 Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter
Laboratory Primate Newsletter VOLUME 48 NUMBER 3 JULY 2009
Printable (PDF) Version of this issue
The Effects of Exposure to an Expanded Environmental Enrichment Program on Select Individual Behaviors in Baboons, by A. K. Goodwin, S. A. James, K. E. Lane, M. C. McDermott, R. L. Rodgerson, & N. A. Ator......1
Female Homosexual Behavior in a New Group of Japanese Macaques: Evolutionary Implications, by P. L. Vasey & C. Reinhart ......8
Looking for Information and Possible Collaboration, by G. Tansey......11
News, Information, and Announcements
Workshop Announcements: Association of Primate Veterinarians ......7
Information Requested or Available ......11
Gorilla Haven Update; Caging Survey; More Interesting Websites
Meeting Announcements ......12
Resources Wanted and Available ......12
Management Innovations in Laboratory Animal Care; Preventing Disease from Animals in Public Setting; Used Equipment Buying and Selling Sites; IPPL News Archive On-Line
Travellers� Health Notes: IAMAT ......13
News Briefs ......14
New Orangutan Population Found in Indonesia; Health Care for Mountain Gorillas and Humans; Carole C. Noon; Use of Experimental Primates Shown; �Ingenious� Ape Mounts Bid to Escape Zoo; Primate Research Center Under Scrutiny
Departments
Recent Books and Articles ......10
Position Available: Atlanta, Georgia......32
* * *
The Effects of Exposure to an Expanded Environmental Enrichment Program on Select Individual Behaviors in Baboons
Amy K. Goodwin, Susan A. James, Kelly E. Lane, Michael C. McDermott, Rebecca L. Rodgerson, and Nancy A. Ator
Johns Hopkins University School of Medicine, Department of Psychiatry and Behavioral Sciences, Division of Behavioral Biology
In our laboratory, we had often discussed our desire to create an area in which our singly housed, adult male baboons could be released to exercise. The opportunity to do so arose when an environmental enrichment grant from the Center for Alternatives to Animal Testing at Johns Hopkins University provided funding for such a project. Thus, the goals of the present study were to create an area large enough for baboons to safely engage in species-specific behavior (i.e., free movement, exploratory behavior, foraging) and to learn whether exposure to this environment would be correlated with changes in target behaviors considered indicators of psychological well-being.
Methods
Subjects: Six adult male baboons (Papio anubis; Primate Imports, New York, NY, or Southwest Foundation for Biomedical Research, San Antonio, TX) served as subjects in the present study. The six baboons (BB, DC, SY, CY, DE and SC) had been in the laboratory for at least three years and had been subjects in behavioral pharmacology studies. During the present study, the behavioral pharmacology studies in which they participated included acute exposure to psychoactive compounds. Subjects were individually housed in stainless steel primate cages equipped with a bench running along one cage wall. The home cages, which provided 10 square feet of floor space (46.5 cubic feet total space), also served as the experimental chambers, so the behavioral pharmacology experiments took place in the home cage. Such experiments typically involve the use of one or more levers, stimulus lights, tones, and food pellet delivery. The baboons had visual and auditory access to other baboons.
The baboons had continuous access to tap water from a spout at the front of their cages and received a daily ration of Lab Diet (®PMI Nutrition International) or Primate diet (®Harlan Teklad) biscuits, one or two pieces of fresh fruits or vegetables daily, and children�s chewable multivitamins. Daily feeding occurred in late morning (never prior to time spent in the enrichment room). The enrichment program already established in the colony, which included human interaction, access to three or more toys (forage boxes, puzzle feeders, mirrors, Kong® toys, wood logs), and music continued without change during the present study.
The overhead lights in the housing room were on for 13 h/day (6:00-19:00 h) and off for the remaining 11 h/day. Natural light also illuminated the room.
Routine physical examinations (under ketamine hydrochloride anesthesia) occurred every two weeks or approximately once per month, depending on the study in which each subject was serving.
All protocols were approved by the Johns Hopkins University Animal Care and Use Committee. Animal care and use and facility maintenance followed the Guide for the Care and Use of Laboratory Animals (1996) and the U. S. Animal Welfare Regulations. Johns Hopkins University is accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International.
Room Construction: It was vital that the time and effort required to create the enrichment room space be manageable and that the expense stay below the $6000 received for the project from the Animal Welfare Enhancement Award. Ultimately, by using resources already available to us, we were able to manage the time and effort required without affecting the normal daily functioning in the laboratory.
The space designated for the enrichment room had been constructed as an animal housing room (i.e., wall and floor surfaces met Guide standards; air was vented externally), and all environmental aspects (e.g., temperature and humidity) were controllable by laboratory personnel. The space was not needed for housing animals when the project began. The room has 145.8 square feet of floor space (663.5 cubic feet of total space), a fully functional sink, and a steel railing on three walls.
A door had to be constructed such that baboons would be able to be safely transferred in and out of the room using the shuttle system described below. In order to accomplish this without permanently altering the room, a door measuring 83.25� by 47.25� was made of aluminum sheets in a frame of 3� x 1.25� 6061 architectural aluminum, and installed in the existing door frame in front of the existing door (see Figure 1). A sub-frame containing a portion of the front wall of an old baboon cage was mounted in the center of the new aluminum door and welded to the larger door frame such that the shuttle cage could be connected to the embedded guillotine-style door. Once the shuttle is attached, the guillotine-style doors on the shuttle cage and the new �cage front� door are opened and the baboon is able to pass from the shuttle into the room. The cage front door is shut behind the baboon. When a baboon is not in the room, the aluminum door functions as any door would, allowing personnel into the room so that it may be cleaned between baboons.

Figure 1: The door constructed to safely transfer baboons in and out of the enrichment room.
Experimental Design and Procedures: The psychological well-being of nonhuman primates must be based on individual needs, thus a single-subject design was used in which each subject served as his own experimental control (Sidman, 1960). A subject�s target behavior was measured before the intervention (i.e., exposure to enrichment room) and then measurements in the home cage continued as baboons were periodically exposed to the enrichment room. Prior to any enrichment room exposure, the frequency of the target behavior in the home cage was recorded using a time-sampling procedure (Martin & Pear, 1992) in which 2-minute observations were conducted every hour for 8 hours (i.e., every hour from 7 a.m. to 3 p.m.) at least three days a week, over at least three weeks (i.e., a minimum of 72 two-minute observations). After subjects began spending time in the enrichment room, the home cage observations occurred once per week on a day when the baboon was not scheduled to be in the enrichment room (e.g., eight 2-minute observations on Fridays).

Figure 2: A baboon in the enrichment room. The wooden structure on the left is the painted and hinged structure used to block baboon access to a sink located in the enrichment room. Shown in the picture are various types of plastic zoo balls, plastic tires, tree branches, mirrors, plastic chains, and cardboard for shredding. In addition, small food items (e.g., raisins, peanuts, cashews, popcorn, sunflower seeds, etc.) are placed throughout the room to encourage foraging behavior.
Three baboons (BB, DC, and SY) were identified as exhibiting behaviors in their home cages for which a decrease in the frequency may indicate improvement in psychological well-being. For baboon BB, the behaviors were pacing and circling in his home cage. For baboons DC and SY, a �huddled� posture, operationally defined as sitting with chin on chest and being unresponsive to normal stimuli (e.g., our saying his name or offering food treats), was identified as such a behavior. In addition, baboon SY was identified as engaging in excessive grooming. For all three baboons, manipulating items/toys provided in the home cage was identified as a behavior for which increases in frequency may signal improvements in psychological well-being and was also recorded.
Periodic cage washes (e.g., every two weeks) require baboons to be transferred out of their home cages and into a temporary cage via a �shuttle�. After the shuttle is attached to the front of a baboon�s home cage, the guillotine style doors on the cage front and shuttle front are lifted and the baboon is able to enter the shuttle. The doors are then shut, and the baboon is transferred to a temporary cage while his home cage is washed. The same procedures are used to transfer the baboons back to their home cages. Although most baboons readily participate in this procedure, some baboons consistently require coaxing to enter the shuttle. Typically, veterinary technicians attempt to coax baboons into the shuttle by placing fruit in it, and when this fails, the back wall of the home cage is cranked forward so baboons have no choice but to enter the shuttle. In these extreme situations, baboons may experience some level of stress associated with the cage wall being cranked forward.
Three baboons (CY, DE, and SC) from the colony were identified as consistently required coaxing and/or cranking of the back wall in their home cage in order to transfer them in the shuttle for routine cage washes. Decreasing the time taken for these baboons to enter the shuttle was presumed to lower the stress levels experienced by these baboons when they require such transfer.
Exposure to the enrichment room began after pre-intervention data had been collected. Between 7 a.m. and 12 p.m. (i.e., prior to daily feedings), individual baboons were transported from their home cages to the enrichment room in the shuttle 2-3 times per week. Two dependent variables were used to measure the effectiveness of exposure to the enrichment room:
After 30 minutes, the shuttle was pushed up against the guillotine door to the room and the doors on the shuttle and the cage door were raised for transportation to the home cage. Baboons typically returned readily to the shuttle. If a baboon did not do so, a technician placed a piece of fruit in the shuttle to coax the baboon into it. As noted above, daily feeding (i.e., primate biscuits and fresh produce) was restricted to post-enrichment room participation, to increase the likelihood that baboons resisting entering the shuttle might be coaxed with the fruit.
Data Analysis: Data are presented for individual baboons. As subjects were also participants in on-going behavioral pharmacology studies, only data collected on days when no drugs were administered were included in this data analysis. As described above, a time-sampling procedure was used to assess the frequency of a specific behavior in the home cage for any given day (Martin and Pear, 1992). Thus, the total number of episodes across the eight 2-min. observations (one each hour for eight consecutive hours) that occurred in one 24-hr period was averaged across days for the periods before and after enrichment room exposure. In addition, the 2-min. observations were combined within each day by adding the frequency of occurrences of any given behavior and then dividing the total frequency by the number of total minutes spent observing in order to express a rate. In this way, the rate for each day served as an independent observation for comparing frequencies before and during exposure to the enrichment room using Student�s t-test (one-tailed). For the individual latencies to enter the transport shuttle, logarithmic transformation of individual latencies were analyzed using Student�s t-test (one-tailed). Changes in the mean frequency of a specific behavior, or the mean latency to enter the shuttle, were deemed statistically significant at the 0.05 level or better for individual baboons after exposure to the enrichment room.
The data are also presented in graph form as (1) the mean (±SD) frequency of each measure for each baboon prior to any exposure in the enrichment room (PRE) and (2) the frequency of that measure following each exposure to the room. In this way, patterns of change over time as a result of repeated exposure to the enrichment room may be detected.
Results
For baboon BB, repetitive pacing/circling in the home cage was identified as the target behavior prior to the start of this study. Using the time-sampling procedure described above, the mean (±SD) frequency of pacing/circling episodes in the home cage prior to any visits to the enrichment room was 2.19 (±1.39) (see Figure 3). After exposure to the enrichment room, the mean (±SD) number of pacing/circling episodes significantly decreased to 0.643 (±0.63) (t=3.948; p=0.0002).
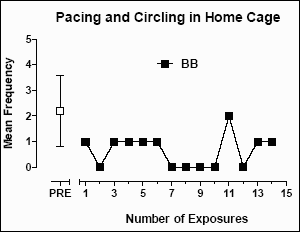
Figure 3: The mean frequency of pacing/circling episodes per two-minute interval in the home cage of baboon BB. Data here, and in Figures 4, 5, and 6, are the mean (±SD) frequency obtained using a time-sampling procedure before exposure to the enrichment room (PRE), and the frequency observed following exposure to the enrichment room.
For baboons DC and SY, a �huddled� posture was identified as a target behavior for which a decrease following exposure to the enrichment room may indicate an improvement in psychological well-being (see Figure 4). As noted previously, the �huddled� posture is defined as sitting with chin on chest and being unresponsive to normal stimuli. For baboon DC, the mean (±SD) frequency of the �huddled� posture prior to exposure to the enrichment room was 5.25 (±1.29). The mean frequency after exposure decreased to 4.21 (±1.31), a statistically significant decrease (t=2.02; p=0.03; see Figure 4). For baboon SY, the frequency of �huddled� posture observed in the home cage also decreased after exposure to the enrichment room. Prior to the first exposure to the enrichment room, the mean (±SD) frequency of a �huddled� posture for SY was 1.88 (±1.37). This was significantly decreased to 0.71 (±0.83) (t=2.92; p=0.003).
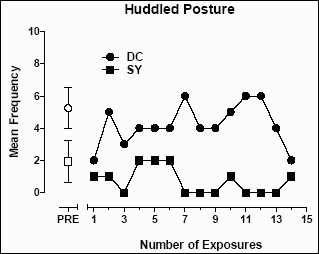
Figure 4: The mean frequency of �huddled� posture per two-minute interval in the home cages of baboons DC and SY.
Excessive grooming was also identified as a target behavior for baboon SY for which a decrease in the frequency may signal an improvement in his psychological well-being (see Figure 5). The mean (±SD) frequency of grooming episodes prior to exposure to the enrichment room was 3.77 (±1.45). The mean (±SD) frequency of grooming episodes after exposure decreased to 2.36 (±0.74), a statistically significant decrease (t=3.34; p=0.0008).
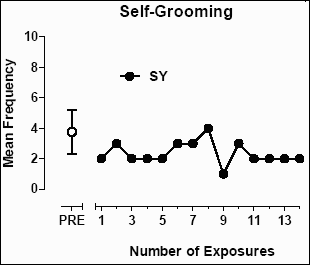
Figure 5: The mean frequency of grooming episodes per two-minute interval in the home cage of baboon SY.
Behaviors for which an increase in frequency might signal improved psychological well-being were also characterized. For baboon BB, there was a significant increase in the frequency of �playing with toys� in his home cage after exposure to the enrichment room (t=4.48; p<0.0001). As shown in Figure 6, the frequency since exposure to the enrichment room generally increased. Prior to enrichment room exposure, the mean (±SD) frequency of baboon BB �playing with toys� in the home cage was 2.27 (±1.91). After the first exposure, the mean (±SD) frequency was 4.9 (±1.54).
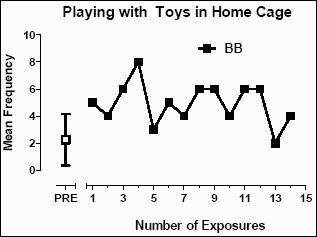
Figure 6: The mean frequency of episodes of �playing with toys� per two-minute interval in the home cage of baboon BB.
The latency to enter the shuttle for transportation from the home cage to the enrichment room was studied in three baboons (DE, CY, SC) known to require coaxing, and often cranking the back wall of the home cage, to get them into the shuttle. As shown in Figure 7, baboon DE required a mean (±SD) latency of 3.45 (±1.82) minutes to enter the shuttle prior to any exposure in the enrichment room. After the first exposure, the mean latency to enter the shuttle significantly decreased (t=7.4; p<0.0001). For baboon CY, the mean (±SD) latency to enter the shuttle prior to exposure was 0.53 ± 0.57 minutes, a significant decrease after exposure (t=2.1; p=0.023).
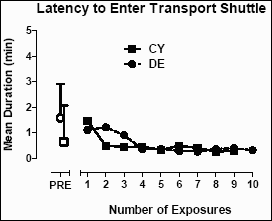
Figure 7: The mean (±SD) latency (min.) to enter the shuttle from the home cages of baboons DE and CY before (PRE) exposure to the enrichment room and after exposure. The maximum latency was five min., after which the back wall of the home cage was cranked forward.
Some of SY�s and DC�s behaviors in the home cage that may signal improvement in psychological well-being when increased (i.e., playing with toys, lip smacking, and grunting) were not significantly changed (data not shown) after exposure to the enrichment room.
Despite coaxing with fruit, one baboon (SC) failed to return to the shuttle after entering the enrichment room. Fortunately, the baboon was one who readily pressed his thigh up against the bars for ketamine injections in the home cage and also did so in the enrichment room. While ketamine is routinely used in lab settings for its sedative effects, it is a drug with potential for abuse, and studies have illustrated that nonhuman primates will self-administer ketamine (Lukas et al., 1984; Moreton et al., 1977). Thus, since intramuscular ketamine clearly served as a reinforcer for this baboon, the veterinary technician was able to sedate the animal with ketamine in order to remove him from the room. When this also needed to be done at the end of his second visit to the room, this baboon was dropped from the study.
Discussion
The objective of the present study was to improve the quality of life for baboons in our lab through exposure to an expanded environmental enrichment program. We could not simply assume, however, that exposure to the enrichment room would result in an improvement in the psychological well-being of our subjects. Thus, we identified three baboons with maladaptive behaviors in their home cages and compared the frequency of these behaviors prior to enrichment room exposure to the frequency of the same behaviors after exposure, and found significant decreases in their frequency.
In addition, we found that two baboons would more readily enter the transport shuttle after exposure to the enrichment room. That is, the stress resulting from cranking the back wall forward and �forcing� baboons out of the cage and into the shuttle for transport no longer occurs for these baboons, since they now readily enter the shuttle. This is important because baboons must be transported out of their home cages for regular cage washes.
Thus, our data support the idea that exposure to the enrichment room improved the psychological well-being of the baboons. In addition, while only one of the baboons showed an increase in the use of toys in his home cage, it is possible the baboons experienced significant increases in their psychological well-being that were undetected by our outcome measures. Moreover, by documenting the smaller objects manipulated (e.g., Kong® toys, balls made of different materials, plastic chains, mirrors) while the baboons were in the enrichment room, we also were able to identify individual toy preferences for individual baboons. This has resulted in more effective enrichment being provided in the home cages.
Another goal was simply to increase the amount of activity in which the baboons were able to engage. Technicians consistently noted in the records that baboons spend a majority of time in the enrichment room moving around and �exploring.�
A discussion of promoting the psychological well-being of nonhuman primates in laboratories would not be complete without mentioning possible causes of abnormal cage behaviors. While it is not possible to know why one baboon engages in maladaptive behaviors when others do not, one theory with an abundance of evidence asserts that removing an infant from his mother�s care too soon can result in the formation of abnormal cage behaviors later in life (Altmann, 2001; Bellanca & Crockett, 2002). Neither information about the age at which a baboon was removed from his mother�s care, nor descriptions of early life experiences, are routinely provided with nonhuman primates upon arrival at research facilities. For example, while the actual age of baboon BB is not known, he was a wild-caught baboon who weighed a mere 8.2 kg and was lacking his canine teeth upon arrival, leaving no doubt he arrived as a juvenile. Moreover, when he arrived at quarantine (and for a period of time after arriving in our facility), BB was housed with a second male baboon that was bigger and dominated BB. Thus, the cage behaviors exhibited by BB later in life may have been shaped as a consequence of being taken from his mother too soon and/or being caged with a dominant older male early in life. While many factors certainly influence the formation of maladaptive behaviors, housing and rearing conditions, early life experience, and colony procedures clearly play a role. Indeed, a retrospective analysis in a colony of rhesus macaques concluded that factors influencing the development of stereotypic and self-injurious behaviors in rhesus macaques included intrinsic factors (i.e., males exhibit more maladaptive behaviors than did females), rearing conditions, housing conditions, colony management practices, and research protocols (Lutz et al., 2003). Regardless of the cause of abnormal cage behaviors, it is important to examine how environmental enrichment may be useful for decreasing these behaviors.
Since we ended our data collection, the enrichment room has been available for all baboons in our colony, and we have encountered baboons that do not readily return to the shuttle for transportation back to their home cages. Specifically, we have had three instances, other than the one reported above, when baboons would not readily exit the enrichment room. In these instances, the technicians tried to coax the baboon into the shuttle by placing fruit in it, but found it required considerable time before the baboons would exit. That is, after a period of 1-4 hours baboons eventually returned to the shuttle, and those baboons are not currently visiting the enrichment room. Other methods for encouraging return to the shuttle, however, are being tried. For example, we have found that turning off the room light was successful with one baboon. Based on our experience during and after the present study, 30-45 minutes seems to be the ideal amount of time in the room after which the majority of baboons will readily exit the room without incident.
It is our hope that other laboratories will use an enrichment room for caged nonhuman primates and will find it a valuable tool for increasing their psychological well-being. Facilities that house very large numbers of nonhuman primates may find it difficult to expend the time, space, and money to offer such enrichment to all animals. While it would be wonderful to give all the animals in a lab access to an enrichment room, it would not be as expensive to at least provide it to those animals who need it most. Indeed, our data support the notion that the psychological well-being of nonhuman primates exhibiting maladaptive behaviors can improve by exposure to an enrichment room.
It should be noted, however, that some unique variables may have contributed to our success. The veterinary technicians in the Division of Behavioral Biology are responsible for the daily care of the same animals, and the baboons serve as subjects in behavioral experiments for a number of years (i.e., studies are not terminal). Thus, the baboons are extremely familiar with their veterinary technicians. In addition, the baboons have a history of shuttling for cage washes and so the act of moving from their home cages to the enrichment room was familiar to them. Nonetheless, exposing baboons with target behaviors indicative of poor psychological well-being to the enrichment room resulted in an apparent improvement in psychological well-being. The ease of replication of the enrichment room in other nonhuman primate colonies is contingent on the availability of space, time, and some financial support.
In conclusion, participation in the expanded enrichment program enhanced the standard enrichment provided in the home cages, and also appeared to improve the psychological well-being of individual baboons with identified maladaptive behaviors in our research program. We consider the time, effort, and expense of creating the enrichment room to have been well placed.
References
Altmann, J. (2001). Baboon mothers and infants. Chicago: University of Chicago Press
Ator, N. A., Weerts, E. M., Kaminski, B. J., Kautz, M. A., & Griffiths, R. R. (2000). Zaleplon and triazolam physical dependence assessed across increasing doses under a once-daily dosing regimen in baboons. Drug and Alcohol Dependence, 61, 69-84.
Bellanca, R. U., & Crockett, C. M. (2002). Factors predicting increased incidence of abnormal behavior in male pigtailed macaques. American Journal of Primatology, 58, 57-69.
Goodwin, A. K., Froestl, W., & Weerts, E. M. (2005). Involvement of gamma-hydroxybutyrate (GHB) and GABA-B receptors in the acute behavioral effects of GHB in baboons. Psychopharmacology, 180, 342-351.
Goodwin, A. K., Griffiths, R. R., Brown, P. R., Froestl, W., Jakobs, C., Gibson, K. M., & Weerts, E. M. (2006) Chronic intragastric administration of gamma-butyrolactone (GBL) produces physical dependence in baboons. Psychopharmacology, 189, 71-82.
Institute of Laboratory Animal Resources (1996). Guide for the care and use of laboratory animals. Washington, DC: National Academy Press.
Lukas, S. E., Griffiths, R. R., Brady, J. V., & Wurster, R. M. (1984). Phencyclidine-analogue self-injection by the baboon. Psychopharmacology, 83, 316-320.
Lutz, C., Well, A., & Novak, M. A. (2003). Stereotypic and self-injurious behavior in rhesus macaques: A survey and retrospective analysis of environment and early experience. American Journal of Primatology, 60, 1-15.
Martin, G., & Pear, J. (1992). Behavior modification: What it is and how to do it (4th ed.). New Jersey: Prentice Hall.
Moreton, J. E., Meisch, R. A., Stark, L., & Thompson, T. (1977). Ketamine self-administration by the rhesus monkey. Journal of Pharmacology and Experimental Therapeutics, 203, 303-309.
Sidman, M. (1960). Tactics of scientific research. New York: Basic Books.
-------------------------------------------
Amy K. Goodwin, Johns Hopkins Univ. Sch. of Med., Behavioral Biology Research Center, 5510 Nathan Shock Dr., Suite 3000, Baltimore, MD 21224 [410-550-2781; fax: 410-550-2780; e-mail: [email protected]].
The establishment of the enrichment room was made possible through an Animal Welfare Enhancement Award made by the Johns Hopkins University Center for Alternatives to Animal Testing. The authors would also like to thank Dan Rodgerson for the technical expertise he lent to the project, and Robert J. Adams, DVM, for helpful comments during the protocol review process and manuscript preparation.
-------------------------------------------
* * *
Workshop Announcements: Association of Primate Veterinarians
The Association of Primate Veterinarians� (APV) 37th Annual Workshop will be held November 5-7, 2009, in Denver, Colorado. The deadline for submitting case reports and/or �What�s Your Diagnosis?� for the Workshop is August 1, 2009. Please keep length of abstract to one page. If you have time constraints and cannot submit your abstract by this date, please submit your intent to present, presentation title, and author(s). Clearly identify the presenting author and provide his or her CV by the August 1 deadline date. For those requiring additional time, full abstract submittals will be required no later than August 15, 2009. Electronic versions (PowerPoint, video) of the case report and/or �What�s Your Diagnosis?� are due no later than October 1, 2009. Electronic abstracts of case reports can be submitted online through the APV Website: <www.primatevets.org>. Please contact Dr. Susanne Rensing [703-245-2200; e-mail: [email protected]] or Dr. Rick Rockar [e-mail: [email protected]t] if you have any questions about case reports.
APV is an international organization consisting of over 400 veterinarians concerned with the health, care and welfare of nonhuman primates. If you are not an APV member and are interested in membership, applications are available online at <www.primatevets.org>. Additional information concerning the Workshop can be found on that same Website.
* * *
Female Homosexual Behavior in a New Group of Japanese Macaques:
Evolutionary Implications
Paul L. Vasey(1) and Christine Reinhart(2)
(1)Dept of Psychology, University of Lethbridge; (2)Dept of Neuroscience, University of Lethbridge
Introduction
Many individuals who work with Japanese macaques (Macaca fuscata) may be unaware that in certain captive and free-ranging groups, females of this species routinely engage in same-sex courtship and series mounting (1) with pelvic thrusting. They do so during temporary, but exclusive, relationships termed �consortships� (Wolfe, 1984). Females sometimes compete with males for same-sex sexual partners and will often choose same-sex sexual partners even when given the simultaneous choice of a sexually motivated male alternative (Vasey, 1998). While it is true that female-female sexual activity increases in frequency as the number of males in the population decreases, such increases likely reflect the fact that there are fewer male sexual competitors under these demographic conditions and more preferred female sexual partners, relative to preferred male sexual partners (Vasey & Gauthier, 2000). Females that engage in these behaviors do not appear to be �masculinized� in terms of their brains (Vasey & Pfaus, 2005) or other aspects of their social (Vasey et al., 2008b) and sexual behavior (Vasey, 2004a; Vasey et al., 2006, 2008a). Same-sex sexual mounting, courtship, and consortship activity in Japanese macaques does not appear to serve any sociosexual function including: attracting male mates, alliance formation, dominance demonstration, acquisition of additional care for offspring, reconciliation, practice for heterosexual copulation, and inter-individual tension reduction (Gouzoules & Goy, 1983; Vasey, 1996, 1998, 2004b, 2006; Vasey et al., 1998).
Several lines of evidence indicate that female-female mounting, courtship, and consortship activity in Japanese macaques are sexually motivated. First, these interactions mirror male-female sexual behavior in many aspects of their expression. For example, the courtship behavior that occurs during homosexual consortships is virtually indistinguishable from that which occurs during heterosexual consortships (Vasey et al., 2008a). Similarly, female-female mounting across the ovarian cycle parallels the pattern found for male-female mounting (O�Neill et al., 2004). Also, females exercise incest avoidance with close female kin (Chapais et al., 1997), despite the fact that these same kin dyads engage in various forms of social affiliation together, such as grooming (Baxter & Fedigan, 1979). Finally, during most same-sex mounts (78.3%), female mounters engage in vulvar, perineal, and anal stimulation (Vasey & Duckworth, 2006).
Here, we report on female homosexual behavior in a new group of Japanese macaques. We discuss the implication of these observations for the proposition, put forward by Vasey & Jiskoot (in press), that female homosexual behavior in Japanese macaques may be associated with genetically distinct free-ranging populations of Japanese macaques on the island of Honshu.
Methods
Fortuitous observations were conducted during November, 2007, at the Primate Research Institute of Kyoto University in Inuyama, Japan. A total of 50 minutes of continuous videotape data were collected on two female Japanese macaques in the Wakasa-B captive, mixed-sex group of Japanese macaques that contained three adult males and 15 adult females. Data were collected on both females simultaneously because they were engaged in series mounting during a homosexual consortship and, as such, they were almost always interacting with each other.
Results
Two adult females were observed to engage in series mounting over a period of 50 minutes. Approximately one mount occurred per minute of observation (n = 52 mounts). Overall, 77% of the observed mounts took the form of ventral-ventral mounting. The remaining mounts (23%) were all sitting mounts during which the mounter sat in a jockey-style position on the back of the mountee. Not all mounts involved pelvic thrusting by the mounter (mean = 0.38 pelvic thrusts/mount). Pelvic thrusting occurred more frequently during sitting mounts (70% of sitting mounts), than during ventral-ventral mounts (30% of ventral-ventral mounts). Both females mounted and were mounted during the observation period; however, one partner adopted the role of mounter during 79% of the interactions.
During inter-mount intervals, females solicited each other using a variety of courtship behaviors including sexual vocalizations, grasping, hands-on-hindquarter solicitations, and body trembling (for a description of individual courtship behaviors see Vasey et al., 2008a). One female vigorously performed sexual solicitations during every observation minute (n = 52). Typically, she would solicit her partner by grasping the partner�s arm, then reclining on her back, pulling the partner onto her ventral surface while vocalizing. This same female performed the greatest number of sexual solicitations, but the least number of mounts (21% of mounts). In contrast, her partner performed sexual solicitations during only six of the 50 observation minutes.
The two consorting females remained either in contact with each other or within arm�s reach during 90% of inter-mount intervals. During 12% of inter-mount intervals, the female that performed the greatest number of sexual solicitations groomed her partner. In contrast, her partner was not observed to groom her.
Twice during the observation period, a third-party female approached and solicited one of the partners who sat alone, while directing an open-mouthed stare threat at the other partner, who sat at a distance and looked away in response to the threat. Solicitations by this third-party female were ignored and the consort partners re-approached each other shortly thereafter and continued their sexual activity.
Discussion
Our observations of female-female series mounting, courtship, and consortship activity in the Wakasa-B group furnishes evidence for the existence of these behaviors in a new group of Japanese macaques. The focal females were observed to engage in frequent mounting and courtship behavior over the course of the entire observation period. Whether a more widespread pattern of female homosexual behavior exists in the captive Wakasa-B colony, and in the wild population from which it was derived, remains an open question. More research will obviously be needed to ascertain whether this is indeed the case.
The focal females employed the ventral-ventral mounting posture at high rates. In contrast, ventral-ventral mounting between females occurs very infrequently in the free-ranging Arashiyama population (Vasey & Duck-worth, 2006) and in the captive Universit� de Montr�al population (Vasey et al., 1998) of Japanese macaques. Similarly, Wolfe (1978) reported that ventral-ventral mounting between females was observed only �occasionally� in the semi-free-ranging Texas (Arashiyama-West) population. Wolfe (1978) also mentioned that when females at her site engaged in ventral-ventral mounting they would rub their perinea against one another, but our focal females did not execute this type of pelvic thrusting. Inter-group variation in female-female mounting behavior (i.e., posture, thrusting) may reflect group-specific, non-adaptive sexual �traditions�.
During the observation period, a third-party female was twice seen to intrude on the homosexual consortship and solicit one of the female partners, while threatening the other. In light of the brief observation period (50 min.), the occurrence of such competitive interactions is striking, given that intrusion by third-party females on female homosexual consortships are extremely rare in the Universit� de Montr�al colony of Japanese macaques (mean = 0.009 intrusions/hr; n = 129 observation hours; Vasey, 2004a). Once again, this sort of inter-group behavioral variation might reflect group-specific differences in non-adaptive sexual �traditions�, which, in this case, may be expressed in the domain of same-sex mate acquisition strategies.
On a broader scale, this case study has implications for understanding the evolution and biogeography of female homosexual consortships in Japanese macaques. Recent mitochondrial DNA research on Japanese macaques indicates genetic divergence into five haplogroups, all of which occur on the island of Honshu (Kawamoto et al., 2007). All of the free-ranging Japanese macaque populations in which female homosexual consortships have been reported fall within a single haplogroup (A1), which is exclusive to Honshu (Vasey & Jiskoot, in press). The original members of the Wakasa-B group were drawn from the wild population in Yazu District, Tottori Prefecture of the Chugoku region of Honshu. As such, members of this group also fall within the A1 haplogroup (location 96 Wakasa, haplotype JN36; Kawamoto et al., 2007). Consequently, our observations of female homosexual consortship interactions in the Wakasa-B group lend further support to the suggestion by Vasey & Jiskoot (in press) that female-female consortship activity in this species may be associated with genetic divergence of the A1 haplogroup on the island of Honshu. As such, we anticipate that the distinctive pattern of female homosexual behavior described here (i.e., same-sex series mounting and courtship during consortships) will characterize A1 haplogroup populations, but will not be ubiquitous in Japanese macaques.
References
Baxter, M. J., & Fedigan, L. M. (1979). Grooming and consort partner selection in a troop of Japanese monkeys (Macaca fuscata). Archives of Sexual Behavior, 8, 445-458.
Chapais, B., Gauthier, C., Prud�homme, J., & Vasey, P. L. (1997). Relatedness threshold for nepotism in Japanese macaques. Animal Behaviour, 53, 533-548.
Gouzoules, H., & Goy, R. W. (1983). Physiological and social influences on mounting behavior of troop-living female monkeys (Macaca fuscata). American Journal of Primatology, 5, 39-49.
Kawamoto, Y., Shotake, T., Nozawa, K., Kawamoto, S., Tomari, K., Kawai, S., Shirai, K., Morimitsu, Y., Takagi, N., Akaza, H., Fujii, H., Hagihara, K., Aizawa, K., Akachi, S., Oi, T., & Hayaishi, S. (2007). Postglacial population expansion of Japanese macaque (Macaca fuscata) inferred from mitochondrial DNA phylogeography. Primates, 48, 27-40.
O�Neill, A. C., Fedigan, L. M., & Ziegler, T. E. (2004). Ovarian cycle phase and same-sex mating behavior in Japanese macaque females. American Journal of Primatology, 63, 25-31.
Vasey, P. L. (1996). Interventions and alliance formation between female Japanese macaques, Macaca fuscata, during homosexual consortships. Animal Behaviour, 52, 539-551.
Vasey, P. L. (1998). Female choice and inter-sexual competition for female sexual partners in Japanese macaques. Behaviour 135, 579-597.
Vasey, P. L. (2004a). Sex differences in sexual partner acquisition, retention and harassment during female homosexual consortships in Japanese macaques. American Journal of Primatology, 64, 397-409.
Vasey, P. L. (2004b). Pre- and post-conflict interactions between female Japanese macaques during homosexual consortships. International Journal of Comparative Psychology, 17, 351-359.
Vasey, P. L. (2006). The pursuit of pleasure: Homosexual behavior, sexual reward and evolutionary history in Japanese macaques. In V. Sommer & P. L. Vasey (Eds.), Homosexual behavior in animals: An evolutionary perspective (pp. 191-219). Cambridge: Cambridge University Press.
Vasey, P. L., & Duckworth, N. (2006) Sexual reward via vulvar, perineal and anal stimulation: A proximate mechanism for female homosexual mounting in Japanese macaques. Archives of Sexual Behavior, 35, 523-532.
Vasey, P. L., & Gauthier, C. (2000). Skewed sex ratios and female homosexual activity in Japanese macaques: An experimental analysis. Primates, 41, 17-25.
Vasey, P. L., & Jiskoot, H. (in press). The biogeography and evolution of female homosexual behavior in Japanese macaques. Archives of Sexual Behavior.
Vasey, P. L., & Pfaus, J. (2005). A sexually dimorphic hypothalamic nucleus in a macaque species with frequent female-female mounting and same-sex sexual partner preference. Behavioral Brain Research, 157, 265-272.
Vasey, P. L., Chapais, B., & Gauthier, C. (1998). Mounting interactions between female Japanese macaques: Testing the influence of dominance and aggression. Ethology, 104, 387-398. Vasey, P. L., Foroud, A., Duckworth, N., & Kovacovsky, S. D. (2006). Male-female and female-female mounting in Japanese macaques: A comparative analysis of posture and movement. Archives of Sexual Behavior, 35, 116-128.
Vasey, P. L., Rains, D., VanderLaan, D. P., Duckworth, N., & Kovacovsky, S. D. (2008a). Courtship behavior during heterosexual and homosexual consortships in Japanese macaques. Behavioral Processes, 78, 401-407.
Vasey P. L., VanderLaan D. P., Rains D., Duckworth N., & Kovacovsky S. D. (2008b). Inter-mount social interactions during heterosexual and homosexual consortships in Japanese macaques. Ethology, 114, 564-574.
Wolfe L. D. (1984). Japanese macaque female sexual behavior: A comparison of Arashiyama East and West. In M. F. Small (Ed.), Female primates: Studies by women primatologists (pp. 141-157). New York: Alan R. Liss.
Wolfe, L. D. (1978). Age and sexual behavior of Japanese macaques (Macaca fuscata). Archvies of Sexual Behavior, 7, 55-68.
-------------------------------------------
Corresponding author: P. L. Vasey, Dept of Psychology, Univ. of Lethbridge, Lethbridge, Alberta, T1K 3M4, Canada [e-mail: [email protected]].
We thank Eiji Enomoto, Mike Huffman, Jean-Baptiste Leca, Sergio Pellis, and the staff of the Primate Research Institute in Inuyama, Japan.
P. L. V.�s research was supported by the University of Lethbridge and a Natural Sciences and Engineering Research Council (NSERC) of Canada Discovery Grant. C. R. was supported by a NSERC Post-Graduate Scholarship and the Alberta Ingenuity Fund.
-------------------------------------------
(1) Series mounting occurs when consort partners engaged in three or more mounts within a 10-minute period. Series mounting (and consortship activity) is deemed to have terminated when consort partners are separated by a distance of more than one meter and there has been no mounting for more than 10 minutes.
-------------------------------------------
* * *
Looking for Information and Possible Collaboration
Ginger Tansey
National Eye Institute, National Institutes of Health
We have identified an adult female cynomolgus macaque (Macaca fascicularis), of Chinese origin, with an interesting retinal lesion. We would like to determine whether this is a genetic condition, in which case it could possibly become a new model for human macular degeneration, which is naturally of great importance to us. In order to do so, we�d like to find other monkeys with the same condition. We expect that this will not be easy.
If you have a cynomolgus macaque of Chinese origin, especially one with alopecia of the head, neck, shoulders and extending onto the body, and particularly with alopecia that began early in life, then the next step is to have someone assess the retina. It is the alopecia that makes us think that the retinal lesion is a genetic condition in our index animal.
Macaques have a retina that looks similar to the retina in humans, so a medical ophthalmologist would be able to identify any interesting lesions right away. In our case, we are interested in lesions of the fundus. If you or your ophthalmologist can identify a potential patient, we�d like to discuss it with you. The next steps would include getting blood and/or tissue samples from your animals, to compare with those of our index animal.
Ultimately, our hope is to identify other monkeys with the same condition to (1) find the gene or genes of interest and (2) develop a breeding colony of macaques with this condition, for gene therapy. The end result could possibly become a treatment for macular degeneration in humans, which would lead to millions of people around the world being spared blindness.
If you think you have a potential candidate for assessment, or have any questions about potential candidates, please contact me at the National Eye Institute, 49/2C72, MSC 4435, 9000 Rockville Pike, Bethesda MD 20892 [301-594-1225; e-mail: [email protected]].
* * *
Information Requested or Available
Gorilla Haven Update
A new update has been posted at <www.gorilla-haven.org> about Joe�s health and Oliver�s future, as well as the future of Gorilla Haven itself...
Caging Survey
Viktor Reinhardt is making an informal survey of the rate of caged macaques living alone in zoos, laboratories, and other places. If you have access to this information for your facility, please indicate:
More Interesting Websites
* * *
Meeting Announcements
The First African Symposium on Zoological Medicine will be held July 18-19th, 2009, at the Johannesburg Zoo, South Africa. Financial assistance is available for veterinarians from other African countries. For more details contact Teresa Slacke at <[email protected]>.
The 2009 Marmoset Research Group of the Americas (MaRGA) Meeting will take place July 29-31, 2009, in Natal, Brazil, at the Universidade Federal do Rio Grande do Norte. For complete information, see <www.cb.ufrn.br/~fisiologia/marga>.
The Laboratory Animal Welfare Training Exchange�s (LAWTE) Biennial Conference will take place in San Antonio, Texas, August 5-7, 2007. Topics to be included are educating researchers on reducing pain and distress in laboratory animals, expectations for animal care training programs, and animal rights/security updates. See the flyer at <www.lawte.org/2009_conference_flyer.pdf> for details.
The 7th World Congress on Alternatives and Animal Use in the Life Sciences will take place August 30 to September 3, 2009, in Rome, Italy. The Congress was given the motto �Calling on Science� in order to emphasize that scientific progress today goes hand in hand with progress towards the reduction, refinement, and replacement of experimental animals (the 3R�s). For complete information, see <www.aimgroup.eu/2009/WC7>.
The Third Orangutan SSP Husbandry Workshop will be held August 31 through September 2, 2009, at Zoo Atlanta, Georgia. See <www.2009orangutanworkshop.org>.
The Association of Zoos & Aquariums� (AZA) 2009 Annual Conference will be held September 12-17, in Portland, Oregon, hosted by the Oregon Zoo. See <aza.org/ConfWork/AC_Intro/index.html>.
The 26th European Association of Zoos and Aquaria Annual Conference will be held September 14-20, 2009, in Copenhagen, Denmark, hosted by the Copenhagen Zoo. See <www.eaza.net/news/frameset_1news.html?page=1calendar> and click on �Conference�.
The 3rd International Congress on Zoo Keeping and the 36th American Association of Zoo Keepers (AAKZ) National Conference will be held September 24-29, 2009 in Seattle, Washington, hosted by the Puget Sound Chapter of AAZK and Woodland Park Zoo. See <www.iczoo.org> and <www.pugetsoundaazk.org>.
The 27th Annual Symposium on Nonhuman Primate Models for AIDS will be held October 28-31, 2009, in Boston, Massachusetts, sponsored by the New England Primate Research Center. For information, see <nhp2009.hms.harvard.edu>.
On October 29, 2009, the Bristol Conservation and Science Foundation (BCSF) and Bristol Zoo Gardens will present the 2nd Annual BCSF Symposium: How Can We Save Primates from Extinction? The one-day symposium will be held in the Clifton Pavilion at Bristol Zoo Gardens, Bristol, U.K., from 10 a.m. to 5.30 p.m. Registration fees are �75 per person and include a buffet-style lunch as well as coffee/tea breaks between the sessions and entry to Bristol Zoo Gardens. For details, see <www.bristolzoo.org.uk/about/conservation/symposium2009>.
The 6th European Zoo Nutrition Conference will be held in Barcelona, Spain, January 28-31, 2010. Please send comments or suggestions for topics and speakers directly to <[email protected]>. Further announcements and information will be posted online at the nutrition area of the European Association of Zoos and Aquaria�s Website, <www.eaza.net>.
The 2010 American Veterinary Medical Association (AVMA) National Meeting will be held in Atlanta, Georgia, July 31th to August 3rd, 2010. See <www.avmaconvention.org>.
* * *
Resources Wanted and Available
Management Innovations in Laboratory Animal Care
Steven M. Niemi, Director of the Center for Comparative Medicine at Massachusetts General Hospital, has introduced a new Website, <www.virtualvivarium.com>, dedicated to management innovations in laboratory animal care.
There are four subject categories available on the home page:
�We look forward to your questions and suggestions, via e-mails to <[email protected]>.�
Preventing Disease from Animals in Public Settings
Health risks such as infectious diseases, exposure to rabies, and injuries are associated with contact with animals in public settings, including county or state fairs, petting zoos, animal swap meets, pet stores, zoologic institutions, circuses, carnivals, educational farms, livestock-birthing exhibits, educational exhibits at schools and child-care facilities, and wildlife photo opportunities. Washing hands is the most important prevention step to reduce the risk for disease transmission associated with animals in public settings. Other recommendations include prohibiting food in animal areas, including transition areas between animal areas and nonanimal areas, providing information about disease risk and prevention to visitors, and proper care of animals.
The National Association of State Public Health Veterinarians, Inc., has a series of recommendations on-line: <www.cdc.gov/mmwr/preview/mmwrhtml/rr5805a1.htm?s_cid=rr5805a1_e> is the basic document. Appendices are:
Thanks to long-time IPPL member Dr. Mich Kabay, all past issues of IPPL News � from 1974 to the present � are now on-line at <www.ippl.org>. There are now over 100 issues accessible. To visit the archive, click on �IPPL in Action� and then on �Archived Newsletters since 1974�. You can enter a word search to locate articles or you can just browse.
* * *
Travelers� Health Notes: IAMAT
The International Association for Medical Assistance to Travelers (IAMAT), a volunteer group, compiles an annual list of doctors around the world who meet the organization�s criteria, who speak English or another second language, and who agree to charge a specific fee. The 2009 Directory lists the current schedule of fees as US$100 for an office visit, US$150 for a house (or hotel) call, and US$170 for night, Sunday, and local holiday calls. These fees do not include consultants, laboratory or surgical procedures, hospitalization, or other expenses. The current listing of doctors and centers includes 93 countries, plus a listing of mental health resources in 15 countries.
IAMAT also publishes and provides to its members pamphlets on immunization and malaria, as well as �World Climate Charts� and a �Traveler Clinical Record� form. IAMAT has a scholarship program for physicians from developing countries to attend travel medicine training courses in North America, and offers lectureship grants to international medical societies, which allow visiting travel and tropical medicine scholars to exchange information and research on the prevention and treatment of tropical diseases.
For information, contact IAMAT, 1287 St. Clair Ave West, Toronto, Ontario M6E 1B8, Canada; 1623 Military Rd, #279, Niagara Falls, NY 14304-1745, U.S.A. [716-754-4883]; 206 Papanui Rd, Christchurch 5, New Zealand; or [e-mail: [email protected]]; or see <www.iamat.org>.
* * *
News Briefs
New Orangutan Population Found in Indonesia
JAKARTA, Indonesia � Conservationists have discovered a new population of orangutans in a remote, mountainous corner of Indonesia � perhaps as many as 2,000 � giving a rare boost to one of the world�s most endangered great apes. A team surveying forests nestled between jagged limestone cliffs on the eastern edge of Borneo counted 219 orangutan nests, indicating a �substantial� number of the animals, said Erik Meijaard, a senior ecologist at the U.S.-based Nature Conservancy. �We can�t say for sure how many,� he said, but even the most cautious estimate would indicate �several hundred at least, maybe 1,000 or 2,000 even.�
There are an estimated 50,000 to 60,000 orangutans left in the wild, 90 percent of them in Indonesia and the rest in neighboring Malaysia. The countries are the world�s top producers of palm oil, used in food, cosmetics and to meet growing demands for �clean-burning� fuels in the U.S. and Europe. Rain forests, where the solitary animals spend almost all of their time, have been clear-cut and burned at alarming rates to make way for lucrative palm oil plantations. The steep topography, poor soil and general inaccessibility of the rugged limestone mountains appear to have shielded the area from development, at least for now, said Meijaard. Its trees include those highly sought after for commercial timber.
The 700-square-mile (2,500-square-kilometer) jungle escaped the massive fires that devastated almost all of the surrounding forests in the late 1990s. The blazes were set by plantation owners and small-scale farmers and exacerbated by the El Nino droughts.
Nardiyono, who headed The Nature Conservancy�s weeklong survey in December, said �it could be the density is very high because after the fires, the orangutans all flocked to one small area.�
Conservationists say the most immediate next step will be working with local authorities to protect the area and others that fall outside of national parks. A previously undiscovered population of several hundred also was found recently on Sumatra, home to around 7,000.
�That we are still finding new populations indicates that we still have a chance to save this animal,� said Paul Hartman, who heads the U.S.-funded Orangutan Conservation Service Program, adding it�s not all �gloom and doom.�
Noviar Andayani, head of the Indonesian Primate Association and Orangutan Forum, said the new discoveries point to how much work still needs to be done to come up with accurate population assessments, considered vital to determining a species� vulnerability to extinction.
�There are many areas that still have not been surveyed,� she said, adding that 18 private conservation groups have just started work on an in-depth census based on interviews with people who spend time in the forests. They include villagers and those working on plantations or within logging concessions.
�We hope this will help fill in a few more gaps,� said Andayani, adding that preliminary tests in areas where populations are known indicate that the new interview-based technique could provide a clearer picture than nest tallies. �Right now the information and data we have about orangutans is still pretty rudimentary.�
Some experts say at the current rate of habitat destruction, the animals could be wiped out within the next two decades. � By Associated Press Writer Robin Mcdowell �April 12
Health Care for Mountain Gorillas and Humans
The University of California, Davis, has launched a new �One Health� program to conserve the world�s remaining 740 mountain gorillas by caring for, not only the gorillas, but also the people and the other animals that share their home in the forests of central Africa. With $750,000 in funding from the David and Lucile Packard Foundation, UC Davis will establish the Mountain Gorilla One Health Program in the School of Veterinary Medicine�s Wildlife Health Center.
The new program will partner with the existing Mountain Gorilla Veterinary Project, begun in 1986, to continue improving gorilla health and survival by addressing human health, livestock health and agricultural issues.
�The concept of �One Health� � that human, animal, and environmental health are inextricably linked and should be considered holistically � is a core principle of the UC Davis Wildlife Health Center,� said wildlife veterinarian and Center assistant director Kirsten Gilardi, who will lead the Program. �We are proud to become partners with the Mountain Gorilla Veterinary Project, one of the few true, on-the-ground, examples of One Health in action anywhere in the world.�
The Mountain Gorilla Veterinary Project�s longtime director, veterinarian Michael Cranfield, will join the UC Davis staff while continuing to oversee the work of the project�s seven veterinarians and 12 technicians and staff members in Rwanda, Uganda, and the Democratic Republic of the Congo.
UC Davis will investigate disease threats facing mountain gorillas, help expand medical care for the humans working in and around the gorilla parks, and improve the health and well-being of livestock to benefit the families who depend on them for nutrition and income.
Combined with anti-poaching patrols and habitat-protection efforts of the Rwandan, Ugandan and Congolese governments and other organizations, the Mountain Gorilla Veterinary Project�s medical program has helped increase the number of mountain gorillas by 17 percent in the past 10 years, making the mountain gorilla the only wild great ape whose numbers are rising, not falling. � from an April 23, 2009, press release
Carole C. Noon
Fort Pierce, Florida � Dr. Carole C. Noon, 59, the founder and director of Save the Chimps Sanctuary in St. Lucie County, died May 2, 2009, after a battle with pancreatic cancer.
Dr. Noon founded Save the Chimps in 1997 to provide a sanctuary for chimpanzees that had been used in the early space program and in laboratory testing. The 150-acre facility in western St. Lucie County is now home to more than 160 chimpanzees. A guestbook may be signed at <www.haisleyfuneralhome.com>. � from <TCPalm.com>.
Use of Experimental Primates Shown
Oxford University has revealed the number of primates it uses for animal experiments, after being ordered to by the Information Commissioner. The university had refused to release the figures in response to a Freedom of Information request, saying it would put researchers at risk.
In 2008 the university held 86 primates with 66 experimented upon, compared to 99 held and 40 experimented on in 2007. The primates used are macaques. The British Union for the Abolition of Vivisection (BUAV) had requested the details.
Michelle Thew, Chief Executive of the BUAV said: �For us one primate being used is too many. We do welcome the release of this information, but the key question for us is why the university fought so hard and so long not to have this basic information made public. We wanted more information about what actually happens to the primates.�
The university said it had refused the request for information, made in 2006, partly on the grounds that it would endanger the safety of its researchers. In April the Information Commissioner, Richard Thomas, ruled that releasing the information would not increase the risk to the health or safety of any person. � BBC News, May 5
�Ingenious� Ape Mounts Bid to Escape Zoo
A zoo in Australia was evacuated Sunday after an �ingenious� 137-pound orangutan short-circuited an electric fence and hopped a wall surrounding her enclosure. The ape, a 27-year-old female named Karta, jammed a stick into wires connected to the fence and then piled up debris to climb a concrete and glass wall at the Adelaide Zoo. Zoo curator Peter Whitehead told reporters Karta sat on top of the fence for about 30 minutes before apparently changing her mind about the escape and climbing back into the enclosure.
�I think when she actually got out and realized where she was ... she realized she shouldn�t be there so then she actually hung onto the wall and dropped back into the exhibit,� Whitehead said. Karta came within a few yards of visitors, who were the first to notice the animal�s escape bid.
Whitehead said the animal was not aggressive, but the zoo was cleared as a precaution, and veterinarians stood by with tranquilizer guns in case of trouble. � Associated Press, May 10
Primate Research Center Under Scrutiny
The University of Louisiana at Lafayette�s New Iberia Research Center was cited for violations of the Animal Welfare Act. An inspection found several areas in which the facility failed to comply with the Animal Welfare Act, which requires commercial animal businesses and research institutions to be licensed or registered with the government and inspected.
Conducted in March by the U.S. Department of Agriculture�s (USDA) Animal and Plant Health Inspection Service (APHIS), the inspection was in response to a complaint filed by the Humane Society of the United States about the way the facility treated its primates. APHIS inspected all areas of care and treatment, citing the facility for issues involving its handling and environmental enhancement of primates.
The inspectors also found problems with New Iberia�s Institutional Animal Care and Use Committee (IACUC), which reviews and approves research protocols. APHIS documented several areas in which IACUC was non-compliant, including incomplete descriptions of protocols and insufficient consideration of alternatives to procedures that may cause distress. APHIS says officials will work with the facility to ensure that corrective actions are taken.
The complete inspection report on USDA�s findings at New Iberia can be found on the APHIS Website, <www.aphis.usda.gov/animal_welfare/efoia>. � DVM Newsmagazine, May 12
* * *
Recent Books and Articles
(Addresses are those of first authors unless otherwise indicated)
Handbooks
* Guideline for Disinfection and Sterilization in Healthcare Facilities, 2008. Rutala, W. A., Weber, D. J., & the Healthcare Infection Control Practices Advisory Committee. Centers for Disease Control, <www.cdc.gov/ncidod/dhqp/pdf/guidelines/Disinfection_Nov_2008.pdf>
. . . �This guideline discusses use of products by healthcare personnel in healthcare settings such as hospitals, ambulatory care and home care; the recommendations are not intended for consumer use of the products discussed.� (They will be of interest to those involved in animal health care, however.)
Magazines and Newsletters
* Animal Behaviour, 2008, 75 [3], <www. sciencedirect.com>.
. . .Contents include: A comparative study of conflict resolution in macaques: Insights into the nature of trait covariation, by B. Thierry, F. Aureli, C. L. Nunn, O. Petit, C. Abegg, & F. B.M. de Waal; The sensory basis of prey detection in captive-born grey mouse lemurs, Microcebus murinus, by M. Piep, U. Radespiel, E. Zimmermann, S. Schmidt, & B. M. Siemers; Sharing the motivation to play: The use of signals in adult bonobos, by E. Palagi; Mating patterns and sexual swellings in pair-living and multimale groups of wild white-handed gibbons, Hylobates lar by C. Barelli, M. Heistermann, C. Boesch, & U. H. Reichard; Precision and power in the analysis of social structure using associations, by H. Whitehead; Dads do not pay for sex but do buy the milk: Food sharing and reproduction in owl monkeys (Aotus spp.), by C. K. Wolovich, S. Evans, & J. A. French; and Can�t get there from here: Inferring kinship from pairwise genetic relatedness, by R. C. Van Horn, J. Altmann, & S. C. Alberts.
* Animal Behaviour, 2008, 75[4], <www. sciencedirect.com>.
. . .Contents include: Revisiting the definition of animal tool use, by R. St Amant & T. E. Horton.
* Animal Behaviour, 2008, 75[5]. <www. sciencedirect.com>.
. . .Contents include: Gaze following in monkeys is modulated by observed facial expressions, by B. M. A. Goossens, M. Dekleva, S. M. Reader, E. H. M. Sterck, & J. J. Bolhuis; Late pregnancy glucocorticoid levels predict responsiveness in wild baboon mothers (Papio cynocephalus), by N. Nguyen, L. R. Gesquiere, E. O. Wango, S. C. Alberts, & J. Altmann; Chimpanzees do not take advantage of very low cost opportunities to deliver food to unrelated group members, by J. Vonk, S. F. Brosnan, J. B. Silk, J. Henrich, A. S. Richardson, S. P. Lambeth, S. J. Schapiro, & D. J. Povinelli; Perception of food amounts by chimpanzees based on the number, size, contour length and visibility of items, by M. J. Beran, T. A. Evans, & E. H. Harris; and Incorporating uncertainty into the study of animal social networks, by D. Lusseau, H. Whitehead, & S. Gero.
* Animal Behaviour, 2008, 75[6], <www. sciencedirect.com>
. . .Contents include: The evolution of teaching, by A. Thornton & N. J. Raihani; Male Barbary macaques eavesdrop on mating outcome: A playback study, by D. Pfefferle, M. Heistermann, J. K. Hodges, & J. Fischer; Punishment and competition over food in captive rhesus macaques, Macaca mulatta, by R. L. Chancellor & L. A. Isbell; and To follow or not to follow: Decision making and leadership during the morning departure in chacma baboons, by S. Stueckle & D. Zinner.
* American Journal of Primatology, 2009, 71[4], <www3.interscience.wiley.com/journal/ 122210925/issue>.
. . .Contents: Reproductive aging in captive and wild common chimpanzees: Factors influencing the rate of follicular depletion, by S. Atsalis & E. Videan; Assessing chimpanzee personality and subjective well-being in Japan, by A. Weiss, M. Inoue-Murayama, K.-W. Hong, E. Inoue, T. Udono, T. Ochiai, T. Matsuzawa, S. Hirata, & J. E. King; Living on the edge: Life-history of olive baboons at Gashaka-Gumti National Park, Nigeria, by J. P. Higham, Y. Warren, J. Adanu, B. N. Umaru, A. M. MacLarnon, V. Sommer, & C. Ross; Quantitative variability of cyanogenesis in Cathariostachys madagascariensis � the main food plant of bamboo lemurs in Southeastern Madagascar, by D. J. Ballhorn, S. Kautz, & F. P. Rakotoarivelo; Captive cotton-top tamarins� (Saguinus oedipus oedipus) use of landmarks to localize hidden food items, by F. L. Dolins; Production and perception of sex differences in vocalizations of Wied�s black-tufted-ear marmosets (Callithrix kuhlii), by A. S. Smith, A. K. Birnie, K. R. Lane, & J. A. French; Dialects in pygmy marmosets? Population variation in call structure, by S. de la Torre & C. T. Snowdon; Visual acuity in the cathemeral strepsirrhine Eulemur macaco flavifrons, by C. C. Veilleux & E. C. Kirk; Methods for calculating activity budgets compared: A case study using orangutans, by M. E. Harrison, E. R. Vogel, H. C. Morrogh-Bernard, & M. A. van Noordwijk; and Noninvasive collection of fresh hairs from free-ranging howler monkeys for DNA extraction, by M. Am�ndola-Pimenta, L. Garc�a-Feria, J. C. Serio-Silva, & V. Rico-Gray.
* American Journal of Primatology, 2009, 71[5], <www3.
interscience.wiley.com/journal/122277859/issue>.
. . .Contents: Editorial, by Paul A. Garber; First record of tool use by wild populations of the yellow-breasted capuchin monkey (Cebus xanthosternos) and new records for the bearded capuchin (Cebus libidinosus), by G. Rodrigues Canale, C. E. Guidorizzi, M. C. Martins Kierulff, & C. A. F. Rodrigues Gatto; Positive reinforcement training in rhesus macaques � Training progress as a result of training frequency, by A.-L. Fernstr�m, H. Fredlund, M. Sp�ngberg, & K. Westlund; The effects of infant births on male-female relationships in Cebus capucinus, by C. R. Sheller, Z. King, & K. Jack; Evaluation of antral follicle growth in the macaque ovary during the menstrual cycle and controlled ovarian stimulation by high-resolution ultrasonography, by C. V. Bishop, M. L. Sparman, J. E. Stanley, A. Bahar, M. B. Zelinski., & R. L. Stouffer; Individual variation in nest size and nest site features of the Bornean orangutans (Pongo pygmaeus), by Y. Rayadin & T. Saitoh; First description of the surgical anatomy of the cynomolgus monkey liver, by C. Vons, S. Beaudoin, N. Helmy, I. Dagher, A. Weber, & D. Franco; Post-conflict third-party affiliation in chimpanzees: What�s in it for the third party? by S. E. Koski & E. H. M. Sterck; Social facilitation of exploratory foraging behavior in capuchin monkeys (Cebus apella), by M. Dindo, A. Whiten, & F. B. M. de Waal; Seasonality of LH, testosterone and sperm parameters in spider monkey males (Ateles geoffroyi), by A. L. Cerda-Molina, L. Hern�ndez-L�pez, R. Chavira-Ram�rez, M. C�rdenas, & R. Mondrag�n-Ceballos; Food choices and habitat use by the Tana River yellow baboons (Papio cynocephalus): A preliminary report on five years of data, by V. K. Bentley-Condit; and Lemurs and kids: Primate conservation awareness through children�s books, by B. Urbani.
* American Journal of Primatology, 2009, 71[6], <www3. interscience.wiley.com/journal/34629/home>.
. . .Contents: Assessing reproductive profiles in female brown mouse lemurs (Microcebus rufus) from Ranomafana National Park, southeast Madagascar, using fecal hormone analysis, by M. B. Blanco & J. S. Meyer; Estimating chimpanzee population size with nest counts: Validating methods in Ta� National Park, by C. Y. Kouakou, C. Boesch, & H. Kuehl; Interactions between zoo-housed great apes and local wildlife, by S. R. Ross, A. N. Holmes, & E. V. Lonsdorf; Dental topography and diets of four Old World monkey species, by J. M. Bunn & P. S. Ungar; The feeding ecology and activity budget of proboscis monkeys, by I. Matsuda, A. Tuuga, & S. Higashi; What role do olfactory cues play in chacma baboon mating? by P. M. R. Clarke, L. Barrett, & S. P. Henzi; Social learning and mother�s behavior in manipulative tasks in infant marmosets, by V. Dell�Mour, F. Range, & L. Huber; Successful behavioral strategy to unite mother and infant rhesus monkeys (Macaca mulatta) after cesarean delivery, by A. M. Ruggiero, M. F. S. X. Novak, R. A. Woodward, & S. J. Suomi; and Characteristics of a group of Hubei Golden Snub-nosed Monkeys (Rhinopithecus roxellana hubeiensis) before and after major snow storms, by Y. Li, X. Liu, M. Liao, J. Yang, & C. B. Stanford.
* The Defender, Spring, 2009, <www.all-creatures .org/saen/nl-2009-sp.pdf>. (Stop Animal Exploitation NOW! <www.all-creatures.org/saen/nl.html>).
* Folia Primatologica, 2009, 80[1], <www.karger.com/fpr>.
. . .Contents: Discovery of sympatric dwarf lemur species in the high-altitude rain forest of Tsinjoarivo, eastern Madagascar: Implications for biogeography and conservation, by M. B. Blanco, L. R. Godfrey, M. Rakotondratsima, V. Rahalinarivo, K. E. Samonds, J.-L. Raharison, & M. T. Irwin; Skewed matrilineal genetic composition in a small wild chimpanzee community, by M. K. Shimada, S. Hayakawa, S. Fujita, Y. Sugiyama, & N. Saitou; Home range estimates vary with sample size and methods, by S. A. Boyle, W. C. Louren�o, L. R. da Silva, & A. T. Smith; and Development of bipedal walking in humans and chimpanzees: A comparative study, by T. Kimura & N. Yaguramaki.
* Great Ape Trust News, March, 2009, Issue 52, <www.greatapetrust.org>.
* International Journal of Primatology, 2009, 30[1].
. . .Contents: Population survey and demographic features of a coastal island population of Alouatta clamitans in Atlantic Forest, southeastern Brazil, by B. Ingberman, R. Fusco-Costa, & E. Leite de Araujo Monteiro-Filho; Diet of Pygathrix nigripes in southern Vietnam, by Hoang Minh Duc, G. S. Baxter, & M. J. Page; Morphological and genetic characterization of Saimiri boliviensis, by E. R. Steinberg, M. Nieves, M. S. Ascunce, A. M. Palermo, & M. D. Mudry; Variation over time in parasite prevalence among free-ranging chimpanzees at Gombe National Park, Tanzania, by J. S. Bakuza & G. Nkwengulila; What do primates hear? A meta-analysis of all known nonhuman primate behavioral audiograms, by M. N. Coleman; Can auditory playback condition predator mobbing in captive-reared Saguinus oedipus? by M. W. Campbell & C. T. Snowdon; Demography and life histories of sympatric patas monkeys, Erythrocebus patas, and vervets, Cercopithecus aethiops, in Laikipia, Kenya, by L. A. Isbell, T. P. Young, K. E. Jaffe, A. A. Carlson, & R. L. Chancellor; Field anesthesia and health assessment of free-ranging Cebus capucinus in Panama, by M. C. Crofoot, T. M. Norton, R. G. Lessnau, T. C. Viner, T. C. Chen, L. M. Mazzaro, & M. J. Yabsley; Understanding behavioral traditions in primates: Are current experimental approaches too focused on food? by C. F. I. Watson & C. A. Caldwell; Effects of yearly change in nut fruiting on autumn home-range use by Macaca fuscata on Kinkazan Island, northern Japan, by Y. Tsuji & S. Takatsuki; Manual laterality for simple reaching and bimanual coordinated task in naturalistic housed Pan troglodytes, by M. Llorente, M. Mosquera, & M. Fabr�; Human ability to recognize kin visually within primates, by A. Alvergne, E. Huchard, D. Caillaud, M. J. E. Charpentier, J. M. Setchell, C. Ruppli, D. F�jan, L. Martinez, G. Cowlishaw, & M. Raymond; and Cerebrospinal fluid monoaminergic metabolites in wild Papio anubis and P. hamadryas are concordant with taxon-specific behavioral ontogeny, by C. J. Jolly, J. E. Phillips-Conroy, J. R. Kaplan, & J. J. Mann.
* International Journal of Primatology, 2009, 30[2].
. . .Contents: Modeling the geographical distribution and fundamental niches of Cacajao spp. and Chiropotes israelita in northwestern Amazonia via a maximum entropy algorithm, by J. P. Boubli & M. G. de Lima; Temporal variation in insect-eating by chimpanzees and gorillas in southeast Cameroon: Extension of niche differentiation, by I. Deblauwe; Nesting behavior of Gorilla gorilla diehli at Kagwene Mountain, Cameroon: Implications for assessing group size and density, by J. L. Sunderland-Groves, A. Ekinde, & H. Mboh; The social system of Lariang tarsiers (Tarsius lariang) as revealed by genetic analyses, by C. Driller, D. Perwitasari-Farajallah, H. Zischler, & S. Merker; Diversity, body mass, and latitudinal gradients in primates, by A. H. Harcourt & B. M. Schreier; Distribution and population densities of diurnal primates in the Karst Forests of Phong Nha � Ke Bang National Park, Quang Binh Province, Central Vietnam, by T. Haus, M. Vogt, B. Forster, Ngoc Thanh Vu, & T. Ziegler; Ranging behavior of proboscis monkeys in a riverine forest with special reference to ranging in inland forest, by I. Matsuda, A. Tuuga & S. Higashi; Ovarian and uterine ultrasonography in Aotus azarai infulatus, by F. O. Barros Monteiro, L. Nassar Coutinho, E. do Socorro de Souza Pompeu, P. H. Gomes de Castro, C. E. Maia, W. L. Assun��o Pereira, & W. R. Russiano Vicente; A high polymorphism level in Rhinopithecus roxellana, by D. Pan, H.-X. Hu, S.-J. Meng, Z.-M. Men, Y.-X. Fu, & Y.-P. Zhang; Sleeping site use by Trachypithecus francoisi at Nonggang Nature Reserve, China, by Z. Qihai, H. Chengming, L. Ming, & W. Fuwen; and Home range size and use in Allocebus trichotis in Analamazaotra Special Reserve, Central Eastern Madagascar, by K. Biebouw.
* Primates, 2009, 50[1], <www.springerlink.com/content/109670>.
. . .Contents: The 60th anniversary of Japanese primatology, by Toshisada Nishida; Chimpanzee social intelligence: Selfishness, altruism, and the mother�infant bond, by S. Hirata; Within-species differences in primate social structure: Evolution of plasticity and phylogenetic constraints, by C. A. Chapman & J. M. Rothman; Emergence, propagation or disappearance of novel behavioral patterns in the habituated chimpanzees of Mahale: A review, by T. Nishida, T. Matsusaka, & W. C. McGrew; Gibbons under seasonal stress: The diet of the black crested gibbon (Nomascus concolor) on Mt. Wuliang, Central Yunnan, China, by P. Fan, Q. Ni, G. Sun, B. Huang, & X. Jiang; Predation on giant flying squirrels (Petaurista philippensis) by black crested gibbons (Nomascus concolor jingdongensis) at Mt. Wuliang, Yunnan, China, by P.-f. Fan & X.-l. Jiang; Seasonality of mating and birth in wild black-and-white snub-nosed monkeys (Rhinopithecus bieti) at Xiaochangdu, Tibet, by Z.-F. Xiang & K. Sayers; A case of spontaneous acquisition of a human sound by an orangutan, by S. A. Wich, K. B. Swartz, M. E. Hardus, A. R. Lameira, E. Stromberg, & R. W. Shumaker; Species, age and sex differences in type and frequencies of injuries and impairments among four arboreal primate species in Kibale National Park, Uganda, by M. E. Arlet, J. R. Carey, & F. Molleman; Group size and composition of black-and-gold howler monkeys (Alouatta caraya) on the Upper Paran� River, Southern Brazil, by L. M. Aguiar, G. Ludwig, & F. C. Passos; The type locality of Pan troglodytes vellerosus (Gray, 1862), and implications for the nomenclature of West African chimpanzees, by J. F. Oates, C. P. Groves, & P. D. Jenkins; Choice of analytical method can have dramatic effects on primate home range estimates, by C. C. Grueter, D. Li, B. Ren, & F. Wei; and Grandmothers care for orphans in a provisioned troop of Japanese macaques (Macaca fuscata), by M. Nozaki.
* Primates, 2009, 50[2], <www.springerlink.com/content/109670>.
. . .Contents: Distribution of potential suitable hammers and transport of hammer tools and nuts by wild capuchin monkeys, by E. Visalberghi, N. Spagnoletti, E. D. Ramos da Silva, F. R. D. Andrade, E. Ottoni, P. Izar, & D. Fragaszy; Using genetics to understand the dynamics of wild primate populations, by L. Vigilant & K. Guschanski; A comparative psychophysical approach to visual perception in primates, by T. Matsuno & K. Fujita; Feeding rate as valuable information in primate feeding ecology, by N. Nakagawa; Interaction studies in Japanese primatology: Their scope, uniqueness, and the future, by M. Nakamura; Distribution of dorsal carriage among simians, by M. Nakamichi & K. Yamada; Geometric characters of the radius and tibia in Macaca mulatta and Macaca fascicularis, by Y. Kikuchi & Y. Hamada; Prevalence of muzzle-rubbing and hand-rubbing behavior in wild chimpanzees in Mahale Mountains National Park, Tanzania, by N. Corp, H. Hayaki, T. Matsusaka, S. Fujita, K. Hosaka, N. Kutsukake, Michio Nakamura, Miho Nakamura, H. Nishie, M. Shimada, K. Zamma, W. Wallauer, & T. Nishida; and Effect of giving birth on the cortisol level in a bonobo groups� (Pan paniscus) saliva, by V. Behringer, W. Clau�, K. Hachenburger, A. Kuchar, E. M�stl, & D. Selzer.
* IPS Bulletin, May, 2009, 35[1].
. . .Contents include reports of projects funded by IPS grants.
* Oryx, 2008, 42[3], <journals.cambridge.org/action/displayJournal?jid=ORX>.
. . .Contents include: Primate surveys and conservation assessments, by A. B. Rylands, E. A. Williamson, M. Hoffmann, & R. A. Mittermeier; Distribution and conservation status of the orang-utan (Pongo spp.) on Borneo and Sumatra: How many remain? by S. A. Wich, E. Meijaard, A. J. Marshall, S. Husson, M. Ancrenaz, R. C. Lacy, C. P. van Schaik, J. Sugardjito, T. Simorangkir, K. Traylor-Holzer, M. Doughty, J. Supriatna, R. Dennis, M. Gumal, C. D. Knott, & I. Singleton; The critically endangered kipunji Rungwecebus kipunji of southern Tanzania: First census and conservation status assessment, by T. R. B. Davenport, D. W. De Luca, T. Jones, N. E. Mpunga, S. J. Machaga, A. Kitegile, & G. Picton Phillipps; In search of the munzala: Distribution and conservation status of the newly-discovered Arunachal macaque Macaca munzaa, by R. Suresh Kumar, N. Gama, R. Raghunath, A. Sinha, & C. Mishra; Monitoring population decline: Can transect surveys detect the impact of the Ebola virus on apes? by C. Devos, P. D. Walsh, E. Arnhem, & M.-C. Huynen; Abundance and community structure of Mentawai primates in the Peleonan forest, north Siberut, Indonesia, by M. Waltert, C. Abegg, T. Ziegler, S. Hadi, D. Priata, & K. Hodges; Seeing the wood for the trees: An assessment of the impact of participatory forest management on forest condition in Tanzania, by T. Blomley, K. Pfliegner, J. Isango, E. Zahabu, A. Ahrends, & N. Burgess; and A population estimate of the endangered chimpanzee Pan troglodytes vellerosus in a Nigerian montane forest: Implications for conservation, by J. Beck & H. Chapman.
Special Journal Issues
* Current Anthropology, 2009, 50[2], <www.journals.uchicago.edu/toc/ca/2009/50/2>.
. . .Contents include: A celebration of Clifford Jolly�s contribution to biological anthropology, by J.�E.�Phillips-Conroy & J.�Rogers; Fifty years of looking at human evolution: Backward, forward, and sideways, by C.�J.�Jolly; Elucidating population histories using genomic DNA sequences, by L.�Vigilant; Seeing the forest through the seeds: Mechanisms of primate behavioral diversity from individuals to populations and beyond, by K.�B.�Strier; The strength of great apes and the speed of humans, by A.�Walker; and Having a Jolly good time�together! Evolution by cooperative interaction, by K.�M.�Weiss.
Anatomy and Physiology
* An anterior temporal face patch in human cortex, predicted by macaque maps. Rajimehr, R., Young, J. C., & Tootell, R. B. H. (NMR Martinos Center, Mass. Gen. Hospital, Harvard Med. Sch., Charlestown, MA 02129 [e-mail: [email protected]]. Proceedings of the National Academy of Sciences, U.S.A., 2009, 106, 1995-2000, <www.pnas.org/content/106/6/1995.full>.
. . . �Increasing evidence suggests that primate visual cortex has a specialized architecture for processing discrete object categories such as faces. Human fMRI studies have described a localized region in the fusiform gyrus [the fusiform face area (FFA)] that responds selectively to faces. In contrast, in nonhuman primates, electrophysiological and fMRI studies have instead revealed 2 apparently analogous regions of face representation: the posterior temporal face patch (PTFP) and the anterior temporal face patch (ATFP). An earlier study suggested that human FFA is homologous to the PTFP in macaque. However, in humans, no obvious homologue of the macaque ATFP has been demonstrated. Here, we used fMRI to map face-selective sites in both humans and macaques, based on equivalent stimuli in a quantitative topographic comparison. This fMRI evidence suggests that such a face-selective area exists in human anterior inferotemporal cortex, comprising the apparent homologue of the fMRI-defined ATFP in macaques.�
* Endocranial volumes of primate species: Scaling analyses using a comprehensive and reliable data set. Isler, K., Kirk, C. E., Miller, J. M. A., Albrecht, G. A., Gelvin, B. R., & Martin, R. D. (Anthro. Inst. & Museum, Univ. of Z�rich-Irchel, Winterthurerstr. 190, CH-8057 Z�rich, Switzerland [e-mail: [email protected]]). Journal of Human Evolution, 2008, 55, 967-978.
. . . �We present a compilation of endocranial volumes (ECV) for 176 nonhuman primate species based on individual data collected from 3813 museum specimens, at least 88% being wild-caught. In combination with body mass data from wild individuals, strong correlations between endocranial volume and body mass within taxonomic groups were found. Errors attributable to different techniques for measuring cranial capacity were negligible and unbiased. The overall slopes for regressions of log ECV on log body mass in primates are 0.773 for least-squares regression and 0.793 for reduced major axis regression. The least-squares slope is reduced to 0.565 when independent contrasts are substituted for species means (branch lengths from molecular studies). A common slope of 0.646 is obtained with logged species means when grade shifts between major groups are taken into account using ANCOVA. In addition to providing a comprehensive and reliable database for comparative analyses of primate brain size, we show that the scaling relationship between brain mass and ECV does not differ significantly from isometry in primates. We also demonstrate that ECV does not differ substantially between captive and wild samples of the same species. ECV may be a more reliable indicator of brain size than brain mass, because considerably larger samples can be collected to better represent the full range of intraspecific variation. We also provide support for the maternal energy hypothesis by showing that basal metabolic rate (BMR) and gestation period are both positively correlated with brain size in primates, after controlling for the influence of body mass and potential effects of phylogenetic relatedness.�
Animal Models
* Trial outcome and associative learning signals in the monkey hippocampus. Wirth, S., Avsar, E., Chiu, C. C., Sharma, V., Smith, A. C., Brown, E., & Suzuki, W. A. (W. A. S., Ctr for Neural Sci., New York Univ., New York, NY 10003 [e-mail: [email protected]]). Neuron, 2009, 61, 930-940, <www.cell.com/neuron/fulltext/S0896-6273(09)00079-8 >.
. . . �In tasks of associative learning, animals establish new links between unrelated items by using information about trial outcome to strengthen correct/rewarded associations and modify incorrect/unrewarded ones. To study how hippocampal neurons convey information about reward and trial outcome during new associative learning, we recorded hippocampal neurons as monkeys learned novel object-place associations. A large population of hippocampal neurons (50%) signaled trial outcome by differentiating between correct and error trials during the period after the behavioral response. About half these cells increased their activity following correct trials (correct up cells) while the remaining half fired more following error trials (error up cells). Moreover, correct up cells, but not error up cells, conveyed information about learning by increasing their stimulus-selective response properties with behavioral learning. These findings suggest that information about successful trial outcome conveyed by correct up cells may influence new associative learning through changes in the cells� stimulus-selective response properties.�
* Extracting kinematic parameters for monkey bipedal walking from cortical neuronal ensemble activity. Fitzsimmons, N. A., Lebedev, M. A., Peikon, I. D., & Nicolelis, M. A. (Dept of Neurobiology, Duke Univ., Durham, NC 27708 [e-mail: [email protected]]). Frontiers in Integrative Neuroscience, 2009, 3[3], <frontiersin.org/integrativeneuroscience/paper/10.3389/neuro.07/003.2009>.
. . . �The ability to walk may be critically impacted as the result of neurological injury or disease. While recent advances in brain�machine interfaces (BMIs) have demonstrated the feasibility of upper-limb neuroprostheses, BMIs have not been evaluated as a means to restore walking. Here, we demonstrate that chronic recordings from ensembles of cortical neurons can be used to predict the kinematics of bipedal walking in rhesus macaques � both offline and in real-time. Linear decoders extracted 3D coordinates of leg joints and leg muscle EMGs from the activity of hundreds of cortical neurons. As more complex patterns of walking were produced by varying the gait speed and direction, larger neuronal populations were needed to accurately extract walking patterns. Extraction was further improved using a switching decoder which designated a submodel for each walking paradigm. We propose that BMIs may one day allow severely paralyzed patients to walk again.�
* Impaired attentional selection following lesions to human pulvinar: Evidence for homology between human and monkey. Snow, J. C., Allena, H. A., Rafalc, R. D., & Humphreys, G. W. (Sch. of Psych., Behav. Brain Sciences, Univ. of Birmingham, Edgbaston, Birmingham B15 2TT, U.K. [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2009, 106, 4054-4059, <www.pnas.org/content/106/10/4054>.
. . . �We examined the contributions of the human pulvinar to goal directed selection of visual targets in three patients with chronic, unilateral lesions involving topographic maps in the ventral pulvinar. Observers completed two psychophysical tasks in which they discriminated the orientation of a lateralized target grating in the presence of vertically-aligned distracters. In experiment 1, where distracter contrast was varied while target contrast remained constant, the patients� contralesional contrast thresholds for discriminating the orientation of grating stimuli were elevated only when the task required selection of a visual target in the face of competition from a salient distracter. Attentional selectivity was restored in the patients in experiment 2 where target contrast was varied while distracter contrast remained constant. These observations provide the first evidence that the human pulvinar plays a necessary role in modulating physical saliency in attentional selection, and supports a homology in global pulvinar structure between humans and monkey.�
* Enhancing SIV-specific immunity in vivo by PD-1 blockade. Velu, V., Titanji, K., Zhu, B., Husain, S., Pladevega, A., Lai, L., Vanderford, T. H., Chennareddi, L., Silvestri, G., Freeman, G. J., Ahmed, R., & Amara, R. R. (R.R.A., Emory Vaccine Ctr, Emory Univ. Sch. of Med., Atlanta, GA 30322 [e-mail: [email protected]]). Nature, 2009, 458, 206-210.
. . . �Chronic immunodeficiency virus infections are characterized by dysfunctional cellular and humoral antiviral immune responses. As such, immune modulatory therapies that enhance and/or restore the function of virus-specific immunity may protect from disease progression. Here we investigate the safety and immune restoration potential of blockade of the co-inhibitory receptor programmed death 1 (PD-1) during chronic simian immunodeficiency virus (SIV) infection in macaques. We demonstrate that PD-1 blockade using an antibody to PD-1 is well tolerated and results in rapid expansion of virus-specific CD8 T cells with improved functional quality. This enhanced T-cell immunity was seen in the blood and also in the gut, a major reservoir of SIV infection. PD-1 blockade also resulted in proliferation of memory B cells and increases in SIV envelope-specific antibody. These improved immune responses were associated with significant reductions in plasma viral load and also prolonged the survival of SIV-infected macaques. Blockade was effective during the early (week 10) as well as late (~week 90) phases of chronic infection even under conditions of severe lymphopenia. These results demonstrate enhancement of both cellular and humoral immune responses during a pathogenic immunodeficiency virus infection by blocking a single inhibitory pathway and identify a novel therapeutic approach for control of human immunodeficiency virus infections.�
* Combined effect of antiretroviral therapy and blockade of IDO in SIV-Infected rhesus macaques. Boasso, A., Vaccari, M., Fuchs, D., Hardy, A. W., Tsai, W.-P., Tryniszewsk, E., Shearer, G. M., & Franchini, G. (Imperial College, Fac. of Med., Dept of Immunology, Chelsea & Westminster Hospital, 369 Fulham Rd, London, SW10 9NH, U.K. [e-mail: [email protected]]). Journal of Immunology, 2009, 182, 4313-4320.
. . . �Increased activity of IDO, which catalyzes the degradation of Trp into kynurenine (Kyn), is observed during HIV/SIV infection, and it may contribute to the persistence of HIV/SIV by suppressing antiviral T cell responses. We administered the IDO inhibitor 1-methyl-D-tryptophan (D-1mT) for 13 days to SIV-infected rhesus macaques receiving antiretroviral therapy (ART). D-1mT treatment increased the plasma levels of Trp, without reducing the levels of Kyn, suggesting only a partial effect on IDO enzymatic activity. Surprisingly, D-1mT significantly reduced the virus levels in plasma and lymph nodes of ART-treated animals with incomplete responsiveness to ART. In SIV-infected animals that were not receiving ART, D-1mT was ineffective in reducing the plasma viral load and had only a marginal effect on the plasma Kyn/Trp ratio. Increased IDO and TGF-β mRNA expression in lymph nodes of ART-treated macaques after D-1mT treatment suggested that compensatory counterregulatory mechanisms were activated by D-1mT, which may account for the lack of effect on plasma Kyn. Finally, D-1mT did not interfere with the ART-induced T cell dynamics in lymph nodes (increased frequency of total CD4 T cells, increase of CD8 T cells expressing the antiapoptotic molecule Bcl2, and reduction of regulatory T cells). Thus, D-1mT appeared to synergize with ART in inhibiting viral replication and did not interfere with the beneficial immunologic effects of ART. Further studies are required to elucidate the immunologic or virologic mechanism by which D-1mT inhibited SIV replication in vivo.�
* Electrophysiological correlates of default-mode processing in macaque posterior cingulate cortex. Hayden, B. Y., Smith, D. V., & Platt, M. L. (Department of Neurobiology, Center for Neuroeconomic Studies, Center for Cognitive Neuroscience, Duke University School of Medicine, Durham, NC 27710 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2009, 106, 5948-5953.
. . . �During the course of daily activity, our level of engagement with the world varies on a moment-to-moment basis. Although these fluctuations in vigilance have critical consequences for our thoughts and actions, almost nothing is known about the neuronal substrates governing such dynamic variations in task engagement. We investigated the hypothesis that the posterior cingulate cortex (CGp), a region linked to default-mode processing by hemodynamic and metabolic measures, controls such variations. We recorded the activity of single neurons in CGp in two macaque monkeys performing simple tasks in which their behavior varied from vigilant to inattentive. We found that firing rates were reliably suppressed during task performance and returned to a higher resting baseline between trials. Importantly, higher firing rates predicted errors and slow behavioral responses, and were also observed during cued rest periods when monkeys were temporarily liberated from exteroceptive vigilance. These patterns of activity were not observed in the lateral intraparietal area, an area linked to the frontoparietal attention network. Our findings provide physiological confirmation that CGp mediates exteroceptive vigilance and are consistent with the idea that CGp is part of the �default network� of brain areas associated with control of task engagement.�
* Clozapine normalizes prefrontal cortex dopamine transmission in monkeys subchronically exposed to phencyclidine. Elsworth, J. D.,�Jentsch, J. D., Morrow, B. A., Redmond,�Jr., D. E., & Roth, R. H. (Dept of Psychiatry, Yale Univ. Sch. of Med., 300 George St, 8300 Suite, New Haven, CT 06511 [e-mail: [email protected]]). Neuropsychopharmacology, 2008, 33, 491-496; <www.nature.com/npp/journal/v33/n3/full/1301448a.html>.
. . . �The mechanism responsible for the therapeutic effects of the prototypical atypical antipsychotic drug, clozapine, is still not understood; however, there is persuasive evidence from in vivo studies in normal rodents and primates that the ability to elevate dopamine neurotransmission preferentially in the prefrontal cortex is a key component to the beneficial effects of clozapine in schizophrenia. Theoretically, such an effect of clozapine would counteract the deficient dopaminergic innervation of the prefrontal cortex that appears to be part of the pathophysiology of schizophrenia. We have previously shown that following repeated, intermittent administrations of phencyclidine to monkeys there is lowered prefrontal cortical dopamine transmission and impairment of cognitive performance that is dependent on the prefrontal cortex; these biochemical and behavioral changes therefore model certain aspects of schizophrenia. We now investigate the effects of clozapine on the dopamine projections to prefrontal cortex, nucleus accumbens, and striatum in control monkeys and in those withdrawn from repeated phencyclidine treatment, using a dose regimen of clozapine that ameliorates the cognitive deficits described in the primate phencyclidine (PCP) model. In normal monkeys, clozapine elevated dopamine turnover in all prefrontal cortical, but not subcortical, regions analyzed. In the primate PCP model, clozapine normalized dopamine (DA) turnover in the dorsolateral prefrontal cortex, prelimbic cortex, and cingulate cortex. Thus, the present data support the hypothesis that the therapeutic effects of clozapine in this primate model and perhaps in schizophrenia may be related at least in part to the restoration of DA tone in the prefrontal cortex.�
* A macaque model of HIV-1 infection. Hatziioannou, T., Ambros, Z., Chung, N. P. Y., Piatak, Jr., M., Yuan, F., Trubey, C. M., Coalter, V., Kiser, R., Schneider, D., Smedley, J., Pung, R., Gathuka, M., Estes, J. D., Veazey, R. S., KewalRamani, V. N., Lifson, J. D., & Bieniasz, P. D. (Aaron Diamond AIDS Research Center, 455 First Ave, New York, NY 10016 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A. , 2009, 106, 4425-4429.
. . . �The lack of a primate model that utilizes HIV-1 as the challenge virus is an impediment to AIDS research; existing models generally employ simian viruses that are divergent from HIV-1, reducing their usefulness in preclinical investigations. Based on an understanding of species-specific variation in primate TRIM5 and APOBEC3 antiretroviral genes, we constructed simian-tropic (st)HIV-1 strains that differ from HIV-1 only in the vif gene. We demonstrate that such minimally modified stHIV-1 strains are capable of high levels of replication in vitro in pig-tailed macaque (Macaca nemestrina) lymphocytes. Importantly, infection of pig-tailed macaques with stHIV-1 results in acute viremia, approaching the levels observed in HIV-1-infected humans, and an ensuing persistent infection for several months. stHIV-1 replication was controlled thereafter, at least in part, by CD8+ T cells. We demonstrate the potential utility of this HIV-1-based animal model in a chemoprophylaxis experiment, by showing that a commonly used HIV-1 therapeutic regimen can provide apparently sterilizing protection from infection following a rigorous high-dose stHIV-1 challenge.�
* Glycerol monolaurate prevents mucosal SIV transmission. Li, O., Estes, J. D., Schlievert, P. M., Duan, L., Brosnahan, A. J., Southern, P. J., Reilly, C. S., Peterson, M. L., Schultz-Darken, N., Brunner, K. G., Nephew, K. R., Pambuccian, S., Lifson, J. D., Carlis, J. V., & Haase, A. T. (A. T. H., Dept of Microbiol., Med. Sch., Univ. of Minnesota, MMC 196, 420 Delaware St S.E., Minneapolis, MN 55455 [e-mail: [email protected]]). Nature , 2009, 458, 1034-1038.
. . . �Although there has been great progress in treating human immunodeficiency virus 1 (HIV-1) infection, preventing transmission has thus far proven an elusive goal. Indeed, recent trials of a candidate vaccine and microbicide have been disappointing, both for want of efficacy and concerns about increased rates of transmission. Nonetheless, studies of vaginal transmission in the simian immunodeficiency virus (SIV)�rhesus macaque (Macacca mulatta) model point to opportunities at the earliest stages of infection in which a vaccine or microbicide might be protective, by limiting the expansion of infected founder populations at the portal of entry. Here we show in this SIV�macaque model, that an outside-in endocervical mucosal signalling system, involving MIP-3α (also known as CCL20), plasmacytoid dendritic cells and CCR5+ cell-attracting chemokines produced by these cells, in combination with the innate immune and inflammatory responses to infection in both cervix and vagina, recruits CD4+ T cells to fuel this obligate expansion. We then show that glycerol monolaurate�a widely used antimicrobial compound with inhibitory activity against the production of MIP-3α and other proinflammatory cytokines�can inhibit mucosal signalling and the innate and inflammatory response to HIV-1 and SIV in vitro, and in vivo it can protect rhesus macaques from acute infection despite repeated intra-vaginal exposure to high doses of SIV. This new approach, plausibly linked to interfering with innate host responses that recruit the target cells necessary to establish systemic infection, opens a promising new avenue for the development of effective interventions to block HIV-1 mucosal transmission.�
* Fictive reward signals in the anterior cingulate cortex. Hayden, B. Y., Pearson, J. M., & Platt, M. L. (Dept of Neurobiol., Duke Univ. Sch. of Med., Duke Univ., Durham, NC 27701 [e -mail: [email protected]]). Science, 2009, 324, 948-950, <www.sciencemag.org/cgi/content/full/324/5929/948>.
. . . �The neural mechanisms supporting the ability to recognize and respond to fictive outcomes, outcomes of actions that one has not taken, remain obscure. We hypothesized that neurons in the anterior cingulate cortex (ACC), which monitors the consequences of actions and mediates subsequent changes in behavior, would respond to fictive reward information. We recorded responses of single neurons during performance of a choice task that provided information about the reward values of options that were not chosen. We found that ACC neurons signal fictive reward information and use a coding scheme similar to that used to signal experienced outcomes. Thus, individual ACC neurons process both experienced and fictive rewards.�
* Generation of transgenic non-human primates with germline transmission. Sasaki, E., Suemizu, H., Shimada, A., Hanazawa, K., Oiwa, R., Kamioka, M., Tomioka, I., Sotomaru, Y., Hirakawa, R., Eto, T., Shiozawa, S., Maeda, T., Ito, M., Ito, R., Kito, C., Yagihashi, C., Kawai, K., Miyoshi, H., Tanioka, Y., Tamaoki, N., Habu, S., Okano, H., & Nomura, T. (Central Inst. for Exper. Animals, 1430 Nogawa, Miyamae-ku, Kawasaki, Kanagawa 216-0001, Japan [e-mail: [email protected]]). Nature, 2009, 459, 523-527, <www.nature.com/nature/journal/v459/n7246>.
. . . �The common marmoset (Callithrix jacchus) is increasingly attractive for use as a nonhuman primate animal model in biomedical research. It has a relatively high reproduction rate for a primate, making it potentially suitable for transgenic modification. Although several attempts have been made to produce nonhuman transgenic primates, transgene expression in the somatic tissues of live infants has not been demonstrated by objective analyses such as polymerase chain reaction with reverse transcription or western blots. Here we show that the injection of a self-inactivating lentiviral vector in sucrose solution into marmoset embryos results in transgenic common marmosets that expressed the transgene in several organs. Notably, we achieved germline transmission of the transgene, and the transgenic offspring developed normally. The successful creation of transgenic marmosets provides a new animal model for human disease that has the great advantage of a close genetic relationship with humans. This model will be valuable to many fields of biomedical research.�
Animal Welfare
* Primate location preference in a double-tier cage: The effects of illumination and cage height. MacLean, E. L., Roberts Prior, S., Platt, M. L., & Brannon, E. M. (Ctr for Cog. Neurosci., Box 90999, Duke Univ., Durham, NC 27708 [e-mail: [email protected]]). Journal of Applied Animal Welfare Science, 2009, 12, 73-81.
. . .Nonhuman primates are frequently housed in double-tier arrangements with significant differences between the environments of the upper and lower-row cages. Although several studies have investigated whether this arrangement alters monkeys� behavior, no studies have addressed the two most notable differences, light and height, individually to determine their relative importance. This experiment examined how rhesus and long-tailed macaques allocated their time between the upper and lower-row cages of a 1-over-1 apartment module under different lighting conditions. In Condition A, monkeys� baseline degree of preference for the upper and lower-row was tested. In Condition B, the lighting environment was reversed by limiting illumination in the upper-row cage and increasing illumination in the lower-row cage. In both conditions, monkeys spent more time in the upper-row cage, thus indicating a strong preference for elevation regardless of illumination. The amount of time that monkeys spent in the lower-row cage increased by 7% under reversed lighting, but this trend was not significant. These results corroborate the importance of providing captive primates with access to elevated areas.
* Environmental standardization: Cure or cause of poor reproducibility in animal experiments? Richter, S. H., Garner, J. P., & W�rbel, H. (H. W., Animal Welfare & Ethology, Justus-Liebig-Univ. of Giessen, Giessen, Germany [e-mail:�[email protected]]). Nature Methods, 2009, 6, 257-261, <www.nature.com/nmeth/journal/v6/n4/full/nmeth.1312.html>.
. . . �It is widely believed that environmental standardization is the best way to guarantee reproducible results in animal experiments. However, mounting evidence indicates that even subtle differences in laboratory or test conditions can lead to conflicting test outcomes. Because experimental treatments may interact with environmental conditions, experiments conducted under highly standardized conditions may reveal local �truths� with little external validity. We review this hypothesis here and present a proof of principle based on data from a multilaboratory study on behavioral differences between inbred mouse strains. Our findings suggest that environmental standardization is a cause of, rather than a cure for, poor reproducibility of experimental outcomes. Environmental standardization can contribute to spurious and conflicting findings in the literature and unnecessary animal use. This conclusion calls for research into practicable and effective ways of systematic environmental heterogenization to attenuate these scientific, economic and ethical costs.�
* Refinements in husbandry, care and common procedures for non-human primates. Ninth report of the BVAAWF/FRAME/RSPCA/UFAW Joint Working Group on Refinement. Jennings, M., & Prescott, M. J. (Eds.). (Res. Animals Dept, RSPCA, Wilberforce Way, Southwater, Horsham, West Sussex RH13 9RS, U.K. [e-mail: [email protected]]). Laboratory Animals, 2009, 43[Suppl. 1], 1-47.
. . .This report, produced by the British Veterinary Association Animal Welfare Foundation /Fund for the Replacement of Animals in Medical Experiments/Royal Society for the Prevention of Cruelty to Animals/Universities Federation for Animal Welfare Joint Working Group on Refinement, sets out practical guidance on refining the husbandry and care of nonhuman primates (hereinafter primates) and on minimizing the adverse effects of some common procedures. It provides a valuable resource to help understand the physical, social and behavioral characteristics and needs of individual primates, and is intended to develop and complement the existing literature and legislative guidelines. Topics covered include refinements in housing, husbandry and common procedures such as restraint, identification, and sampling; with comprehensive advice on issues such as primate communication, assessing and facilitating primate well-being, establishing and maintaining social groups, environmental and nutritional enrichment, and animal passports. The most commonly used species are the key focus of this resource, but its information and recommendations are generally applicable to other species, provided that relevant individual species characteristics are taken into account.
Behavior
* Free-ranging macaque mothers exaggerate tool-using behavior when observed by offspring. Masataka, N., Koda, H., Urasopon, N., & Watanabe, K. (Primate Res. Inst., Kyoto Univ., Inuyama, Japan [e-mail: [email protected]]). PLoS ONE, 2009, 4[3]: e4768.
. . . �The population-level use of tools has been reported in various animals. Nonetheless, how tool use might spread throughout a population is still an open question. In order to answer that, we observed the behavior of inserting human hair or human-hair-like material between their teeth as if they were using dental floss in a group of long-tailed macaques (Macaca fascicularis) in Thailand. The observation was undertaken by video-recording the tool-use of 7 adult females who were rearing 1-year-old infants, using the focal-animal-sampling method. When the data recorded were analyzed separately according to the presence/absence of the infant of the target animal in the target animal�s proximity, the pattern of the tool-using action of long-tailed adult female macaques under our observation changed in the presence of the infant as compared with that in the absence of the infant so that the stream of tool-using action was punctuated by more pauses, repeated more often, and performed for a longer period during each bout in the presence of the infant. We interpret this as evidence for the possibility that they exaggerate their action in tool-using so as to facilitate the learning of the action by their own infants.�
* Spontaneous planning for future stone throwing by a male chimpanzee. Osvath, M. (Lund Univ. Cognitive Science, Kungshuset Lundag�rd, 222 22 Lund, Sweden [e-mail: [email protected]]). Current Biology, 2009, 19 [5], R190.
. . .Planning for a future, rather than a current, mental state is a cognitive process generally viewed as uniquely human. Here, however, is a report on a decade of observations of spontaneous planning by a male chimpanzee in a zoo. The planning actions, which took place in a calm state, included stone caching and the manufacture of discs from concrete, objects later used as missiles against zoo visitors during agitated chimpanzee dominance displays. Such planning implies advanced consciousness and cognition traditionally not associated with nonhuman animals.
* Intersexual food transfer among orangutans: Do females test males for coercive tendency? van Noordwijk, M. A., & van Schaik, C. P. (Anthrop. Inst. & Museum, Univ. of Z�rich, Winterthurerstr. 190, 8057 Z�rich, Switzerland [e-mail: [email protected]]). Journal of Behavioral Ecology and Sociobiology, 2009, 63, 883-890.
. . . �Tolerated transfer of food among adults is rare among primates, except in humans. Here, we present data on a consistent pattern of tolerated intersexual transfer of food (held in hand, foot, or mouth by the owner) among adult orangutans, in two different natural populations (Pongo abelii and Pongo pygmaeus wurmbii), based on ca. 9,000 h of focal observation per site. Although rare, intersexual food transfers were disproportionately from males to sexually active females and involved food that was equally available to both sexes. There was no evidence for direct trading of food for social favors (mating, grooming, or agonistic support) or for sharing under pressure of harassment. However, females frequently protested with loud screams when males, especially unflanged ones, attempted to take food they possessed, and also when males responded aggressively to their taking attempt. Since associations ended sooner when the female emitted noisy calls, a male who did not allow a female to take food from him risked losing the association. These findings support the hypothesis that by taking food, a sexually active female may test the male�s tendency toward violence. Thus, intersexual food taking in orangutans is based on female leverage, resulting in a species-wide female entitlement to male �generosity�. The inhibition of food defense required for this kind of transaction may also form the basis for sharing patterns among species in which nutritional benefits have become important, such as chimpanzees and perhaps human foragers.�
* Wild chimpanzees exchange meat for sex on a long-term basis. Gomes, C. M., & Boesch, C. (Max Planck Inst. for Evolutionary Anthropology, Leipzig, Germany [e-mail: [email protected]]). PLoS ONE, 2009, 4[4], e5116, <www. plosone.org/article/info%3Adoi/10.1371/journal.pone.0005116>.
. . . �Humans and chimpanzees are unusual among primates in that they frequently perform group hunts of mammalian prey and share meat with conspecifics. Especially interesting are cases in which males give meat to unrelated females. The meat-for-sex hypothesis aims at explaining these cases by proposing that males and females exchange meat for sex, which would result in males increasing their mating success and females increasing their caloric intake without suffering the energetic costs and potential risk of injury related to hunting. Although chimpanzees have been shown to share meat extensively with females, there has not been much direct evidence in this species to support the meat-for-sex hypothesis. Here we show that female wild chimpanzees copulate more frequently with those males who, over a period of 22 months, share meat with them. We excluded other alternative hypotheses to exchanging meat for sex, by statistically controlling for rank of the male, age, rank and gregariousness of the female, association patterns of each male-female dyad and meat begging frequency of each female. Although males were more likely to share meat with estrous than anestrous females given their proportional representation in hunting parties, the relationship between mating success and sharing meat remained significant after excluding from the analysis sharing episodes with estrous females. These results strongly suggest that wild chimpanzees exchange meat for sex, and do so on a long-term basis. Similar studies on humans will determine if the direct nutritional benefits that women receive from hunters in foraging societies could also be driving the relationship between reproductive success and good hunting skills.�
* Gaze following and gaze priming in lemurs. Ruiz, A., G�mez, J. C., Roeder, J. J., & Byrne, R. W. (Scottish Primate Research Group and Centre for Social Learning and Cognitive Evolution, School of Psychology, Univ. of St Andrews, St Andrews, KY16 9JP, U.K.). Animal Cognition, 2008, 12, 427-434.
. . . �Although primates have often been found to co-orient visually with other individuals, members of these same species have usually failed to use co-orientation to find hidden food in object-choice experiments. This presents an evolutionary puzzle: what is the function of co-orientation if it is not used for a function as basic as locating resources? Co-orientation responses have not been systematically investigated in object-choice experiments, and requiring co-orientation with humans (as is typical in object-choice tasks) may underestimate other species� abilities. Using an object-choice task with conspecific models depicted in photographs, we provide experimental evidence that two lemur species (Eulemur fulvus, n = 4, and Eulemur macaco, n = 2) co-orient with conspecifics. Secondly, by analyzing together two measures that have traditionally been examined separately, we show that lemurs� gaze following behavior and ultimate choice are closely linked. Individuals were more likely to choose correctly after having looked in the same direction as the model, and thus chose objects correctly more often than chance. We propose a candidate system for the evolutionary origins of more complex gaze following: �gaze priming.��
* Gestural communication of the gorilla (Gorilla gorilla): repertoire, intentionality and possible origins. Genty, E., Breuer, T., Hobaiter, C., & Byrne, R. W. (R. W. B., Scottish Primate Research Group, Sch. of Psych., Univ. of St Andrews, Fife KY16 9JP, Scotland, U.K. [e-mail: [email protected]]). Animal Cognition, 2009, 12, 527-546.
. . . �Social groups of gorillas were observed in three captive facilities and one African field site. Cases of potential gesture use, totalling 9,540, were filtered by strict criteria for intentionality, giving a corpus of 5,250 instances of intentional gesture use. This indicated a repertoire of 102 gesture types. Most repertoire differences between individuals and sites were explicable as a consequence of environmental affordances and sampling effects: overall gesture frequency was a good predictor of universality of occurrence. Only one gesture was idiosyncratic to a single individual, and was given only to humans. Indications of cultural learning were few, though not absent. Six gestures appeared to be traditions within single social groups, but overall concordance in repertoires was almost as high between as within social groups. No support was found for the ontogenetic ritualization hypothesis as the chief means of acquisition of gestures. Many gestures whose form ruled out such an origin, i.e. gestures derived from species-typical displays, were used as intentionally and almost as flexibly as gestures whose form was consistent with learning by ritualization. When using both classes of gesture, gorillas paid specific attention to the attentional state of their audience. Thus, it would be unwarranted to divide ape gestural repertoires into �innate, species-typical, inflexible reactions� and �individually learned, intentional, flexible communication�. We conclude that gorilla gestural communication is based on a species-typical repertoire, like those of most other mammalian species but very much larger. Gorilla gestures are not, however, inflexible signals but are employed for intentional communication to specific individuals.�
Care
* Dose-finding study of Fluoxetine and Venlafaxine for the treatment of self-injurious and stereotypic behavior in rhesus macaques (Macaca mulatta). Fontenot, M. B., Musso, M. W., McFatter, R. M., & Anderson, G. M. (Div. of Behavioral Sci., New Iberia Res. Ctr, Univ. of Louisiana, Lafayette, LA 70560). Journal of the American Association for Laboratory Animal Science, 2009, 48, 176-184.
. . .The short-term effects on rates and durations of self-injurious behavior (SIB) and self-directed stereotypies associated with various doses of fluoxetine (FLX) and venlafaxine (VEN) were examined in rhesus macaques. Adult male macaques (n = 17; age, 7 to 15 yr) with at least one episode of severe SIB within the past 5 years were randomized to treatment with FLX (n = 6), VEN (n = 6), or placebo (PLC, n = 5), administered by voluntary consumption of medication provided in fruit-flavored tablets. After 4-wk baseline and 4-wk placebo lead-in phases, doses were increased monthly for four months (FLX: 0.5, 2.0, 4.0, and 8.0 mg/kg; VEN: 2.0, 4.0, 8.0, and 16.0 mg/kg). Animals in the PLC condition received similar nonmedicated fruit-flavored tablets. Focal behavioral observations, plasma drug levels, and neurochemical data were obtained. Results indicated that rates and percentage of time spent self-biting declined at all doses of FLX, with the greatest effect seen at 2.0 mg/kg. For VEN, percentage of time spent self-biting was significantly lower only at the 4.0 mg/kg dose. Treatment-induced reductions in platelet serotonin and cerebrospinal fluid 5-hydroxyindoleacetic acid (CSF 5HIAA) concentrations were substantially greater in the FLX-treated condition than in the VEN-treated condition. Plasma FLX and norfluoxetine levels increased with FLX dose; plasma levels of VEN were low and not dose-related. Fluoxetine at a dose of 2.0 mg/kg daily was most efficacious in reducing SIB, and the observed reductions in platelet serotonin and CSF 5HIAA levels indicated substantial bioeffect at this dose. Treatment with VEN was marked by noncompliance, low bioeffect, and low efficacy.
* L-Tryptophan and correlates of self-injurious behavior in small-eared bushbabies (Otolemur garnettii). Watson, S. L., McCoy, J. G., Fontenot, M. B., Hanbury, D. B., & Ward, C. P. (Univ. of So. Mississippi, Dept of Psych., Hattiesburg, MS 39406-0001). Journal of the American Association for Laboratory Animal Science, 2009, 48, 185-191.
. . .Self-injurious behavior (SIB) among captive primates is a recurring problem for those who manage such facilities. Its prevalence highlights the need for research evaluating the effectiveness of potential treatment approaches. In the present study, four weeks of dietary supplementation with L-tryptophan (100 mg daily) was evaluated for the treatment of self-inflicted wounds in 22 small-eared bushbabies, a prosimian primate, with a history of SIB. The treatment significantly reduced stereotypy and was associated with a reduction in wound area and severity. In terms of physiologic measures, preexisting high levels of cortisol were reduced in bushbabies with SIB, whereas serotonin concentrations were increased after four weeks of treatment. Results indicate that L-tryptophan as a dietary supplement may be a viable adjunct to standard husbandry procedures for animals exhibiting maladaptive behaviors such as stereotypy and SIB.
* Positive reinforcement training to enhance the voluntary movement of group-housed sooty mangabeys (Cercocebus atys atys). Veeder, C. L., Bloomsmith, M. A., McMillan, J. L., Perlman, J. E., & Martin, A. L. (Yerkes NPRC, Emory Univ., Atlanta, GA 30329). Journal of the American Association for Laboratory Animal Science, 2009, 48, 192-195.
. . . �Positive reinforcement training (PRT) has successfully been used to train diverse species to execute behaviors helpful in the everyday care and wellbeing of the animals. Because little information is available about training sooty mangabeys, we analyzed PRT with a group of 30 adult males as they were trained to shift from one side of their enclosure to the other. Over a four-month period we conducted 57 training sessions totaling 26.5 hours of training and recorded compliance information. During training, compliance increased from 76% of the animals during the first five training sessions to 86% of the animals shifting during the last five sessions. This result indicated progress but fell short of our goal of 90% compliance. After 25 training sessions, problem-solving techniques were applied to help the consistently noncompliant animals become more proficient. The techniques included reducing social stress by shifting animals so that noncompliant monkeys could shift into an unoccupied space, using more highly preferred foods, and �jackpot�-sized reinforcement. To determine whether social rank affected training success, animals were categorized into high, medium, and low dominance groups, based on seven hours of behavioral observations. A Kruskal-Wallis test result indicated a significant difference in compliance according to the category of dominance. Although training a group this large proved challenging, the mangabeys cooperated more than 90% of the time during follow-up sessions. The training program improved efficiency in caring for the mangabeys.�
* The effects of age and sex on interest toward movies of conspecifics in Japanese macaques (Macaca fuscata). Tsuchida, J., & Izumi, A. (Dept of Behavioral & Brain Sci., Primate Res. Inst., Kyoto Univ., Inuyama, Aichi 484-8506, Japan). Journal of the American Association for Laboratory Animal Science, 2009, 48, 286-291.
. . . �Previous studies have suggested that images of conspecifics are useful for environmental enrichment for nonhuman primates, but whether the age and sex of the animals alter the effectiveness of such images is unclear. We investigated preferences to movies in Japanese macaques (male and female; age, two to 19 yr). Each monkey was housed individually in a cage outfitted with a touch-sensitive computer display. A subject monkey that touched the display was shown one of 30 movies that were recorded at an open enclosure containing their conspecifics. During the experimental sessions, 25 of 38 subjects touched the display at least once. The response duration was longer when monkeys appeared in the movies. The response duration decreased with age in male monkeys but not female monkeys. The results suggested the movies of conspecifics are useful for environmental enrichment, but further consideration seems appropriate for various subpopulations, particularly aged monkeys.�
Conservation
* The lion-tailed macaque Macaca silenus (Primates: Cercopithecidae): Conservation history and status of a flagship species of the tropical rainforests of the Western Ghats, India. � Singh, M., Kaumanns, W., Singh, M., Sushma, H. S., & Molur. S. (Biopsychology Lab., Univ. of Mysore, Manasagangotri, Mysuru, Karnataka 570006, India [e-mail: [email protected]]). Journal of Threatened Taxa, 2009, 1, 151-157, <www.threatenedtaxa.org>.
. . .��The lion-tailed macaque is a threatened species inhabiting the rainforests of the Western Ghats mountain range in southern India. Once assessed to be less than a thousand individuals remaining in the wild habitats, the population is now estimated to be between 3000 and 3500 individuals. However, the rainforest habitats of the species are highly fragmented. During the past three decades or less, the population of this species has severely declined due to habitat degradation and illegal hunting in several areas of its occurrence. In situ conservation programs included notification of certain areas as lion-tailed macaque conservation regions. Several captive breeding programs have been initiated in order to have a viable captive population of the species. However, the analysis reveals that both in situ and ex situ conservation programs have not achieved the desired success and the species is even more endangered than it was a few decades ago. In this article, we discuss these conservation programs and suggest further measures for effective conservation of lion-tailed macaques.�
* Present distribution, population status, and conservation of western hoolock gibbons Hoolock hoolock (Primates: Hylobatidae) in Namdapha National Park, India, Kumar, A., Mary, P. P., & Bagchie, P. (Dept of Forestry, NE Regional Inst. of Science & Tech., Deemed Univ., Nirjuli, Itanagar, Arunachal Pradesh 791109, India [e-mail: [email protected]]). Journal of Threatened Taxa, 2009, 1, 203-210, <www.threatenedtaxa.org>.
. . .A survey on the present distribution, population status and conservation of western hoolock gibbon was conducted from September, 2006, to April, 2007, in Namdapha National Park, Arunachal Pradesh, northeastern India. The data were recorded from 12 localities in the Park: Gibbon land, Baghnallah, Deban, 15th Mile, 16th Mile, Hawaghar, 19th Mile, Haldibari, Hornbill camp, Baranallah, Firmbase camp, and Embyong. A total of 50 individuals in 20 groups were recorded during the census by using direct and indirect methods. Out of 20 groups, nine groups were observed through direct visual observation. The remaining 11 groups were estimated by using indirect observation methods such as songs, calls, and branch shaking. The composition of the population was 19 adult males (38%), 19 adult females (38%), and 12 immatures (24%). The group size was estimated as 2.5 individuals per group. Anthropogenic disturbances observed in the gibbon habitat were habitat loss, hunting and poaching, canopy gaps, livelihood issues for local people, and livestock grazing.
* Occurrence of the Madras tree shrew Anathana ellioti (Waterhouse) (Scandentia: Tupaiidae) in the Biligirirangan Hills, Karnataka, India. Srinivasan, U., Prashanth, N. S., Lakshminarayanan, S., Varma, K., Karthikeyan, S., Vellal, S., Cavale, G., Mandanna, D., Ross, P., & Thapa (Post-grad. Progr. in Wildlife Bio. & Conservation, Wildlife Conservation Society � India Program & Natl Ctr for Biol. Sci., Bangalore, Karnataka, India [e-mail: [email protected]]). Journal of Threatened Taxa, 2009, 1, 283-286, <www.threatenedtaxa.org>.
. . . �The Madras Tree Shrew is endemic to peninsular India. It inhabits deciduous forests and has until now not been reported from the Biligiriranga Swamy Temple Wildlife Sanctuary (BRT WLS) or the contiguous hill ranges or from anywhere else in the state of Karnataka. We provide details of nine independent sightings of this species from six locations in the BRT WLS between 2003 and 2008. Photographs of the three individuals from different locations are also presented. These records indicate an extension of the range of this species in the BRT WLS, and possibly the forests contiguous to the protected area.�
Diet and Nutrition
* Utilization of forest flora by Phayre�s leaf-monkey Trachypithecus phayrei (Primates: Cercopithecidae) in semi-evergreen forests of Bangladesh. Aziz, M. A., & Feeroz, M. M. (Dept of Zoology, Jahangirnagar Univ., Savar, Dhaka 1342, Bangladesh [e-mail: [email protected]]). Journal of Threatened Taxa, 2009, 1, 257-262, <www.threatenedtaxa.org>.
. . .Trachypithecus phayrei was observed to use 29 floral species belonging to 14 plant taxa for feeding, resting, and sleeping in the semi-evergreen forest of Bangladesh. Principal food items were young and mature leaves (47%), shoots (19%), flowers and buds (16%), fruits and seeds (14%), and petioles (4%). T. phayrei preferred leaves in December (76%), fruits and seeds in June (57%), and flowers and buds in April (41%); respective lowest preferences were in April (49%), February (10%), and January (9%). There were two feeding peaks: early morning (0600-0800 hr) and late afternoon (1400-1600 hr). The studied group of T. phayrei divided into sub-troops during foraging and feeding.
Disease
* Serologic evaluation of clinical and subclinical secondary hepatic amyloidosis in rhesus macaques (Macaca mulatta). MacGuire, J. G., Christe, K. L., Yee, J. L., Kalman-Bowlus, A. L., & Lerche, N. W. (California NPRC, Davis, CA 95616 [e-mail: [email protected]]). Comparative Medicine, 2009, 59, 168-173.
. . .Secondary hepatic amyloidosis in nonhuman primates carries a grave prognosis once animals become clinically ill. The purpose of this study was to establish serologic parameters that potentially could be used to identify rhesus macaques undergoing subclinical development of secondary hepatic amyloidosis. A retrospective analysis was completed by using serum biochemical profiles from 26 histologically diagnosed amyloidotic macaques evaluated at 2 stages of disease, clinical and subclinical (3 to 32 mo. prior to clinical signs of disease). Standard serum biochemistry values for cases were compared with institutional age- and gender-specific references ranges by construction of 95% confidence intervals for the difference between means. In addition, 19 histologically diagnosed amyloidotic macaques and 19 age-matched controls were assayed for changes in various parameters by using routinely banked, frozen (-80�C) sera available from clinical and subclinical time points. Clinically amyloidotic animals displayed increased levels of alkaline phosphatase, aspartate aminotransferase, lactate dehydrogenase, gamma glutamyltranspeptidase, and macrophage colony-stimulating factor and significantly decreased quantities of albumin and total cholesterol. Subclinical amyloidotic animals displayed increased levels of alkaline phosphatase, aspartate aminotransferase, lactate dehydrogenase, and serum amyloid A and decreased concentrations of albumin and total cholesterol. The serologic parameters studied indicate a temporal relationship of these factors not previously described, show a clear pattern of disease progression, and could be useful in subclinical disease detection.
* Treatment of giardiasis in common marmosets (Callithrix jacchus) with tinidazole. Kramer, J. A., Hachey, A. M., Wachtman, L. M., & Mansfield, K. G. (K. G. M., New England PRC, Southborough, MA 01772 [e-mail: [email protected]]). Comparative Medicine, 2009, 59, 174-179.
. . .Giardia intestinalis is a common protozoan parasite that can infect many laboratory animal primates, although its role as a contributor to the induction of gastrointestinal disease remains unclear. This study sought to investigate the prevalence of Giardia in a colony of common marmosets by using a Giardia antigen-capture assay and to address the possible eradication of this infection by using tinidazole, an antiprotozoal similar to metronidazole but requiring fewer doses. Among 31 colony marmosets, 13 (42%) were positive for Giardia. Two doses of oral tinidazole eliminated the infection in all animals. Repeat testing of the 13 Giardia-positive monkeys 1 year later showed that 11 remained negative and that treated animals had a significant increase in weight at 1 year. Giardia antigen is common in common marmoset feces, and treatment using oral tinidazole is possible and highly effective.
* Replication-deficient Ebolavirus as a vaccine candidate. Halfmann, P., Ebihara, H., Marzi, A., Hatta, Y., Watanabe, S., Suresh, M., Neumann, G., Feldmann, H., & Kawaoka, Y. (Y. K., Dept of Pathobiol. Sci., Sch. of Vet. Med., Univ. of Wisconsin, 2015 Linden Dr., Madison, WI 53706 [e-mail: [email protected]]). Journal of Virology, 2009, 83, 3810-3815, <jvi.asm.org/cgi/content/full/83/8/3810>.
. . . �Ebolavirus causes severe hemorrhagic fever, with case fatality rates as high as 90%. Currently, no licensed vaccine is available against Ebolavirus. We previously generated a replication-deficient, biologically contained Ebolavirus, EbolaΔVP30, which lacks the essential VP30 gene, grows only in cells stably expressing this gene product, and is genetically stable. Here, we evaluated the vaccine potential of EbolaΔVP30. First, we demonstrated its safety in STAT-1-knockout mice, a susceptible animal model for Ebolavirus infection. We then tested its protective efficacy in two animal models, mice and guinea pigs. Mice immunized twice with EbolaΔVP30 were protected from a lethal infection of mouse-adapted Ebolavirus. Virus titers in the serum of vaccinated mice were significantly lower than those in nonvaccinated mice. Protection of mice immunized with EbolaΔVP30 was associated with a high antibody response to the Ebolavirus glycoprotein and the generation of an Ebolavirus NP-specific CD8+ T-cell response. Guinea pigs immunized twice with EbolaΔVP30 were also protected from a lethal infection of guinea pig-adapted Ebolavirus. Our study demonstrates the potential of the EbolaΔVP30 virus as a new vaccine platform.�
* Dating the age of the SIV lineages that gave rise to HIV-1 and HIV-2. Wertheim, J. O., & Worobey, M. (Dept of Ecology & Evol. Biol., Univ. of Arizona, Tucson, AZ 85721 [e-mail: [email protected]]). PLoS Computational Biology, 2009, 5[5]: e1000377, <www.ploscompbiol.org/article/info%3Adoi%2F10.1371%2Fjournal.pcbi.1000377>.
. . . �Great strides have been made in understanding the evolutionary history of simian immunodeficiency virus (SIV) and the zoonoses that gave rise to HIV-1 and HIV-2. What remains unknown is how long these SIVs had been circulating in non-human primates before the transmissions to humans. Here, we use relaxed molecular clock dating techniques to estimate the time of most recent common ancestor for the SIVs infecting chimpanzees and sooty mangabeys, the reservoirs of HIV-1 and HIV-2, respectively. The date of the most recent common ancestor of SIV in chimpanzees is estimated to be 1492 (1266�1685), and the date in sooty mangabeys is estimated to be 1809 (1729�1875). Notably, we demonstrate that SIV sequences sampled from sooty mangabeys possess sufficient clock-like signal to calibrate a molecular clock; despite the differences in host biology and viral dynamics, the rate of evolution of SIV in sooty mangabeys is indistinguishable from that of its human counterpart, HIV-2. We also estimate the ages of the HIV-2 human-to-human transmissible lineages and provide the first age estimate for HIV-1 group N at 1963 (1948�1977). Comparisons between the SIV most recent common ancestor dates and those of the HIV lineages suggest a difference on the order of only hundreds of years. Our results suggest either that SIV is a surprisingly young lentiviral lineage or that SIV and, perhaps, HIV dating estimates are seriously compromised by unaccounted-for biases.�
* Complexity of the inoculum determines the rate of revesion of SIV Gag CD8 T cell mutant virus and outcome of infection. Loh, L., Reece, J., C., Fernandez, C. S., Alcantara, S., Center, R., Howard, J., Purcell, D. F. J., Balamurali, M., Petravic, J., Davenport, M. P., & Kent, S. J. (Dept of Microbiology & Immunology, Univ. of Melbourne, Melbourne, Victoria, Australia [e-mail: [email protected]]). PLoS Pathogens, 2009, 5[4], <www.ncbi.nlm.nih.gov/pubmed/19360124>.
. . . �Escape mutant (EM) virus that evades CD8+ T cell recognition is frequently observed following infection with HIV-1 or SIV. This EM virus is often less replicatively �fit� compared to wild-type (WT) virus, as demonstrated by reversion to WT upon transmission of HIV to a na�ve host and the association of EM virus with lower viral load in vivo in HIV-1 infection. The rate and timing of reversion is, however, highly variable. We quantified reversion to WT of a series of SIV and SHIV viruses containing minor amounts of WT virus in pigtail macaques using a sensitive PCR assay. Infection with mixes of EM and WT virus containing ≥10% WT virus results in immediate and rapid outgrowth of WT virus at SIV Gag CD8 T cell epitopes within 7 days of infection of pigtail macaques with SHIV or SIV. In contrast, infection with biologically passaged SHIVmn229 viruses with much smaller proportions of WT sequence, or a molecular clone of pure EM SIVmac239, demonstrated a delayed or slow pattern of reversion. WT virus was not detectable until ≥8 days after inoculation and took ≥8 weeks to become the dominant quasispecies. A delayed pattern of reversion was associated with significantly lower viral loads. The diversity of the infecting inoculum determines the timing of reversion to WT virus, which in turn predicts the outcome of infection. The delay in reversion of fitness-reducing CD8 T cell escape mutations in some scenarios suggests opportunities to reduce the pathogenicity of HIV during very early infection.�
* High frequencies of resting CD4+ T cells containing integrated viral DNA are found in rhesus macaques during acute lentivirus infections. Nishimura, Y., Sadjadpour, R., Mattapallil, J. J., Igarashi, T., Lee, W., Buckler-White, A., Roederer, M., Chun, T.-W., & Martin, M. A. (Lab. of Molecular Microbiology, NIH, Bethesda, MD 20892 [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2009, 106, 8015-8020, <www.pnas.org/content/106/19/8015.full>.
. . . �We and others have reported that the vast majority of virus-producing CD4+ T cells during the acute infection of rhesus macaques with simian immunodeficiency virus (SIV) or CXCR4 (X4)-using simian/human immunodeficiency viruses (SHIVs) exhibited a nonactivated phenotype. These findings have been extended to show that resting CD4+ T lymphocytes collected from SIV- or X4-SHIV-infected animals during the first 10 days of infection continue to release virus ex vivo. Furthermore, we observed high frequencies of integrated viral DNA (up to 5.1 � 104 DNA copies per 105 cells) in circulating resting CD4+ T cells during the first 10 days of the infection. Integration of SIV DNA was detected only in memory CD4+ T cells and SHIVs preferentially integrated into resting na�ve CD4+ T cells. Taken together, these results show that during the acute infection large numbers of resting CD4+ T cells carry integrated nonhuman primate lentiviral DNA and are the major source of progeny virions irrespective of coreceptor usage. Prompt and sustained interventions are therefore required to block the rapid systemic dissemination of virus and prevent an otherwise fatal clinical outcome.�
* Selection of a simian-human immunodeficiency virus strain resistant to a vaginal microbicide in macaques. Dudley, D. M., Wentzel, J. L., Lalonde, M. S., Veazey, R. S., & Arts, E. J. (E. J. A., Div. of Infect. Dis., BRB 1029, Case Western Reserve Univ., 10900 Euclid Ave., Cleveland, OH 44106 [e-mail: [email protected]]). Journal of Virology, 2009, 83, 5067-5076.
. . .PSC-RANTES binds to CCR5, inhibits human immunodeficiency virus type 1 (HIV-1) entry, and has been shown as a vaginal microbicide to protect rhesus macaques from a simian-human immunodeficiency virus chimera (SHIVSF162-p3) infection in a dose-dependent manner. In this study, env gene sequences from SHIVSF162-p3-infected rhesus macaques treated with PSC-RANTES were analyzed for possible drug escape variants. Two specific mutations located in the V3 region of gp120 (K315R) and C-helical domain of gp41 (N640D) were identified in a macaque (m584) pretreated with a 100 �M dose of PSC-RANTES. These two env mutations were found throughout infection (through week 77) but were found at only low frequencies in the inoculating SHIVSF162-p3 stock and in the other SHIVSF162-p3-infected macaques. HIV-1 env genes from macaque m584 (envm584) and from inoculating SHIVSF162-p3envp3) were cloned into an HIV-1 backbone. Increases in 50% inhibitory concentrations to PSC-RANTES with envm584 were modest (sevenfold) and most pronounced in cells expressing rhesus macaque CCR5 as compared to human CCR5. Nonetheless, virus harboring envm584, unlike inoculating virus envp3, could replicate even at the highest tissue culture PSC-RANTES concentrations (100 nM). Dual-virus competitions revealed a dramatic increase in fitness of chimeric virus containing envm584 (K315R/N640D) over that containing envp3, but again, only in rhesus CCR5-expressing cells. This study is the first to describe the immediate selection and infection of a drug-resistant SHIV variant in the face of a protective vaginal microbicide, PSC-RANTES. This rhesus CCR5-specific/PSC-RANTES resistance selection is particularly alarming given the relative homogeneity of the SHIVSF162-p3 stock compared to the potential exposure to a heterogeneous HIV-1 population in human transmission.
* Multisystemic eosinophilia resembling hypereosinophilic syndrome in a colony-bred owl monkey (Aotus vociferans). Gozalo, A. S., Rosenberg, H. F., Elkins, W. R., Montoya, E. J., & Weller, R. E. (Comp. Med. Branch, NIAID., NIH, Bethesda, MD 20892). Journal of the American Association for Laboratory Animal Science, 2009, 48, 303-306.
. . . �In animals, multisystemic eosinophilic disease is a rare condition characterized by eosinophilic and lymphoplasmacytic infiltrates in various organs. This disorder resembles the human disease known as hypereosinophilic syndrome, a condition defined by prolonged peripheral eosinophilia in the absence of recognizable etiology and associated with end-organ damage. In this report we describe a research-na�ve, colony-born, juvenile female owl monkey who presented clinically with severe respiratory distress and histologically with multiple end-organ infiltration with phenotypically mature eosinophils, plasma cells, and lymphocytes. No tumors or infectious agents were noted either macroscopically or microscopically. Cultures from lung samples revealed no bacteria or fungi. Histologic examination of lung, heart, thymus, liver, spleen, kidney, adrenal, pancreas, stomach, small intestine, and colon revealed no migrating nematode larvae, other parasites, or foreign material that might trigger eosinophilia, nor was there any evidence of or history consistent with an allergic etiology. Given that we ruled out most exogenous and endogenous triggers of eosinophilia, the signs, symptoms, and pathologic findings support the diagnosis of multisystemic eosinophilic disease. To our knowledge, this report is the first description of presumptive hypereosinophilic syndrome in a nonhuman primate.�
* Tacrolimus ointment: A novel and effective topical treatment of localized atopic dermatitis in a rhesus macaque (Macaca mulatta). Torreilles, S. L., Luong, R. H., Felt, S. A., & McClure, D. E. (Dept of Comp. Med., Stanford Univ. Sch. of Med., Stanford, CA 94305). Journal of the American Association for Laboratory Animal Science, 2009, 48, 307-311.
. . .An adult, male, rhesus macaque presented with pruritus and a focal, exudative, inflamed, erythematous skin lesion of approximately 2 cm in diameter on the ventral aspect of the mandible. The lesion resolved after 10 days of treatment with 1% chlorhexidine solution and triple-antibiotic ointment. However, the skin lesion subsequently recurred several times over a 2-month period. A punch biopsy was performed, and histological changes were most consistent with a diagnosis of atopic dermatitis. Treatment with topical tacrolimus ointment, an immunosuppressive drug, proved successful in the resolution of all clinical signs after 4 months. According to a literature review, this article is the first report of the use of tacrolimus ointment as a topical treatment of atopic dermatitis in a rhesus macaque.
Evolution, Genetics, and Taxonomy
* Age of Zhoukoudian Homo erectus determined with 26Al/10Be burial dating. Shen, G., Gao, X., Gao, B., & Granger, D. E. (G.S., College of Geographical Sciences, Nanjing Normal Univ., Nanjing, Jiangsu 210046, China [e-mail: [email protected]]; or D.E.G., Dept of Earth & Atmospheric Sciences, Purdue Univ., West Lafayette, IN 47907-2051 [e-mail: [email protected]]). Nature, 2009, 458, 198-200, <www.nature.com/nature/journal/v458/n7235/full/nature07741.html>.
. . . �The age of Zhoukoudian Homo erectus, commonly known as �Peking Man�, has long been pursued, but has remained problematic owing to the lack of suitable dating methods. Here we report cosmogenic 26Al/10Be burial dating of quartz sediments and artefacts from the lower strata of Locality 1 in the southwestern suburb of Beijing, China, where early representatives of Zhoukoudian Homo erectus were discovered. This study marks the first radioisotopic dating of any early hominin site in China beyond the range of mass spectrometric U-series dating. The weighted mean of six meaningful age measurements, 0.77 ± 0.08 million years (Myr, mean ± s.e.m.), provides the best age estimate for lower cultural layers 7�10. Together with previously reported U-series dating of speleothem calcite and palaeomagnetic stratigraphy, as well as sedimentological considerations, these layers may be further correlated to S6�S7 in Chinese loess stratigraphy or marine isotope stages (MIS) 17�19, in the range of ~0.68 to 0.78 Myr ago. These ages are substantially older than previously supposed and may imply early hominins� presence at the site in northern China through a relatively mild glacial period corresponding to MIS 18.�
* Successive radiations, not stasis, in the South American primate fauna. Hodgson, J. A., Sterner, K. N., Matthews, L. J., Burrell, A. S., Jani, R. A., Raaumb, R. L., Stewart, C.-B., & Disotell, T. R. (T. R. D., Dept of Anthropology, New York University, 25 Waverly Place, New York, NY 10003 [e-mail: [email protected]). Proceedings of the National Academy of Sciences, U.S.A. , 2009, 106, 5534-5539.
. . . �The earliest Neotropical primate fossils complete enough for taxonomic assessment, Dolichocebus, Tremacebus, and Chilecebus, date to approximately 20 Ma. These have been interpreted as either closely related to extant forms or as extinct stem lineages. The former hypothesis of morphological stasis requires most living platyrrhine genera to have diverged before 20 Ma. To test this hypothesis, we collected new complete mitochondrial genomes from Aotus lemurinus, Saimiri sciureus, Saguinus oedipus, Ateles belzebuth, and Callicebus donacophilus. We combined these with published sequences from Cebus albifrons and other primates to infer the mitochondrial phylogeny. We found support for a cebid/atelid clade to the exclusion of the pitheciids. Then, using Bayesian methods and well-supported fossil calibration constraints, we estimated that the platyrrhine most recent common ancestor (MRCA) dates to 19.5 Ma, with all major lineages diverging by 14.3 Ma. Next, we estimated catarrhine divergence dates on the basis of platyrrhine divergence scenarios and found that only a platyrrhine MRCA less than 21 Ma is concordant with the catarrhine fossil record. Finally, we calculated that 33% more change in the rate of evolution is required for platyrrhine divergences consistent with the morphologic stasis hypothesis than for a more recent radiation. We conclude that Dolichocebus, Tremacebus, and Chilecebus are likely too old to be crown platyrrhines, suggesting they were part of an extinct early radiation. We note that the crown platyrrhine radiation was concomitant with the radiation of two South American xenarthran lineages and follows a global temperature peak and tectonic activity in the Andes.�
* Early modern human diversity suggests subdivided population structure and a complex out-of-Africa scenario. Gunza, P., Bookstein, F. L., Mitteroecker, P., Stadlmayr, A., Seidler, H., & Weber, G. W. (G. W. W., Dept of Anthropology., Univ. of Vienna, Althanstr. 14, A-1090 Vienna, Austria [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2009, 106, 6094-6098, <www.pnas.org/content/106/15/6094.full>.
. . . �The interpretation of genetic evidence regarding modern human origins depends, among other things, on assessments of the structure and the variation of ancient populations. Because we lack genetic data from the time when the first anatomically modern humans appeared, between 200,000 and 60,000 years ago, instead we exploit the phenotype of neurocranial geometry to compare the variation in early modern human fossils with that in other groups of fossil Homo and recent modern humans. Variation is assessed as the mean-squared Procrustes distance from the group average shape in a representation based on several hundred neurocranial landmarks and semilandmarks. We find that the early modern group has more shape variation than any other group in our sample, which covers 1.8 million years, and that they are morphologically similar to recent modern humans of diverse geographically dispersed populations but not to archaic groups. Of the currently competing models of modern human origins, some are inconsistent with these findings. Rather than a single out-of-Africa dispersal scenario, we suggest that early modern humans were already divided into different populations in Pleistocene Africa, after which there followed a complex migration pattern. Our conclusions bear implications for the inference of ancient human demography from genetic models and emphasize the importance of focusing research on those early modern humans, in particular, in Africa.�
* The foot of Homo floresiensis. Jungers, W. L., Harcourt-Smith, W. E. H., Wunderlich, R. E., Tocheri, M. W., Larson, S. G., Sutikna, T., Due, R. A., & Morwood, M. J. (Dept of Anat. Sci., Stony Brook Univ. Med. Ctr, Stony Brook, NY 11794-8081 [e-mail: william.jungers@ stonybrook.edu]). Nature, 2009, 459, 81-84, <doi:10.1038/nature07989>.
. . . �Homo floresiensis is an endemic hominin species that occupied Liang Bua, a limestone cave on Flores in eastern Indonesia, during the Late Pleistocene epoch. The skeleton of the type specimen (LB1) of H. floresiensis includes a relatively complete left foot and parts of the right foot. These feet provide insights into the evolution of bipedalism and, together with the rest of the skeleton, have implications for hominin dispersal events into Asia. Here we show that LB1�s foot is exceptionally long relative to the femur and tibia, proportions never before documented in hominins but seen in some African apes. Although the metatarsal robusticity sequence is human-like and the hallux is fully adducted, other intrinsic proportions and pedal features are more ape-like. The postcranial anatomy of H. floresiensis is that of a biped, but the unique lower-limb proportions and surprising combination of derived and primitive pedal morphologies suggest kinematic and biomechanical differences from modern human gait. Therefore, LB1 offers the most complete glimpse of a bipedal hominin foot that lacks the full suite of derived features characteristic of modern humans and whose mosaic design may be primitive for the genus Homo. These new findings raise the possibility that the ancestor of H. floresiensis was not Homo erectus but instead some other, more primitive, hominin whose dispersal into southeast Asia is still undocumented.�
* Insular dwarfism in hippos and a model for brain size reduction in Homo floresiensis. Weston, E. M., & Lister, A. M. (Dept of Palaeontology, Natural History Museum, London SW7 5BD, U.K. [e-mail: [email protected]]). Nature, 2009, 459, 85-88, <doi:10.1038/nature07922>.
. . . �Body size reduction in mammals is usually associated with only moderate brain size reduction, because the brain and sensory organs complete their growth before the rest of the body during ontogeny. On this basis, �phyletic dwarfs� are predicted to have a greater relative brain size than �phyletic giants�. However, this trend has been questioned in the special case of dwarfism of mammals on islands. Here we show that the endocranial capacities of extinct dwarf species of hippopotamus from Madagascar are up to 30% smaller than those of a mainland African ancestor scaled to equivalent body mass. These results show that brain size reduction is much greater than predicted from an intraspecific �late ontogenetic� model of dwarfism in which brain size scales to body size with an exponent of 0.35. The nature of the proportional change or grade shift observed here indicates that selective pressures on brain size are potentially independent of those on body size. This study demonstrates empirically that it is mechanistically possible for dwarf mammals on islands to evolve significantly smaller brains than would be predicted from a model of dwarfing based on the intraspecific scaling of the mainland ancestor. Our findings challenge current understanding of brain�body allometric relationships in mammals and suggest that the process of dwarfism could in principle explain small brain size, a factor relevant to the interpretation of the small-brained hominin found on the Island of Flores, Indonesia.�
* Elucidating geological and biological processes underlying the diversification of Sulawesi tarsiers. Merker, S., Driller, C., Perwitasari-Farajallah, D., Pamungkas, J., & Zischler, H. (Inst of Anthropology, Johannes-Gutenberg Univ. Mainz, Colonel-Kleinmann-Weg 2 (SB II), 55099 Mainz, Germany [e-mail: [email protected]]). Proceedings of the National Academy of Sciences, U.S.A., 2009, 106, 8459-8464, <www.pnas.org/content/106/21/8459.full>.
. . . �Because of their exceptionally long independent evolution, a range diminution of their Eocene relatives, and a remarkable subsequent diversification in Southeast Asia, tarsiers are of particular importance to evolutionary primatologists. Little is known, however, of the processes shaping the radiation of these small enigmatic primates�especially on the Indonesian island of Sulawesi, their center of endemism. Geological reconstructions and progress in applying DNA sequence information to divergence dating now provide us with the tools and background to comprehend tarsier dispersal. Here, we describe effects of plate-tectonic movements, Pleistocene sea level changes, and hybridization on the divergence of central Sulawesi tarsiers. We analyzed 12 microsatellites, the cytochrome b gene, the hypervariable region I of the mitochondrial control region, and the sex-determining region on the Y-chromosome from 144 specimens captured along a transect crossing a species boundary and a contact zone between 2 microplates. Based on these differentially inherited genetic markers, geographic information, and recordings of vocalizations, we demonstrate that the species boundary coincides with a tectonic suture. We estimate the most recent common ancestor of the 2 taxa to have lived 1.4 Mya, we describe asymmetrical introgressive hybridization, and we give evidence of unbiased dispersal in one species and male-biased dispersal in another species. This study exemplifies that the distribution of tarsier acoustic forms on Sulawesi is consistent with the allocation of genetic variability and that plate-tectonic and glacial events have left traceable marks in the biogeography of this island's unique fauna.�
Nutrition
* Nutritional goals of wild primates. Felton, A. M., Felton, A., Lindenmayer, D. B., & Foley, W. J. (Fenner School of Environment & Society, Australian Natl Univ., Canberra, ACT 0200, Australia [e-mail: [email protected]]). Functional Ecology, 2009, 23, 70-78 <www3.interscience.wiley.com/cgi-bin/fulltext/121640886>.
. . . �Primates meet their nutritional goals by prioritizing certain nutritional parameters when choosing the types and quantities of different foods. There are five major models applied in primate nutritional ecology, each of which proposes that diet selection subserves a different primary nutritional goal: (i) energy maximization; (ii) nitrogen (protein) maximization; (iii) avoidance or regulation of intake of plant secondary metabolites; (iv) limitations on the intake of dietary fiber; and (v) nutrient balancing. Here, we review the evidence in support of each of these nutritional goals as drivers of primate diet selection. We discuss some of the costs and benefits associated with different methodological approaches used in primate nutritional ecology. New approaches developed outside of primatology have provided better frameworks for understanding the nutritional goals of some primate species. We suggest that the field of primate nutritional ecology needs to take greater advantage of the techniques developed by nutritional ecologists working in other fields. Specifically, we recommend (i) the increased application of the Geometric Framework for nutrition, (ii) the application of methodological approaches that enable the estimation of nutrient and energy availability from food sources, and (iii) continuous follows of individual primates in the wild for determining primary nutritional goals.�
------------------------------
We would like to acknowledge Primate-Science as a source for information about new books.
------------------------------
* * *
Position Available: Atlanta, Georgia
Emory University is seeking an accomplished clinical veterinarian to join a highly active, nationally renowned transplant medicine group. Working closely with clinical veterinarians at the Yerkes National Primate Research Center, this person will be responsible for organizing, implementing, and tracking transplant studies utilizing nonhuman primates. �We are looking for a candidate with strong clinical and surgical skills and, ideally with a primatology and laboratory animal background. This is a clinical research position and includes a faculty appointment at a junior or senior level. This is a new, long-term position that we would like to fill as quickly as possible. Interested parties should immediately contact Dr Jim Else, Associate Director for Veterinary Resources at Yerkes [e-mail: [email protected]], with a CV and a letter detailing their interest.�
* * *
Contents of Volumes 43-45 (2004-2006)
Contents of Volumes 46-present (2007-2009)
* * *
All correspondence concerning the Newsletter should be addressed to:
Judith E. Schrier, Psychology Department, Box 1853, Brown University
Providence, Rhode Island 02912. [401-863-2511; FAX: 401-863-1300]
e-mail address: Judith_Schrier@brown.edu
Current and back issues of the Newsletter are available on the World Wide Web at
http://www.brown.edu/primate
The Newsletter is supported by Brown University
Cover photograph of ring-tailed lemurs (Lemur catta), taken at the San Diego Zoo by Paul Wilde, 1997
Copyright (c) 2009 by Brown University
Last updated: June 23, 2009