Direction Selectivity in motion-sensitive neurons of the visual
pathway
and miscellaneous topics/demos in perception of motion
Motion is a vector defined by direction and speed.
2006: We will review 2 paper from Fried, Munch & Werblin:
"Mechanisms and circuitry underlying directional selectivity in the retina",
Nature 420: 411 (2002)
link
to Nature 2002 paper and
Notes on 2002 Nature paper: Voltage
clamping is used to separate excitatory and inhibitory inputs to a DS cell: During
clamping the amount of current needed to "clamp" the voltage of the cell
is measured while the cell is stimulated. To begin with, the key result is Fig 1(e),
where inhibition is seen off-center from the physical position of the DS g-cell
being recorded from (the "spatial profile" they call it). Time is now
right-to-left. Compare to Fig 1(b). In Fig 1(f) the same result is obtained with
flashing spots, not moving bars. They estimate the inhibitory offset at 150 microns
of distance on the retina: the cell responsible for the "inhibitory lobe, the
starburst amacrine, has a dendritic field that extends the distance required.
Fig 2 shows simultaneous recordings from
DS cell and starburst amacrines. Fig 2(d, e) has DS g-cell processes in green,
starburst amacrines in cyan.
"Directional selectivity is formed at multiple levels by laterally offset
inhibition in rabbit retina", Neuron 46: 117 (2005)
link
to Neuron 2005 paper
Notes on 2005 Neuron paper: Are any amacrines cells already
direc. selec. before reacting with DS G-cells?
linked from:
http://www.hms.harvard.edu/bss/neuro/bornlab/nb204/papers
Previously we had considered, on the topic of directional selectivity,
a paper by Werblin's post-doc advisor, Horace Barlow, and W. Levick, reviewed below.
Notes on Barlow & Levick
(1965)
Horace Barlow & Bill Levick, "The mechanism of directionally
selective units in the rabbit's retina," J. Physiol. 179:
477 (1965), Barlow and Levick recorded from rabbit retinal ganglion cells and showed
that a population of the cells are selective for the direction of movement a small
target takes, moving through a cell's "receptive field" of a 10 or so
square degrees of visual angle. In the Discussion section of their paper Barlow
and Levick put forth two models, or hypotheses, for explaining direction selectivity,
and associate parts of the models with more distal cells in the retina, such as
amacrine cells.
Definition of receptive field (RF): The region of space in which
visual stimuli can influence firing of a single neuron, such as ganglion cell of
rabbit retina. Testing with small ON and OFF stationary stimuli is one way to map
spatial extent of RF.
Basic Observation: Response in spikes/sec to one direction of movement,
but not the opposite direction.
Fig 2: No significant trend in latencies across RF: How can asymmetric latencies
explain DS?
Figs 3, 4: DS occurs throughout the RF
Fig 5: the slit experiment: the subunit of DS appears to be a fraction of the total
RF, and is "duplicated" a dozen times across the field.
Fig 6: Collision: Two slit timing experiment: response the the second slit depends
on which slit went first.
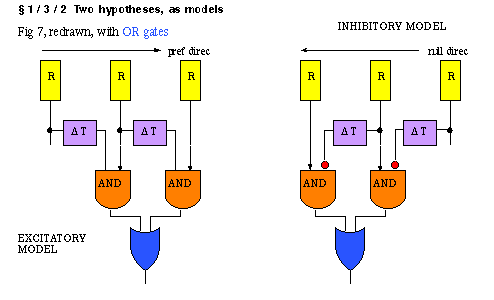
What is a critical test, to decide between the two models? Full field sustained
stimulus: excitatory should respond in sustained way, inhibitory model in transient
way. Inhibitory model provides the correct prediction.
Another approach: What is delta T? the order of msec...one test: response to v.
slow movement in null direction: response breaks through. Another test: response
to 2 slits timed for null and preferred apparent motion:
Maturana and Frenk, in pigeons: Collision test of preferred
and null simultaneously moving:
after Barlow: Nigel Daw tried a pharmacology test in
rabbit retina, antagonizing inhibition with picrotoxin delivered through an artery
headed to retina.
%-------------OTHER TOPICS IN PERCEPTION OF MOTION, below
----------------- -------------
Demo: Pulfrich effect
Both eyes are yoked to be fixating the same point. The filtered
eye is reporting what it saw about 25 msec earlier than the unfiltered eye (increased
retinal delay). The diagrams below illustrate why you have the perception of nearness
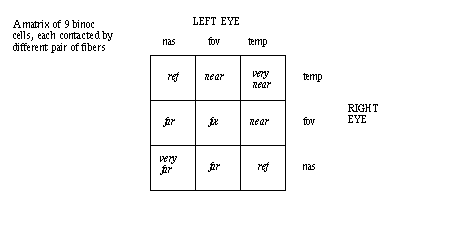
for left-right motion and left eye filter. In the CNS are 3x3 cells, each receiving
a different left-right pair of inputs.
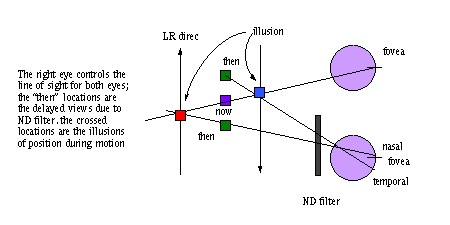
An explanation in terms of binocular units in the visual cortex
is given below:
Juggling in strobe light
What can you learn about perception of motion-apparent motion-from the time
between strobe flashes at the "cut off" frequency below which juggling
breaks up? That interval is about 100 msec. Presumably the brain has the ability
to correctly predict the path of movement if the sample rate is greater than
10 Hz.
Illusion of apparent motion and color. jump
to link. fixate on cross in middle. How long does it take to fatigue the
"magenta" pathway?
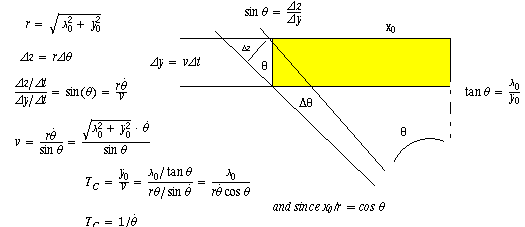
Optic flow
Computing time to collision by knowing only theta, theta dot and x0. Solve for
time to collision = Tc.
Certainly Tc is y0/v, but we need to compute what they are in the terms given.
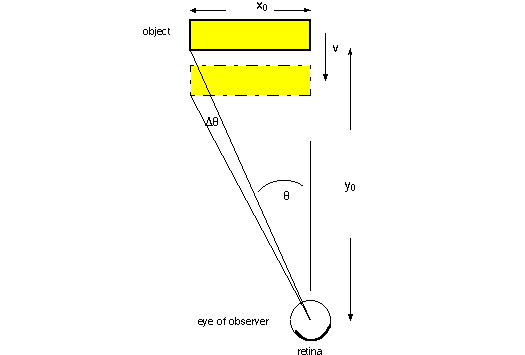
with a derivation of the solution shown in the detail below.
Cortical area MT ("medial temporal"): most of
the neurons there responds best to motion, of particular speed, direction, with
some limitation of receptive field. See asgn area
MT.