. When presented with portions of carrot pieces poured from a cup into opaque boxes, rhesus picked the box with the greatest number of portions for comparisons of 1 vs. 2, 2 vs. 3, and 3 vs. 4, but not for comparisons of 4 vs. 5 and 3 vs. 6. Additional experiments indicate that rhesus based their decisions on both the number of portions and the total amount of food. These results show that the capacity to individuate non-solid portions is not unique to humans, and does not depend on structures of natural language. Further, the fact that rhesusĺ ability to represent non-solid portions is constrained by the same 4-item limit typically ascribed to the system of parallel individuation that operates over solid objects suggests that the visual system recruits common working memory processes for retaining information about solid objects and non-solid portions. Results are discussed with respect to theories of visual processing, as well as to the role that the human language faculty may have played in both the evolution and development of quantification.
. . .
ôRecent evidence for different tool kits, proposed to be based upon culture-like transmission, have been observed across different chimpanzee communities in Western Africa. In light of these findings, the reported failures by seven captive juvenile chimpanzees tested with 27 tool use tasks (Povinelli, 2000) seem enigmatic. Here we report successful performance by a group of nine captive, enculturated chimpanzees, and limited success by a group of six semi-enculturated chimpanzees, on two of the Povinelli tasks, the Flimsy Tool task, and the Hybrid Tool task. All chimpanzees were presented with a rake with a flimsy head and a second rake with a rigid head, either of which could be used to attempt to retrieve a food reward that was out of reach. The rigid rake was constructed such that it had the necessary functional features to permit successful retrieval, while the flimsy rake did not. Both chimpanzee groups in the present experiment selected the functional rigid tool correctly to use during the Flimsy Tool task. All animals were then presented with two ôhybrid rakesö A and B, with one half of each rake head constructed from flimsy, non-functional fabric, and the other half of the head made of wood. Food rewards were placed in front of the rigid side of Rake A and the flimsy side of Rake B. To be successful, the chimps needed to choose the rake that had the reward in front of the rigid side of the rake head. The fully enculturated animals were successful in selecting the functional rake, while the semi-enculturated subjects chose randomly between the two hybrid tools. Compared with findings from Povinelli, whose non-enculturated animals failed both tasks, our results demonstrate that chimpanzees reared under conditions of semi-enculturation could learn to discriminate correctly the necessary tool through trial-and-error during the Flimsy Tool task, but were unable to recognize the functional relationship necessary for retrieving the reward with the ôhybridö rake. In contrast, the enculturated chimpanzees were correct in their choices during both tasks. These results provide the first empirical evidence for the differential effects of enculturation on subsequent tool use capacities in captive chimpanzees.ö
. . .
ôHuman cooperation is unparalleled in the animal world and rests on an altruistic concern for the welfare of genetically unrelated strangers. The evolutionary roots of human altruism remain poorly understood. Recent evidence suggests a discontinuity between humans and other primates because individual chimpanzees do not spontaneously provide food to other group members, indicating a lack of concern for their welfare. Here, we demonstrate that common marmoset monkeys (Callithrix jacchus) do spontaneously provide food to nonreciprocating and genetically unrelated individuals, indicating that other-regarding preferences are not unique to humans and that their evolution did not require advanced cognitive abilities such as theory of mind. Because humans and marmosets are cooperative breeders and the only two primate taxa in which such unsolicited prosociality has been found, we conclude that these prosocial predispositions may emanate from cooperative breeding.ö
. . .
ôUntil recently, Plasmodium knowlesi malaria in humans was misdiagnosed as Plasmodium malariae malaria. The objectives of the present study were to determine the geographic distribution of P. knowlesi malaria in the human population in Malaysia and to investigate 4 suspected fatal cases. Sensitive and specific nested polymerase chain reaction was used to identify all Plasmodium species present in (1) blood samples obtained from 960 patients with malaria who were hospitalized in Sarawak, Malaysian Borneo, during 2001ľ2006; (2) 54 P. malariae archival blood films from 15 districts in Sabah, Malaysian Borneo (during 2003ľ2005), and 4 districts in Pahang, Peninsular Malaysia (during 2004ľ2005); and (3) 4 patients whose suspected cause of death was P. knowlesi malaria. For the 4 latter cases, available clinical and laboratory data were reviewed. P. knowlesi DNA was detected in 266 (27.7%) of 960 of the samples from Sarawak hospitals, 41 (83.7%) of 49 from Sabah, and all 5 from Pahang. Only P. knowlesi DNA was detected in archival blood films from the 4 patients who died. All were hyperparasitemic and developed marked hepatorenal dysfunction. Human infection with P. knowlesi, commonly misidentified as the more benign P. malariae, are widely distributed across Malaysian Borneo and extend to Peninsular Malaysia. Because P. knowlesi replicates every 24 h, rapid diagnosis and prompt effective treatment are essential. In the absence of a specific routine diagnostic test for P. knowlesi malaria, we recommend that patients who reside in or have traveled to Southeast Asia and who have received a ôP. malariaeö hyperparasitemia diagnosis by microscopy receive intensive management as appropriate for severe falciparum malaria.ö
. . .
ôEbola virus (EBOV), a public health concern in Africa and a potential biological weapon, is classified as a biosafety level-4 agent because of its high mortality rate and the lack of approved vaccines and antivirals. Basic research into the mechanisms of EBOV pathogenicity and the development of effective countermeasures are restricted by the current biosafety classification of EBOVs. We therefore developed biologically contained EBOV that express a reporter gene instead of the VP30 gene, which encodes an essential transcription factor. A Vero cell line that stably expresses VP30 provides this essential protein in trans and biologically confines the virus to its complete replication cycle in this cell line. This complementation approach is highly efficient because biologically contained EBOVs lacking the VP30 gene grow to titers similar to those obtained with wild-type virus. Moreover, EBOVs lacking the VP30 gene are indistinguishable in their morphology from wild-type virus and are genetically stable, as determined by sequence analysis after seven serial passages in VP30-expressing Vero cells. We propose that this system provides a safe means to handle EBOV outside a biosafety level-4 facility and will stimulate critical studies on the EBOV life cycle as well as large-scale screening efforts for compounds with activity against this lethal virus.ö
. . .
ôThe antiretroviral restriction factor TRIM5 has recently emerged as an important mediator of innate immunity and species-specific inhibition of retroviral replication in mammals. Selection pressure from pathogenic infection has driven rapid evolution of TRIM5 genes, leading to the antiviral specificities we see today. Remarkably, the New World owl monkey (Aotus trivirgatus) encodes a TRIM5 protein in which the antiviral determinants in the B30.2 domain have been replaced by cyclophilin A (CypA) encoded by a retrotransposed cDNA. The owl monkey TRIMCyp protein restricts infection by a subset of lentiviruses that recruit CypA to their capsids, including HIV-1 and feline immunodeficiency virus. Here, we show that the Old World monkey, rhesus macaque (Macaca mulatta), also encodes a TRIMCyp protein that has arisen independently from that in owl monkeys. The rhesus TRIMCyp is encoded by a single, but common, allele (Mamu7) of the rhesus TRIM5 gene, among at least six further alleles that encode full-length TRIM5 proteins with no homology to CypA. The antiviral specificity of the rhesus TRIMCyp is distinct, restricting infection of HIV-2 and feline immunodeficiency virus but not HIV-1. Restriction by rhesus TRIMCyp is before reverse transcription and inhibited by blocking CypA binding, with cyclosporine A, or by mutation of the capsid CypA binding site. These observations suggest a mechanism of restriction that is conserved between TRIMCyp proteins. The lack of activity against HIV-1 suggests that Mamu7 homozygous animals will be null for TRIM5-mediated restriction of HIV-1 and could contribute to improved animal models for HIV/AIDS.ö
. . .
ôThe host range of retroviruses is influenced by antiviral proteins such as TRIM5, a restriction factor that recognizes and inactivates incoming retroviral capsids. Remarkably, in owl monkeys (omk), a cyclophilin A (CypA) cDNA has been transposed into the TRIM5 locus, resulting in the expression of a TRIM5-CypA fusion protein (TRIMCyp) that restricts retroviral infection based on the retroviral capsid-binding specificity of CypA. Here, we report that the seemingly improbable genesis of TRIMCyp has, in fact, occurred twice, and pigtailed macaques (pgt) express an independently generated TRIMCyp protein. The omkTRIMCyp and pgtTRIMCyp proteins restrict infection by several lentiviruses, but their specificities are distinguishable. Surprisingly, pgtTRIMCyp cannot bind to or restrict HIV-1 capsids as a consequence of a point mutation close to the Cyp:capsid-binding interface that was acquired during or after transposition of pgtCypA. However, the same mutation confers on pgtTRIMCyp the ability to restrict FIV in the presence of cyclosporin A, a drug that normally abolishes the interaction between pgtTRIMCyp or omkTRIMCyp and lentiviral capsids. Overall, an intuitively unlikely evolutionary event has, in fact, occurred at least twice in primates and represents a striking example of convergent evolution in divergent species.ö
. . .
ôPrimates have evolved a variety of restriction factors that prevent retroviral replication. One such factor, TRIM5α, mediates a postentry restriction in many Old World primates. Among New World primates, Aotus trivirgatus exerts a similar early restriction mediated by TRIMCyp, a TRIM5ľcyclophilin A (CypA) chimera resulting from a CypA retrotransposition between exons 7 and 8 of the TRIM5 gene. Macaca nemestrina do not express TRIM5α
; therefore, we asked whether these animals and related Old World primates express TRIMCyp. RT-PCR of total RNA from M. nemestrina and Macaca fascicularis yielded three TRIMCyp amplification products, one of which is predicted to encode a TRIMCyp chimera containing a full-length CypA. Unlike A. trivirgatus, genomic sequencing of M. nemestrina and M. fascicularis identifies a CypA retrotransposition in the 3ĺ untranslated region of the TRIM5 locus. There is ≈78% homology between the predicted protein sequences of Old World and New World primate TRIMCyp, with most of the differences found in the TRIM5-derived sequence. Notably, exon 7 is absent from both M. nemestrina and M. fascicularis TRIMCyp. Neither M. nemestrina nor M. fascicularis TRIMCyp could restrict HIV-1 or simian immunodeficiency virus SIVmac in an in vitro infectivity assay. The discovery of TRIMCyp in both M. nemestrina and M. fascicularis indicates that TRIMCyp expression may be more common among Old World primates than previously believed. Convergent evolution of TRIMCyp in both Old World and New World primates suggests that TRIMCyp may have provided evolutionary advantages.ö
. . .
ôGiven the evolutionary importance of gene duplication to the emergence of species-specific traits, we have extended the application of cDNA array-based comparative genomic hybridization (aCGH) to survey gene duplications and losses genome-wide across 10 primate species, including human. Using human cDNA arrays that contained 41,126 cDNAs, corresponding to 24,473 unique human genes, we identified 4159 genes that likely represent most of the major lineage-specific gene copy number gains and losses that have occurred in these species over the past 60 million years. We analyzed 1,233,780 gene-to-gene data points and found that gene gains typically outnumbered losses (ratio of gains/losses = 2.34) and these frequently cluster in complex and dynamic genomic regions that are likely to serve as gene nurseries. Almost one-third of all human genes (6696) exhibit an aCGH-predicted change in copy number in one or more of these species, and within-species gene amplification is also evident. Many of the genes identified here are likely to be important to lineage-specific traits including, for example, human-specific duplications of the AQP7 gene, which represent intriguing candidates to underlie the key physiological adaptations in thermoregulation and energy utilization that permitted human endurance running.ö
. . .
ôDerivation of embryonic stem (ES) cells genetically identical to a patient by somatic cell nuclear transfer (SCNT) holds the potential to cure or alleviate the symptoms of many degenerative diseases while circumventing concerns regarding rejection by the host immune system. However, the concept has only been achieved in the mouse, whereas inefficient reprogramming and poor embryonic development characterize the results obtained in primates. Here, we used a modified SCNT approach to produce rhesus macaque blastocysts from adult skin fibroblasts, and successfully isolated two ES cell lines from these embryos. DNA analysis confirmed that nuclear DNA was identical to donor somatic cells and that mitochondrial DNA originated from oocytes. Both cell lines exhibited normal ES cell morphology, expressed key stem-cell markers, were transcriptionally similar to control ES cells and differentiated into multiple cell types in vitro and in vivo. Our results represent successful nuclear reprogramming of adult somatic cells into pluripotent ES cells and demonstrate proof-of-concept for therapeutic cloning in primates.ö
. . .
ôThe molecular changes responsible for the evolution of modern humans have primarily been discussed in terms of individual nucleotide substitutions in regulatory or protein coding sequences. However, rates of nucleotide substitution are slowed in primates, and thus humans and chimpanzees are highly similar at the nucleotide level. We find that a third source of molecular evolution, gene gain and loss, is accelerated in primates relative to other mammals. Using a novel method that allows estimation of rate heterogeneity among lineages, we find that the rate of gene turnover in humans is more than 2.5 times faster than in other mammals and may be due to both mutational and selective forces. By reconciling the gene trees for all the gene families included in the analysis, we are able to independently verify the numbers of inferred duplications. We also use two methods based on the genome assembly of rhesus macaque to further verify our results. Our analyses identify several gene families that have expanded or contracted more rapidly than is expected even after accounting for an overall rate acceleration in primates, including brain-related families that have more than doubled in size in humans. Many of the families showing large expansions also show evidence for positive selection on their nucleotide sequences, suggesting that selection has been important in shaping copy-number differences among mammals. These findings may help explain why humans and chimpanzees show high similarity between orthologous nucleotides yet great morphological and behavioral differences.ö
. . .
The role of Pleistocene forest refugia and rivers in the evolutionary diversification of tropical biota has been the subject of considerable debate. A range-wide analysis of gorilla mitochondrial and nuclear variation was used to test the potential role of both refugia and rivers in shaping genetic diversity in current populations. Results reveal strong patterns of regional differentiation that are consistent with refugial hypotheses for central Africa. Four major mitochondrial haplogroups are evident with the greatest divergence between eastern (A, B) and western (C, D) gorillas. Coalescent simulations reject a model of recent eastľwest separation during the last glacial maximum but are consistent with a divergence time within the Pleistocene. Microsatellite data also support a similar regional pattern of population genetic structure. Signatures of demographic expansion were detected in eastern lowland (B) and Gabon/Congo (D3) mitochondrial haplogroups and are consistent with a history of postglacial expansion from formerly isolated refugia. Although most mitochondrial haplogroups are regionally defined, limited admixture is evident between neighboring haplogroups. Mantel tests reveal a significant isolation-by-distance effect among western lowland gorilla populations. However, mitochondrial genetic distances also correlate with the distance required to circumnavigate intervening rivers, indicating a possible role for rivers in partitioning gorilla genetic diversity. Comparative data are needed to evaluate the importance of both mechanisms of vicariance in other African rainforest taxa.
. . .
ôTraits that were adaptive under previous conditions may no longer have fitness benefits. However, some spe-cies still retain certain antipredator behaviors even though they do not coexist with predators that their ancestors once faced. Studies have examined the responses of a variety of na´ve species to these predators, but none have specifically investigated whether na´ve primates retain antipredator behaviors against felid predators. We studied the pig-tailed langur (Simias concolor) to determine whether it still recognizes felids as predators even though dangerous felids do not exist on the islands on which it inhabits. The responses of the langurs to playbacks of the vocalizations of felids (an ancestral predator), elephants (an unknown animal but not a predator), humans (a known predator), and pigs and birds (known animals but not predators) were compared. Langurs fled more slowly and looked at the speaker less in response to the felid and elephant calls than they did in response to human voices. Similar numbers of langurs fled in response to all playback treatments except the pig and bird. The results suggest that langurs are afraid of novel vocalizations but have not retained specific acoustic knowledge of felid predator vocalizations. For long-lived species that have extended periods of learning, being able to modify general behavioral responses, such as antipredator behaviors, based on experience may be more adaptive than having fixed behavioral strategies.ö
. . .
ôLemurs and the other strepsirrhine primates are of great interest to the primate genomics community due to their phylogenetic placement as the sister lineage to all other primates. Previous attempts to resolve the phylogeny of lemurs employed limited mitochondrial or small nuclear data sets, with many relationships poorly supported or entirely unresolved. We used genomic resources to develop 11 novel markers from nine chromosomes, representing about 9 kb of nuclear sequence data. In combination with previously published nuclear and mitochondrial loci, this yields a data set of more than 16 kb and adds about 275 kb of DNA sequence to current databases. Our phylogenetic analyses confirm hypotheses of lemuriform monophyly and provide robust resolution of the phylogenetic relationships among the five lemuriform families. We verify that the genus Daubentonia is the sister lineage to all other lemurs. The Cheirogaleidae and Lepilemuridae are sister taxa and together form the sister lineage to the Indriidae; this clade is the sister lineage to the Lemuridae. Divergence time estimates indicate that lemurs are an ancient group, with their initial diversification occurring around the Cretaceous-Tertiary boundary. Given the power of this data set to resolve branches in a notoriously problematic area of primate phylogeny, we anticipate that our phylogenomic toolkit will be of value to other studies of primate phylogeny and diversification. Moreover, the methods applied will be broadly applicable to other taxonomic groups where phylogenetic relationships have been notoriously difficult to resolve.ö
. . .
Archeological research in the African rainforest reveals unexpected results in the search for the origins of hominoid technology. The ancient Panin sites from C˘te dĺIvoire constitute the only evidence of prehistoric ape behavior known to date anywhere in the world. Recent archeological work has yielded behaviorally modified stones, dated by chronometric means to 4,300 years of age, lodging starch residue suggestive of prehistoric dietary practices by ancient chimpanzees. The ôChimpanzee Stone Ageö pre-dates the advent of settled farming villages in this part of the African rainforest and suggests that percussive material culture could have been inherited from an common human-chimpanzee clade, rather than invented by hominins, or have arisen by imitation, or resulted from independent technological convergence.
. . .
ôAn accurate estimate for orangutan nest decay time is a crucial factor in commonly used methods for estimating orangutan population size. Decay rates are known to vary, but the decay process and, thus, the temporal and spatial variation in decay time are poorly understood. We used established line-transect methodology to survey orangutan nests in a lowland forest in East Kalimantan, Indonesia, and monitored the decay of 663 nests over 20 months. Using Markov chain analysis, we calculated a decay time of 602 days, which is significantly longer than times found in other studies. Based on this, we recalculated the orangutan density estimate for a site in East Kalimantan; the resulting density is much lower than previous estimates (previous estimates were 3 to 8 times higher than our recalculated density). Our data suggest that short-term studies where decay times are determined using matrix mathematics may produce unreliable decay times. Our findings have implications for other parts of the orangutan range where population estimates are based on potentially unreliable nest decay rate estimates, and we recommend that for various parts of the orangutan range census estimates be reexamined. Considering the high variation in decay rates there is a need to move away from using single-number decay time estimates and, preferably, to test methods that do not rely on nest decay times as alternatives for rapid assessments of orangutan habitat for conservation in Borneo.ö
. . .
Ages were determined at two hominid localities from the Chad Basin in the Djurab Desert (Northern Chad). In the Koro Toro fossiliferous area, KT 12 locality (16░00ĺN, 18░53ĺE) was the site of discovery of Australopithecus bahrelghazali (Abel) and, in the Toros-Menalla fossiliferous area, TM 266 locality (16░15ĺN, 17░29ĺE) was the site of discovery of Sahelanthropus tchadensis (Touma´). At both localities, the evolutive degree of the associated fossil mammal assemblages allowed a biochronological estimation of the hominid remains: early Pliocene (3ľ3.5 Ma) at KT 12 and late Miocene (≈ 7 Ma) at TM 266. Atmospheric 10Be, a cosmogenic nuclide, was used to quasicontinuously date these sedimentary units. The authigenic 10Be/9Be dating of a pelite relic within the sedimentary level containing Abel yields an age of 3.58 ▒ 0.27 Ma that points to the contemporaneity of Australopithecus bahrelghazali (Abel) with Australopithecus afarensis (Lucy). The 28 10Be/9Be ages determined, within the anthracotheriid unit containing Touma´ bracket, the age of Sahelanthropus tchadensis to lie between 6.8 and 7.2 Ma. This chronological constraint is an important cornerstone both for establishing the earliest stages of hominid evolution and for new calibrations of the molecular clock.
. . .
Serum iron and ferritin concentrations, total iron-binding capacity, and transferrin saturation were assessed as indicators of iron metabolic status in three genera of lemurs, to determine whether these variables are useful for screening for iron overload. Blood samples were collected weekly for 3 weeks and assayed for serum iron and ferritin concentrations and total iron-binding capacity. Liver biopsy specimens were evaluated histologically and assayed for total iron, nonheme iron, and trace mineral concentrations. Deposition of iron was scored on Prussian blueľstained slides. Hepatic iron content ranged from 497 to 12,800 Pg/g dry weight (median, 2,165 Pg/g). Differences were seen in mean hepatic iron content across genera, with ruffed lemurs having the highest concentrations and ring-tailed lemurs having the lowest. Iron accumulation in the liver was mild, and cellular pathologic changes associated with iron storage disease were not detected in any lemur. Ferritin concentration was the only variable that correlated significantly with hepatic iron content in all 3 genera of lemurs; however, both transferrin saturation and serum iron concentration were correlated with hepatic iron concentration in ring-tailed and ruffed lemurs. Serum ferritin concentration was the only variable that was consistently correlated with hepatic iron content in all 3 genera. Mean hepatic iron content varied across genera, suggesting that the propensity for lemurs to develop iron overload in captivity may vary across taxa.
. . .
ôWhen females mate with multiple males, paternal care is generally expected to be negligible, because it may be difficult or impossible for males to discriminate their own offspring from those of other males, and because engaging in paternal care may reduce male mating opportunities. Consequently, males in multimale societies are not pre-dicted to provide direct benefits to their offspring. We have recently demonstrated, however, that males in a typical multimale primate society (yellow baboons, Papio cynocephalus) discriminate their own offspring from those of other males and provide care to them in the form of repeated support during agonistic encounters. This observation raises the question of whether fathers enhance offspring fitness in this species. Here we use 30 years of data on age at maturity for 118 yellow baboons with known fathers. We show that the fatherĺs presence in the offspringĺs social group during the offspringĺs immature period accelerated the timing of physiological maturation in daughters. Sons also experienced accelerated maturation if their father was present during their immature period, but only if the father was high ranking at the time of their birth. Because age at reproductive maturity has a large impact on lifetime reproductive success, our results indicate a direct effect of paternal presence on offspring fitness. This relationship in turn suggests that the multiple roles that males play in multimale animal societies have not been sufficiently examined or appreciated and that paternal effects may be more pervasive than previously appreciated.ö
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; FAX: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
http://www.brown.edu/primate
ACKNOWLEDGMENTS
Last Updated: March 19, 2008